


Abstract
Ethanol (EtOH) reversibly activates large conductance, Ca++-activated K+ (BK) channels in rat neurohypophysial terminals, an effect that probably contributes to the inhibition of vasopressin release by this drug. Heterogeneity in the terminal channel population makes it difficult to determine the mechanisms underlying this activation. Here, we report the effects of EtOH on the steady-state activity of BK channels cloned from mouse brain (mslo, α subunit) and expressed inXenopus oocytes. EtOH reversibly increased mslochannel activity in excised patches, showing a potency (EC50 = 24 mM) similar to that reported using native channels. EtOH activation was observed under conditions that make it highly improbable that it is mediated by freely diffusible second messengers, or secondary to G-protein modulation. Rather, it probably results from a functional interaction between the drug and the channel α subunit. Activation occurred without increase in the number of functional channels present in the patch and resulted from actions that were a function of EtOH concentration: at ≤10 mM, activation was due to a decrease in the channel mean closed time, whereas between 25 and 100 mM, activation was due to both a decrease in the mean closed time and an increase in the mean open time. The characteristic high unitary conductance and ionic selectivity of BK channels were unaltered by the drug. Whereas the voltage dependence of channel gating remained unchanged, channel activation mediated by the response of the Ca++-sensing site(s) to increases in the concentration of intracellular calcium, [Ca++]ic, was reduced by EtOH. This finding is consistent with EtOH and [Ca++]ic behaving functionally as partial and full agonists of mslo channels, respectively. Because the potentiation of mslo activity by the drug decreased as Ca++ levels were increased, EtOH-activation of BK channels would be most evident when [Ca++]ic is near resting levels, rather than during periods of high activity and Ca++ influx.
CAK channel activation increases K+ efflux, which repolarizes/hyperpolarizes the membrane potential, decreasing neuronal excitability (Lang and Ritchie, 1987). It has been speculated that drug-induced increases in CAK currents might contribute to the depressant action of EtOH and other sedative/hypnotic drugs on central neurons (Krnjevic, 1972). Several groups have reported increases in CAK responses by EtOH at relevant concentrations. Ca++-dependent afterhyperpolarization and Ca++-dependent Rb86 efflux were used as indexes of CAK responses in these studies (Carlen et al., 1982; Harris and Cadwell, 1985; Madsen and Edeson, 1990), which makes it difficult to address which CAK conductance was involved and the mechanisms underlying the increase in current. We recently found that EtOH, at circulating concentrations found after moderate drinking, reversibly activates BK channels in rat neurohypophysial terminals (Dopico et al., 1996a), which probably contributes to the decreased vasopressin release and plasma levels found after moderate drinking (Dopico et al., 1995a).
BK channel activity is increased with cell depolarization and/or elevated [Ca++]ic (Magleby and Pallota, 1983;Latorre et al., 1989). Thus, it is possible that EtOH activates BK channels by increasing the sensitivity of the channel to voltage and/or [Ca++]ic. In neuroterminals, heterogeneity in the response of native BK channels to changes in [Ca++]ic or transmembrane voltage (Wanget al., 1992; Dopico et al., 1996a) prevented us from testing this hypothesis.
It is postulated that native BK channels from mammalian tissue are heterooligomeric channels that consist of pore-forming (α) and regulatory (β) subunits (McManus et al., 1995). BK channel α subunits have been cloned from cDNA obtained from different sources, including Drosophila melanogaster (dslo) and mouse brain (mslo). These cloned channels have been expressed inXenopus oocytes and characterized electrophysiologically (Atkinson et al., 1991; Butler et al., 1993).
In this work, we used single-channel recordings from excised patches to examine the acute action of EtOH on the activity of mslo BK channels expressed in Xenopus oocytes (where no endogenous β subunits have been reported). Thus, we can address whether the drug interacts (directly or via membrane-bound mediators) with the α subunit, independently of modulatory subunits. A population of cloned channels with nonvariant voltage and/or Ca++-sensitivity also allows us to determine the role of changes in the voltage- and/or Ca++-sensitivity of the channel in EtOH action. Because the electrophysiological properties examined are associated with functional domains in the α subunit (Weiet al., 1994), EtOH-modified properties also allow preliminary speculation on which regions of this subunit are targeted by EtOH.
Results show that EtOH reversibly activates mslo channels with a potency (EC50 = 24 mM) very close to that reported for native channels and at blood levels considered legal intoxication. EtOH-induced activation results from drug actions on channel gating—in particular, a marked increase in the frequency of channel openings. Channel activation by EtOH is not accompanied by changes in the channel unitary conductance, ion selectivity or voltage dependence of gating. Although EtOH activation of mslo channels is not affected by voltage, it is critically modulated by [Ca++] at the intracellular side of the membrane. Preliminary data were presented in abstract form (Dopico et al., 1995b; Dopico et al., 1996b).
Methods
Use of Xenopus oocytes and injection.
Xenopus laevis females were purchased from commercial breeders in the United States and were maintained in artificial pond water on a 12-h light/dark cycle. In this habitat, they do not show seasonal breeding behavior, so oocytes are available throughout the year. Stages V and VI oocytes were predominantly used, because they transcribe mRNA into channels efficiently. Oocytes were removed and kept in a Ca++-free ND-96 solution (mM): 82.5 NaCl, 2 KCl, 1 MgCl2, 5 HEPES, pH 7.5, containing 2 mg/ml collagenase (type IV, Sigma, St. Louis, MO) for 2 to 4 h, in order to remove the follicular layer. After defolliculation, oocytes were transferred to a Ca++-containing ND-96 saline (mM): 96 NaCl, 2 KCl, 1.8 CaCl2, 5 HEPES, pH 7.5, supplemented with 2.5 mM sodium pyruvate and 2 mg/ml gentamicin. Injection of mRNA was conducted using a Drummond micropipette modified for microinjection (Drummond Scientific Co., Broomall, PA). Injection volumes of mRNA (0.1–1 μg/μl) ranged from 18.6 to 50.6 nl. The use of varying concentrations and volumes, as well as different intervals between injection and electrophysiological recording (24–72 h), gave us crude control over the amount of BK channel protein expressed in the oocyte’s membrane.
RNA preparation and analysis.
BK channels were expressed inXenopus oocytes by injection of mRNA transcribed in vitro from complementary DNA (cDNA) coding for the mbr5 variant of the mslo BK channel, derived from the Slowpokelocus of mouse brain (Butler et al., 1993). Briefly, polyadenylated RNA transcripts were synthesized from Sal 1 linearizedmslo templates using T3 RNA polymerase. Full-length cDNA from mslo was a generous gift from Dr. Lawrence Salkoff (Washington University).
Single-channel recordings.
