


Abstract
Although flavopiridol, a semisynthetic flavone, was initially thought to be a specific inhibitor of cyclin-dependent kinases, it has now been shown that flavopiridol mediates antitumor responses through mechanism(s) yet to be defined. We have shown previously that flavopiridol abrogates tumor necrosis factor (TNF)-induced nuclear factor-κB (NF-κB) activation. In this report, we examined whether this flavone affects other cellular responses activated by TNF. TNF is a potent inducer of activator protein-1 (AP-1), and flavopiridol abrogated this activation in a dose- and time-dependent manner. Flavopiridol also suppressed AP-1 activation induced by various carcinogens and inflammatory stimuli. When examined for its effect on other signaling pathways, flavopiridol inhibited TNF-induced activation of various mitogen-activated protein kinases, including c-Jun NH2-terminal kinase (JNK), p38 mitogen-activated protein kinase (MAPK), and p44/p42 MAPK. It is noteworthy that this flavone also suppressed TNF-induced activation of Akt, a cell survival kinase, and expression of various antiapoptotic proteins, such as IAP-1, IAP-2, XIAP, Bcl-2, Bcl-xL, and TRAF-1. Flavopiridol also inhibited the TNF-induced induction of intercellular adhesion molecule-1, c-Myc, and c-Fos, all known to mediate tumorigenesis. Moreover, TNF-induced apoptosis was enhanced by flavopiridol through activation of the bid-cytochrome-caspase-9-caspase-3 pathway. Overall, our results clearly suggest that flavopiridol interferes with the TNF cell-signaling pathway, leading to suppression of antiapoptotic mechanisms and enhancement of apoptosis.
Flavopiridol is a semisynthetic flavonoid closely related to a compound originally isolated from the stem bark of Dysoxylum binectariferum (also called rohitukine), a plant indigenous to India. The parent compound is identical to flavopiridol except that a methyl group replaces the chlorophenyl moiety at position 2 (Naik et al., 1988). Flavopiridol has been shown to be a potent inhibitor of cyclin-dependent kinase (CDK) 1, CDK 2, CDK 4, and CDK 7 (Carlson et al., 1996). It inhibits CDKs by competing with ATP at the nucleotide-binding site on CDKs, as indicated by kinetics studies (Losiewicz et al., 1994) and X-ray crystallography of the CDK 2-flavopiridol complex (De Azevedo et al., 1996). The tyrosine phosphorylation of CDK 2 is also inhibited by this flavone (Worland et al., 1993). Through inhibition of CDKs, flavopiridol induces arrest of cell growth at the G1 and G2 phases of the cell cycle (Carlson et al., 1996).
Because of its ability to suppress the growth of breast carcinoma (Carlson et al., 1996), lung carcinoma (Bible and Kaufmann, 1996), chronic B cell leukemia and lymphoma (König et al., 1997; Arguello et al., 1998), multiple myeloma (Gojo et al., 2002), and head and neck squamous cell carcinoma (Patel et al., 1998), flavopiridol is currently in clinical trials for the treatment of different cancers (Karp et al., 2003; Morris et al., 2006; Fornier et al., 2007). Flavopiridol has also been shown to enhance the activity of other growth-suppressing agents, such as tumor necrosis factor (TNF), doxorubicin, and etoposide (Bible and Kaufmann, 1997; Cartee et al., 2003; Takada and Aggarwal, 2004).

Effect of flavopiridol on TNF-induced AP-1 activation. A, HL60 cells were preincubated at 37°C with the indicated concentrations of flavopiridol for 8 h and then treated with 0.1 nM TNF at 37°C for 30 min. Nuclear extracts were then prepared and assayed for AP-1 activation by EMSA. B, HL60 cells were preincubated at 37°C with 100 nM flavopiridol for the indicated times and then treated with 0.1 nM TNF at 37°C for 30 min. Nuclear extracts were then prepared and assayed for AP-1 activation by EMSA. C, effect of flavopiridol on the activation of AP-1 induced by different concentrations of TNF. HL60 cells were incubated with 100 nM flavopiridol for 8 h, treated with different concentrations of TNF for 30 min, and then subjected to EMSA for AP-1 activation. D, AP-1 induced by TNF is composed of c-Fos and c-Jun subunits. Nuclear extracts from untreated or TNF-treated cells were incubated with indicated antibodies, unlabeled AP-1 oligonucleotide probe, or mutant oligo-probe and then assayed for AP-1 activation by EMSA. E, effect of flavopiridol on AP-1 activation induced by different activators. Flavopiridol blocks AP-1 activation induced by TNF, LPS, doxorubicin, interleukin-1β (IL-1β), PMA, okadaic acid, and H2O2. HL60 cells were preincubated with 100 nM flavopiridol for 8 h, treated with 0.1 nM TNF, 10 μg/ml LPS, and IL-1β for 30 min, 15 ng/ml PMA for 1 h, 500 μMH2O2 for 2 h, 1 μg/ml doxorubicin and 500 nM okadaic acid for 6 h and then analyzed for AP-1 activation. The cell viability (C.V.) was determined by the trypan blue exclusion assay.
Research in the last few years has indicated that in addition to inhibiting CDK activity, flavopiridol can also alter the expression of various proteins such as Bcl-2 (König et al., 1997), Mcl-1 (Gojo et al., 2002), cyclin D1 (Takada and Aggarwal, 2004), and vascular endothelial growth factor (Rapella et al., 2002). Most of these genes are known to be regulated by the nuclear transcription factor NF-κB (Kumar et al., 2004).
We have recently reported that flavopiridol can suppress TNF-induced NF-κB activation (Takada and Aggarwal, 2004). TNF is also a very potent activator of AP-1, JNK, p38 MAPK, p44/p42 MAPK, and Akt (Aggarwal, 2003). Whether flavopiridol affects cellular responses activated by TNF, other than NF-κB, is not known. Also unclear is how the expression of antiapoptotic gene products and the apoptotic effects of TNF are affected by flavopiridol. We therefore investigated in detail the effect of flavopiridol on different signals transduced by TNF. We found that flavopiridol abrogated the TNF-induced activation of AP-1, JNK, p38 MAPK, p44/p42 MAPK, and Akt and suppressed the expression of antiapoptotic gene products, leading to enhancement of apoptosis through bid cleavage, cytochrome c release, and caspase activation.
