


Abstract
The role of M2 and M3 receptors in the contractile and phosphoinositide responses elicited to oxotremorine-M was investigated in the guinea pig colon. Under standard conditions, both the contractile and phosphoinositide responses were insensitive to pertussis toxin and irreversibly antagonized by alkylation of M3 receptors withN-(2-chloroethyl)-4-piperidinyl diphenylacetate. After treatment with N-(2-chloroethyl)-4-piperidinyl diphenylacetate, the remaining contractile response was sensitive to pertussis toxin and weakly antagonized by the M2- and M4-selective antagonist AF-DX 116. In contrast, the residual phosphoinositide response was unaffected by pertussis toxin. The pertussis toxin sensitivity of the remaining contractile response suggests that the M2 receptor is mediating the contraction, whereas its weak antagonism by AF-DX 116 suggests that an alternate muscarinic subtype mediates the response. To explain this enigma, we investigated a mathematical model for receptor action based on an interaction between two receptor subtypes (M2 and M3). This model predicts that a response mediated by both the M2 and M3 receptor can be pertussis toxin sensitive yet exhibit an antagonistic profile indicative of an M3 response.
M2and M3 muscarinic receptor subtypes are abundantly expressed in smooth muscle throughout the gastrointestinal tract (Eglen et al., 1996; Ehlert et al., 1997). Radioligand binding (Giraldo et al., 1988; Michel and Whiting, 1990), mRNA hybridization (Maeda et al., 1988), and immunoprecipitation studies (Wall et al., 1991) have shown that the M2 receptor is the more abundant of the two receptors in the intestine (Ehlert et al., 1997). Interestingly, a large body of pharmacological evidence shows that the less abundant M3 receptor mediates contraction of gastrointestinal smooth muscle under standard conditions (i.e., no other contractile or relaxant agents present). The M3 receptor is known to stimulate phospholipase Cβ in a variety of cell types, including smooth muscle. Presumably, this mechanism mobilizes calcium and initiates contraction in smooth muscle.
The M2 receptor is known to mediate a pertussis toxin-sensitive inhibition of adenylyl cyclase activity in the guinea pig ileum and colon (Candell et al., 1990; Zhang and Buxton, 1991;Thomas and Ehlert, 1994). Muscarinic agonists have been shown in both ileum and colon to oppose the increase in cAMP elicited by forskolin, isoproterenol, and a variety of other compounds that stimulate cAMP accumulation (Griffin and Ehlert, 1992; Ostrom and Ehlert, 1997; Sawyer and Ehlert, 1998). The M2 receptor also elicits an indirect contraction in both the guinea pig ileum and colon by preventing the relaxant effects of forskolin and isoproterenol on histamine-induced contraction (Thomas et al., 1993; Thomas and Ehlert, 1994; Ostrom and Ehlert, 1997; Sawyer and Ehlert, 1998). The M2-mediated indirect contraction is pertussis toxin sensitive (Sawyer and Ehlert, 1998), unlike the standard M3-mediated contraction.
Extensive alkylation of M3 receptors by the selective irreversible antagonistN-(2-chloroethyl)-4-piperidinyl diphenylacetate (4-DAMP mustard) (Thomas et al., 1993) renders the contractile response of the guinea pig colon sensitive to pertussis toxin treatment (Sawyer and Ehlert, 1998). This sensitivity suggests that the M2 receptor may directly participate in the contractile response of the colon after most of the M3 receptors have been inactivated with 4-DAMP mustard. When expressed in high abundance, M2receptors have been shown to mediate a weak, pertussis toxin-sensitive, phosphoinositide response by coupling to Gi(Ashkenazi et al., 1987; Lai et al., 1991; Dell’Acqua et al., 1993). Such a mechanism would be difficult to detect in the face of a large M3 phosphoinositide response via Gq. Thus, it is important to consider whether M2 receptors in the colon are capable of eliciting contraction through the phosphoinositide pathway after most of the M3 phosphoinositide response has been inactivated.
In the present report, we investigated this hypothesis and have shown that although the muscarinic contractile response is extremely sensitive to pertussis toxin after 4-DAMP mustard treatment, the phosphoinositide response is not. Thus, our results provide no evidence for the direct coupling of M2 receptors to phosphoinositide hydrolysis. Instead, our results are consistent with a mathematical model in which the M2 receptor is capable of eliciting contraction when there is a low level of M3 receptor activation. This model accounts for the finding that the muscarinic contractile response after 4-DAMP mustard treatment is sensitive to pertussis toxin yet nevertheless relatively insensitive to the M2-selective antagonist AF-DX 116.
