


Abstract
SUMOylation, one of post-translational modifications, is covalently modified on lysine residues of a target protein through an enzymatic cascade reaction similar to protein ubiquitination. Along with identification of many SUMOylated proteins, protein SUMOylation has been proven to regulate multiple biologic activities including transcription, cell cycle, DNA repair, and innate immunity. The dysregulation of protein SUMOylation and deSUMOylation modification is linked with carcinogenesis and tumor progression. The SUMOylation-associated enzymes are usually elevated in various cancers, which function as cancer biomarkers to relate to poor outcomes for patients. Considering the significance of protein SUMOylation in regulating diverse biologic functions in cancer progression, numerous small-molecule inhibitors targeting protein SUMOylation pathway are developed as potentially clinical anticancer therapeutics. Here, we systematically summarize the latest progresses of associations of small ubiquitin-like modifier (SUMO) enzymes with cancers and small-molecular inhibitors against human cancers by targeting SUMOylation enzymes. We also compared the pros and cons of several special anticancer inhibitors targeting SUMO pathway. As more efforts are invested in this field, small-molecule inhibitors targeting the SUMOylation modification pathway are promising for development into novel anticancer drugs.
Introduction
The small ubiquitin-like modifier (SUMO) proteins, approximately with 11 kDa, covalently modify lysine (Lys) amino acid of a target substrate protein. Four major SUMO paralogues, including SUMO1, SUMO2, SUMO3, and SUMO4, are expressed in mammalian cells (Enserink, 2015). Most SUMOylation sites of a target protein reside in a consensus motif with the ψKxE or ψKxE/D sequence (ψ is an aliphatic residue, K is the target Lys, E/D is Asp or Glu, and x is any amino acid) (Yang et al., 2017). Except binding in consensus motif, SUMOs can noncovalently interact with proteins through targeting specific SUMO-interacting motifs (Hecker et al., 2006).
SUMOylation regulates multiple biologic processes including transcription, cell cycle, DNA repair, and innate immunity (Seeler and Dejean, 2003; Bettermann et al., 2012). A multistep enzymatic cascade reaction is involved in the biochemical process of protein SUMOylation (Fig. 1). First, four SUMO paralogues are synthesized as preproteins that are cleaved by sentrin-specific proteases (SENPs) to expose a diglycine (GG) motif on the carboxy-terminus. After that, an ATP-requiring activation, mediated by a heterodimeric SUMO-activating enzyme E1 (SUMO E1), generates a SUMO-SUMO E1 thioester. Then SUMO is transferred to the SUMO-conjugating enzyme E2 (SUMO E2) to form a thioester again. Next, a SUMO ligase E3 (SUMO E3) stabilizes the interaction between SUMO E2 and the substrate, and brings about an isopeptide bond between the SUMO C-terminus and a Lys within the target protein (Yang et al., 2017). Generally, SENPs, SUMO E1, SUMO E2, and SUMO E3 are four key enzymes to in turn perform cleaving, activating, transferring, and ligating of SUMO to Lys residue of a substrate protein.