Materials and Methods
Reagents. Flavopiridol was obtained from the Drug Synthesis and Chemistry Branch, Developmental Therapeutics Program, Division of Cancer Treatment and Diagnosis, National Cancer Institute (Bethesda, MD). A solution of flavopiridol (1 mM) was prepared in dimethyl sulfoxide and then diluted to desired concentrations before use with the media. In all experiments, concentration of dimethyl sulfoxide was kept lower than 0.1%. Bacteria-derived human recombinant TNF, purified to homogeneity with a specific activity of 5 × 107 U/mg, was kindly provided by Genentech (South San Francisco, CA). Penicillin, streptomycin, RPMI 1640 medium, Dulbecco's modified Eagle's medium, and fetal bovine serum were obtained from Invitrogen (Carlsbad, CA). PMA, okadaic acid, H2O2, and agarose (type VII) were obtained from Sigma Chemical (St. Louis, MO). The antibodies against c-Fos, c-Jun, cyclin D1, JNK1, p44/p42 MAPK, p38 MAPK, Akt, inhibitor-of-apoptosis protein 1/2 (IAP1/2), Bcl-2, Bcl-xL, TRAF1, ICAM-1, c-Myc, cytochrome c, Bid, caspase-9, caspase-3, and PARP were obtained from Santa Cruz Biotechnology (Santa Cruz, CA). Phospho-specific Akt, phospho-specific p44/p42 MAPK, and phospho-specific p38 MAPK antibodies were purchased from Cell Signaling (Danvers, MA). X-chromosome-linked inhibitor-of-apoptosis protein (XIAP) antibody was obtained from BD Biosciences (San Diego, CA).
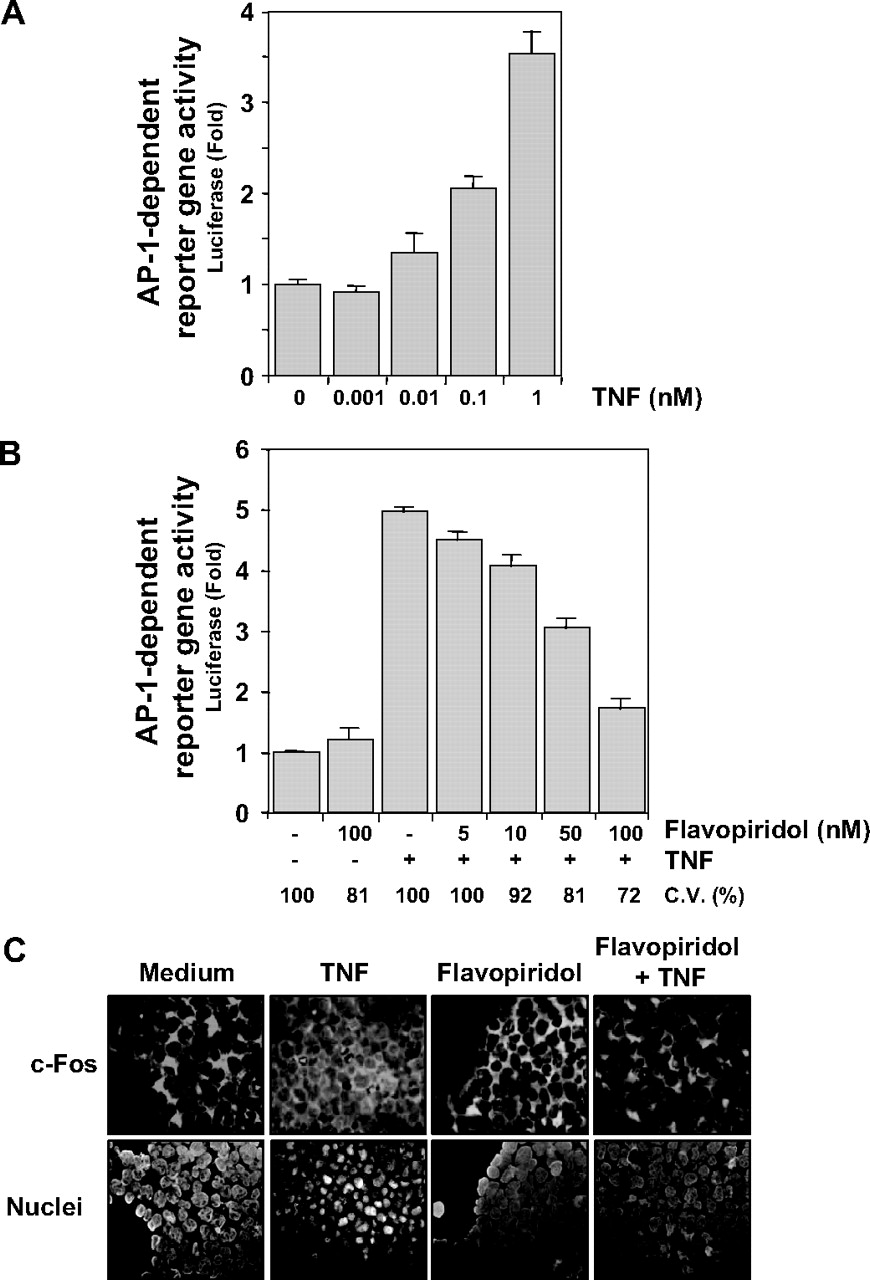
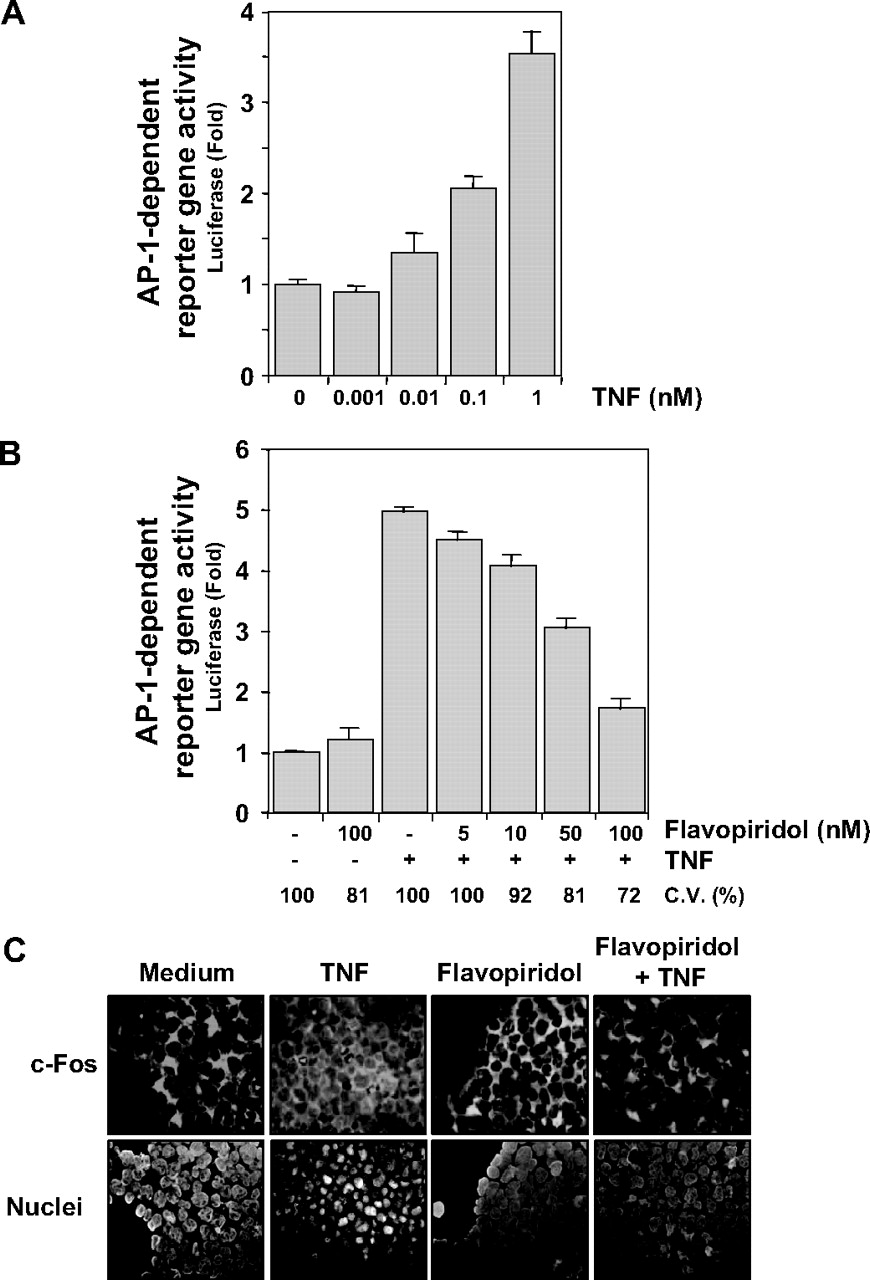
Flavopiridol inhibits TNF-induced AP-1-dependent reporter gene expression. A, cells were transiently transfected with an AP-1-dependent luciferase reporter plasmid for 24 h and then washed and treated with the indicated concentrations of TNF for a further 24 h. Whole-cell lysates were prepared and assayed for luciferase activity as described under Materials and Methods. B, cells were transiently transfected with an AP-1-dependent luciferase reporter plasmid for 24 h, washed and treated with the indicated concentrations of flavopiridol for 8 h, and then treated with 1 nM TNF for a further 24 h. Whole-cell lysates were prepared and assayed for luciferase activity as described under Materials and Methods. The cell viability (C.V.) was determined by MTT methods as described under Materials and Methods. C, immunocytochemical analysis of AP-1 subunit c-Fos localization after treatment with TNF in the absence or presence of 100 nM flavopiridol. Cells were incubated with flavopiridol for 8 h and then treated with 1 nM TNF for 20 min. Cells were subjected to immunocytochemical analysis as described under Materials and Methods.
Cell Lines. HL60 (human myeloid leukemia), A293 (human embryonic kidney), and H1299 (human lung adenocarcinoma) cells were obtained from American Type Culture Collection. HL60 cells were cultured in RPMI 1640 medium; A293 cells were cultured in Dulbecco's modified Eagle's medium. Both media were supplemented with 10% fetal bovine serum, 100 U/ml penicillin, and 100 μg/ml streptomycin.