Before recordings, oocytes were placed into a dish containing a hypertonic solution (mM): 200 K+ aspartate, 20 KCl, 1 MgCl2, 10 ethylene glycol-bis(β-aminoethyl ether) N,N,N′,N′-tetraacetic acid (EGTA), 10 HEPES, pH 7.4, for 10 min. With this treatment the oocytes shrunk, which allowed us to remove the vitelline layer with forceps, exposing the oocyte membrane for subsequent patch recording. Then the oocytes were placed back into ND-96 saline (in this case without gentamicin; for composition, see above) for 10 to 15 min before recording. Single-channel recordings were obtained from excised I/O membrane patches using standard patch-clamp techniques. All solutions were made with ultrapure 18 MΩ and bidistilled water and with high-grade salts. Unless otherwise stated, recordings were obtained in “symmetric conditions”—that is, the same solution bathed both the extracellular and the cytosolic sides of the patch membrane (mM): 145 K+gluconate, 5 EGTA, 1 MgCl2, 15 HEPES, 10 glucose, pH 7.35, containing varying amounts of free Ca++. The free Ca++ concentration was adjusted to the desired nominal value by adding CaCl2. The calculation of the final, nominal [Ca++]free in all solutions was done according to Fabiato (1988). In some cases, the extracellular surface of the patch was exposed to a [Ca++]free of 3 to 10 μM. This relatively high concentration of Ca++, together with the Mg++ present in this solution, helped to increase gigaseal formation. In experiments to study the Ca++-sensitivity of BK channels, the intracellular surface of the patch was exposed to (bath) solutions that differed in their final [Ca++]free (0.01–1 μM).
Electrodes were pulled from glass capillaries (Drummond Scientific Co.). The shank of each electrode was coated with Sylgard (Dow Corning Co., Midland, MI) to reduce capacitance and electrical noise. Immediately before recording, the tip of the electrode was fire-polished on a microforge (Narashige, Kyoto, Japan) to give resistances of 8 to 10 MΩ when filled with extracellular solution (for composition, see above). An Ag/AgCl electrode pellet was used as the ground electrode. After excision from the oocyte, the membrane patch was exposed to a stream of bath solution containing the desired concentration of EtOH and/or [Ca++]freeflowing from a micropipette (1-mm diameter, WPI Inc.). Deionized (100% purity) EtOH (American Bioanalytical, Natick, MA) was freshly diluted in bath solution immediately before each experiment. Superfusion with dextrose isosmotically replacing EtOH was used as control for EtOH superfusion. All experiments were carried out at room temperature (20°C).
Single-channel currents were recorded using a patch-clamp amplifier (EPC7, List Electronics, Darmstadt, Germany) at a bandwidth of 3 kHz and were low-pass filtered at 1.5 kHz using an eight-pole Bessel filter (model 902LPF, Frequency Devices, Haverhill, MA). Data were acquired and stored using an A/D converter and an IBM-compatible computer. Data acquisition and analysis were performed using pCLAMP software, version 6.0.2 (Axon Instruments, Foster City, CA). Data with noisy or drifting baseline were not considered for analysis (less than 1% of records were rejected). Data were sampled at a bandwidth of 5 kHz. Durations of open and closed times were measured with half-amplitude threshold analysis. A maximum-likelihood minimization routine was used to fit exponential curves to the distribution of open and closed times. Determination of the minimum number of exponential terms for adequate fit was established using a standard F statistic table (significance level <.01).
As an index of channel activity we used NPo, the product of the total number of functional channels present in the patch membrane (N) and the probability that a particular channel is open under steady-state conditions (Po).NPo values were calculated from the area under the curve (AUC) of the Gaussian fit of all-points amplitude histograms. Assuming a Poisson distribution, NPo = ∑xi, with i = 1 … n, wheren is the maximum number of simultaneous conducting channels during the observation period, and x is the AUC corresponding to each opening. For construction of these histograms and of idealized records, data obtained from multiple episodes at the desired step potential were sampled for a total time of no less than 10 s. In multichannel patches of unknown N, and in the presence of “overlapping” openings, knowingNPo and the number of openings (#o) during a long period of observation (T) enabled us to calculate the mean open time (to) from the relationship
Similarly, the mean closed time (tc) could be obtained from
The single-channel conductance (γ) was obtained from the unitary current amplitude (i)-voltage relationship (i/Vplot). Voltages given correspond to the potential at the intracellular side of the patch. Data are expressed as mean ± S.E.M.
Results
EtOH-induced increase of mslo channel steady-state activity in excised I/O patches is reversible and concentration-dependent.
The acute application of EtOH to the cytosolic surface of I/O patches reversibly increased the steady-state activity (NPo) of mslo channels expressed in Xenopus oocytes (fig.1). A flow or osmotic effect was ruled out by exposing the excised patch to a stream of solution with dextrose isosmotically replacing EtOH (control perfusion), which did not change channel activity. Similarly to what we previously described for the activation by EtOH of BK channels in neurohypophysial terminals (Dopicoet al., 1996a), whereas EtOH’s action was almost immediate in onset (<6 s after switching the perfusion to EtOH-containing solutions), it took approximately 5 to 6 min of drug-free perfusion for channel NPo to return to pre-EtOH levels in all cases. Because the Ca++-sensing sites of the channel seem to be located at the intracellular side of the membrane (McManus, 1991), the fact that EtOH activation was observed in I/O patches more than 15 min after patch excision, with [Ca++]ic highly buffered, indicates that this activation is not mediated by changes in the levels of [Ca++]ic or other freely diffusible cytosolic second messengers. Because EtOH activation was observed during I/O recordings using symmetric [Ca++] at positive potentials (i.e., with Ca++ flowing outward), EtOH-induced increase in mslo NPo was not caused by a transient elevation of [Ca++] at the intracellular side of the patch in the vicinity of the mslo channel as a consequence of an EtOH-induced transmembrane flux of Ca++. Consequently, EtOH’s activation of mslo channels expressed in oocytes after injection of the α subunit seems to be due to a direct interaction of EtOH with this protein or some closely associated entity in the patch membrane that, eventually, interacts with themslo α subunit.

EtOH (50 mM) reversibly increases the activity (NPo) of mslo channels expressed inXenopus oocytes. Representative single-channel recordings obtained from I/O patches before (upper panels), during (middle panels), and 6.5 min after (lower panels) exposure of the cytosolic side of the patch to EtOH. Channel openings are shown as upward deflections; arrows indicate the baseline level. NPo values were obtained from all-points histograms, constructed from 72 to 126 s of recording under each particular condition. The composition of the solution facing the intracellular and extracellular sides of the patch (symmetric conditions) is described in “Methods.” In this case, free [Ca++]ic∼ 32 nM. The membrane potential was set to +20 mV.
