


Abstract
Receptor activity modifying proteins (RAMPs) constitute a group of three proteins, designated as RAMP1, 2, and 3, which are able to effect functional changes in some members of the G protein-coupled receptor family. Thus, RAMP1 or RAMP3 can modify the calcitonin receptor (CTR) to also function as a high-affinity amylin receptor-like phenotype. To examine the RAMP/CTR interaction, individual RAMPs were coexpressed with either of the two human CTR (hCTR) isoforms, the insert negative (hCTRI1−) or the insert positive (hCTRI1+), in Chinese hamster ovary (CHO-P) or African monkey kidney (COS-7) cells. CHO-P cells provide an environment conducive to a low, but significant, level of amylin binding with either hCTR isoform alone, unlike in COS-7, where RAMP coexpression is imperative for amylin binding. Also, in CHO-P, hCTRI1− induced amylin binding with all three RAMPs, in contrast to COS-7, where only RAMP1 or RAMP3 generate an amylin receptor phenotype. hCTRI1+ induced high-affinity amylin binding with any RAMP in either cell line. In COS-7 cells, hCTRI1+/RAMP-generated receptor displayed high- and low-affinity states, in contrast with the single-state binding seen with hCTRI1−/RAMP-generated receptor, whereas in CHO-P cells a two-affinity state receptor phenotype was evident with both hCTR isoforms. Endogenous RAMP expression is low and similar between cell lines. The results suggest that CTR/RAMP interaction in these cells is complex with other cellular factors such as the levels of different G proteins and/or receptor/RAMP stoichiometry following heterologous coexpression contributing to the ultimate receptor phenotype.
Amylin, a peptide hormone secreted by the pancreatic β-cells, has effects on a number of tissues. In skeletal muscle, amylin inhibits insulin-stimulated glucose uptake and glycogen synthesis (Leighton and Cooper, 1988; Kreutter et al., 1993). In the gastrointestinal system, it regulates gastric acid secretion and gastric emptying (Rossowski et al., 1997). In the kidney, it is known to regulate ion transport and activity of renin (Cooper and Wookey, 1997). Amylin also has anorectic effects following direct infusion into some regions of the brain and following peripheral administration (reviewed by Sexton and Perry, 1996). Consistent with its functional targets, high-affinity binding sites for amylin have been demonstrated in brain, kidney, and skeletal muscle (Beaumont et al., 1993; Sexton et al., 1994; Wookey et al., 1996; Chai et al., 1997).
Recently, we demonstrated that high-affinity amylin receptor-like phenotypes are generated from the calcitonin receptor (CTR) gene product when coexpressed with receptor activity modifying proteins (RAMPs) (Christopoulos et al., 1999). Originally discovered during attempts to clone the receptor for calcitonin gene-related peptide (CGRP), RAMPs comprise a family of three proteins, designated RAMP1, 2, and 3, each of ∼16-kDa molecular mass. RAMPs are single transmembrane domain proteins with a short intracellular domain and a relatively long extracellular domain (McLatchie et al., 1998). The demonstration of CTR/RAMP derivation of amylin receptor phenotypes followed the discovery by McLatchie et al. (1998) that high-affinity CGRP or adrenomedullin receptors are generated by the coexpression of either RAMP1 or RAMPs 2 or 3, respectively, with calcitonin receptor-like receptor (CRLR).
In many species, CTRs exist in multiple forms, resulting from alternate splicing of the CTR gene transcript. The two most common isoforms of the human CTR (hCTR) are identical in amino acid sequence except for the presence or absence of a 16-amino-acid insert in the putative first intracellular domain. These isoforms are designated as hCTRI1+ (or hCTR-1) and hCTRI1− (or hCTR-2), respectively. We found that the coexpression of hCTRI1− with RAMP1 or RAMP3, but not RAMP2, in COS-7 cells resulted in a significant induction of specific 125I-rat amylin binding, and amylin-induced cAMP response. The RAMP1-derived amylin receptor phenotype also had high affinity for CGRP, whereas the RAMP3-derived phenotype had reduced affinity for CGRP (Christopoulos et al., 1999). Confocal microscopic localization of c-myc-tagged RAMP1 indicated that, when transfected alone, RAMP1 was essentially localized in the cytosol. Cotransfection with hCTR induces cell surface expression of RAMP1, presumably associated with CTR (Christopoulos et al., 1999). Thus, it appears that the functional expression of receptors for peptides of the calcitonin (CT)/amylin/CGRP/adrenomedullin family depends on the interaction of CTR or CRLR with RAMPs.
The insert-negative and -positive isoforms of the hCTR are differentially expressed in a variety of tissues, although the hCTRI1− is the predominant isoform (Kuestner et al., 1994). Little difference in CT binding is observed between these two isoforms, but they differ markedly in signaling properties (Nussenzveig et al., 1994; Moore et al., 1995; Naro et al., 1998). It remains unclear whether RAMP-receptor interactions vary among different receptor isoforms, and the extent to which other components of the cellular background may influence receptor phenotype. In this study, we have compared the pharmacological profile of amylin and CT receptor phenotypes derived from the two major hCTR isoforms and individual RAMPs coexpressed in either COS-7 or Chinese hamster ovary (CHO-P) cells. We found that the resulting amylin receptor-like phenotypes vary in their pharmacological characteristics dependent on the hCTR isoform involved and the host cell environment in which they are expressed.