Electrophoretic Mobility Shift Assays. To measure AP-1 activation, we performed EMSA. In brief, nuclear extracts prepared from TNF-treated cells (2 × 106/ml) were incubated with 32P-end-labeled AP-1 consensus oligonucleotide 5′-CGCTTGATGACTCAGCCGGAA-3′ (bold indicates AP-1 binding site) for 30 min at 37°C, and the DNA-protein complex formed was separated from free oligonucleotide on 5% native polyacrylamide gels. The specificity of binding was also examined by competition with the unlabeled oligonucleotide. For supershift assays, nuclear extracts prepared from TNF-treated HL60 cells were incubated with antibodies against either the c-Fos or the c-Jun subunits of AP-1 for 15 min at 37°C before the complex was analyzed by EMSA. Antibodies against cyclin D1 and preimmune serum were included as negative controls. The dried gels were visualized, and radioactive bands were quantified using a PhosphorImager and ImageQuant software (GE Healthcare, Chalfont St. Giles, Buckinghamshire, UK).
Luciferase Assay. To determine the effect of flavopiridol on TNF-induced AP-1-dependent luciferase reporter gene expression, A293 cells were seeded at a concentration of 1 × 105 cells in a six-well plate and transfected with 0.5 μg of DNA consisting of AP-1 promoter-luciferase reporter plasmid by FuGENE6 (Roche Applied Science, Indianapolis, IN). After 12 h, cells were incubated with 100 nM flavopiridol for 8 h, treated with TNF for 24 h, and harvested. Luciferase activity was then measured using the Promega luciferase assay system (Madison, WI) and detected using a Victor 3 microplate reader (PerkinElmer Life and Analytical Sciences, Waltham, MA).
Nuclear Localization of c-Fos by Immunocytochemistry. To determine the effect of flavopiridol on TNF-induced nuclear translocation of c-Fos, treated HL-60 cells were plated on a poly-l-lysine-coated glass slide by centrifugation using a Shandon Cytospin 4 (Thermo Fisher Scientific, Waltham, MA), air-dried, fixed with 4% paraformaldehyde, and permeabilized with 0.2% Triton X-100. After being washed in PBS, slides were blocked with 5% normal goat serum for 1 h and then incubated with rabbit polyclonal anti-c-Fos antibody at a 1:200 dilution. After overnight incubation at 4°C, the slides were washed, incubated with goat anti-rabbit IgG-Alexa Fluor 594 (Invitrogen, Carlsbad, CA) at a 1:200 dilution for 1 h, and counterstained for nuclei with Hoechst 33342 (50 ng/ml) for 5 min. Stained slides were mounted with mounting medium purchased from Sigma Chemical and analyzed using a fluorescence microscope (Labophot-2; Nikon, Tokyo, Japan). Pictures were a captured using Photometrics Coolsnap CF color camera (Nikon, Lewisville, TX) and MetaMorph version 4.6.5 software (Molecular Devices, Sunnyvale, CA).