EtOH’s activation of mslo channels was concentration-dependent from 10 to 100 mM, and it showed a plateau at 100 to 200 mM (fig. 2A). At the highest concentration tested, individual patches showed high variability; higher concentrations than 200 mM resulted in loss of gigaseals. The EC50, obtained from figure 2a by linear interpolation, was 24.3 ± 4.8 mM, very close not only to the EC50 for BK channel activation of native channels (Dopico et al.,1996a), but also to the circulating level of alcohol that constitutes legal intoxication in the United States (20–22 mM).

A) The activation of mslo channels by EtOH is concentration-dependent, reaching a “plateau” at 100 to 200 mM EtOH. The EC50, obtained by linear interpolation, is 24.3 ± 4.8 mM. Results are expressed as the ratio ofNPo values obtained in the presence and absence of EtOH, determined in the same patch. Each point of the graph is the mean ± S.E. of 4 to 20 determinations; each determination was obtained in a different patch. In all cases, the patch membrane potential was set at 20 to 40 mV, and free [Ca++]ic was 31.6 to 100 nM. B) Logit-log plot of mslo channel activity as a function of [EtOH]. Maximum NPo (maxNPo) was calculated as the mean of NPo values obtained at 100 and 200 mM EtOH, because the EtOH-induced increase inNPo values at those concentrations reached a plateau, as shown in panel A. Data plotted were obtained at 10, 25 and 50 mM EtOH. The Hill coefficient (n, slope of the graph) is 0.87, not significantly different from 1.
The presence of a maximal response in mslo NPoto EtOH allowed us to construct a logit-log plot (wherein responses to an agonist are related to the maximal response observed, in a double logarithmic plot) (Taylor and Insel, 1990) to assess the response ofmslo channel activity to EtOH. In this type of plot, the intercept on the abscissa is the EC50, and the slope gives the equivalent of n, the Hill coefficient (fig. 2B). We obtained an EC50 of 21 mM and a slope of 0.87. The fact that n was not significantly different from 1 indicates that the functional interaction between EtOH and the mslo α subunit does not require cooperativity among multiple binding sites.
EtOH modifies channel gating properties.
We have shown above a reversible increase in channel NPo by EtOH. In the vast majority of cases, we dealt with multichannel patches in whichN was unknown. Even in this situation, an increase inNPo by EtOH may be interpreted as an increase inPo. Because of the fast and reversible nature of the activation, it is unlikely that EtOH causes an increase inNPo by introducing more channel-formingmslo α subunits into the patch. Also, in excised (i.e., “cell free”) patches, the source of these additional channels would have to be the limited area of membrane under the rim of the pipette, which also makes this possibility unlikely. Nevertheless, we could not rule out a priori the possibility that EtOH might increase channel NPo by unmasking “silent” channel proteins already present in the patch. However, we were able to observe an increase in channel (N)Po by EtOH in I/O patches containing only one functional mslo channel protein (as described below for fig.4). Furthermore, the increase in channel NPo by 50 mM EtOH (∼190% of control) observed in multichannel patches of undetermined N (fig. 3) at 40 mV and 100 nM free [Ca++]ic was similar to the increase in Po produced by the same concentration of the drug in a single-channel patch (∼183% of control; dwell-time distributions for this channel are shown in fig. 4) under identical recording conditions. Thus, the increase in channelPo produced by EtOH totally accounts for the drug-induced increase in channel NPo. In conclusion, the reversible increase in channel activity produced by EtOH occurs without any change in N.

Dwell-time distributions of mslo channels in control (panel A, open times; panel B, closed times) and 50 mM EtOH (panel C, open times; panel D, closed times) obtained from a single-channel patch. The membrane potential was set at +40 mV, and the free [Ca++]ic was 100 nM. Exponential curves were fitted using a maximum-likelihood minimization routine. The different components (τ) of each exponential distribution are indicated by dotted lines. In both control and EtOH, both the open-times distribution and the closed-times distributions could be well fitted with two exponentials. A statistical comparison of different fitting models was done using a standard Fstatistical table (P < .01). Both the actual time value (in milliseconds) and the contribution to the total distribution (as percentages in parentheses) of each component are shown at the right of each panel. These percentages reflect the relative contribution of each component to the exit from the state (open or closed) considered.
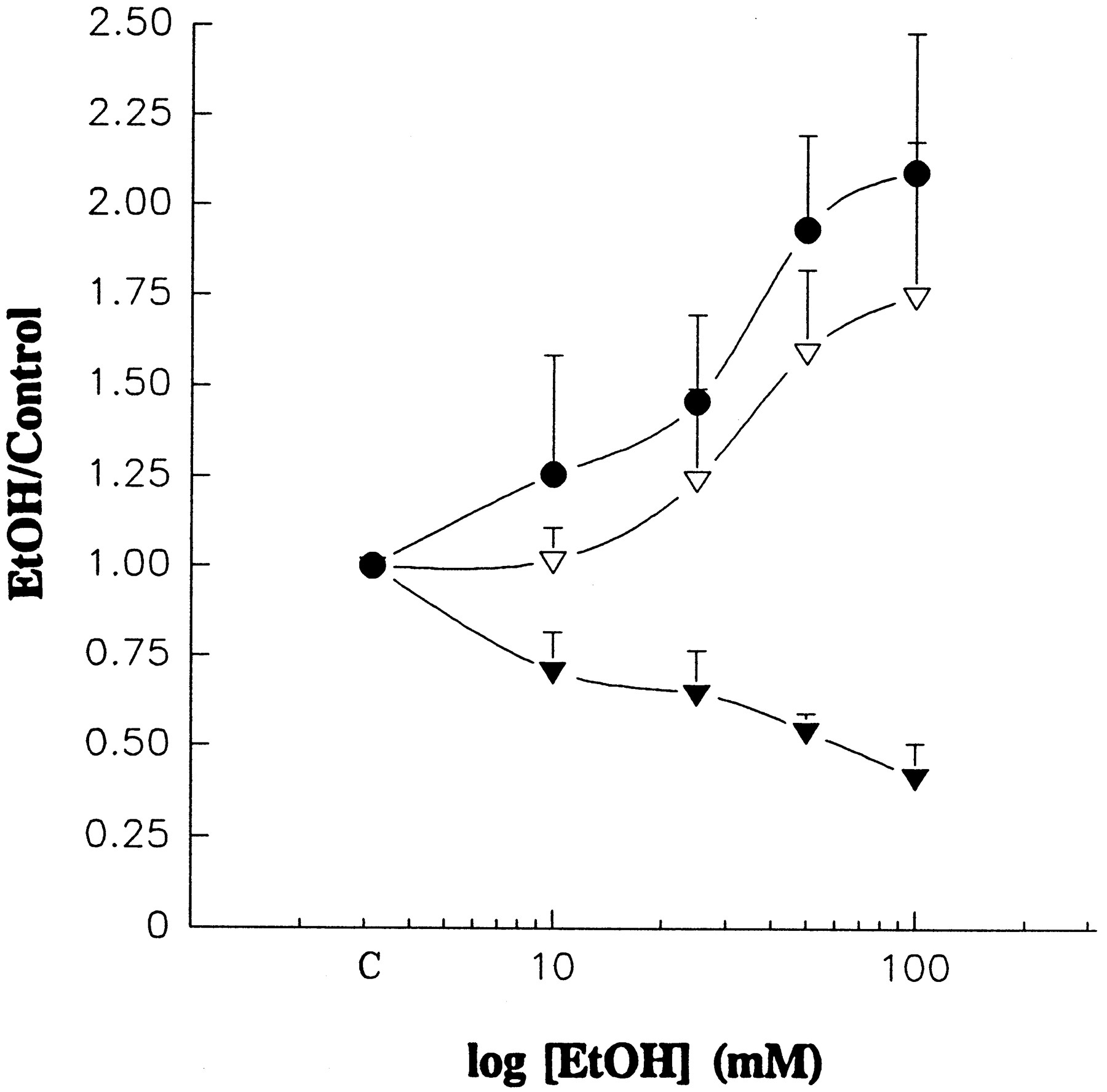
Changes in mean open time (to, hollow triangles), mean closed time (tc, filled triangles) and steady-state activity (NPo, filled circles) of mslo channels as a function of [EtOH]. Results are expressed as ratios of values obtained in the presence and absence of EtOH. Each point of the graph is the mean ± S.E. of 3 to 9 determinations; each determination was obtained in a different patch. Individual points at a given EtOH concentration for all three variables were simultaneously determined in the same patch. In all patches, the patch membrane potential was set at 40 mV, and free [Ca++]ic was 100 nM. The composition of the solution facing the intracellular and extracellular sides of the patch (symmetric conditions) is described in “Methods.” Because data are expressed as ratios, there is a limit for ratios describing decreases induced by the drug (i.e., EtOH-induced decreases intc ratios are limited to values between 1 and 0), whereas EtOH-induced increases in NPo andto ratios can attain values above 1. C: control values.