Materials and Methods
Hormones and Chemicals.
Salmon CT (sCT), human CT (hCT), human CGRP (hCGRP), and rat amylin were obtained from Bachem (Torrance, CA). Dulbecco's modified Eagle's medium (DMEM), fetal bovine serum (FBS), HEPES, gentamicin, and LipofectAMINE were obtained from Life Technologies-BRL (Grand Island, NY). Minocycline was from ICN Biomedicals (Costa Mesa, CA). BSA was obtained from Commonwealth Serum Laboratories (Parkville, Australia).125I-rat amylin (specific activity 2000 Ci/mmol) and 125I-hCT (specific activity 2000 Ci/mmol) were purchased from Amersham (Buckinghamshire, UK). All other chemicals were reagent grade or better.
Plasmid DNA.
The cDNAs of hCTRI1− and hCTRI1+ isoforms were cloned into the expression plasmid vectors pZem228CC (Kuestner et al., 1994) and pcDNA1 (Invitrogen, San Diego, CA), respectively. RAMP1, 2, and 3 cDNAs were cloned into the expression vector plasmid pcDNA3neo (Invitrogen).
Cell Culture.
African green monkey kidney (COS-7) cells and CHO-P cells, a variant of CHO cells stably transfected with P-selectin (a gift from Dr. A. Albestin, Howard Florey Institute of Experimental Medicine, Parkville, Victoria, Australia), were routinely maintained at 37°C in a humidified atmosphere of 95% air, 5% CO2, in DMEM containing 5% (v/v) FBS, 15 mM HEPES, 80 mg/l gentamicin, and 1 mg/l minocycline.
For binding studies, cells were seeded into 24-well plates at approximately 2.5 × 104cells/cm2 surface area. Cells were transfected 48 h postseeding, when they were approximately 70 to 80% confluent.
Transfection.
For RAMP dose-response studies, cells grown in 24-well plates were transfected with plasmid carrying the hCTR cDNA (0.1 μg/well) without RAMP or vector DNA, or with increasing amounts (0.05–0.25 μg/well) of RAMP or vector DNA (pcDNA3).
For competition binding studies, cells grown as above were cotransfected with plasmid carrying the hCTR cDNA (0.1 μg/well) with RAMP cDNA, or vector DNA (0.15 μg/well). Transfections were performed using LipofectAMINE (Life Technologies-BRL) according to the manufacturer's instructions. DNA for transfection was purified using Qiagen maxi prep columns (Qiagen, Clifton Hill, Australia). DNA resuspended in serum-free DMEM was mixed with LipofectAMINE (5 μl/μg of DNA) and left at room temperature for 30 min to allow the DNA/liposome complex formation. Growth medium was removed and the DNA/liposome complex was added to cells, followed by the addition of a sufficient volume of serum-free DMEM to make a final volume of 250 μl of medium/well. Transfected cells were incubated at 37°C in 95% air, 5% CO2 incubator. After a 4- to 5-h incubation, 250 μl of DMEM containing 10% FBS was added to each well, and incubation continued for an additional 48 h before performing binding assays.
Radioligand Binding.
Receptor/RAMP or receptor/vector cotransfected cells in 24-well plates were incubated in binding buffer (DMEM containing 0.1% w/v BSA) with ∼80 pM125I-hCT or ∼70 pM125I-rat amylin, in the absence (total binding) or presence of increasing concentrations (10−13to 10−6 M) of unlabeled peptide. Nonspecific binding was defined as binding in the presence of 10−6 M unlabeled peptide. Following incubation for 60 min at 37°C, cells were rinsed with ice-cold PBS (140 mM NaCl, 2 mM KCl, 1 mM KH2PO4, 8 mM Na2HPO4, pH 7.3), solubilized with 0.5 M NaOH, and counted on a Packard gamma counter at ∼75% counting efficiency.
RNA Extraction and Northern Analysis.
Total RNA was extracted from COS-7 and CHO-P cells by the guanidine isothiocyanate/phenol/chloroform extraction method (Chomczynski and Sacchi, 1987). The RNA (20 μg/lane) was size-fractionated on a formaldehyde/agarose (1% w/v) gel, transferred onto a nylon-based membrane (zeta probe; Bio-Rad, Hercules, CA), and cross-linked to the membrane by UV light exposure.
Prehybridization was carried out at 55°C for 15 min in 8 ml (1 ml/5 cm2 membrane area) of prehybridization buffer (0.5 M Na2HPO4, pH 7.2; 7% SDS; 1 mM EDTA). Hybridization was carried out at 55°C overnight in the same buffer to which was added 32P-labeled RAMP1, 2, or 3 probe. The probe in each instance was a cDNA fragment obtained after restriction digestion of human RAMP1, 2, or 3 cDNA or mouse RAMP1, 2, or 3 cDNA. The restriction-digested DNA was electrophoresed on 1% (w/v) agarose, the gel portion containing the appropriate fragment excised. DNA was extracted from the gel using Bresaclean (Bresatec, Thebarton, SA, Australia), and labeled with [32P]α-dCTP by random primer labeling using a Prime-a-gene kit (Promega, Madison, WI). After hybridization, membranes were washed at 42°C for 30 min in wash solution A (40 mM Na2HPO4, pH 7.2; 5% SDS; 1 mM EDTA) followed by a 30-min wash at 42°C in solution 2 (40 mM Na2HPO4; 1% SDS; 1 mM EDTA). The membranes were exposed to X-ray film (Kodak Bio-max) for periods ranging from 3 to 5 days at −80°C with intensifying screens.
