


Abstract
The vitamin D Receptor (VDR) belongs to the superfamily of steroid/thyroid hormone receptors that is activated by 1α,25-dihydroxyvitamin D3. Traditional targets for 1α,25-dihydroxyvitamin D3 action include tissues involved in the maintenance of calcium homeostasis and bone development and remodeling. Peroxisome proliferator-activated receptor γ coactivator-1α (PGC-1α), a transcriptional coactivator that plays a role in mitochondrial biogenesis and energy metabolism, is predominantly expressed in kidney, heart, liver, and skeletal muscle. Because VDR and PGC-1α display an overlapping pattern of expression, we investigated the possibility that PGC-1α could serve as a coactivator for VDR. Transient cotransfection assays demonstrate that PGC-1α augments ligand-dependent VDR transcription when either full-length VDR or Gal4 DNA binding domain-VDR-ligand binding domain chimeras were analyzed. Furthermore, mammalian two-hybrid assays, coimmunoprecipitation analyses, and biochemical coactivator recruitment assays demonstrate a ligand-dependent interaction between the two proteins both in cells and in vitro. The coactivation potential of PGC-1α requires an intact AF-2 domain of VDR and the LXXLL motif in PGC-1α. Taken together, these results indicate that PGC-1α serves as a coactivator for VDR.
The hormonally active form of vitamin D, 1α,25-dihydroxyvitamin D3 (1,25(OH)2D3) or calcitriol, and its synthetic analogs mediate their biological actions via the vitamin D receptor (VDR; NR1I1), a member of the nuclear receptor superfamily (van Leeuwen et al., 2001). The actions of 1,25(OH)2D3 include maintenance of calcium homeostasis and bone development and remodeling (DeLuca, 2004). In addition, vitamin D3 has also been shown to function in the differentiation of leukemic cells (Abe et al., 1981; Bar-Shavit et al., 1983; Mangelsdorf et al., 1984), inhibit the growth and proliferation of various cancerous cells (Colston et al., 1981; Dokoh et al., 1984; Skowronski et al., 1993), and possess immunosuppressive activity (Manolagas et al., 1985). The physiological and pharmacological actions of these compounds have indicated the application of VDR ligands in inflammation, autoimmune diseases, osteoporosis, cancers, and dermatological indications (Pinette et al., 2003).
The VDR protein is modular in nature and can be functionally divided into three distinct regions: an N-terminal region containing a ligand-independent activation function-1, a highly conserved central region containing the DNA binding domain (DBD), and the C-terminal region of the receptor containing a multifunctional domain harboring the ligand binding domain (LBD), the RXR heterodimerization motif, and a ligand-dependent activation function-2 (AF2) (Mangelsdorf et al., 1995; Giguere, 1999; Burris, 2001). The binding of a ligand to the receptor induces a conformational change within the LBD of VDR, resulting in the release of bound corepressor proteins and permitting the association of coactivator proteins that mediate a series of events leading to transcriptional activation of target genes (Glass and Rosenfeld, 2000; Savkur and Burris, 2004). The traditional target tissues exhibiting hormone-dependent VDR actions include tissues such as the bone, kidney, small intestine, parathyroid glands, skeletal muscle, heart, skin, breast, colon, prostate, and gonads. VDR response elements (VDREs) have been identified in the promoters of vitamin D3-responsive genes that are essential for the hormonal responsiveness of the tissues including calbindin-D9K (Darwish and DeLuca, 1992), osteocalcin (Kerner et al., 1989; Demay et al., 1992a), osteopontin (Noda et al., 1990; Ozono et al., 1990), parathyroid hormone (Demay et al., 1992b), calbindin-D28K (Gill and Christakos, 1993), the calcidiol (25-hydroxyvitamin D3) 24-hydroxylase (Zierold et al., 1994, 1995) and the insulin-like growth factor binding protein-3 (Peng et al., 2004).
It has been demonstrated that PGC-1α is a potent transcription coactivator for several nuclear receptors and transcription factors (Puigserver et al., 1998; Wu et al., 1999, 2002, 2003; Knutti et al., 2000; Tcherepanova et al., 2000; Vega et al., 2000; Delerive et al., 2002; Huss et al., 2002; Oberkofler et al., 2003; Kanaya et al., 2004; Lau et al., 2004; Zhang et al., 2004; Savkur et al., 2005). PGC-1α is primarily expressed in kidney, heart, and skeletal muscle tissues (Esterbauer et al., 1999) and induces mitochondrial biogenesis when expressed ectopically in skeletal and cardiac myocytes (Lehman et al., 2000; Wu et al., 1999). Because both VDR and PGC-1α possess an overlapping tissue distribution, we reasoned that PGC-1α might serve as a potential coactivator of VDR and enhance VDR-mediated transcription. In this study, we demonstrate that PGC-1α augments ligand-mediated VDR transcription and serves as a coactivator of VDR. The coactivation activity of PGC-1α requires an intact AF-2 domain of VDR and the LXXLL motif in PGC-1α. Mammalian two-hybrid analyses, coimmunoprecipitation analyses, and biochemical coactivator recruitment assays demonstrate a ligand-dependent interaction between the two proteins both in cultured cells and in vitro.