An increase in channel Po can be explained by an increase in the mean open time (to), an increase in the frequency of openings (i.e., a decrease in the mean closed time, tc) or both. Even in multichannel patches of unknown N, and in the presence of “overlapping” openings (a situation that often occurred in the presence of EtOH, an activator of the channel), knowledge ofNPo and of the number of openings during a long period of observation allowed us to calculateto, as described in “Methods.” The determination of tc in multichannel patches cannot be interpreted, because the closed intervals that contribute to the mean value may be terminated by the reopening of the channel that closed last or by the opening of another channel, which leads to artificially low tc values. However, making the series of assumptions explained in “Methods” enabled us to evaluate changes in tc as a function of [EtOH] from multichannel patches.
Figure 3 shows changes in three different variables (NPo, to andtc) as a function of [EtOH] from multichannel patches, at fixed potential (40 mV) and free [Ca++]ic (100 nM). Individual points at a given EtOH concentration for all three variables were obtained from the same patch. At low EtOH concentrations (10 mM), increases inNPo are not accompanied by increases into, which indicates that at this low concentration, the increase in NPo by the drug is totally explained by the observed decrease intc (−30%). On the other hand, at concentrations greater than 10 mM, increases into are also observed, which indicates that within this range of concentrations, the increase inNPo by EtOH is due to a combination of increases in to (24.1 to 74.8%) and decreases intc (−35.3 to −58.3%), both phenomena being concentration-dependent.
To address which specific components of the channel lifetime distributions were affected by EtOH, we examined the effect of the drug in patches where N = 1. This condition was obtained by decreasing the amount of mRNA injected and the interval between injection and recording. The N value was estimated by holding the patch at depolarized voltages and applying solutions containing a high free [Ca++] (usually 10 μM) to the cytosolic side of the membrane, effectively raisingPo to approximately 1 (Po ≥ 0.89). Under this condition, the maximal level of openings observedduring a long period of time (>10 min) was considered to represent N. The ideal situation would be to obtain routinely patches in which N = 1; this would make it possible to test kinetic models while manipulating voltage, [Ca++]ic and [EtOH] in various combinations. Unfortunately, probably because of channel clustering in the oocyte membrane, we were able to obtain this condition only in very few cases (2 out of 24), the more common situation being patches with either N ≥ 5 or no channels at all. The clustering ofmslo α subunits in the oocyte membrane is consistent with previous reports about clustering of native BK channels in Rana pipiens hair cells, demyelinated axons from Xenopus and mouse N1E-115 neuroblastoma cells (Roberts et al., 1990;Leinders and Vijverberg, 1992; Wu et al., 1993).
Dwell-time distributions from a single-channel patch are shown in figure 4. Openings could be well fitted with two exponentials, with time constants of 1.42 and 4.93 ms and contributions to the total fit area (i.e., to the total time spent in open states) of 63.5 and 35.6%, respectively (fig. 4A). In the presence of 50 mM EtOH, we also detected two similar components 1.40 and 4.69 ms in duration, which now contribute 25.4 and 74.6% to the total time spent in open states, respectively (fig. 4C). When the duration of these components was weighted, we obtained for this particular patch to values of 2.67 and 3.85 ms, in the absence and presence of 50 mM EtOH. Thus the increase into produced by EtOH essentially results from a change in the relative contributions of the two time constants to the total fit area, being the channel driven away from brief open states toward longer open states. The resulting increase into (+44.9%) is very close to that calculated from multichannel patches, shown in figure 3. This similarity suggests that the EtOH-induced changes in the duration and relative contributions of the components of the open-time distribution obtained from single-channel patches may also account for the overall increase in to produced by the drug in multichannel patches.
Figure 4 also shows the distribution of closed times. In the absence of ethanol, this distribution could be well fitted with two exponentials, with time constants of 1.64 and 30.74 ms and contributions to the total fit area (i.e., the total time spent in closed states) of 64.7 and 35.3%, respectively (fig. 4B). Whereas the relative contributions of these components to the fit were barely modified by 50 mM EtOH, the drug reduced the duration of the brief component to 1.51 ms and, more important, dramatically reduced the duration of the long component to 14.94 ms (fig. 4D). These changes result in an EtOH-induced decrease in tc from 11.91 to 5.56 ms (−53.3%), which matches the decrease in tcproduced by the drug in multichannel patches (fig. 3). The results reported in figures 3 and 4 indicate that 10 mM EtOH destabilizes the long closed state of the channel, whereas 25 to 100 mM EtOH modifies transitions leading away from both open and closed states, destabilizing the long closed state and shifting the channel population from briefer to longer openings. The resulting increase in bothto and frequency of channel openings explains the overall increase in steady-state activity (Po) produced by the drug.
EtOH does not affect high permeability and selectivity for K+.