Reverse Transcription-Polymerase Chain Reaction (RT-PCR).
First strand cDNA was synthesized by incubating 5 μg of total RNA at 42°C for 2 h in the presence of Moloney-murine leukemia virus reverse transcriptase, random hexamer oligonucleotides, and reverse transcriptase buffer (Promega). The RT products were used in PCR amplification using appropriate primer sets bearing nucleotide sequences that were based on human RAMP1, 2, or 3 cDNAs. Each PCR comprised a single denaturation step of 5 min at 95°C followed by varying numbers of cycles of 1 min at 95°C, 1 min at 55°C, and 2 min at 72°C. From the initial reaction volume of 50 μl, aliquots of 10 μl each were removed for analysis between 25 and 40 cycles. The PCR products were electrophoresed on a 1% (w/v) agarose gel alongside a size marker, stained with ethidium bromide, and photographed under UV transillumination.
Data Analysis.
To account for differences in transfection efficiency and binding levels between experiments, data were expressed as percentages of specific binding, where nonspecific binding was defined as binding in the presence of 10−6 M sCT. Competition binding curves were analyzed using GraphPad Prism (version 3; GraphPad Software, San Diego, CA) using either a one- or two-site mass action model. The curve fits were constrained with a lower limit of 0.1 to 1 as binding data was entered as a percentage of specific binding. A P value of less than .05 was considered significant.
The effect of RAMP cotransfection on 125I-amylin or 125I-hCT binding was analyzed by two-tailed paired t test (RAMP versus vector control) using the program SigmaStat (version 2; SPSS Inc., Chicago, IL).
Results
RAMPs 1, 2, and 3 Induce Specific 125I-Amylin Binding in COS-7 Cells Cotransfected with hCTRI1+
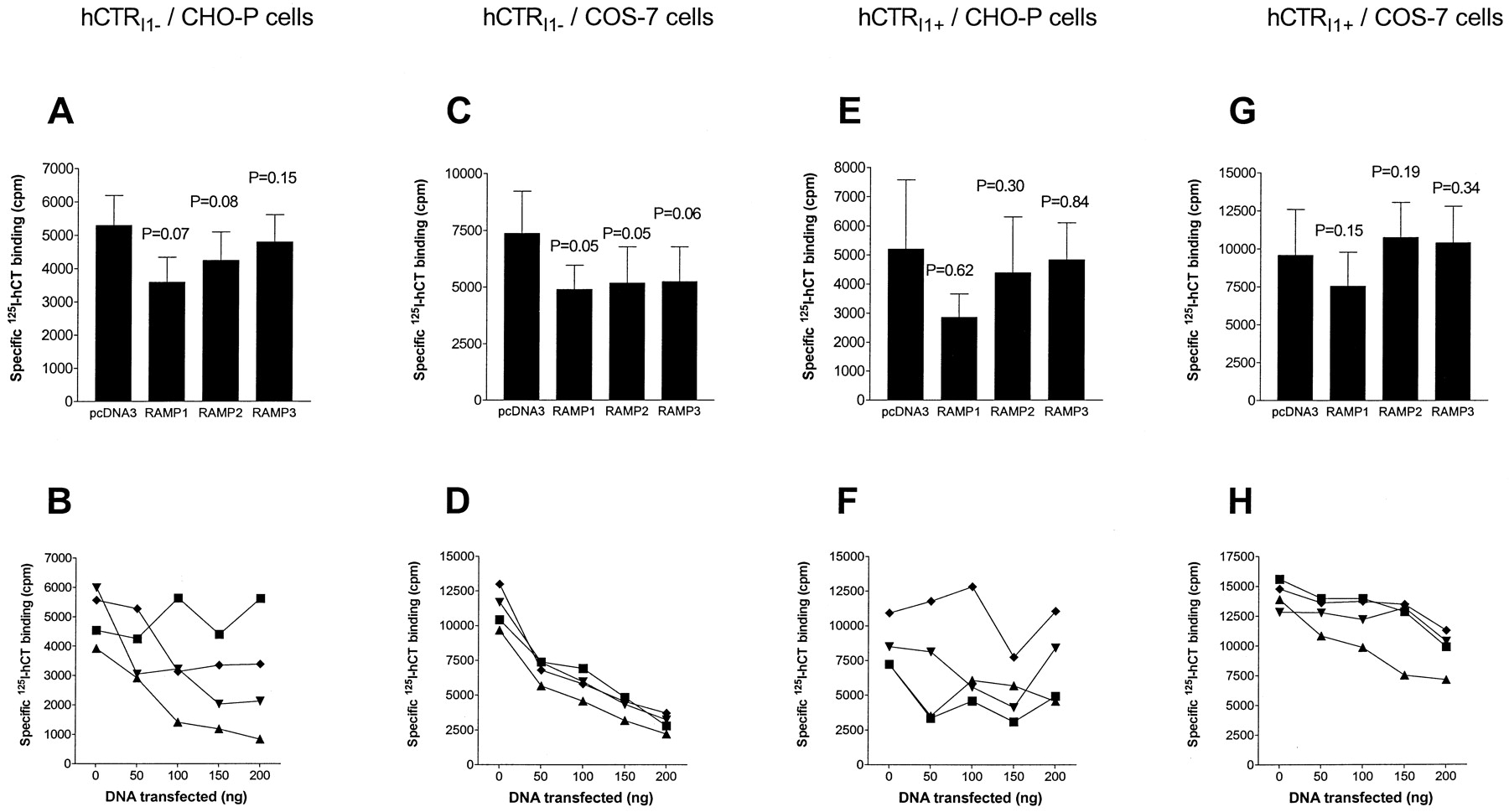
Cotransfection of hCTRI1+ and each of RAMP-1, -2, or -3 in COS-7 cells led to a marked induction of specific125I-amylin binding (Fig.1, A and B). This differed from results with the insert-negative isoform where only RAMPs 1 and 3 generated a significant change in amylin binding (Christopoulos et al., 1999).