Materials and Methods
Plasmid Construction. Full-length VDR and VDRΔAF2 were constructed by cloning PCR-amplified DNA fragments into the BamHI and EcoRV sites of pcDNA3.1D/V5-His-TOPO (Invitrogen, Carlsbad, CA). Gal4–VDR-LBDΔAF2 was constructed from Gal4–VDR-LBD (pM–VDR-LBD; encompassing amino acids 89–427 of VDR) (Bettoun et al., 2003) by PCR amplification, and subcloned into the EcoRI and XbaI sites of the pM vector (BD Biosciences Clontech, Palo Alto, CA). Full-length VDR and VDRΔAF2 fused to the Gal4-DBD were constructed by PCR amplification and cloned into the pM vector between the EcoRI and XbaI sites. The expression plasmid for PGC-1α has been described previously (Delerive et al., 2002). The plasmid expressing poly-His-tagged VDR-LBD (encompassing amino acids 89–427 of VDR) has been described previously (Bettoun et al., 2003). Plasmids expressing GST–PGC-1α and VP16–PGC-1α fusion constructs (encoding amino acids 100–411 of PGC-1α) have been described previously (Wu et al., 2003). The OPN-tk-Luc reporter plasmid containing three copies of the osteopontin VDRE has been described previously (Bettoun et al., 2003). The pG5-Luc reporter plasmid was purchased from Promega (Madison, WI). The sequence of all plasmids was verified by DNA sequencing.
Cell Culture and Transient Cotransfection Assays. HEK293 cells were maintained in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin at 37°C under 5% CO2. Before transfections, cells were plated into 96-well plates at a density of 25 × 103 cells/well in the medium supplemented with 10% dextran charcoal-stripped fetal bovine serum. Transfections were performed using FuGENE 6 (Roche Molecular Biochemicals, Indianapolis, IN). Each transfection contained 25 ng of luciferase reporter plasmid and various combinations of other expression plasmids as described in the figure legends. Twenty-four hours after transfection, fresh media containing either 0.1% DMSO or 1,25(OH)2D3 (100 nM in 0.1% DMSO) was added as indicated. Twenty-four hours after treatment, the luciferase activity was measured using the standard luciferase substrate reagents (Promega).
Adenovirus Infection and Gene Expression. Adenoviral constructs expressing PGC-1α and green fluorescent protein (GFP) have been described previously (Savkur et al., 2005). Caco-2 cells (American Type Culture Collection, Manassas, VA) were plated in six-well plates at a density of 5 × 105 cells/well. Cells were infected at an approximate multiplicity of infection of 10 to achieve 100% infection. Forty-eight hours after infection, fresh media containing either 0.1% DMSO or 1,25(OH)2D3 (100 nM in 0.1% DMSO) was added as indicated. Twenty-four hours after treatment, cells were harvested and RNA and cDNA were prepared using ABI 6100 reagents and methodologies (Applied Biosystems, Foster City, CA). The expression of 1,25 dihydroxyvitamin D3-24 hydroxylase (24-hydroxylase) was quantified using standard quantitative-PCR techniques and normalized to the levels of GAPDH.
Immunoprecipitation Assay. Interaction of VDR and PGC-1α in mammalian cells was assessed using immunoprecipitation assays as described previously for PGC-1α and FXR (Savkur et al., 2005) with the following modifications. HEK293 cells plated in 150-mm dishes at a density of 15 × 106 cells/dish were transfected with plasmids expressing amino-terminal myc-His-tagged PGC-1α or full-length VDR alone or in combination. After 24 h of treatment in the absence or presence of ligand [0.1% DMSO or 1,25(OH)2D3 (1 μM in 0.1% DMSO)], cells were lysed followed by immunoprecipitation with a monoclonal anti-myc antibody (Santa Cruz Biotechnology, Inc., Santa Cruz, CA). Samples were then subjected to Western blot analysis using a monoclonal anti-VDR antibody (Santa Cruz Bio-technology, Inc.).