Western Blot Analysis. To determine the effect of flavopiridol on the levels of protein expression in TNF-treated HL60 cells, we prepared whole-cell extracts using lysis buffer (20 mM Tris, pH 7.4, 250 mM NaCl, 2 mM EDTA, pH 8, 0.1% Triton X-100, 0.01 mg/ml aprotinin, 0.005 mg/ml leupeptin, 0.4 mM phenylmethylsulfonyl fluoride, and 4 mM NaVO4) and performed SDS-polyacrylamide gel electrophoresis (PAGE). After electrophoresis, the proteins were electrotransferred to nitrocellulose membranes, blotted with each antibody, and detected using ECL reagent (GE Healthcare). The density of the bands was measured using NIH Image (http://rsb.info.nih.gov/nih-image/).
JNK Assay. To determine the effect of flavopiridol on the kinase activity of JNK in HL60 cells, JNK complex from whole-cell extracts was precipitated with antibody against JNK1, followed by treatment with protein A/G-Agarose beads (Pierce, Rockford, IL). After 2 h of incubation, the beads were washed with lysis buffer and then assayed in kinase assay mixture containing 50 mM HEPES, pH 7.4, 20 mM MgCl2, 2 mM dithiothreitol, 20 μCi of [γ-32P]ATP, 10 μM unlabeled ATP, and 2 μg of substrate GST-c-Jun (1-79). The immunocomplex was incubated at 30°C for 30 min and then boiled with SDS sample buffer for 5 min. Finally, the protein was resolved on 10% SDS-PAGE, the gel was dried, and the radioactive bands were visualized using the PhosphorImager. To determine the total amount of JNK1 in each sample, whole-cell extracts were subjected to Western blot analysis using anti-JNK1 antibody.
Measurement of Cytochrome c Release. To determine the effect of flavopiridol on TNF-induced cytochrome c release, HL60 cells were treated with 100 nM flavopiridol for 8 h and then treated with 1 nM TNF for the indicated times; the cytosolic extracts were then prepared as described previously (Yang et al., 1997). In brief, the cells were washed with PBS, resuspended in the buffer containing 0.25 M sucrose, 30 mM Tris-HCl, pH 7.9, 1 mM EDTA, 1 mM phenylmethylsulfonyl fluoride, 2 mM sodium orthovanadate, 10 mM NaF, 2 μg/ml leupeptin, and 2 μg/ml aprotinin and then homogenized gently with a glass Dounce homogenizer for 20 strokes. The homogenates were centrifuged at 425g for 10 min to remove nuclei, and the supernatants were centrifuged at 20,817g for 30 min to remove mitochondria and other insoluble fragments. The supernatants were again centrifuged as above to ensure complete removal of mitochondria. Protein (50 μg) was subjected to 15% SDS-PAGE, and then Western blot analysis was performed using anti-cytochrome c antibody.
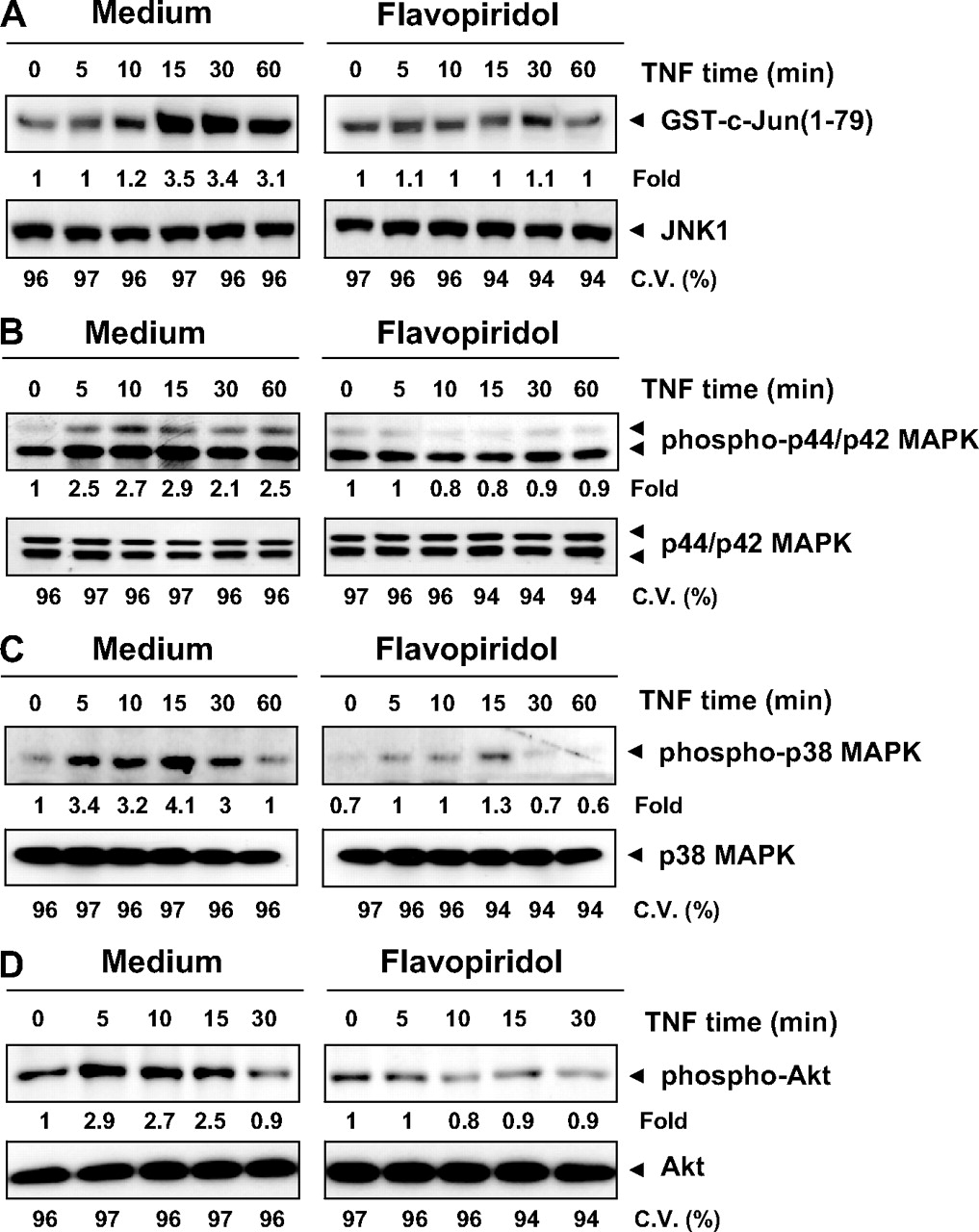
Effect of flavopiridol on TNF-induced JNK, p44/p42 MAPK, p38 MAPK, and Akt activation. A, HL60 cells were incubated with 100 nM flavopiridol for 8 h and then treated with 1 nM TNF for the indicated times. Whole-cell extracts were prepared and immunoprecipitated with anti-JNK1 antibody, and then the kinase assay was performed using GST-c-Jun (1-79) as a substrate. Whole-cell extracts were also subjected to Western blot analysis using anti-JNK1 antibody. B, HL60 cells were incubated with 100 nM flavopiridol for 8 h and treated with 1 nM TNF for the indicated times. Whole-cell extracts were prepared, and then Western blot analysis was performed using anti-phospho-p44/p42 MAPK and p44/42 MAPK antibodies. C, HL60 cells were incubated with 100 nM flavopiridol for 8 h and treated with 1 nM TNF for the indicated times. Whole-cell extracts were prepared, and then Western blot analysis was performed using antiphospho-p38 MAPK and p38 MAPK antibodies. D, HL60 cells were incubated with 100 nM flavopiridol for 8 h and treated with 1 nM TNF for the indicated times. Whole-cell extracts were prepared, and then Western blot analysis was performed using anti-phospho-Akt and Akt antibodies. The cell viability (C.V.) was determined by the trypan blue exclusion asssay.