Effect of RAMP cotransfection on the expression of specific 125I-rat amylin binding in COS-7 (A and B) or CHO-P (C–F) cells transfected with either hCTRI1+ (A–D) or hCTRI1− (E and F). Cells were transfected with 100 ng of receptor DNA and increasing amounts of RAMP DNA or vector control (B, D, F; representative data from experiments repeated three to four times). Pooled data for cells transfected with 100 ng of receptor DNA and 150 ng of RAMP or vector DNA are shown in A, C, and E (n = 5–8). *P < .05, pairedt test for RAMP versus vector control. Vector (pcDNA3) control, ▪; RAMP1, ▾; RAMP2, ▴; RAMP3, ♦.
Only low level specific amylin binding was seen with receptor plus vector (see the legend to Fig. 2A), and this was predominantly to a CT receptor-like phenotype (Table1), which was similar to that characterized with 125I-hCT (see Table 4). Comparison of RAMP-induced phenotype indicated that interaction with sCT and rat amylin was similar for all three RAMPs (Fig. 2, B–D). Unlike interaction with the hCTRI1− isoform (Christopoulos et al., 1999), binding could be segregated into a high- and low-affinity component, with the majority of sites corresponding to the low-affinity site (Table 1; Fig. 2). Human CT interacted with essentially a single class of sites, with the RAMP2-induced phenotype displaying higher affinity than the RAMP1- or RAMP3-induced phenotypes (Fig. 2, C versus B and D; Table 1). All three RAMPs induced a high- and low-affinity CGRP interaction (Fig. 2, B–D). The number of high-affinity CGRP sites, however, was lower in the RAMP2-induced phenotype, whereas the affinity of the RAMP3-induced phenotype was lower (for both sites), than the RAMP1 or RAMP2 phenotypes (Table 1; Fig. 2D). The affinity of peptides for RAMP1- and RAMP3-induced phenotype from hCTRI1− (Christopoulos et al., 1999) was similar to the low-affinity sites revealed with hCTRI1+.

Competition of 125I-rat amylin binding to COS-7 cells cotransfected with 100 ng of hCTRI1+ DNA and 150 ng of pcDNA3 (A), RAMP1 (B), RAMP2 (C), or RAMP3 (D) DNA. sCT, ▪; hCT, ▴; rat amylin, ▾; hCGRPα, ♦. Data are mean ± S.E. of three separate experiments. B, total binding; Bo, total binding in the absence of competing peptide; N, nonspecific binding (measured in the presence of 10−6 M sCT). Actual Bo values (cpm) for individual experiments were pcDNA3 (1,600; 1,300; 2,700); RAMP1 (14,600; 9,600; 15,300); RAMP2 (14,400; 10,600; 16,300); RAMP3 (10,700; 4,700; 10,700). Actual N values (cpm) ranged from 400 to 1200.
pIC50 values (M) for peptides in competition for binding of125I-rat amylin to COS-7 cells transfected with hCTRl1+(100 ng) and RAMP or vector DNA (150 ng)
pIC50 values (M) for peptides in competition for binding of125I-hCT to COS-7 cells transfected with hCTRl1+ (100 ng) and RAMP or vector DNA (150 ng)
RAMPs 1, 2, and 3 Induce Specific 125I-Amylin Binding in CHO-P Cells Cotransfected with Either hCTRI1+ or hCTRI1−
In contrast to COS-7 cells, cotransfection of either hCTRI1+ or hCTRI1− with any of the RAMPs in CHO-P cells led to an increase in specific125I-amylin binding (Fig. 1, C–F).
Analysis of hCTRI1+/RAMP Phenotypes in CHO-P Cells
Competition binding analysis of the hCTRI1+-based phenotypes in CHO-P cells indicated that binding was segregated into high- and low-affinity sites (Fig.3; Table2). Multi-site binding was apparent for all four peptides used to characterize the receptors and, as such, CHO-P binding displayed more complexity than the binding to COS-7 cells (compare Figs. 2 and 3). The interaction of hCT was similar across all hCTRI1+/RAMP combinations, with respect to both ligand binding affinities and the proportion of receptors in high- versus low-affinity states (Table 2). sCT displayed similar affinities in all receptor/RAMP combinations, although the number of high-affinity sites was greater than with the vector control (Table 2). Analysis of amylin competition binding indicated that RAMP1 and RAMP3 increased the affinity of both the low- and high-affinity sites relative to the vector control. RAMP2 also increased the affinity of the low-affinity site, but to a lesser extent than RAMP1 (P < .05). No change to the high-affinity site was seen (Table 2). For CGRP interaction with different hCTRI1+/RAMP combinations, no significant change was seen in the high-affinity site, except for an increase in the proportion of sites in the high-affinity state with RAMP3. RAMP1 increased the affinity of the low-affinity site, as did RAMP2, but to a lesser extent than RAMP1 (P < .05). The hCTRI1+/RAMP3 low-affinity site was equivalent to that seen with receptor plus vector and was significantly lower in affinity than the RAMP1-induced site (Table 2).