Coactivator Recruitment Assay. Biochemical interaction between polyhistidine-tagged VDR-LBD and GST–PGC-1α was assayed using the AlphaScreen technology as described previously (Savkur et al., 2005). In brief, the assay was performed in white, low-volume, 384-well plates using a final volume of 15 μl containing final concentrations of 100 nM baculovirus-expressed poly-Histagged VDR-LBD protein (encompassing amino acids 89–427 of VDR), 5 nM GST–PGC-1α fusion protein (encoding amino acids 100–411 of PGC-1α), and 10 μg/ml of both Ni+2-chelate donor beads and anti-GST acceptor beads in an assay buffer containing 25 mM HEPES, pH 7.0, 100 nM NaCl, 0.1% bovine serum albumin, and 2 mM dithiothreitol. Different concentrations of 1,25(OH)2D3 dissolved in DMSO (maintained at a final concentration of 1%) were added as indicated. Dose-response curves were analyzed using Prism software (GraphPad Software Inc., San Diego, CA).
Results
To determine whether PGC-1α functions as a coactivator and enhances VDR-mediated transcription, a transient transfection assay was performed using a plasmid that expresses full-length VDR and a luciferase reporter gene driven by the herpes simplex virus-thymidine kinase promoter preceded by three repeats of the VDRE derived from the human osteopontin (OPN) gene (5′-GGGTCGTATGGTTCA-3′) (Ozono et al., 1990). Cotransfection was performed in the absence or presence of the VDR ligand, 1,25(OH)2D3 (Fig. 1A). Increasing amounts of a plasmid expressing PGC-1α in the absence of the ligand resulted in a 1.5-fold activation of the reporter gene at the highest concentration of the transfected plasmid (50 ng). Addition of 1,25(OH)2D3 resulted in an approximate 15-fold increase in the transcription of the reporter gene in the absence of PGC-1α. Increasing amounts of the PGC-1α–expressing plasmid resulted in an approximate 100-fold activation of the reporter in the presence of 1,25(OH)2D3. Thus, efficacious coactivation of VDR by PGC-1α is highly dependent on the presence of the ligand.
Because the interaction between PGC-1α and other NRs has been mapped to the LBD of the nuclear receptors, we created a chimeric receptor by fusing the LBD of VDR to the yeast Gal4 DBD to further study the coactivator function of PGC-1α. This Gal4-DBD/VDR-LBD fusion receptor (Gal4–VDR-LBD) was coexpressed with a cognate Gal4 responsive reporter gene in the absence or presence of 1,25(OH)2D3 (Fig. 1B). Overexpression of PGC-1α has no effect on the basal activity of the reporter gene, even at the highest concentration of the expressed PGC-1α (50 ng). In the absence of transfected PGC-1α, transcription from the reporter gene increases approximately 5-fold in the presence of the ligand. Overexpression of PGC-1α resulted in an increase in transcription to approximately 500-fold in the presence of the ligand. Thus, dose-dependent augmentation of the chimeric Gal4–VDR-LBD mediated transcription by PGC-1α confirms that PGC-1α serves as coactivator for VDR-mediated transcription in a ligand-dependent manner. Furthermore, our results also demonstrate that the LBD and hinge region of VDR (encompassing amino acids 89–427) is sufficient for the ligand-dependent coactivation.

PGC-1α coactivates VDR-mediated transcription in a ligand-dependent manner. A, HEK293 cells were cotransfected with 25 ng of a plasmid expressing full-length VDR, 25 ng of a luciferase reporter gene driven by the minimal thymidine kinase promoter preceded by three copies of the VDRE derived from the human osteopontin gene (OPN-tk-Luc), and increasing amounts of a plasmid expressing PGC-1α (2.5, 5, 10, 25, and 50 ng). Twenty-four hours after transfection, cells were treated with either 0.1% DMSO or 1,25(OH)2D3 (in 0.1% DMSO) as indicated. Twenty-four hours after treatment, the luciferase activity was measured using the standard luciferase substrate reagents. B, HEK293 cells were cotransfected with 25 ng of a plasmid expressing Gal4–VDR-LBD, 25 ng of the pG5-Luc reporter plasmid, and increasing amounts of a plasmid expressing PGC-1α (2.5, 5, 10, 25, and 50 ng). Twenty-four hours after transfection, cells were treated with either 0.1% DMSO or 1,25(OH)2D3 as indicated. Twenty-four hours after treatment, the luciferase activity was measured using the standard luciferase substrate reagents. All experiments were performed in triplicate, and the data are displayed as the mean ± S.D. of a single experiment, representative of at least three independent experiments. C, after infection with an adenoviral vector directing the expression of either PGC-1α or GFP, Caco-2 cells were treated with either 0.1% DMSO or 1,25(OH)2D3 as indicated. After 24 h of treatment, the expression of an endogenous VDR target gene, 1,25-dihydroxyvitamin D3-24 hydroxylase (24-hydroxylase) was assessed by quantitative-reverse transcription PCR and normalized to GAPDH. *, significantly different (P < 0.05 using the t test) from control (GFP) in the presence of the ligand.