A remarkable property of all BK channels is the combination of a large conductance and an exquisite selectivity for K+: they are practically impermeable to Na+(Latorre, 1994). We previously observed, using native BK channels, that the enhancement of BK currents by EtOH is completely explained by an increase in channel NPo, with no major changes in single-channel conductance and reversal potential in symmetric conditions (Dopico et al., 1996a). In the present study, we further evaluated BK channel ion conduction properties not only using symmetric [K+] but also substituting Na+ for K+, in order to establish whether EtOH modifies the high selectivity of the channel for K+ over Na+. Figure 5 showsi/V plots obtained in the absence and presence of 50 mM EtOH, a concentration at which channel NPo was increased about 160% over control values (fig. 2a). In the absence of EtOH, the single-channel conductance, obtained from the slope of thei/V plot in symmetric 145 mM [K+] was 240.1 ± 6.5 pS (r = 0.999), with a reversal potential near 0 mV (−0.7 mV). The reversal potential shifted in a positive direction to +28.6 mV (r = 0.998) with 100 mM [Na+]/45 mM [K+] at the intracellular side, almost identical to the theoretical value of +30.6 mV predicted by the Nernst equation for a channel highly selective for K+. Thus, mslo channels exhibit the combination of large unitary conductance and high selectivity for K+ over Na+ that is characteristic of BK channels. Figure 5 also shows that this characteristic feature was unmodified by EtOH. In the presence of the drug, the single-channel conductance in symmetric 145 mM [K+] was 235.7 ± 7.2 pS (r = 0.999), with a reversal potential of −1.3 mV. The shift in the reversal potential to +28.2 mV (r = 0.995) when 100 mM [Na+]/45 mM [K+] was used at the intracellular side of the patch was almost identical to that observed in controls, following the Nernst prediction. Therefore, EtOH-activatedmslo channels retain a typical characteristic of BK channels: the combination of large conductance and high selectivity for K+, remaining almost impermeable to Na+.

Unitary current-voltage (i/V) relationships of mslo BK channels for both symmetric 145 mM [K+] (circles) and 145 [K+]o/45 mM [K+]i (Na+ substituting for K+) (triangles), in the absence (filled figures) and presence (hollow figures) of 50 mM EtOH, from I/O patches. For composition of the solutions, please see “Methods.” In controls, the single-channel conductance (γ), obtained from the slope of thei/V plot in symmetric [K+], is 240.1 ± 6.5 pS (r = 0.999) with a reversal potential near 0 mV (−0.71 mV). The reversal potential shifted in a positive direction to +28.6 mV (r = 0.998) with 45 mM [K+]i, a value very close to the theoretical value of +30.6 mV predicted by the Nernst equation for a channel highly selective for K+. In the presence of EtOH, γ in symmetric 145 mM [K+] is 235.7 ± 7.2 pS (r = 0.999), not significantly different from that in controls, and the reversal potential is −1.31 mV. The shift in the reversal potential to +28.2 mV (r = 0.995) with 45 mM [K+]i is almost identical to that observed in controls, which indicates that EtOH does not modify the high selectivity of K+ over Na+ that is characteristic of BK channels. Individual points at each potential were obtained from all-points histograms. Plots for symmetric and asymmetric [K+] have been fitted using first- and second-order regressions, respectively. Each data point is the mean ± S.E. of at least three determinations; each determination was obtained in a different patch.
EtOH differentially modulates the voltage- and Ca++-sensitivity of the channel.
The study of EtOH’s action on cloned channels, compared to natural BK channels, has the advantage of providing a population of channels that show very defined functional characteristics—in particular, voltage/Ca++-sensitivity. Dual sensitivities to activating [Ca++]ic and transmembrane depolarizing voltage play a key role in the modulation of BK channel gating, without modifying single-channel conductance or ion selectivity (Magleby and Pallota, 1983; Latorre et al., 1989; McManus, 1991). We have demonstrated above that EtOH increases mslo channel activity by modifying channel gating without altering either single-channel conductance or the selectivity of the channel for K+. We next tested whether the drug enhances mslo channel activity by increasing the sensitivity of the channel to voltage and/or [Ca++]ic.
For a fixed intracellular [Ca++], if the voltage activation is described by a Boltzmann relationship, a plot of the natural log of NPo (or ofPo if N = 1) as a function of voltage should be linear at low values of Po(Singer and Walsh, 1987). Low values of Po were achieved by working at low [Ca++]ic (10–316 nM). In figure 6 we plot the natural log of NPo as a function of voltage, at lowPo. In this type of plot, the reciprocal of the slope (a measure of voltage sensitivity), is the potential needed to produce an e-fold change in NPo(Singer and Walsh, 1987). Figure 6 shows not only the expected increase of mslo channel NPo as the potential was set at more positive values, but also the linearity of the plot (r > 0.99) at low NPo values, which is characteristic of a Boltzmann relationship. The voltage-sensitivity of mslo channels found here was quantitatively homogenous among different channels from different patches: 18.1 ± 3.6 mV per e-fold change inNPo (n = 5). The reciprocal of the slope in the NPo-V relationship gives an effective valence (z) of 1.43 ± 0.28, calculated from 1/slope = RT/zF, where R, Tand F have their usual meaning. These values are within the range of values reported in the literature for this and other BK clones encoded by slo genes and for native BK channels (Toroet al., 1990; Butler et al., 1993; DiChiara and Reinhardt, 1995; Dopico et al., 1996a). More important, figure 6 shows that 50 mM EtOH did not modify the slope of this relationship. This representative result, in which control and EtOH values were obtained in the same patch, was confirmed in four other patches from different oocytes, the voltage-sensitivity ofmslo channels being 17.5 ± 2.2 mV pere-fold change in NPo(z = 1.47 ± 0.18) in the presence of 50 mM EtOH (not statistically different from the control value shown above; P > .9; n = 5). The lack of change in the slope after exposure to EtOH indicates that the drug does not modify the voltage-sensitivity of the channel, although the probability of the channel being open at a given potential is markedly increased. In other words, EtOH displaces the equilibrium between conducting and nonconducting states of the mslo channel without changing the voltage dependence of the gating process.

Representative plot of NPo as a function of voltage from mslo channels in the presence (hollow circles) and absence (filled circles) of 50 mM EtOH, obtained in the same I/O patch. Given a fixed intracellular [Ca++] (in this case, [Ca++]free ∼100 nM), when the voltage activation is described by a Boltzmann relationship, a plot of the natural log of NPo as a function of voltage should be linear at low values of Po. The reciprocal of the slope, a measure of voltage sensitivity, is the potential needed to produce an e-fold change inNPo: 21.5 mV (r = 0.995) and 22.1 mV (r = 0.993) in the presence and absence of EtOH, respectively. This representative result was confirmed in four other patches from different oocytes, the voltage sensitivity ofmslo channels being 17.52 ± 2.18 vs.18.07 ± 3.61 mV/e-fold change inNPo, in the presence and absence of 50 mM EtOH, respectively (not statistically significant, P > .9). Thus EtOH activates mslo channels without modifying the voltage dependence of activation.