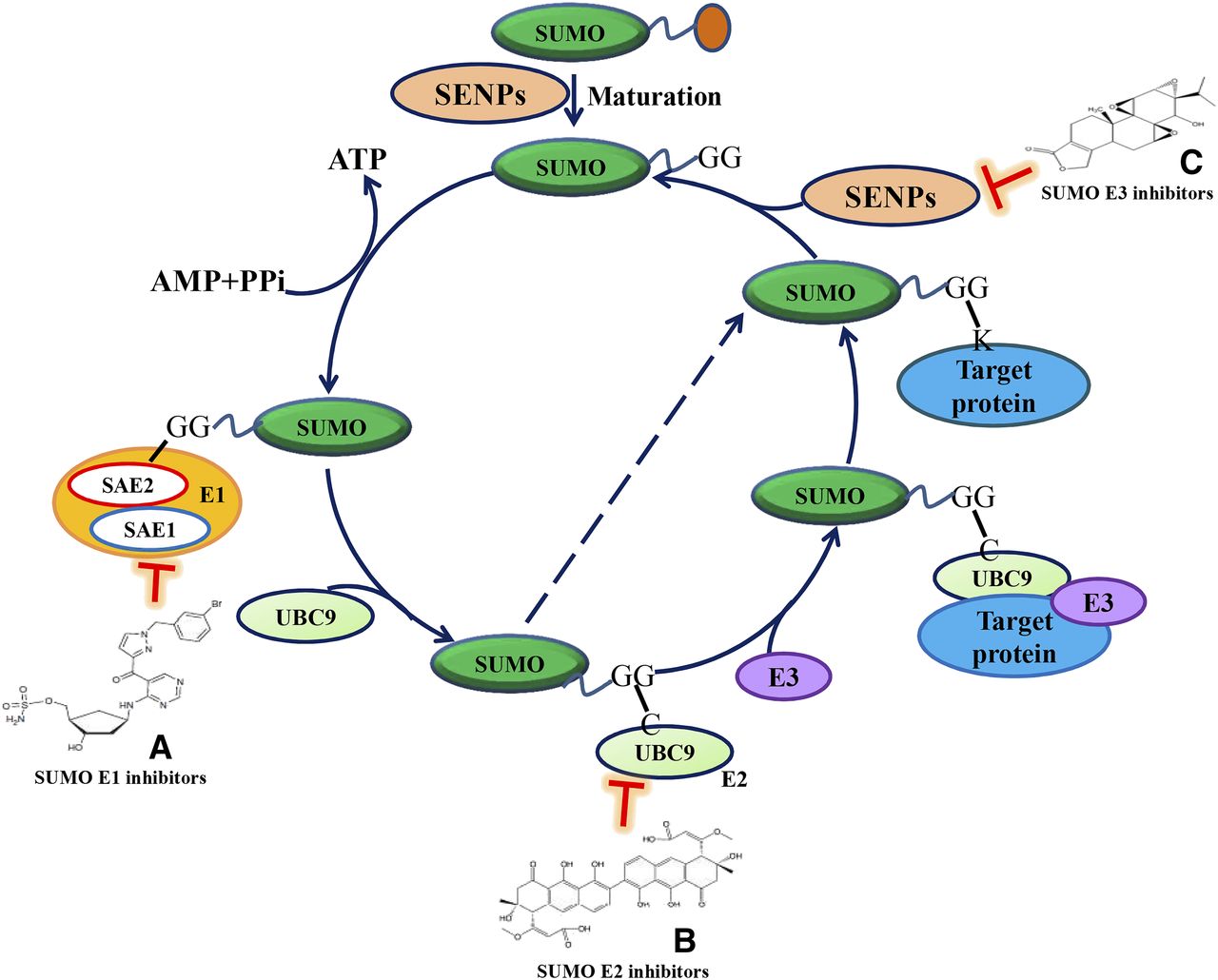
Small-molecule inhibitors that target the protein SUMOylation process. Protein SUMOylation is a multistep enzymatic cascade reaction. First, all SUMO paralogues are synthesized as preproteins that are cleaved by SENPs to expose a carboxy-terminal diglycine (GG) motif. An ATP-requiring activation step by the heterodimeric SUMO E1 generates a SUMO-SUMO E1 thioester. SUMO is then transferred to the SUMO E2, again forming a thioester. This last step usually requires a SUMO E3 to form an isopeptide bond between the SUMO C terminus and a Lys within the target protein. SUMO E2 promotes SUMOs onto Lys residues of target substrate even in the absence of an SUMO E3. The small-molecule inhibitors targeting SUMOylation-associated enzymes can block the subsequent SUMO modification biochemical process. (A–C) Chemical structure of three inhibitors (ML-792, spectomycin B1, and triptolide).