To determine whether the increase in PGC-1α expression would affect the expression of a VDR target gene in a cell line expressing endogenous VDR, an adenoviral vector directing the expression of either PGC-1α or GFP was used to infect Caco-2 cells. After infection, cells were treated with 1,25(OH)2D3 followed by assessment of expression of the calcidiol (25-hydroxyvitamin D3-24-hydroxylase) mRNA. As depicted in Fig. 1C, treatment of the cells with 1,25(OH)2D3 results in a 570-fold induction in the expression of 24-hydroxylase mRNA in control cells (GFP-adenoviral-infected cells). In the cells in which PGC-1α was overexpressed, the levels of 24-hydroxylase mRNA were enhanced by 60%, consistent with our observation that PGC-1α functions as a coactivator of VDR. PGC-1α overexpression alone, in the absence of ligand, did not result in enhancement of 24-hydroxylase mRNA expression, confirming the ligand dependence of the coactivation.
It has previously been demonstrated that the C-terminal LBD of VDR that contains the activation function-2 (AF-2) domain is required for its response to 1,25(OH)2D3 (Nayeri et al., 1996; Nakajima et al., 1998). To investigate whether the AF-2 domain was necessary for the ligand-dependent coactivation of VDR by PGC-1α, cotransfection assays were performed using either a full-length VDR expression vector lacking the AF-2 or a Gal4–VDR-LBD chimeric receptor lacking the AF-2 in combination with their cognate luciferase reporter plasmids. As illustrated in Fig. 2A, wild-type VDR mediated transcriptional activity was increased in the presence of the ligand, 1,25(OH)2D3. However, deletion of helix 12 of VDR abolished the transcriptional activity in response to the ligand and also to overexpression of PGC-1α. The requirement for the AF-2 domain was also reiterated when the wild-type Gal4–VDR-LBD chimeric receptor and the helix 12 deletion constructs were assessed with the ligand and overexpressed PGC-1α (Fig. 2B). Thus, an intact AF-2 domain is necessary for VDR-mediated transcription in response to both the ligand and PGC-1α coactivation.

The AF-2 domain of VDR is required for ligand-dependent coactivation by PGC-1α. A, HEK293 cells were cotransfected with 25 ng of plasmids expressing full-length VDR or the AF-2 deletion (VDRΔAF2), 25 ng of the OPN-tk-Luc reporter plasmid, and 25 ng of a plasmid expressing PGC-1α in the absence or presence of 1,25(OH)2D3 as described in Fig. 1. The luciferase activity was measured 24 h after treatment. B, HEK293 cells were cotransfected with 25 ng of plasmids expressing chimeric wild-type Gal4–VDR-LBD or the AF-2 deletion (Gal4–VDR-LBDΔAF2), 25 ng of the pG5-Luc reporter plasmid, and 25 ng of a plasmid expressing PGC-1α in the absence or presence of 1,25(OH)2D3 as described in Fig. 1. The luciferase activity was measured 24 h after treatment. All experiments were performed in triplicate, and the data are displayed as the mean ± S.D. of a single experiment, representative of at least three independent experiments. Immunoblot analyses revealed no significant deviation in the expression of wild-type and mutant receptors (data not shown).