Competition of 125I-rat amylin binding to CHO-P cells cotransfected with 100 ng of hCTRI1+ DNA and 150 ng of pcDNA3 (A), RAMP1 (B), RAMP2 (C), or RAMP3 (D) DNA. sCT, ▪; hCT, ▴; rat amylin, ▾; hCGRPα, ♦. Data are mean ± S.E. of four separate experiments. B, total binding; Bo, total binding in the absence of competing peptide; N, nonspecific binding (measured in the presence of 10−6 M sCT). Actual Bo values (cpm) for individual experiments were pcDNA3 (2400, 1900, 2300, 3300); RAMP1 (5900, 6500, 5600, 9700); RAMP2 (3400, 3400, 3400, 7700); RAMP3 (5500, 3500, 3900, 5800). Actual N values (cpm) ranged from 200 to 1200.
pIC50 values (M) for peptides in competition for binding of125I-rat amylin to CHO-P cells transfected with hCTRl1+(100 ng) and RAMP or vector DNA (150 ng)
Comparison of hCTRI1+-Based Phenotypes in CHO-P and COS-7 Cells
Comparison of the phenotypes displayed in COS-7 and CHO-P cells indicated that interaction of RAMP-induced phenotypes with either sCT or amylin was similar in both cell types (Table 1 versus 2). However, significant differences were observed in the interaction of hCT with hCTRI1+/RAMPs in the different cellular backgrounds. Although both high- and low-affinity sites were seen in CHO-P cells, only the low-affinity site was observed in COS-7 cells. Nonetheless, the affinity of hCT for the RAMP2-induced site in COS-7 cells was higher than the low-affinity site seen in CHO-P cells (P < .05, Table 1 versus 2). The behavior of CGRP in the two cellular backgrounds differed also, with the high-affinity state induced by RAMPs 1 or 3 in CHO-P cells having higher affinity than the high-affinity site in COS-7 cells (P < .05).
Analysis of hCTRI1−/RAMP Phenotypes in CHO-P Cells
Competition binding analysis of the hCTRI1−-derived phenotypes in CHO-P cells, like hCTRI1+ profiles, revealed complex binding indicative of two receptor sites (Fig. 4; Table 3). In this the pattern of binding differs substantially from hCTRI1− phenotypes in COS-7 cells where essentially monophasic curves were observed (Christopoulos et al., 1999). The interaction of sCT and hCT with hCTRI1− and either RAMPs or vector control was not significantly different. In hCTRI1−/vector-transfected cells, no high-affinity amylin binding site could be discerned, although considerable variance across experimental groups was seen (Fig. 4A; Table 3). A high-affinity site was revealed in hCTRI1−/RAMP1 and hCTRI1−/RAMP3 cotransfections, but not with hCTRI1−/RAMP2 (Fig. 4, B–D; Table 3). The low-affinity amylin site, in all receptor/RAMP combinations, had significantly higher affinity than the vector control (Table 3). Although interaction of CGRP with hCTRI1−/RAMP1 phenotype could be segregated into a high- and low-affinity component, the higher affinity site had reduced affinity compared with hCTRI1−/vector and hCTRI1−/RAMP3 high-affinity sites, and also to the high-affinity CGRP sites seen with hCTRI1+/RAMPs in COS-7 and CHO-P cells. This site, however, is not significantly different from either the hCTRI1+/RAMP low-affinity CGRP site or the hCTRI1−/RAMP1 site in COS-7 cells (Christopoulos et al., 1999), leading us to believe that these sites are equivalent.

Competition of 125I-rat amylin binding to CHO-P cells cotransfected with 100 ng of hCTRI1− DNA and 150 ng of pcDNA3 (A), RAMP1 (B), RAMP2 (C), or RAMP3 (D) DNA. sCT, ▪; hCT, ▴; rat amylin, ▾; hCGRPα, ♦. Data are mean ± S.E. of three separate experiments. B, total binding; Bo, total binding in the absence of competing peptide; N, nonspecific binding (measured in the presence of 10−6 M sCT). Actual Bo values (cpm) for individual experiments were pcDNA3 (1350, 2700, 900); RAMP1 (6800, 4300, 5600); RAMP2 (4400, 4200, 3600); RAMP3 (5300, 4700, 5300). Actual N values (cpm) ranged from 300 to 1000.
pIC50 values (M) for peptides in competition for binding of125I-rat amylin to CHO-P cells transfected with hCTRl1− (100 ng) and RAMP or vector DNA (150 ng)
The low-affinity binding sites identified in CHO-P cells with hCTRI1−/RAMP1 or hCTRI1−/RAMP3, for all competing ligands, were not significantly different from the binding sites identified in COS-7 cells cotransfected with hCTRI1− and RAMP1 or RAMP3.