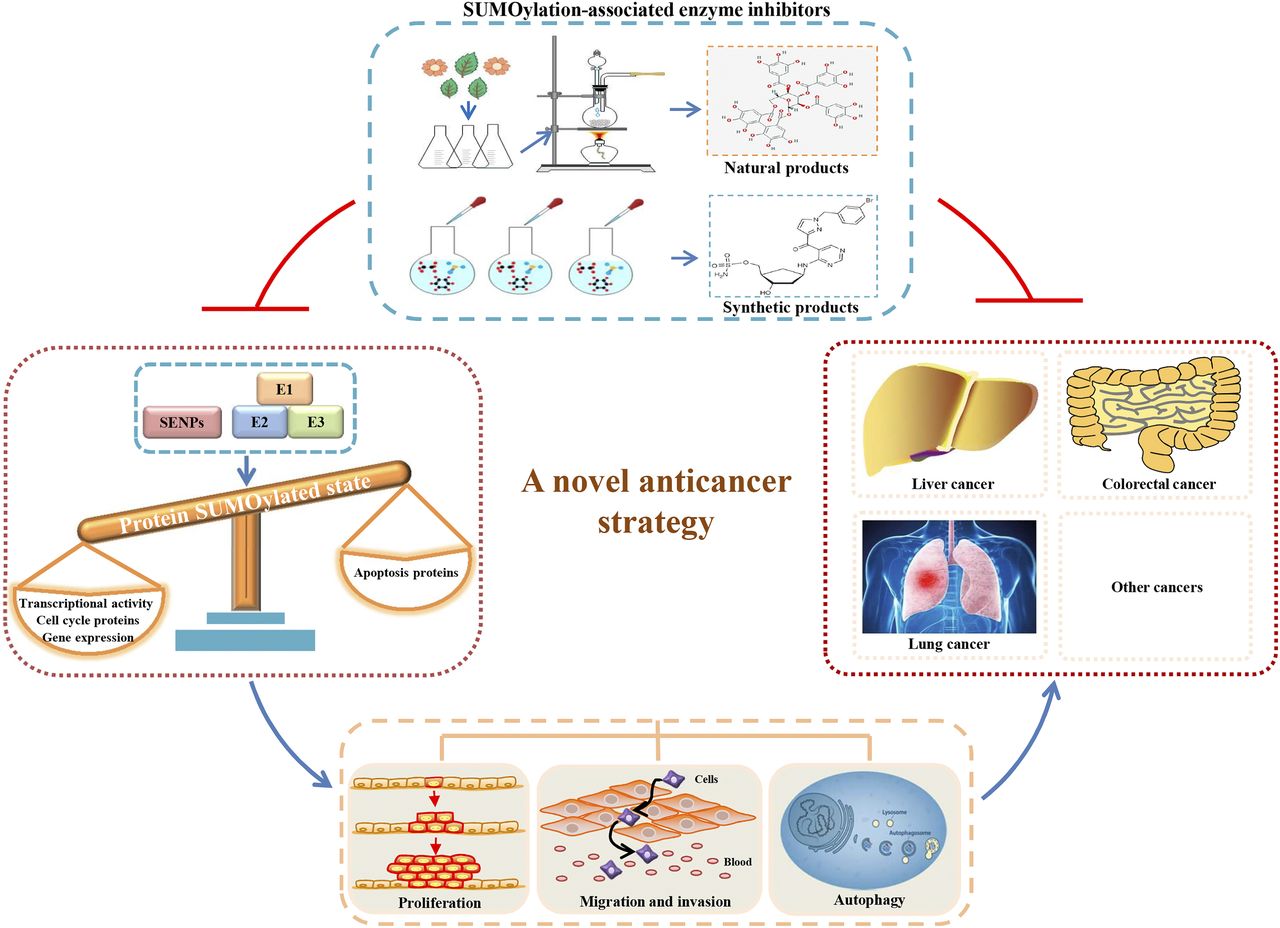
Protein SUMOylation is a dynamic regulation in physiologic conditions, and the imbalance of SUMOylation and de-SUMOylation associates with the occurrence and progression of cancer and other diseases (Fig. 2) (Liang et al., 2017; Seeler and Dejean, 2017; Yang et al., 2017). To date, due to the importance of SUMOylation in regulating diverse biologic systems in carcinogenesis, several novel small-molecule inhibitors targeting SUMOylation have been developed as promising anticancer compounds. In this review, we summarize the latest research progresses of small-molecule inhibitors by targeting SUMOylation-associated enzymes as highly potent anticancer compounds.