The binding of a ligand to the LBD of NRs induces a conformational change that permits the recognition of specific motifs contained within coactivator proteins. This motif, commonly referred to as the NR box or LXXLL motif, is necessary and sufficient to permit most NR-coactivator interactions (Heery et al., 1997; Savkur and Burris, 2004). Previous studies have demonstrated that an intact LXXLL motif in PGC-1α is required for its coactivation of several NRs (Tcherepanova et al., 2000; Vega et al., 2000; Knutti et al., 2001; Delerive et al., 2002; Wu et al., 2002, 2003; Oberkofler et al., 2003; Schreiber et al., 2003; Savkur et al., 2005). To determine whether the NR box of PGC-1α mediates the ligand-dependent coactivation of VDR, cotransfection assays were performed with the chimeric Gal4–VDR-LBD and plasmids expressing the wild-type PGC-1α or a mutant PGC-1α in which the leucines in the LXXLL motif were mutated to alanines (LXXLL→AXXAA). Expression of the wild-type PGC-1α resulted in a 25-fold enhancement of transcriptional activity in the presence of 1,25(OH)2D3 (Fig. 3A). However, mutation of the central LXXLL (L2) motif of PGC-1α resulted in an ∼90% reduction in the coactivation potential of PGC-1α. To ascertain whether the LXXLL motif of PGC-1α mediated ligand-dependent coactivation of the full-length VDR protein, cotransfection assays were performed with either the wild-type PGC-1α or the PGC-1αAXXAA mutant. As observed with the chimeric Gal4–VDR-LBD receptor, coexpression of wild-type PGC-1α with the full-length VDR resulted in a 4.5-fold enhancement of transcriptional activity in the presence of 1,25(OH)2D3 (Fig. 3B). However, coexpression of the PGC-1αAXXAA mutant resulted in a ∼75% reduction in the coactivation potential compared with the wild-type PGC-1α. The observation that the L2 LXXLL motif of PGC-1α is the critical determinant in receptor recognition is consistent with our previous studies with RXR, TRβ, PPARγ, and FXR (Delerive et al., 2002; Wu et al., 2002, 2003; Savkur et al., 2005).
To determine whether the coactivation of VDR by PGC-1α is a result of a direct interaction between the two proteins in vivo, a mammalian two-hybrid assay was performed using a plasmid expressing a Gal4-DBD–full-length VDR chimera (Gal4–VDR) and a VP16–PGC-1α chimera (spanning amino acids 100–411 of PGC-1α) along with the luciferase reporter plasmid harboring five copies of the Gal4 response element (Fig. 4A). Wild-type Gal4–VDR mediated transcriptional activity was increased in the presence of 1,25(OH)2D3. This activation was further enhanced by ∼3.5-fold by expression of VP16–PGC-1α, indicating that the region of PGC-1α encompassing amino acids 100 to 411, spanning the critical LXXLL motif, was sufficient to mediate the ligand-dependent interaction with VDR. However, deletion of helix 12 of VDR abolished the interaction with PGC-1α, demonstrating that the AF-2 domain of VDR was essential for this interaction. The interaction of VDR and PGC-1α was also assessed by coimmunoprecipitation. Amino-terminal myc-tagged PGC-1α was cotransfected with full-length VDR into HEK293 cells. After a 24-h treatment either in the absence or presence of 1,25(OH)2D3, the cells were lysed and immunoprecipitated with a myc antibody, and the presence of VDR in the immunoprecipitate was revealed by Western blotting with a monoclonal antibody against VDR. The results shown in Fig. 4B indicate that VDR and PGC-1α do interact with each other in vivo and consistent with our previous results, the interaction is ligand-dependent. To examine whether this was a result of a direct interaction, both proteins were expressed and their ability to interact was assessed in a biochemical assay. A biochemical coactivator recruitment assay using the Alpha-Screen technology was performed using E. coli expressed GST–PGC-1α fusion protein (encompassing amino acids 100–411 of PGC-1α), and the hinge/LBD of VDR that was expressed in a baculovirus as a poly-His-tagged fusion protein. As illustrated in Fig. 4C, increasing concentrations of 1,25(OH)2D3 resulted in a dose-dependent increase in the interaction between the VDR LBD and PGC-1α. The results from the coactivator recruitment assay and the cotransfection experiments using the Gal4–VDR-LBD chimeric receptor also demonstrate that the hinge/LBD of VDR is sufficient for the PGC-1α-mediated coactivation of VDR both in vitro and in vivo. Thus, coactivation of VDR by PGC-1α is mediated via a direct interaction that requires an intact AF-2 domain of VDR and the LXXLL motif in PGC-1α.