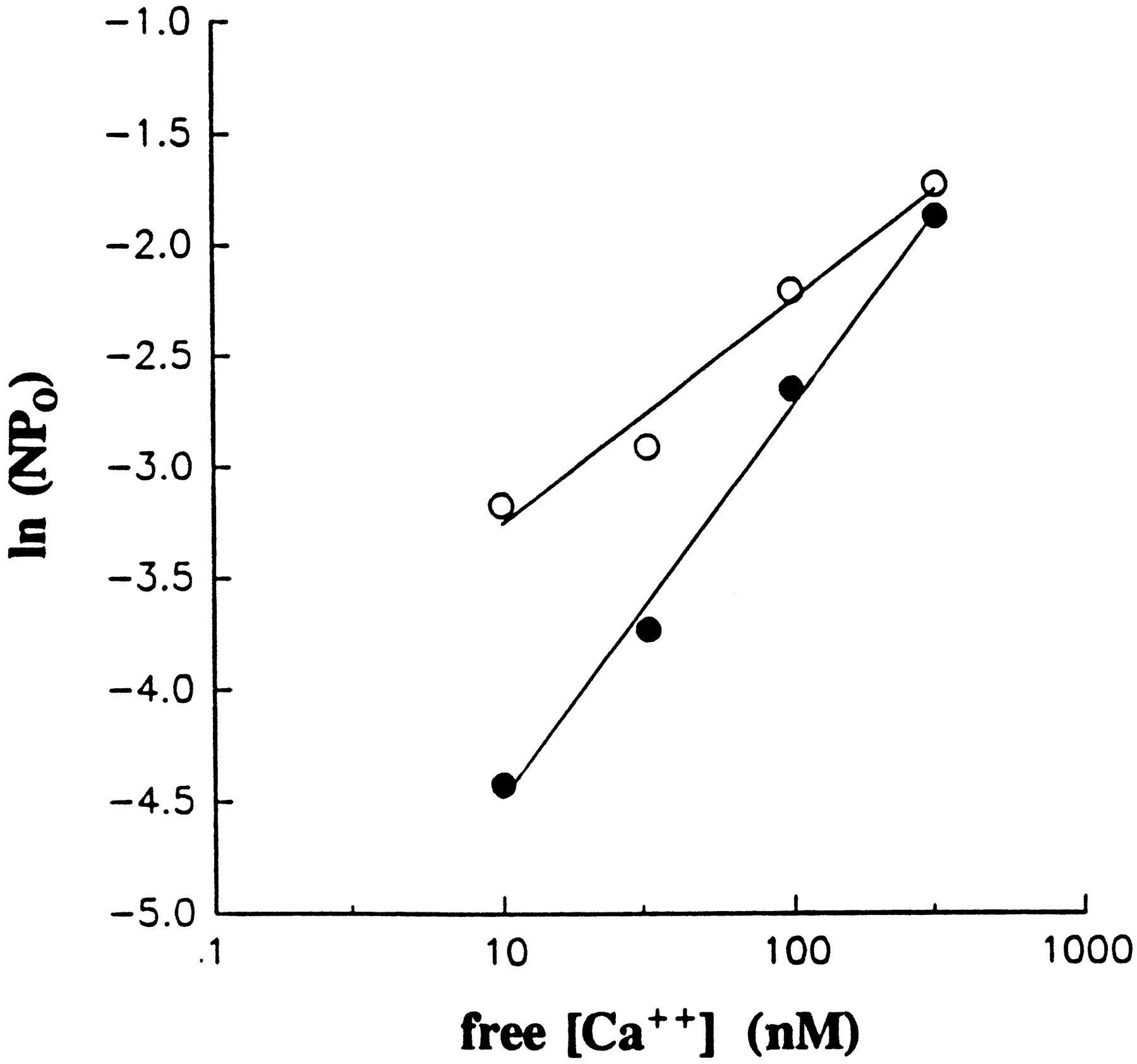
The EtOH-induced parallel shift of the voltage activation curve toward negative potentials shown in figure 6 is consistent with the possibility that the drug increases the Ca++-sensitivity ofmslo channels (Barrett et al., 1982; DiChiara and Reinhardt, 1995). To explore this possibility directly, we evaluated EtOH action on channel activity at a range of free [Ca++]ic at a fixed potential. For BK channels, given a fixed voltage, a plot of the natural log of (N)Po as a function of [Ca++]ic should be linear at low values of activity (Barrett et al., 1982). Figure7 shows a representative plot ofNPo as a function of free [Ca++]ic, at +40 mV in the presence and absence of 50 mM EtOH. The plots show that EtOH increases the apparent Ca++-sensitivity of mslo channels, because channels in the presence of the drug require less cytosolic free Ca++ to any given level of activity. However, in spite of this shift to the left, EtOH decreased the slope of theNPo-[Ca++]icrelationship: 1.75 (r = 0.997) vs. 1.04 (r = 0.977) in the absence and presence of 50 mM EtOH, respectively. Therefore, for a given [EtOH], the increase inmslo channel NPo is an inverse function of [Ca++]ic. In the case depicted in figure 7, EtOH increased channel activity to 292.9% of control values at 10 nM free [Ca++]ic, whereas the drug increased channel NPo to only 122.8% of control values at 316.2 nM free [Ca++]ic. Data similar to those shown in figure 7 were obtained in six other patches from different oocytes, with slopes of 1.82 ± 0.21 vs.0.77 ± 0.23 in the absence and presence of 50 mM EtOH, respectively (n = 7; P < .02, paired two-tailedt test). The decrease in the slope of theNPo-free [Ca++]icrelationship may be interpreted as EtOH altering the capability of the Ca++-sensing site(s) to respond to increases in [Ca++]ic. Alternatively, this result might be expected if the drug interfered with one or more Ca++-buffer equilibria, resulting in increases in nominal free Ca++ in the solutions (and, eventually, channel activation); such an effect would be more pronounced at low free [Ca++]ic and Po, with smaller effects as nominal Ca++ andPo were increased. We feel that the latter possibility is unlikely, because 1) EtOH-induced activation exhibited slow recovery (5–6 min), and 2) increasing the levels of EGTA to 20 mM while maintaining the same nominal free Ca++ concentration (100 nM) did not affect the increase in NPo by 50 mM EtOH (178% of control, n = 2). Thus, when considering the overall effects of EtOH on cellular excitability, it is important to take into account that the efficacy of EtOH in increasing BK channel activity is a function of the cytosolic Ca++concentration.