Novel anticancer drug discovery strategy based on SUMOylation-associated enzyme inhibitors. The high expression of SUMO E1, SUMO E2, SUMO E3, and SENPs in cancer cells breaks the functional balance of SUMOylated and deSUMOylated proteins, which changes gene transcription activity, gene expression, and cell biologic behaviors including cell proliferation, migration, invasion, and autophagy, eventually leading to the occurrence and progression of cancers. The natural or synthetic inhibitors targeting SUMOylation-associated enzymes have good anticancer activities through inhibiting protein SUMOylation.
Small-Molecule Inhibitors Targeting Protein SUMOylation-Associated Enzymes
The expression of SENPs, SUMO E1, SUMO E2, and SUMO E3 is usually increased in numerous cancers (Coppola et al., 2009; Bellail et al., 2014; Seeler and Dejean, 2017). The SUMO pathway components are involved in various tumors (Coppola et al., 2009; Bellail et al., 2014; Liu et al., 2015; Seeler and Dejean, 2017). The SUMO pathway components are involved in various human cancers (Table 1), so small-molecule inhibitors targeting protein SUMOylation-associated enzymes exhibit promising antitumor activities.
SUMOylation-associated enzymes in cancer cells
SENP and Its Anticancer Inhibitors.
SUMOylation is a reversible and dynamic process between protein SUMOylation and deSUMOylation for normal cellular activities (Yang et al., 2017). SENPs, as cysteine proteases, cleave the precursor or inactive form of SUMO at the C terminus via its hydrolase activity to expose two GG residues. Alternatively, SENPs can catalyze the deconjugation of SUMO molecules from a target protein. SENPs include six members, including SENP1, SENP2, SENP3, SENP5, SENP6, and SENP7. Among these SENPs, SENP1 is widely increased in various cancers (Table 1) (Zuo and Cheng, 2009; Xu et al., 2011). It plays an important role in cancer occurrence (Bawa-Khalfe et al., 2007), development (Bawa-Khalfe et al., 2010), and metastasis (Wang et al., 2013). For instance, SENP1 enhances vascular endothelial growth factor production by regulating the stability of hypoxia-inducible factor 1 (Xu et al., 2010), which is essential for cancer cell proliferation (Xu et al., 2011). In addition, the up-regulation of SENP1 is positively linked with lymph node metastasis and tumor aggressiveness of pancreatic ductal adenocarcinoma (Ma et al., 2014).
Except for SENP2, which decreases in bladder cancer (Tan et al., 2013) and hepatocellular carcinoma (HCC) (Shen et al., 2012), other SENP members such as SENP3 and SENP5 are also up-regulated in multiple cancers, including neuroblastoma (Xiang-Ming et al., 2016), multiple myeloma (Xu et al., 2015), gastric cancer (Ren et al., 2014), oral squamous cell carcinoma (Sun et al., 2013), and breast cancer (Cashman et al., 2014). Generally, the abnormal expression levels of SENPs can serve as tumor biomarkers.
To date, several SENP1 inhibitors have exhibited good anticancer activities in vitro (Table 2). These inhibitors mainly include benzodiazepine-based peptidomimetic covalent compounds (Qiao et al., 2011), SUMO-derived peptide-based covalent inhibitors (Albrow et al., 2011), and the noncovalent 2-(4-chlorophenyl)-2-oxoethyl 4-benzamidobenzoates (Chen et al., 2012b) and 1-[4-(N-benzylamino) phenyl]-3-phenylureas (Uno et al., 2012).
Small-molecule inhibitors for SENP1/2
Inhibitors’ chemical structure from corresponding literature or PubChem.