Hoechst Staining. To determine the effect of flavopiridol on TNF-induced chromosomal fragmentation, HL60 cells were treated with 100 nM flavopiridol for 8 h and treated with 1 nM TNF for 16 h. Cells were fixed with 2% glutalaldehyde, stained with Hoechst 33342 (50 ng/ml), and analyzed under a fluorescence microscope; pictures were captured using a Photometrics Coolsnap CF color camera.
MTT Assay. The effect of flavopiridol on the cytotoxic effects of TNF, cisplatin, and paclitaxel was determined by the MTT dye reduction method. In brief, HL60 cells (5000 cells/well) were incubated with 100 nM flavopiridol for 8 h in triplicate in a 96-well plate and then treated with 1 nM TNF, 30 μg/ml cisplatin, and 1 nM paclitaxel for 24 h at 37°C. MTT solution was then added to each well. After a 2-h incubation at 37°C, lysis buffer (20% SDS, 50% dimethylformamide) was added, the cells were incubated overnight at 37°C, and the optical density was then measured at 570 nm using a 96-well multiscanner (MRX Revelation; Dynex Technologies, Chantilly, VA). Percentage cytotoxicity was calculated by dividing the absorbance of treated group with that of untreated group and multiplied with 100. The resulting value was subtracted from 100 to obtain the percentage cytotoxicity.
Trypan Blue Exclusion Assay. Cell suspension was mixed with equal volume of 0.4% isotonic trypan blue solution. Total cell number and fraction of nonviable, dye-accumulating cells were counted after 2 min in Fuchs-Rosenthal hemocytometer under light microscope.
Clonogenic Assay. H1299 cells were seeded in six-well plates at 500 cells/well in RPMI 1640 medium containing 10% serum. After 12 h, cells were treated with medium containing indicated concentrations of flavopiridol and TNF (1 nM). The medium with flavopiridol and TNF was replaced after every 4 days. After 10 days of incubation, colonies were stained with 0.3% crystal violet solution (dissolved in 1:1 mixture of methanol and H2O) for 2 min, washed once with Dulbecco's phosphate-buffered saline, air-dried, and manually counted. Each point was a mean of three replicate wells.
In another set of experiments, HL60 cells (1 × 103) were plated in RPMI 1640 medium containing 0.33% agarose type VII, 10% serum, various concentrations of flavopiridol and TNF (1 nM). Cells were incubated for 10 days. Colonies (> 50 cells) were counted using an inverted microscope.

Flavopiridol inhibited TNF-induced AP-1-dependent gene products. A, flavopiridol suppressed TNF-induced expression of antiapoptotic proteins. HL60 cells were incubated with 100 nM flavopiridol for 8 h and then treated with 1 nM TNF for the indicated times. Whole-cell extracts were prepared and analyzed by Western blot analysis using the indicated antibodies. B, flavopiridol suppressed TNF-induced ICAM-1, c-Myc, and c-Fos expressions. HL60 cells were incubated with 100 nM flavopiridol for 8 h and then treated with 1 nM TNF for the indicated times. Whole-cell extracts were prepared and analyzed by Western blot using the indicated antibodies. The cell viability (C.V.) was determined by the trypan blue exclusion assay.
Results
The goal of this study was to investigate the effect of flavopiridol on TNF-induced signaling. For most experiments, we used HL60 cells because they express both types of TNF receptors. For most studies, cells were treated with 100 nM or less flavopiridol for 8 h. The cell viability under these conditions, as determined by trypan blue exclusion, was greater than 90%. Pretreatment of cells with 100 nM flavopiridol for 8 h lead to cell viability of 97, 94, 89, 71, and 59% after 0, 3, 6, 12, and 24 h of TNF (1 nM) treatment, respectively.
Flavopiridol Inhibited TNF-Induced AP-1 Activation. TNF is one of the most potent activators of AP-1 (Karin et al., 1997). To determine the effect of flavopiridol on TNF-induced AP-1 activation, HL60 cells were treated with the indicated concentrations of flavopiridol, exposed to TNF, and then examined for AP-1 activation by DNA-binding activity using EMSA. As shown in Fig. 1 A, TNF activated AP-1 and flavopiridol pretreatment abolished TNF-induced AP-1 activation in a dose-dependent manner. We also pretreated cells with 100 nM flavopiridol for the indicated times, treated them with TNF, and then analyzed them for AP-1 activation. We found that flavopiridol inhibited the TNF-induced AP-1 activation in a time-dependent manner (Fig. 1B). Cells were also pretreated with flavopiridol, treated with increasing concentrations of TNF up to 10,000 pM, and then analyzed for AP-1 activation. Flavopiridol inhibited even the AP-1 activation induced by 10,000 pM TNF (Fig. 1C). It is noteworthy that, although not seen earlier, flavopiridol also suppressed the basal AP-1 activity in this experiment.
As indicated by the supershift analysis, TNF-induced AP-1 consisted of c-Fos and c-Jun subunits (Fig. 1D). Neither non-specific antibody against cyclin D1 nor preimmune serum had any effect on the supershift. Formation of the AP-1 band could be prevented by unlabeled oligonucleotide.
AP-1 is also activated by a wide variety of other agents in addition to TNF, including LPS, doxorubicin, interleukin-1β, phorbol ester, okadaic acid, and H2O2. Therefore, we examined the effect of flavopiridol on the activation of AP-1 by these various agents. We found that flavopiridol suppressed the activation of AP-1 induced by all these various agents (Fig. 1E). These results suggest that flavopiridol may act at a general step of convergence in the signal transduction pathway leading to AP-1 activation.