Comparison of hCTRI1− and hCTRI1+Phenotypes in CHO-P Cells
Although similar in many respects, a number of significant differences were seen in the interaction of peptides with hCTRI1−- and hCTRI1+-derived phenotypes. For example, sCT had higher affinity for hCTRI1−/RAMP1 phenotype compared with hCTRI1+/RAMP1, although interaction of sCT was similar between other receptor/RAMP combinations. In contrast, no high-affinity amylin or CGRP binding sites were identified in hCTRI1−/RAMP2 combinations. Also, as discussed above, CGRP appears to lack the high-affinity interaction with hCTRI1−/RAMP1, which is apparent with hCTRI1+/RAMP1.
Analysis of Effect of RAMP Cotransfection on 125I-hCT Binding in COS-7 or CHO-P Cells Transfected with hCTRs
In general, cotransfection of RAMPs with hCTRs tended to decrease125I-hCT binding, with the greatest effect seen with hCTRI1− in COS-7 cells. However, considerable between experiment variation occurred and analysis of pooled data at 150 ng RAMP revealed no significant differences (Fig.5).

Effect of RAMP cotransfection on the expression of specific 125I-hCT binding in COS-7 cells (C, D, G, H) or CHO-P cells (A, B, E, F) transfected with either hCTRI1−(A–D) or hCTRI1+ (E–H). Cells were transfected with 100 ng of receptor DNA and increasing amounts of RAMP DNA or vector control [B, D, F, H; representative data from a single experiment (n = 2–4)]. Pooled data for cells transfected with 100 ng of receptor DNA and 150 ng of RAMP or vector DNA are shown in A, C, E, and G (n = 4–7). Vector (pcDNA3) control, ▪; RAMP1, ▾; RAMP2, ▴; RAMP3, ♦.
Competition Binding Analysis
COS-7 Cells.
In hCTRI1+-transfected COS-7 cells, cotransfection with RAMP had little effect on any of the competing peptides, with only a small decrease in sCT affinity seen with RAMP2 (Table 4). Similarly, RAMP cotransfection had little impact on hCTRI1−-transfected cells (Table5). Comparison of hCTRI1+- and hCTRI1−-derived phenotypes in COS-7 cells indicated the presence of a high-affinity sCT binding site in hCTRI1−-transfected cells that was not apparent in hCTRI1+-transfected cells (Table 4 versus 5).
pIC50 values (M) for peptides in competition for binding of125I-hCT to COS-7 cells transfected with hCTRl1− (100 ng) and RAMP or vector DNA (150 ng)
CHO-P Cells.
In hCTRI1+-cotransfected CHO-P cells, high- and low-affinity sites were observed for all peptides, with the exception of CGRP competition for binding in hCTRI1+/vector cotransfections (Table6). RAMP cotransfection had relatively little effect on CT peptide binding, inducing only a small increase in the proportion of high-affinity hCT sites with RAMP2 and a decrease in the number of high-affinity sites with RAMP3. All three RAMPs increased the proportion of high-affinity amylin and CGRP binding sites, with RAMP1 having the greatest impact (Table 6).
pIC50 values (M) for peptides in competition for binding of125I-hCT to CHO-P cells transfected with hCTRl1+ (100 ng) and RAMP or vector DNA (150 ng)
Few significant differences occurred between hCTRI1+-transfected CHO-P cells and hCTRI1+-transfected COS-7 cells. In CHO-P cells, both high- and low-affinity sCT binding was seen, whereas binding in COS-7 cells was principally to a single low-affinity site. The other differences seen were higher affinity, for both high- and low-affinity amylin sites with hCTRI1+/RAMP3 in CHO-P cells versus COS-7 cells and a greater proportion of high-affinity CGRP binding sites with hCTRI1+/RAMP1 in CHO-P cells (P < .05; Table 4 versus 6).
In hCTRI1−-cotransfected CHO-P cells, high- and low-affinity sites were seen for all peptides in all transfection combinations (Table 7). RAMP1 cotransfection increased the proportion of high-affinity sCT sites, whereas all three RAMPs increased the proportion of high-affinity amylin sites (Table 7). RAMPs 1 and 2 also increased the proportion of high-affinity CGRP sites, although the hCTRI1−/RAMP2 CGRP site had lower affinity than the vector control (Table 7). The only other significant difference was an increase in the affinity of the low-affinity hCT site with hCTRI1−/RAMP3 cotransfection. The profiles of hCTRI1−-derived phenotypes in COS-7 and CHO-P cells were generally similar, although the proportion of high-affinity amylin and CGRP sites tended to be greater in CHO-P-transfected cells.
pIC50 values (M) for peptides in competition for binding of125I-hCT to CHO-P cells transfected with hCTRl1− (100 ng) and RAMP or vector DNA (150 ng)
Expression of Endogenous RAMPs in COS-7 and CHO-P Cells
Northern analysis of endogenous RAMP mRNA expression in COS-7 and CHO-P cells failed to yield any appreciable signal, indicating that the expression of endogenous RAMPs is low in both cell lines (data not shown). Control Northerns using rat tissue or transfected cell RNA confirmed that the probes could detect RAMP mRNA.