To improve anticancer potency, several compounds have been designed to obtain micromolar levels of antitumor activities. For example, compound 13m has 3.5 μM of IC50 through in silico screening and rational drug design (Zhao et al., 2016), and the compound 3 is obtained with 3.3 μM of IC50 (Xie et al., 2016). However, these inhibitors need great improvement in order for them to be useful for in-depth in vivo studies.
The SENP1 inhibitors not only are designed through chemical synthesis but also are derived from natural products. Triptolide, extracted from medicinal herbs, down-regulates SENP1 and inhibits prostate cancer cell proliferation (Huang et al., 2012). Similarly, a natural pentacyclic triterpenoid named Momordin Ιc also induces cell cycle arrest and cell apoptosis to suppress prostate cancer cell growth by inhibiting of SENP1 (Wu et al., 2016). So SENP1 may serve as an attractive drug target for developing new cancer therapeutics.
In addition to SENP1, SENP2 has been confirmed to be a molecular target for chemical drugs that effectively enhance SUMO conjugation and suppress SENP2 activity. For example, compound 1, 2, 5-oxadiazole, has potential for development into a novel SENP2-targeting therapeutic agent against various diseases (Kumar et al., 2014). In addition, cytoprotective molecules are being developed as novel stroke therapies by quantitative high-throughput screening (Bernstock et al., 2018b).
SUMO-E1 and Its Anticancer Inhibitors.
SUMO E1, an activating enzyme in SUMOylation pathway, is a heterodimer consisting of SUMO-activating enzyme subunit 1 (SAE1) and subunit 2 (SAE2), which has a particular significance due to its enhancing cell metastasis in tumorigenesis (Table 1). The increased SUMO enzymes are closely associated with pathogenesis of HCC; higher levels of SUMO E1 indicate lower survival rates of HCC patients (Lee and Thorgeirsson, 2004). Knock-down of SAE2 inhibits cancer malignancy and increases chemotherapy sensitivity of small cell lung cancer (Liu et al., 2015). Breast cancer patients with lower expression of SAE1/SAE2 have significantly fewer cases of metastatic cancer and increased survival compared with patients with higher SAE1/SAE2 levels (Kessler et al., 2012). SAE1/SAE2 is also up-regulated in lung adenocarcinoma (Inamura et al., 2007).
There has been growing interest in investigating selective SUMO E1–targeting inhibitors (Table 3). For example, several natural products—including ginkgolic acid (Fukuda et al., 2009a), kerriamycin B (Fukuda et al., 2009b), and davidiin (Takemoto et al., 2014)—are confirmed to inhibit SUMO E1 by blocking formation of SUMO E1–SUMO intermediate. Among these natural products, ginkgolic acid is the most widely used and commercially available chemical regent that targets global SUMOylation, but its inhibitory effects on cells can vary greatly depending on the assay and a measured substrate. Tannic acid has been identified as targeting human liver receptor homolog-1 as a general nontoxic SUMOylation inhibitor via cell-based screening (Suzawa et al., 2015).
Small-molecule inhibitors for SUMO-activating enzyme E1
Inhibitors’ chemical structure from corresponding literature or PubChem.
Most natural SUMO E1 inhibitors function in the micromolar range except for davidiin, which inhibits at submicromolar concentrations. To address this limitation, other synthetic SAE inhibitors—compound 21 (Kumar et al., 2013) and ML-792 ([(1R,2S,4R)-4-[[5-[1-[(3-bromophenyl)methyl]pyrazole-3-carbonyl]pyrimidin-4-yl]amino]-2-hydroxycyclopentyl]methyl sulfamate) (He et al., 2017)—have been developed to inhibit cancer cell growth with good selective activity. ML-792 forms a covalent adduct with SUMO at its C terminus, which inhibits the enzyme activity of SAE2 at nanomolar concentrations (He et al., 2017). Therefore, ML-792 selectively blocks SAE activity in treating MYC-amplified tumors.
SUMO-E2 and Its Anticancer Inhibitors.
