


Visual Overview
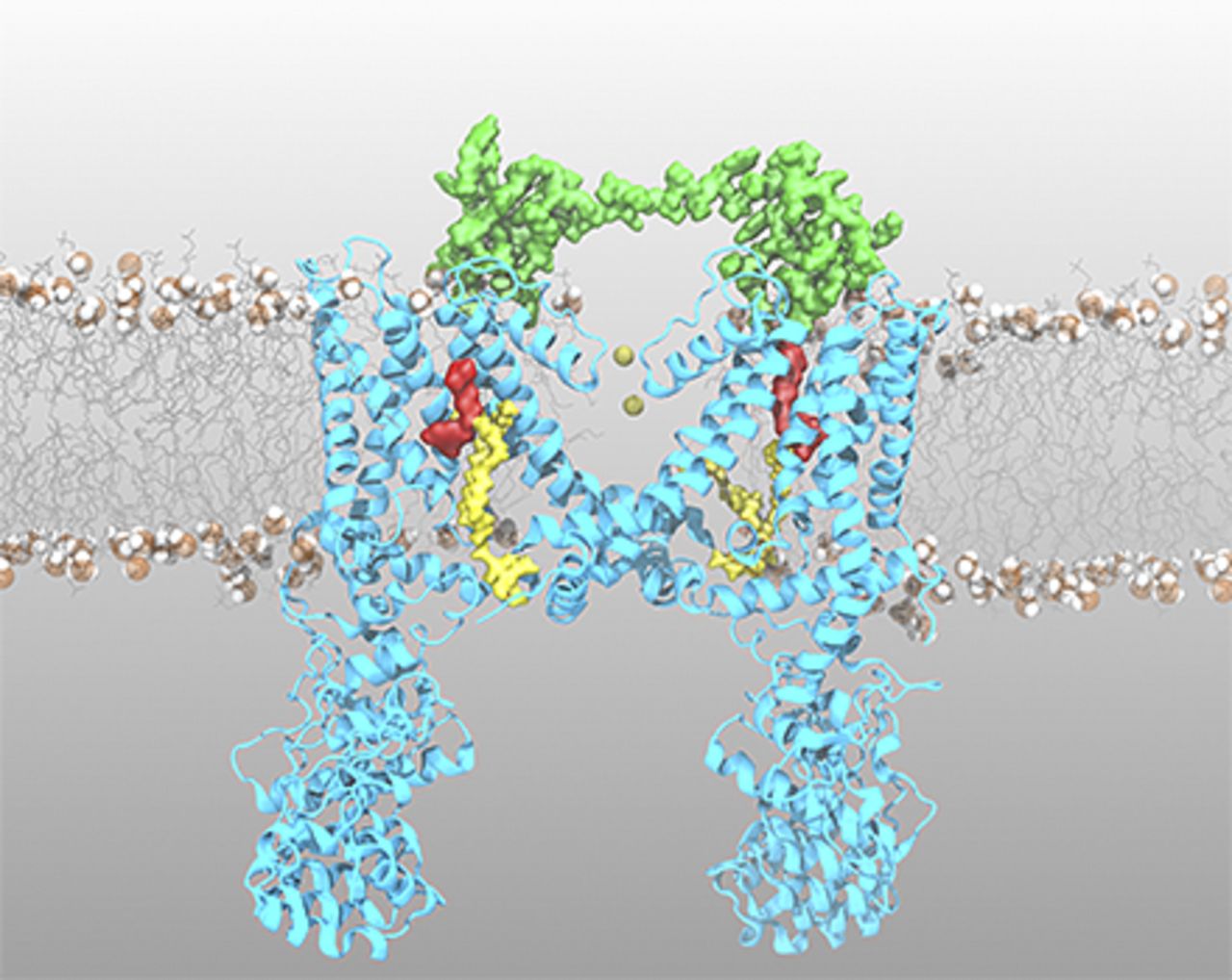
Abstract
The transient receptor potential vanilloid 1 (TRPV1) ion channel is a polymodal receptor that mediates the flux of cations across the membrane in response to several stimuli, including heat, voltage, and ligands. The best known agonist of TRPV1 channels is capsaicin, the pungent component of “hot” chili peppers. In addition, peptides found in the venom of poisonous animals, along with the lipids phosphatidylinositol 4,5-biphosphate, lysophosphatidic acid, and cholesterol, bind to TRPV1 with high affinity to modulate channel gating. Here, we discuss the functional evidence regarding ligand-dependent activation of TRPV1 channels in light of structural data recently obtained by cryoelectron microscopy. This review focuses on the mechanistic insights into ligand binding and allosteric gating of TRPV1 channels and the relevance of accurate polymodal receptor biophysical characterization for drug design in novel pain therapies.
Footnotes
- Received March 26, 2016.
- Accepted June 16, 2016.
This research was supported by the National Fund of Science and Technology [Grants 1150273, 1131003, and RI-130006]. The Centro Interdisciplinario de Neurociencia de Valparaíso is a Millennium Institute supported by the Millennium Scientific Initiative of the Ministerio de Economía, Fomento y Turismo.
- Copyright © 2016 by The American Society for Pharmacology and Experimental Therapeutics
MolPharm articles become freely available 12 months after publication, and remain freely available for 5 years.Non-open access articles that fall outside this five year window are available only to institutional subscribers and current ASPET members, or through the article purchase feature at the bottom of the page.
|





{kind=link}