RT-PCR of COS-7 and CHO-P total RNA yielded unique products of 410, 456, and 400 bp that were the expected sizes for RAMP1, 2, and 3, respectively (Fig. 6). The yield of RAMP2 was comparatively lower than that of RAMP1 or RAMP3, and the yields of RAMP1 and RAMP3 were also similar across the two cell lines.

RT-PCR amplification of RAMP1, RAMP2, and RAMP3 from CHO-P (left) and COS-7 (right) mRNA. Five micrograms of total RNA was reverse transcribed using random hexamer primers and then were subjected to increasing cycles of PCR amplification using human RAMP1-, RAMP2-, or RAMP3-specific primer sets. PCR products were electrophoresed on 1% agarose and stained.
Discussion
High-affinity amylin receptor-like phenotypes result from the coexpression of the CTR gene product with RAMPs. This has been demonstrated in COS-7, CHO-K1 (Christopoulos et al., 1999), and rabbit arterial endothelial cells (RAEC) (Muff et al., 1999). In this study, we compared the ligand binding characteristics of two isoforms of hCTR when coexpressed with individual RAMPs in COS-7 and CHO-P cells. The results, in addition to confirming the previous findings, show several interesting features relating to the nature of receptor/RAMP interactions.
The receptor isoforms used in this study differ only by the presence (hCTRI1+) or absence (hCTRI1−) of a 16-amino-acid insert in the predicted first intracellular loop. Previously we revealed that the I1− isoform yields two distinct amylin receptor phenotypes when coexpressed with either RAMP1 or RAMP3 in COS-7 cells. The RAMP1-derived receptor phenotype also showed high affinity for CGRP, whereas in the RAMP3-derived phenotype, the affinity for CGRP was greatly reduced (Christopoulos et al., 1999). The combination of RAMP2/hCTRI1− did not substantially alter amylin binding in either COS-7 (Christopoulos et al., 1999) or RAEC (Muff et al., 1999), although we have demonstrated recently that low level specific amylin binding can be induced in COS-7 cells (Zumpe et al., 2000). In contrast, the present results demonstrate that the hCTRI1+ yields amylin receptor phenotypes with all three RAMPs in COS-7 cells. The receptor phenotype generated from the I1+ isoform in COS-7 displays two-site (high- and low-affinity) binding, in contrast to the single site binding seen with hCTRI1−/RAMP in the same cells. The low-affinity sites for hCTRI1+/RAMP-1 or -3 (which comprised the majority of sites) were equivalent to the single affinity sites seen for hCTRI1−/RAMP-1 or -3 combinations, and in general follow the pattern of interaction seen with hCTRI1− in COS-7 cells with RAMP1 generating a receptor phenotype with high affinity for both amylin and CGRP, whereas RAMP3-derived phenotype has relatively poor affinity for CGRP.
In this study, we also demonstrate that RAMP2 is capable of modulating the phenotype of the CTR gene product and that the resultant phenotype is distinct from that of RAMPs 1 and 3. This differs from RAMP interaction with CRLR where the RAMP2- or RAMP3-generated adrenomedullin receptors are essentially equivalent (Fraser et al., 1999). Although varying with receptor isoform and cell background, the RAMP2-derived amylin receptor phenotype tended to have a lower affinity interaction with amylin or CGRP and higher affinity interaction with hCT, than the corresponding RAMP1 phenotype.
Amylin binding properties of hCTRs expressed in CHO-P cells differ from those in COS-7 cells in a number of respects. First of all, both isoforms of hCTR, in the absence of RAMPs, have a low level (∼1–1.5% of total cpm added) of 125I-amylin binding in CHO-P cells, whereas specific amylin binding with hCTR alone is negligible in COS-7 cells. Secondly, in CHO-P cells, significant increases in 125I-amylin binding occur when either receptor isoform is coexpressed with any of the three RAMPs. However, this increase (2- to 3.5-fold of the basal) is less than that observed in COS-7 cells where, with the exception of hCTRI1−/RAMP2, at least a 5-fold increase in125I-amylin binding is seen. The mean125I-hCT binding was not significantly different between COS-7 and CHO-P cells (see Fig. 5). This similarity in hCT binding between cell lines was consistently maintained in individual experiments where levels of receptor expression were varied (results not shown). These results suggest that the differences in amylin binding between cell types are not entirely attributable to differences in transfection efficiency. Nonetheless, an effect due to RAMP/receptor stoichiometry cannot be ruled out. The third distinctive feature in CHO-P cells is that ligand binding, following hCTR/RAMP coexpression, is more complex, and the biphasic nature of the competition for both125I-amylin and 125I-hCT more prominent. As discussed earlier, the I1+ isoform shows greater evidence of multistate binding than I1− in both cell lines, but this is greater in CHO-P than in COS-7. Effect of host-cell environment on the receptor phenotype is also illustrated by the pharmacological variation between the amylin receptor phenotypes generated from the C1a isoform of rat CTR (the rat equivalent of insert-negative isoform) and RAMPs expressed in either HEK293 or COS-7 cells (Christopoulos et al., 1999). In HEK293, amylin binding was induced by RAMP1 only, although, in these cells CTR is stably expressed with additional transient expression of RAMPs, whereas in COS-7 and CHO-P both receptor and RAMPs are transiently expressed.