Ubiquitin-conjugating enzyme 9 (Ubc9) is a sole SUMO E2 in the SUMOylation pathway, which promotes SUMO transferring into Lys residues of a target substrate even in the absence of SUMO E3 (Gareau and Lima, 2010). Ubc9 is increased in many cancers (Seeler and Dejean, 2017), including advanced melanomas, head and neck tumor, lung tumor, HCC, colon cancer, breast cancer, and glioblastoma (Ahn et al., 2001; Moschos et al., 2007, 2010; Wu et al., 2009; Chen et al., 2011; Tomasi et al., 2012; Li et al., 2013; Yang et al., 2013). Inhibition of Ubc9 expression sensitizes cytotoxic effects of chemotherapeutic drugs against melanoma cells (Ahn et al., 2001; Moschos et al., 2007). Ubc9 stability and its expression are increased by cell division cycle 2 (Cdc2)-mediated phosphorylation in HCC (Tomasi et al., 2012). Therefore, detection of Ubc9 expression level could be helpful to early diagnosis of adenocarcinoma, HCC, and other cancers.
Suppression of Ubc9 by small-molecule inhibitors represents a potential or emerging strategy. We have summarized several Ubc9 inhibitors that suppress human malignant cells in vitro (Table 4). For instance, GSK145A was identified as an Ubc9 inhibitor using a high-throughput assay (Brandt et al., 2013). 2-D08 (2′,3′,4′-trihydroxyflavone) has been identified as a potent SUMOylation inhibitor by an electrophoretic mobility shift assay (Kim et al., 2013). In addition, spectomycin B1, an antibiotic against gram-positive bacteria (Staley and Rinehart, 1994) that has been identified as a novel SUMOylation inhibitor with direct binding to Ubc9, reduces estrogen receptor-dependent gene expression and suppresses proliferation of breast-cancer cells (Hirohama et al., 2013). Spectomycin B1 is a potential therapeutic agent against hormone-dependent breast cancers (Hirohama et al., 2013). The development of other Ubc9 chemical inhibitors is ongoing through a small-molecule microarray approach (Zlotkowski et al., 2017).
Small-molecule inhibitors for SUMO-conjugating enzyme E2
SUMO-E3 and Its Anticancer Inhibitors.
SUMO E3 includes the protein inhibitor of activated STAT (PIAS), Ran binding protein 2 (RanBP2), the polycomb protein (Pc2), and other members (Martin et al., 2007). SUMO conjugation is always enhanced in the presence of SUMO E3, but in the absence of SUMO E3 Ubc9 can also promote SUMOs onto Lys residues of a target substrate. Most of current understanding on SUMO E3 functions mainly comes from studies of the PIAS family (Coppola et al., 2009; Chen et al., 2012a), which regulates protein stability and signaling transduction pathways, and controls inflammation (Rabellino et al., 2017). RanBP2, a nuclear pore complex protein that belongs to the second type of SUMO E3, has also been found to increase in multiple myeloma (Felix et al., 2009) and small cell lung cancer (Horio et al., 2010). Therefore, SUMO E3 ligase could be a potential biomarker for cancer and act as a drug target for cancer therapy. However, no small-molecule inhibitor has been designed to effectively inhibit SUMO-E3 enzymes up to now.
Pros and Cons of SUMOylation-Targeting Small-Molecule Inhibitors
SUMOylation-associated enzyme inhibitors have been derived from natural products or synthetic chemicals. Besides previous biologic activities, a few natural products have been found to have novel anticancer potentials via acting on the SUMO process. For example, kerriamycin B and spectomycin B1 are typical antibiotics against gram-positive bacteria, and ginkgolic acid and tannic acid have good anti-inflammatory or immunosuppressive activities (Staley and Rinehart, 1994; Antonoff et al., 2009). These reagents have been confirmed to have anticancer activities by inhibition of protein SUMOylation. Although these compounds have yet to be approved for any clinical trials against tumors, they hold great promise for development into efficient therapeutics, thus extending new clinical applications to old drugs. As they are already used in clinical applications, these drugs would present few novel side effects if their disease treatment spectrum were enlarged.