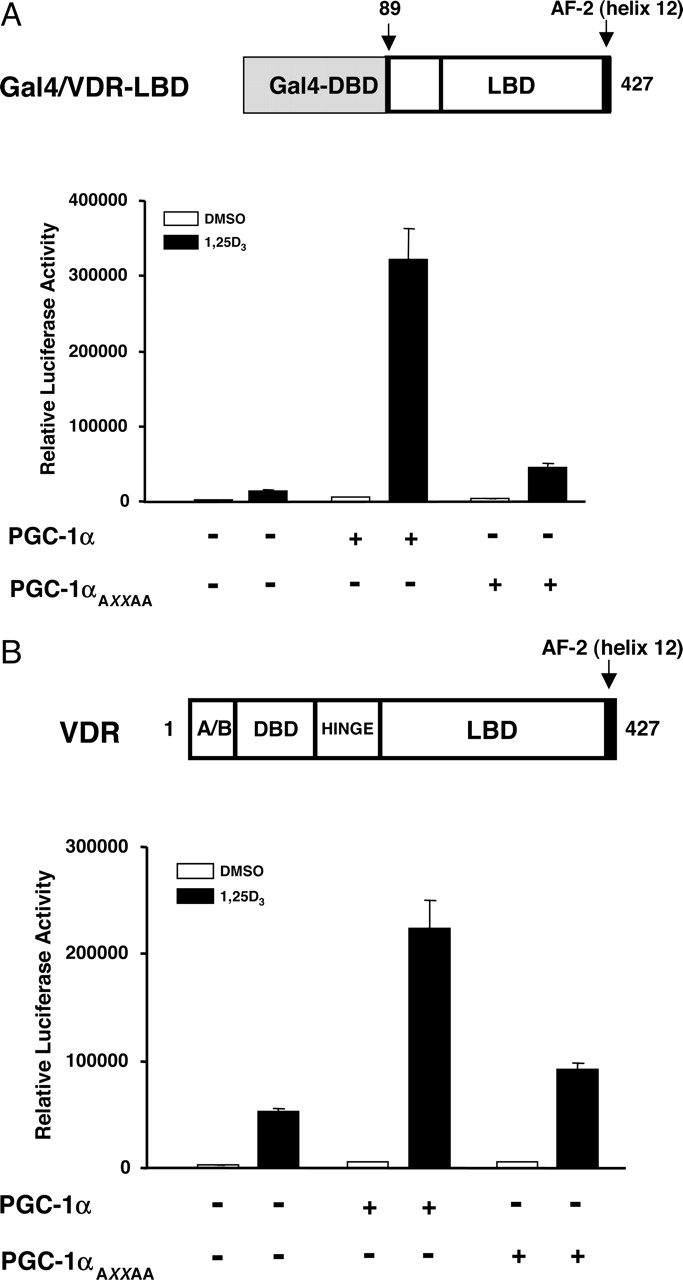
The NR box of PGC-1α is essential for the coactivation of VDR. A, HEK293 cells were cotransfected with 25 ng of the plasmid expressing wild-type Gal4–VDR-LBD, 25 ng of the pG5-Luc reporter plasmid, and 25 ng of plasmids expressing either the wild-type PGC-1α or the mutated PGC-1α harboring an AXXAA substitution of the LXXLL motif in the absence or presence of 1,25(OH)2D3 as described in Fig. 1. The luciferase activity was measured 24 h after treatment. B, HEK293 cells were cotransfected with 25 ng of a plasmid expressing full-length VDR, 25 ng of the OPN-tk-Luc reporter plasmid, and 25 ng of plasmids expressing either the wild-type PGC-1α or the PGC-1αAXXAA mutant in the absence or presence of 1,25(OH)2D3 as described in Fig. 1. The luciferase activity was measured 24 h after treatment. All experiments were performed in triplicate, and the data are displayed as the mean ± S.D. of a single experiment, representative of at least three independent experiments. Expression of mutant PGC-1α was compared with the wild-type by Western blots and showed no significant difference in expression (data not shown).