Flavopiridol Repressed TNF-Induced AP-1-Dependent Reporter Gene Expression. Although we showed by the DNA-binding assay that flavopiridol blocks AP-1 activation, we also determined the effect of flavopiridol on TNF-induced AP-1-dependent reporter gene expression. We transiently transfected the cells with the AP-1-regulated luciferase reporter construct and then stimulated them with TNF. We found that TNF induced AP-1-regulated reporter gene expression in a dose-dependent manner (Fig. 2A). Flavopiridol suppressed the TNF-induced AP-1-dependent reporter gene activation in a dose-dependent manner (Fig. 2B).
Flavopiridol Inhibited TNF-Induced Nuclear Translocation of c-Fos. Nuclear translocation of c-Fos is required for its transcriptional activity (Karin et al., 1997). An immunocytochemical assay showed that, in untreated cells, c-Fos is localized in the cytoplasm, TNF induces nuclear translocation, and flavopiridol suppresses the nuclear translocation (Fig. 2C).
Flavopiridol Suppressed TNF-Induced JNK, p44/p42 MAPK, and p38 MAPK Activation. TNF is also a potent activator of JNK, p38 MAPK, and p44/p42 MAPK (Aggarwal, 2003). We examined whether flavopiridol can modulate the activation of these kinases induced by TNF. Cells were pretreated with flavopiridol and then treated with TNF for the indicated times. Whole-cell extracts were prepared, immunoprecipitated with anti-JNK1 antibody, and then analyzed for JNK activity. As shown in Fig. 3A, TNF induced JNK activation in a time-dependent manner, and flavopiridol blocked it. Wholecell extracts were also subjected to Western blot analysis using anti-p44/p42 MAPK and p38 MAPK. TNF induced these MAPKs in a time-dependent manner, and flavopiridol suppressed them (Fig. 3, B and C). Thus, flavopiridol suppressed the activation of all three MAPKs induced by TNF.
Flavopiridol Suppressed TNF-Induced Akt Activation. Akt is one of the most important cell survival kinases activated by TNF (Aggarwal, 2003). To determine the effect of flavopiridol on TNF-induced activation of Akt, whole-cell extracts were also analyzed by Western blot using antiphospho-specific-Akt antibody (Fig. 3D). TNF induced phosphorylation of Akt, and this activation was suppressed by flavopiridol. The basal level of phospho-Akt was not significantly affected by the flavone.
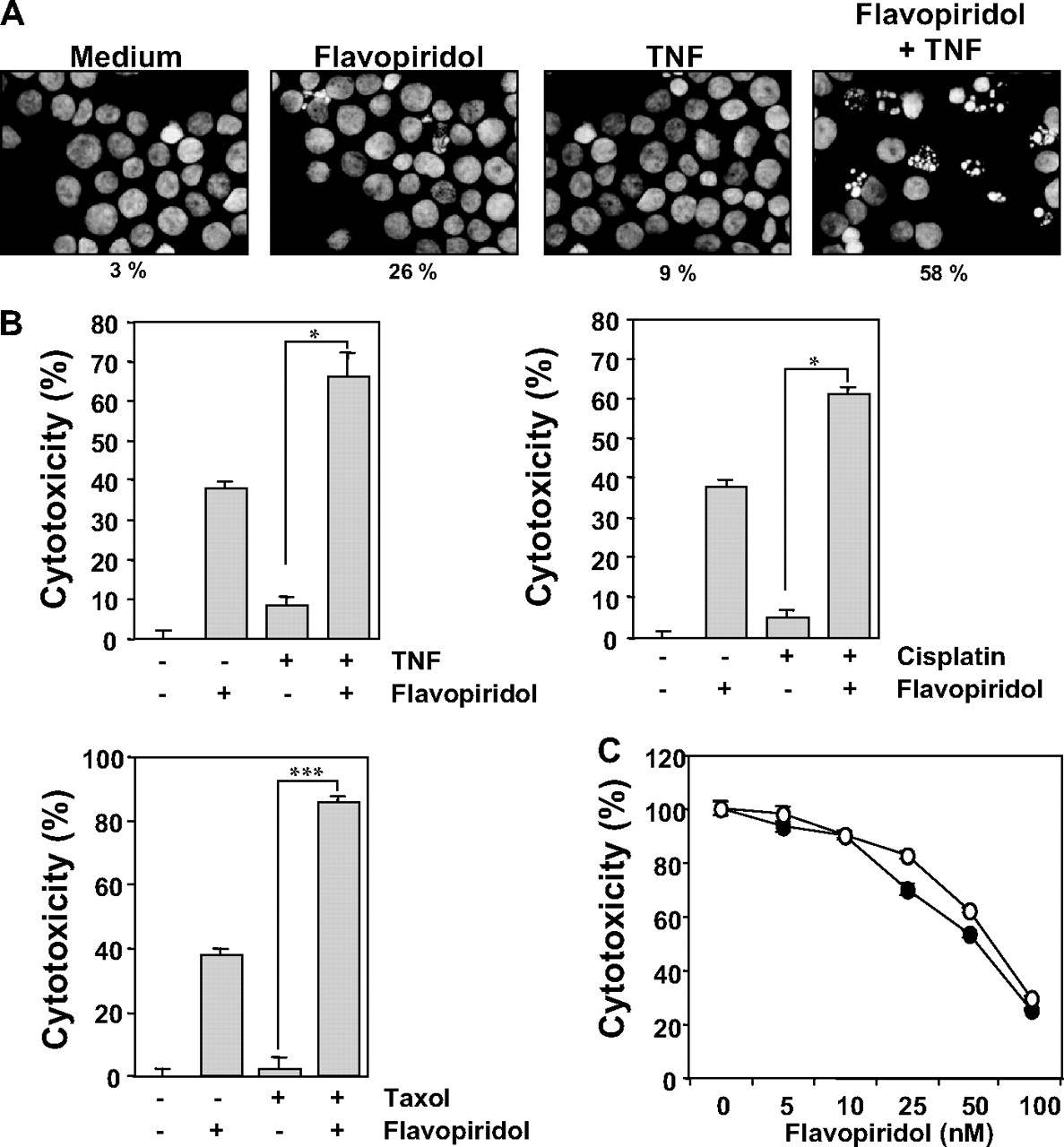
Flavopiridol suppressed cell proliferation and enhanced TNF-, cisplatin- and paclitaxel-induced apoptosis. A, HL60 cells were incubated with 100 nM flavopiridol for 8 h and treated with 1 nM TNF for 16 h. Cells were fixed with glutalaldehyde and stained with Hoechst 33342, and the nuclei were analyzed under a fluorescence microscope as described under Materials and Methods. B, HL60 (5000 cells/well) were seeded in triplicate in 96-well plates, pretreated with 100 nM flavopiridol for 8 h, and then incubated with 1 nM TNF, 30 μg/ml cisplatin, or 1 nM paclitaxel for 24 h. Cell viability was then analyzed by the MTT method as described under Materials and Methods. Data represent the mean of three measurements ± S.D. ***, p < 0.001; *, p < 0.05. C, HL60 (5000 cells/well) were seeded in triplicate in 96-well plates, and then treated with indicated concentration of flavopiridol for 72 h (○). Cells in another plate were exposed to flavopiridol for 8 h, washed with PBS and incubation continued in flavopiridol-free medium for total of 72 h (•). Cell viability was then analyzed by the MTT method as described under Materials and Methods.