The SUMOylation pathway is an ideal drug target to overcome oncogenic mechanisms that promote a proapoptotic state, but most SUMOylation inhibitors display weak anticancer activity and uncharacterized selectivity in their current stage (Table 5). Therefore, many more drug-like chemical scaffolds are needed to uncover the therapeutic potential of SUMOylation–associated enzyme inhibitors, along with their structural description.
The pros and cons of SUMO inhibitors
Future Perspectives
Protein post-translational modification (PTM) is a step in protein biosynthesis, including phosphorylation, glycosylation, ubiquitination, SUMOylation, and other modifications (Kessler and Edelmann, 2011). Among these PTMs, SUMOylation, a competitor of ubiquitination, has become a research hot spot in recent years (Yang et al., 2017). Mass spectrometry is the most sensitive method for identification and quantification of PTMs (Liang et al., 2012), including protein SUMOylation (McManus et al., 2016; Lamoliatte et al., 2017; Lumpkin et al., 2017). More and more SUMOylated proteins have been shown to be involved in the pathogenesis of multiple diseases, including brain ischemia and cancers (Liang et al., 2017; Yang et al., 2017; Bernstock et al., 2018a). Protein SUMOylation mediates in protein-protein interactions, is responsive to DNA damage, changes protein intracellular localization, or directs changes of protein activities (Dou et al., 2011; Hickey et al., 2012). Thus, the SUMOylation pathway has emerged as a promising therapeutic target for drug discovery because of its crucial role in various cancers.
Compared with conventional high-throughput screening, virtual filtering plays a noteworthy role in the identification and optimization of small-molecule inhibitors targeting the SUMO pathway. Through virtual screening in combination with biologic assay (Fukuda et al., 2009a,b; Kumar et al., 2013; Takemoto et al., 2014; Suzawa et al., 2015; He et al., 2017; Bernstock et al., 2018b). GSK145A, 2-D08, and spectomycin B1 inhibit Ubc9 (Brandt et al., 2013; Hirohama et al., 2013; Kim et al., 2013), and several compounds exhibit potential activity against tumors. However, the discovery phase of SUMOylation-associated enzyme inhibitors as a potential therapeutic remains in its infancy. Many of the available inhibitors of SUMOylation-associated enzyme display weak activity and uncharacterized selectivity.
It is worth mentioning that a novel inhibitor, ML-792, specifically inhibits SAE at nanomolar concentrations without suppressing any other ubiquitin-activating enzymes or various other ATP-using enzymes (He et al., 2017). Therefore, the SUMO inhibitors with high selectivity facilitate further exploration of the novel cellular functions of protein SUMOylation, which opens a new avenue for specifically targeting SUMOylation as a potential clinical anticancer therapy.
Authorship Contributions
Wrote or contributed to the writing of the manuscript: Yang, Xia, Wang, Zhao, Sheng, Ye, He, Zhou, Zhu, Xu, Liang.
Footnotes
- Received February 26, 2018.
- Accepted May 16, 2018.
This work was supported by the National 863 High Tech Foundation [Grant 2014AA020608], the National Key Basic Research Program of China [Grants 2013CB911303 and 2011CB910703], the National Natural Science Foundation of China [Grants 31470810 and 31071235], the Science and Technology Department of Sichuan Province [Grant 2017JY0232], and the Health and Family Planning Commission of Sichuan Province [Grant 17ZD045].
Abbreviations
- HCC
- hepatocellular carcinoma
- ML-792
- [(1R,2S,4R)-4-[[5-[1-[(3-bromophenyl)methyl]pyrazole-3-carbonyl]pyrimidin-4-yl]amino]-2-hydroxycyclopentyl]methyl sulfamate
- PTM
- protein post-translational modification
- SAE1/2
- SUMO-activating enzyme subunit 1/2
- SENP(s)
- sentrin-specific proteases
- SUMO
- small ubiquitin-like modifier
- SUMO E1
- SUMO-activating enzyme E1
- SUMO E2
- SUMO-conjugating enzyme E2
- SUMO E3
- SUMO ligase E3
- Ubc9
- ubiquitin-conjugating enzyme 9
- Copyright © 2018 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}