Discussion
Vitamin D3 exerts a variety of biological functions such as calcium homeostasis, cell proliferation, and cell differentiation. Most of the actions are mediated through the transcriptional control of the target genes by the action of the VDR. Binding of a ligand to the VDR results in the release of the existing corepressor proteins and permits the recruitment of coactivator proteins that results in a series of events leading to transcriptional regulation of target genes (van Leeuwen et al., 2001). The target organs exhibiting hormone-dependent VDR actions include tissues such as bone, kidney, cardiac and skeletal muscle. In this study, we demonstrate that PGC-1α, which exhibits an overlapping expression pattern with VDR, serves as an efficient ligand-dependent coactivator of VDR. PGC-1α coactivation of VDR activity in either full-length or Gal–VDR-LBD chimeric receptor is dependent on the presence of the ligand. This mode of action of PGC-1α with VDR differs from its interaction with PPARγ, which is primarily ligand-independent (Puigserver et al., 1998) but still displays a significant ligand-dependent component (Wu et al., 2003). Other receptors such as retinoid-related orphan receptor-α and estrogen-related receptor-α also display a ligand-independent mode of interaction with PGC-1α (Schreiber et al., 2003; Lau et al., 2004); however, the majority of receptors that have now been examined display a ligand-dependent mode of interaction with PGC-1α. The NRs that have been shown to interact with PGC-1α in a ligand-dependent manner include TRα, TRβ, ERα, RXR, FXR, and PPARα; however, some of these receptors still retain a limited degree of ligand independence (Tcherepanova et al., 2000; Vega et al., 2000; Delerive et al., 2002; Wu et al., 2002; Savkur et al., 2005). Our results demonstrate that PGC-1α interacts with either the full-length VDR or the Gal–VDR-LBD chimeric receptor in a ligand-dependent manner. Both systems reveal that the coactivation requires the AF-2 domain of the receptor and the NR box of PGC-1α. Analysis of the amino acid sequence of PGC-1α indicated the presence of three functional NR boxes comprising the conserved LXXLL motifs (L1, L2, and L3) (Knutti et al., 2001; Huss et al., 2002). However, it has been demonstrated that the second LXXLL motif (L2; amino acids 144–148) is the primary contributor to the interaction of PGC-1α with several nuclear receptors, including TRβ1, RXR, FXR, PPARα, PPARγ, glucocorticoid receptor, and ERα (Knutti et al., 2000; Tcherepanova et al., 2000; Vega et al., 2000; Delerive et al., 2002; Wu et al., 2002, 2003; Savkur et al., 2005). Consistent with these observations, we demonstrate that the intact L2 LXXLL motif in PGC-1α plays a critical role in its coactivation of VDR. Furthermore, results from the one-hybrid assay using the Gal–VDR-LBD chimeric receptor and the coactivator recruitment assay using the poly-His-tagged VDR-LBD fusion protein indicate that the hinge/LBD of VDR is sufficient for its coactivation by PGC-1α.
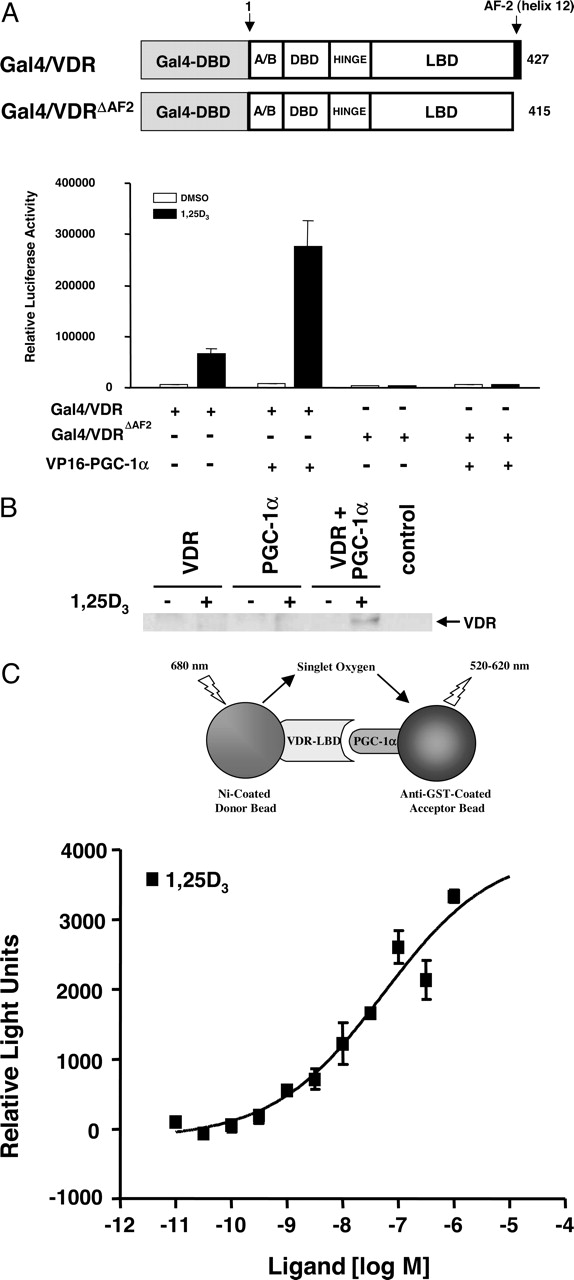
VDR and PGC-1α directly interact in a ligand-dependent manner both in vivo and in vitro. A, cotransfection and measurement of luciferase activity was performed as described in Fig. 1 but with 25 ng of the plasmid expressing wild-type Gal4–VDR or Gal4–VDRΔAF2 mutant, 25 ng of the pG5-Luc reporter plasmid, and 25 ng of a VP16–PGC-1α (amino acids 100–411 of PGC-1α) expressing plasmid in the absence or presence of 1,25(OH)2D3. All experiments were performed in triplicate, and the data are displayed as the mean ± S.D. of a single experiment, representative of at least three independent experiments. B, HEK293 cells were untransfected (control) or transfected with a plasmid expressing full-length VDR (VDR) or myc-tagged PGC-1α alone (PGC-1α) or with both the plasmids (VDR+PGC-1α). After 24 h of treatment in the absence (-) or presence (+) of 1,25(OH)2D3 cells were lysed, followed by immunoprecipitation with