Flavopiridol Repressed TNF-Induced Antiapoptotic Gene Products. TNF induces the expression of the antiapoptotic proteins IAP1/2 (Wang et al., 1998), XIAP (Stehlik et al., 1998), Bcl-2 (Catz and Johnson, 2001), Bcl-xL (Tamatani et al., 1999), and TRAF1 (Wang et al., 1998). We investigated whether flavopiridol can modulate the expression of these antiapoptotic gene products induced by TNF. Cells were pretreated with flavopiridol and then treated with TNF for the indicated times. Whole-cell extracts were prepared and analyzed by Western blot using the indicated antibodies. As shown in Fig. 4A, TNF induced these antiapoptotic proteins in a time-dependent manner, and flavopiridol suppressed the expression.
Flavopiridol Repressed the TNF-Induced ICAM-1, c-Myc, and c-Fos Expression. That ICAM-1 and c-Myc expression are induced by TNF has been reported (Duyao et al., 1992; van de Stolpe et al., 1994). The expression of c-Fos is tightly regulated by AP-1 (Rauscher et al., 1988). We investigated whether flavopiridol can modulate the TNF-induced tumorigenic gene products ICAM-1, c-Myc, and c-Fos. Cells were pretreated with flavopiridol and then exposed to TNF for the indicated times. Whole-cell extracts were prepared and analyzed by Western blot for the expression of ICAM-1, c-Myc, and c-Fos (Fig. 4B). TNF induced ICAM-1, c-Myc, and c-Fos expressions in a time-dependent manner, and flavopiridol blocked these expressions. The results further support the role of flavopiridol in blocking TNF-induced protein expression.
Flavopiridol Potentiates TNF-Induced Apoptosis. Because most of the antiapoptotic gene products induced by TNF are down-regulated by flavopiridol, we also investigated whether flavopiridol modulates TNF-induced apoptosis. Cells were pretreated with flavopiridol, treated with TNF, fixed and stained with Hoechst 33342, and then examined for nuclei status. We found that TNF alone and flavopiridol alone did not induce significant chromatin condensation; however, treatment with both reagents together markedly induced chromatin condensation (Fig. 5A). We also examined TNF-, cisplatin- and paclitaxel (Taxol)-induced cytotoxicity by the MTT method. TNF, cisplatin, and paclitaxel by themselves had minimal effects on the viability of the cells. Flavopiridol itself, although it had minimum effect at 8 h, was significantly cytotoxic to HL60 cells when treated for 32 h. However, pre-exposure to flavopiridol potentiated the effects of TNF, cisplatin, and paclitaxel (Fig. 5B).
The necessary length of treatment with flavopiridol was also determined. For this, HL60 cells were seeded and then exposed to flavopiridol for either 8 or 72 h and then examined for cell viability at the end of total 72 h. As shown in Fig. 5C, flavopiridol induced cytotoxicity equally whether exposed for 8 or 72 h, indicating that continuous presence of the drug is not needed.

Flavopiridol enhances TNF-induced Bid cleavage, cytochrome c release and caspase activation. A, HL60 cells were treated with 100 nM flavopiridol alone for the indicated times (left). HL60 cells were incubated with 100 nM flavopiridol for 8 h and treated with 1 nM TNF for the indicated times (right). Whole-cell extracts were prepared, and then Western blot analysis was performed using anti-Bid antibody. B, HL60 cells were incubated with 100 nM flavopiridol for 8 h and treated with 1 nM TNF for the indicated times. Cytoplasmic extracts were prepared as described under Materials and Methods and subjected to Western blot analysis using anti-cytochrome c antibody. C, HL60 cells were treated with 100 nM flavopiridol alone, 1 nM TNF, or both for the indicated times. Whole-cell extracts were prepared, and then Western blot analysis was performed using the indicated antibodies.
Flavopiridol Potentiates TNF-Induced Cytochrome c Release and Caspase Activation. Among the cytokines, TNF is one of the most potent inducers of apoptosis (Rath and Aggarwal, 1999). However, apoptotic effects of TNF require inhibition of protein synthesis in most cell lines (Sugarman et al., 1985). Numerous cell survival proteins have been identified that can suppress TNF-induced apoptosis. In the presence of cycloheximide, TNF is known to induce bid-cleavage through caspase-8 activation; this cleaved bid could induce cytochrome c release from mitochondria, which then could activate caspase-9, leading to caspase-3 activation and PARP cleavage (Rath and Aggarwal, 1999). We investigated whether suppression of TNF-induced cell survival proteins by flavopiridol leads to enhancement of TNF-induced caspase activation in HL60 cells. Cells were pretreated with flavopiridol, treated with TNF for the times indicated in the figure legend, and then examined for bid cleavage (Fig. 6A), cytochrome c release (Fig. 6B), caspase-9 activation (Fig. 6C, lane 1), caspase-3 activation (Fig. 6C, lane 2), and PARP cleavage (Fig. 6C, lane 3). TNF or flavopiridol alone had minimal effect, but the combination enhanced bid cleavage, cytochrome c release, caspase-9 activation, caspase-3 activation, and PARP cleavage in a time-dependent manner. These results thus suggest that flavopiridol pretreatment sensitizes the cells to TNF-induced caspase activation, most likely through the suppression of cell survival proteins shown in Fig. 4.
Flavopiridol Potentiated the Effect of TNF in Growth Inhibition of Tumor Cells. Whether flavopiridol potentiates the effect of TNF in a clonogenic assays was examined in H1299 and HL60 cells. Cells were exposed to different concentrations of flavopiridol alone or with TNF, cultured for 10 days, and then counted for the colonies. The exposure to flavopiridol resulted in dose-dependent reduction in colony formation compared with that of control cells (Fig. 7A). TNF enhanced the inhibition of colony formation induced by flavopiridol in H1299 (Fig. 7B) and HL60 cells (Fig. 7C). These results demonstrate that flavopiridol enhances the effect of TNF for inhibition of tumor colony formation.