Representative plot of mslo channelNPo as a function of [Ca++]ic, at a fixed potential (+40 mV) in the presence (hollow circles) and absence (filled circles) of 50 mM EtOH, obtained in the same I/O patch. A plot of the natural log ofNPo as a function of [Ca++]free should be linear at low values ofPo. The slope, a measure of the response in channel activity upon recognition of Ca++-sensing sites to increases in [Ca++]ic, is markedly decreased by EtOH: 1.748 (r = 0.997) vs. 1.044 (r = 0.977) in the presence and absence of the drug, respectively. Data similar to those for this representative case were obtained in six other patches from different oocytes, the slopes being 1.824 ± 0.210 vs. 0.769 ± 0.225 in the absence and presence of 50 mM EtOH, respectively (n = 7; P < .02, paired two-tailed t test).
Discussion
Present results demonstrate that acute exposure to EtOH reversibly increases the activity of cloned BK (mslo) channels expressed in Xenopus oocytes after injection of the channel α subunit. Although the maximal degree of potentiation ofmslo channel activity by EtOH is slightly lower than that observed previously in the neurohypophysial BK channel, for which a maximal level of activation (400–450% of controls) occurred at a concentration of 50 mM EtOH, the EC50 found in this study (24 mM) is not only close to legal intoxicating levels in United States but also is almost identical to the EC50 obtained for activation by EtOH of the nerve terminal BK channel (22 mM). In both systems, EtOH-induced BK channel activation occurs immediately after exposure of the cytosolic side of the patch to the drug and decreases to pre-EtOH values after 5 to 6 min of washout with drug-free solution. Furthermore, in both native and cloned channels, the drug modifies similar gating processes: 50 mM EtOH drives the channel away from brief open states toward long open states and destabilizes the long closed state of the channel. In both systems, these modifications, which result in the overall increase in channel activity produced by the drug, occur without altering the channel unitary conductance. These striking qualitative and quantitative similarities support the validity of studying the action of EtOH on cloned channels encoded byslo genes expressed in Xenopus oocytes. Currently, the subunit composition and sequence(s) of the neurohypophysial BK channel are unknown. However, the similarities in the activation by EtOH of two BK channels embedded in two probably different environments (the neurohypophysial and the oocyte membrane) suggest a common site(s) for the action of EtOH on BK channels (probably the α subunit itself; see below).
The EtOH-induced increase in mslo channelNPo is entirely due to actions on channel gating; it occurs without changes in the number of functional channels (in this case, α subunits) present in the patch. Drug actions on gating are a function of EtOH concentration. At [EtOH] = 10 mM, the increase in NPo is totally explained by a decrease in tc. On the other hand, at [EtOH] ≥ 25 mM, activation results from a combination of concentration-dependent increases in to and frequency of channel openings. It is interesting to note that although the overall effect is the opposite (i.e., decrease of channel activity), EtOH’s actions on l-type Ca++ channel gating in neurohypophysial terminals show a similar dependence on drug concentration: EtOH-induced changes into contribute to the overall decrease inNPo only at [EtOH] ≥ 25 mM. In this system, EtOH’s effects on gating are consistent with the interaction of a single drug molecule with a single site (Wang et al., 1994). Although it is possible that the concentration-dependent differential effects of EtOH on mslo channels reflect the interaction of the drug with multiple sites of different affinities, the fact that we found a Hill coefficient not significantly different from 1 for EtOH activation (fig. 2B) indicates that if multiple sites are involved in the functional interaction between EtOH and the channel, this interaction occurs without cooperativity (either positive or negative) among these sites.
The distribution of open and closed times shown in figure 4 indicates that the channel in the absence of the drug exhibits at least two open and two closed states, which is consistent with previous models proposed for BK channels (Barrett et al.,1982; Toro et al., 1990; DiChiara and Reinhardt, 1995). Because of the filtering (1.5 kHz) used to construct idealized records, we worked in the millisecond scale, so the number of open and closed states proposed here may be underestimated. Whether EtOH modifies briefer and/or longer events or switches the channel into a different gating “mode” (a phenomenon that usually occurs in the range of tens of seconds to minutes) remains to be determined. Despite the fact that no additional components in the distribution of either open or closed times were introduced by EtOH, it is possible that a unique EtOH-BK channel dwell time(s) exists but, as a result of fast dissociation, cannot be resolved.
EtOH increased mslo channel activity by shifting the ln(NPo)-[Ca++]icrelationship to the left while decreasing its slope. The finding that potentiation of mslo activity by EtOH decreased as Ca++ levels were increased could be explained by an elevated Po in the presence of high [Ca++]ic, which would leave little room for further activation by EtOH. However, even at the highest [Ca++]ic used, NPovalues were low, as confirmed by the linearity of the plot of ln(NPo) vs.[Ca++]ic (fig. 7). The decrease in slope indicates an altered Ca++ dependence of channel gating in the presence of EtOH. In the absence of the drug, an approximate two power relationship (limiting slope = 1.82) was observed between free [Ca++]ic and channelNPo, which suggests that channel activation requires either the strong cooperative binding of two Ca++ions or the weaker cooperativity of more than two Ca++ions. These data are consistent with previous findings obtained in describing the Ca++ dependence of channel gating after injection of dslo or hslo α subunits intoXenopus oocytes (DiChiara and Reinhardt, 1995). Mechanistically, these results can be interpreted as meaning that the binding of two Ca++ ions is required to open themslo channel or that, alternatively, only one Ca++ is required to open the channel, which, once opened, dwells in open states longer as [Ca++]ic is increased. On the other hand, in the presence of 50 mM EtOH, the limiting slope in the free [Ca++]ic-NPorelationship was well below 1 (0.77). Because EtOH does not change the number of α subunits (see “Results”), this finding indicates that the drug introduces apparent negative cooperativity in the interaction between Ca++ ions in the cytosolic side of the patch and Ca++-sensing site(s).
One possibility to explain the decreasing augmentation of channelNPo by EtOH with increasing [Ca++]ic is that EtOH behaves functionally as a “partial” agonist of the mslo channel, for which [Ca++]ic is a “full” agonist. Two observations are consistent with this hypothesis: 1) EtOH “antagonism” of Ca++ on mslo channels (i.e., the decrease in the slope of theNPo-[Ca++]icrelationship produced by the drug) and EtOH “agonism” (i.e., mslo channel activation) occur at the same EtOH concentration (50 mM), a requirement for partial agonists (Kenakin, 1987); 2) Under conditions in which the basal activation of the system is kept low, the putative partial agonist should function as a competitive antagonist (Kenakin, 1987). To test this proposition, we evaluated the action of 50 mM EtOH on mslo channel NPo in I/O patches exposed to a high nominal free [Ca++]ic while keepingPo values low by recording at negative potentials (−80 mV). Under these recordings conditions, 50 mM EtOH failed to increase channel activity: NPo in the presence of the drug was 103.4 ± 14.4% of control (pre-EtOH, obtained in the same patch) values (n = 4). Whether this postulated partial agonism is a consequence of the interaction of EtOH with a shared site at the receptor or reflects interactions with two (or more) discrete sites (one or more that explain activation and another or others that explain the decreased response in channel activity to increases in [Ca++]ic), cannot be determined from functional studies such as those described here. In the absence of binding studies, a variety of possibilities (not mutually exclusive) may be considered in an effort to explain this action of EtOH: an actual decrease in the number of Ca++ binding sites, introduction of negative cooperativity in the actual binding of Ca++ ions to a conserved number of Ca++recognition sites, and alteration in the Ca++ dependence of channel gating per se (that is, EtOH modifies steps subsequent to Ca++ interaction with recognition sites).