a monoclonal myc-antibody. The samples were then subjected to Western blotting using a monoclonal VDR antibody. C, E. coli-expressed GST–PGC-1α (amino acids 100–411 of PGC-1α) was incubated with baculovirus-expressed poly-His-tagged VDR-LBD (amino acids 89–427 of VDR) in 384-well plates using a final volume of 15 μlinthe presence of increasing concentration of 1,25(OH)2D3. Assay plates were covered with a clear seal and incubated in the dark for 2 h, after which the plates were read for 1 s/well in a Fusion microplate analyzer (PerkinElmer Life and Analytical Sciences) using the manufacturer's standard AlphaScreen detection protocol. All experiments were performed in triplicate, and the data are displayed as the mean ± S.D. of a single experiment, representative of at least three independent experiments.
PGC-1α is predominantly expressed in the kidney, liver, and skeletal and cardiac muscle (Esterbauer et al., 1999) and induces mitochondrial biogenesis when overexpressed in skeletal and cardiac myocytes (Wu et al., 1999; Lehman et al., 2000). Furthermore, expression of PGC-1α is regulated in skeletal muscle cells (Handschin et al., 2003). It is interesting that VDR has been demonstrated to play a role in the development of skeletal muscle (Demay, 2003; Endo et al., 2003) and in the regulation of myocardial development (O'Connell et al., 1997). Thus, our results demonstrating coactivation of VDR by PGC-1α suggest an additional role for PGC-1α in regulating the genes involved in skeletal and cardiac development that are under the influence of VDR. The expression of VDR and PGC-1α also overlaps in the kidney. Renal VDR is required for vitamin D3-mediated reabsorption of calcium and plays an important role in the regulation of serum calcium (Healy et al., 2003). The kidney is also an important tissue for the regulation of the metabolism of 1,25(OH)2D3, because the expression of several genes, such as the vitamin D-dependent calcium binding proteins (calbindin-D9K and calbindin-D28K) and the 24-hydroxylase are regulated by VDR (Darwish and DeLuca, 1992; Gill and Christakos, 1993; Zierold et al., 1994, 1995). Thus, coactivation of VDR by PGC-1α suggests that PGC-1α could play an additional role in VDR-mediated regulation of the genes involved in calcium metabolism.
In summary, our data suggest that PGC-1α acts as an authentic coactivator for VDR. Our results demonstrate that PGC-1α modulation of VDR activity is highly ligand-dependent and requires an intact AF-2 domain of VDR and the LXXLL motif of PGC-1α. Given the role of VDR in kidney as well as cardiac and skeletal muscle development, the expression of PGC-1α in these tissues suggests the potential involvement of PGC-1α in a wide array of physiological pathways involved in the regulation of genes during the development of these tissues and organs. Furthermore, because the expression of PGC-1α is regulated by environmental stimuli, such as diet and lower temperatures, it follows that the function of VDR could be influenced in response to these external stimuli in the tissues that exhibit an overlap in the expression of PGC-1α.
Footnotes
-
Article, publication date, and citation information can be found at http://molpharm.aspetjournals.org.
-
doi:10.1124/mol.105.012708.
-
ABBREVIATIONS: 1,25(OH)2D3, 1α,25-dihydroxyvitamin D3; OPN, osteopontin; PGC-1α, peroxisome proliferator-activated receptor γ coactivator-1α; VDR, vitamin D receptor; DBD, DNA binding domain; LBD, ligand binding domain; AF, activation function; VDRE, vitamin D response element; PCR, polymerase chain reaction; GST, glutathione S-transferase; HEK, human embryonic kidney; DMSO, dimethyl sulfoxide; GFP, green fluorescent protein; PPAR, peroxisome proliferator-activated receptor; FXR, farnesoid X receptor; RXR, 9-cis retinoid X receptor; NR, nuclear receptor; TR, thyroid receptor; ER, estrogen receptor.
- Received March 9, 2005.
- Accepted May 20, 2005.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}