Discussion
TNF is one of the most pleiotropic cytokines, mediating multiple cellular responses (Aggarwal, 2003). The proinflammatory effects of TNF are mediated through the activation of NF-κB (Aggarwal, 2003). Previous studies from our laboratory have shown that flavopiridol can abrogate TNF-induced NF-κB activation (Takada and Aggarwal, 2004). The current study was designed to investigate the effect of flavopiridol on the activation of other cellular responses to TNF. We found that this flavone abolished TNF-induced AP-1 activation and that of various MAPKs, suppressed the activation of Akt, abrogated the expression of antiapoptotic and tumorigenic gene products, and enhanced apoptosis. Flavopiridol also suppressed the AP-1 activation induced by LPS, doxorubicin, interleukin-1β, phorbol ester, okadaic acid, and H2O2.

Flavopiridol potentiates the effect of TNF for growth inhibition of tumor cells. A, H1299 (500 cells/well) were exposed to varying doses of flavopiridol alone and with 1 nM TNF; and incubated for an additional 10 days before counting colony numbers. B, a graphic representation of the data with H1299 cells. C, HL60 (1000 cells/well) were seeded in RPMI 1640 medium containing 0.33% of agarose, exposed to varying doses of flavopiridol alone and with 1 nM TNF and incubated for an additional 10 days before counting colony numbers. Determinations were made in triplicate.
We found that flavopiridol suppressed the activation of AP-1 induced by a wide variety of stimuli. How this flavone suppresses AP-1 activation is not clear. We found that flavopiridol suppressed nuclear translocation of c-Fos. In addition, TNF-induced JNK activation was also suppressed by this flavone. Both of these mechanisms may contribute to the suppression of AP-1 activation. AP-1 has been shown to regulate a wide range of cellular processes, including cell proliferation, death, survival, differentiation tumor promotion, and drug resistance (Shaulian and Karin, 2002). It is possible that some of the previously reported effects of flavopiridol are due to suppression of AP-1.
In current study, we used 100 nM flavopiridol for TNF-induced AP-1 suppression. Flavopiridol is currently undergoing phase III clinical trials for the treatment of various refractory neoplasms, both as a single agent and in combination with other antineoplastic drugs (Senderowicz, 2002). The dose used in the present report corresponds to the clinically achievable pharmacological concentrations of the flavopiridol (Senderowicz, 2002; Thomas et al., 2002). The range of doses of flavopiridol used in our study (50-100 nM) are comparable with those shown to be active in other tumor systems also (Bible and Kaufmann, 1996; Patel et al., 1998; Takada and Aggarwal, 2004).
We found that flavopiridol also suppressed TNF-induced p44/p42 MAPK activation, which has been linked to cell proliferation (Aggarwal, 2003). Antiproliferative effects linked to flavopiridol (Senderowicz and Sausville, 2000) could also be due to inhibition of this kinase. In addition, we found that flavopiridol inhibited activation of Akt and JNK1. Li et al. (2004) reported that inhibition of Akt activation blocked tumor promoter 5-MCDE-induced activation of JNK, p44/p42 MAPK, and AP-1 activation, and the dominant-negative mutant of Akt blocked JNK1 and AP-1 activation by 5-MCDE (Li et al., 2004). B[a]PDE-induced AP-1 transactivation has also been shown to be inhibited by pretreatment of cells with the PI-3K inhibitors wortmannin and LY294002, and an overexpression of the dominant-negative mutant of PI-3K impaired B[a]PDE-induced activation of PI-3K and Akt and AP-1 transactivation (Li et al., 2004). Furthermore, they also found that an overexpression of the dominant-negative Akt mutant blocked B[a]PDE-induced activation of Akt, AP-1, and JNK. Thus suppression of Akt by flavopiridol may also contribute to its chemopreventive activities.
We found that flavopiridol blocked AP-1-dependent reporter gene expression. Several genes that are involved in various diseases are regulated by AP-1. These include cell cycle-regulating proteins, inflammatory cytokines, metalloproteinases, angiogenesis, and invasion (Bakiri et al., 2000; Passegué and Wagner, 2000; Szabowski et al., 2000). Thus it is possible that flavopiridol mediates its immunosuppressive effects through suppression of AP-1-regulated genes. Indeed, flavopiridol has been reported to suppress cyclooxygenase-2, cyclin D1, and metaroproteinase-9 (Takada and Aggarwal, 2004). Because AP-1-regulated gene products have also been implicated in tumorigenesis, flavopiridol may prove useful in suppressing tumorigenesis.
We found that several gene products linked with suppression of apoptosis and tumorigenesis were also down-regulated by flavopiridol. This finding correlated with the increase in apoptosis induced by TNF and chemotherapeutic agents. Bid cleavage, cytochrome c release, activation of capase-3 and capase-9, and PARP cleavage were all enhanced when TNF was used in combination with flavopiridol. These results are consistent to those reported previously in which flavopiridol was found to potentiate the apoptotic effects of TNF, TNF-related apoptosis-inducing ligand, and phorbol ester (Cartee et al., 2002; Kim et al., 2003). Although most of the studies with flavopiridol were carried out using TNF, our results indicate that this drug can modulate the apoptosis induced by other agents. Overall, our results indicate that flavopiridol could inhibit multiple pathways activated by TNF, modulate the gene expression, and increase apoptosis. This provides the rationale for combining flavopiridol with cytokines and chemotherapeutic agents for the treatment of cancer.
Acknowledgments
We thank Michael Worley for providing valuable editorial comments. Dr. Takada is an Odyssey Program Special Fellow at The University of Texas M. D. Anderson Cancer Center. Dr. Aggarwal is Ransom Horne, Jr., Distinguished Professor of Cancer Research.
Footnotes
- Received August 30, 2007.
- Accepted February 19, 2008.
This work was supported by the Clayton Foundation for Research (to B.B.A.), National Cancer Institute core grant CA16672, and the Theodore N. Law Award for Scientific Achievement Fund from The University of Texas M. D. Anderson Cancer Center (to Y.T.).
ABBREVIATIONS: CDK, cyclin-dependent kinase; TNF, tumor necrosis factor; NF-κB, nuclear factor-κB; AP-1, activator protein-1; JNK, c-Jun N-terminal kinase; MAPK, mitogen-activated protein kinase; PMA, phorbol 12-myristate 13-acetate; ICAM, intercellular adhesion molecule-1; PARP, poly(ADP-ribose) polymerase; EMSA, electrophoretic mobility shift assay; PAGE, polyacrylamide gel electrophoresis; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium; LPS, lipopolysaccharide; IAP, inhibitor-of-apoptosis protein; XIAP, X-chromosome-linked inhibitor-of-apoptosis protein; 5-MCDE, (±)-anti-5-methylchrysene-1,2-diol-3,4-epoxide; B[a]PDE, (±)-benzo[a]pyrene-7,8-diol-9,10-epoxide.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}