Although a competitive interaction between EtOH and Ca++has been demonstrated to underlie EtOH’s inhibition of Ca++-dependent ATPase activity in erythrocytes (IC50 ∼ 3 M) (Lopez and Kosk-Kosicka, 1995), our results are the first to show a functional antagonism between Ca++at physiological intracellular levels (about 10–316 nM) and EtOH at pharmacologically relevant concentrations. EtOH activation of BK channels would be most evident in a resting neuron and/or when [Ca++]ic is limited by intracellular storage mechanisms. EtOH is known to inhibit Ca++ entry by blocking voltage-dependent Ca++ channels (Wang et al.,1994; Mullikin-Kilpatrick et al., 1995). EtOH-blockade of Ca++ channels will maintain [Ca++]ic near resting levels, maximizing the activation of BK channels by the drug. Furthermore, BK channels exposed to EtOH retain their high permeability and selectivity for K+ over Na+ (fig. 5), and these properties allow BK channel activity to contribute effectively to membrane repolarization or hyperpolarization. Thus, EtOH activation of BK channels and blockade of voltage-dependent Ca++ channels would act synergistically to result in repolarization or hyperpolarization of the cellular membrane and, eventually, decreased excitability.
EtOH-induced activation of mslo channels occurs in I/O patches when highly buffered Ca++ solutions are used at both sides of the patch, in the absence of cyclic nucleotides or other “regenerating” systems. This result indicates that EtOH-inducedmslo channel activation is not secondary to EtOH’s modulation of freely diffusible second messengers (including cytosolic Ca++) but, rather, probably results from a functional interaction between EtOH and the channel or a closely associated entity in the membrane patch. It is currently postulated that native BK channels from mammalian tissues consist of two structurally distinct subunits, termed α and β (McManus et al., 1995). It is unlikely that the β subunit is the subunit mainly responsible in making the BK channel activatable by EtOH. First, EtOH activation was observed when BK channel activity was evoked by injection of the channel α subunit into Xenopus oocytes, an expression system where no endogenous BK β subunits have to our knowledge been observed. Second, preliminary data obtained in I/O patches from oocytes injected with the α subunit of the dslo BK channel also show a reversible, equipotent increase in NPo by EtOH (Dopico et al., 1996b), and BK β subunits do not seem to regulate dslo channel activity (Meera et al.,1996). Third, bslo (Moss et al., 1996) andmslo α subunits are about 98% homologous, yet onlymslo is potentiated by EtOH when they are expressed in the oocyte. Therefore, it is unlikely that an endogenous β subunit is important for EtOH’s action on BK channels (Dopico and Treistman, 1996).
The use of mslo α allows us to speculate on which specific regions in this subunit EtOH functionally interacts with.Mslo electrophysiological properties have been related to functional domains (termed “core” and “tail”) in the channel protein: 1) both the core (S1–S8) and the tail (S9–S10) are required for function; 2) the core determines ion selectivity and permeability (properties linked to the pore-lining sequence, a region between S5 and S6 termed H5), and voltage dependence of gating (a region defined by S4); 3) the tail contributes to determining apparent Ca++-sensitivity (Wei et al., 1994). The lack of change in the voltage dependence of mslo channel gating in the presence of acute EtOH shown here indicates that EtOH does not functionally interact with the voltage sensor. In addition, the unmodified K+ permeability and selectivity ofmslo channels in the presence of EtOH indicate that the drug does not alter two key properties associated with the H5 region. Together, these data indicate that acute EtOH does not modify several of the major electrophysiological characteristics of mslochannels determined by the core. However, an interaction between EtOH and the core cannot be totally ruled out because the drug increased the channel to, which is at least partially determined by this domain (Wei et al., 1994). On the other hand, the EtOH-induced alteration in the response of the Ca++-sensing site(s) to increases in [Ca++]ic shown here with mslochannels suggests a functional interaction of the drug with Ca++ sensing sites of the channel α subunit, possibly with the S9 to S10 region (tail domain).
Although the simplest explanation for our data is a direct interaction of EtOH with selective regions of the α subunit, we cannot ignore the possibility that EtOH is affecting membrane-bound modulators of BK channel activity, such as G proteins (Scornick et al., 1993;Mullikin-Kilpatrick et al., 1995), protein kinase or phosphatase activities (Reinhardt et al., 1991), redox status (Loguercio et al., 1993; Lee et al.,1994), lipids (Bolotina et al., 1989; Kirber et al., 1992; Abadji et al., 1994), or water-lipid or lipid-protein interfaces (Barry and Gawrich, 1994). The recording ofmslo channel activity in I/O patches long after patch excision (>15 min) in the absence of nucleotides in the solutions facing both sides of the patch makes some of these elements unlikely mediators of EtOH action. However, it is intriguing that washout of EtOH’s effects in both native and cloned channels was significantly slower than the onset of these effects, requiring ≥5 min, a result that suggests the need for further experiments to probe the basis for this observation.
Acknowledgments
Authors wish to thank Drs. José R. Lemos and Hélène Widmer for their critical reading of the manuscript, Dr. Joshua J. Singer for helpful discussion, and Andrew Wilson and Lynda Zorn for their excellent technical assistance.
Footnotes
-
Send reprint requests to: Steven N. Treistman, Ph.D., Department of Pharmacology and Molecular Toxicology, University of Massachusetts Medical School, 55 Lake Avenue North, Worcester, MA 01655.
-
↵1 This work was supported by grants from the Alcoholic Beverage Medical Research Foundation (A.M.D.) and National Institutes of Health AA08003 (S.N.T.).
-
↵2 Present affiliation: Department of Physiological Sciences, University of California at Los Angeles, CA 90095.
- Abbreviations:
- EtOH
- ethanol
- BK
- large conductance, Ca++-activated K+
- CAK
- Ca++-dependent K+
- [Ca++]ic
- intracellular calcium
- EGTA
- 10 ethylene glycol-bis(β-aminoethyl ether) N,N,N′,N′-tetraacetic acid
- I/O
- inside-out
- N
- number of functional channels present in the patch membrane
- Po
- probability that a particular channel is open
- T
- period of recording
- to
- channel mean open time
- #o
- number of channel openings during T
- tc
- channel mean closed time
- Pc
- probability that a particular channel is closed
- #c
- number of channel closures during T
- γ
- single-channel conductance
- i
- unitary current amplitude
- Received April 10, 1997.
- Accepted September 15, 1997.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}