Although it failed to achieve statistical significance, binding of125I-hCT to hCTRI1− in both cell types was decreased by cotransfection with each of the RAMPs. This is similar to the observation of Muff and colleagues (1999) in RAEC, where RAMP1 or RAMP3 cotransfection with hCTRI1− demonstrated a significant inverse relationship between 125I-amylin and125I-hCT binding. In contrast, much less effect was seen on binding of 125I-hCT to hCTRI1+ transfected cells following RAMP cotransfection.
We examined whether there is differential expression of endogenous RAMPs between the two cell lines that might explain these differences. Endogenous RAMP1, 2, or 3 mRNA, in both cell types, was undetectable by Northern analyses using either human, mouse, or CHO-P-derived-RAMP cDNA probes, even under low-stringency hybridization conditions. However, it is evident that mRNA for each of the three RAMPs is expressed in low abundance in both COS-7 and CHO-P cells as revealed by RT-PCR amplification. Buhlmann et al. (1999) also failed to detect RAMP1 mRNA in COS-7 cells by Northern analysis, and detected only a very weak signal for RAMP2 mRNA. The minimal expression of endogenous RAMPs in these cells is consistent with their poor ability to bind amylin with CTR in the absence of heterologously coexpressed RAMPs. Based on RT-PCR, there is no obvious difference in the levels of mRNA for each of the RAMPs in the two cell lines. However, variation in the receptor/RAMP stoichiometric ratios in the two cell lines may potentially contribute to the between-cell-type variation.
These findings suggest that the difference between the two host cell environments in determining the nature of the receptor phenotypes generated from hCTR and RAMPs involves an additional component(s) of the cellular background. The way in which each of the two receptor isoforms interacts with this component may be relevant to the resulting receptor phenotype. RAMPs were first identified by McLatchie et al. (1998) as a family of low molecular weight proteins capable of associating with CRLR to generate either a CGRP receptor or an adrenomedullin receptor. The mechanism by which RAMPs convert a particular receptor to another is not well understood, although they have been shown to be involved in the regulation of terminal glycosylation and trafficking of CRLR to the cell surface (McLatchie et al., 1998; Fraser et al., 1999). However, unlike CRLR, CTR in all cells studied is efficiently expressed at the cell surface in the absence of RAMP cotransfection (Christopoulos et al., 1999). Potential host-cell factors would include the level and type of G protein. It is well known that the amount of G protein available for ternary complex formation influences the affinity states of GPCRs for their ligands. This is particularly important in heterologous expression systems that invariably result in overexpression of receptors in relation to available amounts of G protein (Kenakin, 1997), as illustrated with secretin receptors expressed in COS cells. Under normal conditions, only 1.8% of receptors formed high-affinity complexes with125I-secretin. After cotransfection with cDNA for Gαs protein, this increased to 15% (Ishihara et al., 1991).
The major functional difference observed between the hCTRI1− and hCTRI1+ is in their ability to modulate second messenger pathways. Our results, indicating essentially no difference in 125I-hCT binding between the two receptor isoforms in the same host environment, agree with earlier findings that the presence or absence of the 16-amino-acid insert has little impact on peptide binding affinities (Nussenzveig et al., 1994; Moore et al., 1995). However, whereas hCTRI1− signals effectively through activation of adenylate cyclase-, phospholipase C-, or phospholipase D-dependent pathways, the adenylate cyclase responsiveness is decreased and the phospholipase C- and D-dependent signaling is abolished in the hCTRI1+ isoform (Moore et al., 1995; Naro et al., 1998). This difference in cellular response is likely due to differences in the efficiency of G protein coupling (or uncoupling). Thus, variation in the formation of receptor/G protein/RAMP complexes may, at least in part, contribute to the differences in observed phenotype between receptor isoforms and host cell environment.
In conclusion, the concept of G protein-coupled receptor/RAMP interaction leading to the generation of different receptor phenotypes has now been extended to include both CRLR and CTR gene products. In this study we examined the result of this interaction in different host cell environments using two RNA splice variants of the same receptor. It was found that the pharmacological characteristics of the receptors resulting from RAMP/receptor coexpression do indeed vary depending on which isoform of the receptor is studied and the cellular background in which it is expressed. The results indicate that RAMP/receptor interaction is complex and that other cellular factors, such as the level and/or type of G protein, are important in the derivation of receptor phenotype.
Footnotes
-
Send reprint requests to: Dr. Patrick M. Sexton, Molecular Pharmacology Laboratory, Department of Pharmacology, The University of Melbourne, 3010, Victoria, Australia. E-mail:p.sexton{at}pharmacology.unimelb.edu.au
-
↵1 This work was supported by grants from the National Health and Medical Research Council of Australia. P.M.S. is a Research Fellow of the National Health and Medical Research Council of Australia.
- Abbreviations:
- CTR
- calcitonin receptor
- CGRP
- calcitonin gene-related peptide
- CT
- calcitonin
- hCTR
- human calcitonin receptor
- RAMP
- receptor activity modifying protein
- CHO
- Chinese hamster ovary
- CHO-P cells
- a variant of CHO cells stably transfected with P-selectin
- CRLR
- calcitonin receptor-like receptor
- FBS
- fetal bovine serum
- sCT
- salmon calcitonin
- hCT
- human calcitonin
- DMEM
- Dulbecco's modified Eagle's medium
- RT-PCR
- reverse transcription-polymerase chain reaction
- RAEC
- rabbit arterial endothelial cells
- Received January 10, 2000.
- Accepted March 14, 2000.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}