Experimental Procedures
Contractile Measurements.
Male Hartley guinea pigs (300–400 g) were asphyxiated with CO2 followed immediately by exsanguination, and segments of colon (1–2 cm) were harvested 1 cm from the cecum. Each segment was rapidly cleaned with Krebs-Ringer-bicarbonate (KRB; 124 mM NaCl, 5 mM KCl, 1.3 mM MgCl2, 26 mM NaHCO3, 1.2 mM KH2PO4, 1.8 mM CaCl2, 10 mM glucose) buffer to remove its contents. The segments were connected to a force transducer and mounted longitudinally in an organ bath containing 50 ml of KRB buffer at 37°C gassed with O2/CO2(19:1). The colonic segments were allowed to equilibrate for 40 min at a resting tension equivalent to a load of 1.5g (optimal pretension was determined after constructing a pretension versus force of contraction curve) before measurement of isometric contractions with a force transducer and polygraph. A test dose of the highly efficacious muscarinic agonist oxotremorine-M was then added to each bath. Once each tissue reached a sustained contraction, each bath was washed with KRB buffer and allowed to incubate for 5 min before the addition of two more test doses. These three test doses were used to ensure reproducibility of the preparations. Segments of colon that did not contract to at least 60% of that elicited by 100 mM KCl were discarded. After the last 5-min incubation, the KRB buffer was replaced with 50 ml of Ca2+-free KRB buffer (124 mM NaCl, 5 mM KCl, 1.3 mM MgCl2, 26 mM NaHCO3, 1.2 mM KH2PO4, 1 mM EGTA, 10 mM glucose). The colon was incubated in Ca2+-free medium for 10 min to inhibit myogenic contraction and cause full relaxation. During this period, a resting tension of 1.5gwas maintained. Subsequently, the Ca2+-free KRB buffer was replaced with 50 ml of K+-deficient KRB buffer (124 mM NaCl, 1.3 mM MgCl2, 26 mM NaHCO3, 1.2 mM KH2PO4, 1.8 mM CaCl2, 10 mM glucose) to inhibit spontaneous contractions. After the addition of the K+-deficient KRB buffer, a large contraction was observed that declined to resting tension within 7 to 10 min. Cumulative agonist concentration-response curves were measured by adding 9 to 18 geometrically spaced (0.33 log unit) concentrations of oxotremorine-M to each of the organ baths. The EC50 value was determined from this curve as described below. The K+-deficient KRB buffer was washed from the bath, and 50 ml of KRB buffer was added. Colonic segments were allowed to incubate for 30 min before any further measurements were made. The above procedure (excluding three test doses) was repeated before each EC50 measurement made with the same tissue.
Contractile Experiments Using 4-DAMP Mustard-Treated Colonic Segments.
Some colonic segments were incubated with 40 nM concentration of the aziridinium ion of 4-DAMP mustard for 1 h in the presence of 1.0 μM [[2-[(diethylamino)methyl]-1-piperidinyl]acetyl]-5,11-dihydro-6H-pyrido[2,3b][1,4]benzodiazepine-6-one (AF-DX 116) to selectively alkylate M3 muscarinic receptors (Thomas et al., 1993). The colon segments were washed thoroughly to remove AF-DX 116 and any unreacted 4-DAMP mustard before measurement of concentration-response curves as described above. Some tissues were washed with KRB buffer, and the aziridinium ion of 4-DAMP mustard (40 nM) and AF-DX 116 (1.0 μM) were added again for an additional hour (i.e., total incubation time of 2 h) before measurement of concentration-response curves. In other experiments, colonic segments were exposed to 4-DAMP mustard for a total of 3 or 4 h using the same washing procedure described above at the end of each hour of exposure. In all experiments, 4-DAMP mustard was first converted to its aziridinium ion by incubation for 30 min at 37°C in 10 mM NaKPO4, pH 7.4, as previously described (Thomas et al., 1992).
Phosphoinositide Hydrolysis Assay.
Male guinea pigs (300–400 g) were sacrificed as described above, and an approximately 16-cm segment of colon was rapidly harvested 1 cm distal the cecum. The colon was placed in ice-cold KRB gassed with O2/CO2 (19:1) and cleaned to remove the contents. The colonic segment was cut longitudinally, and the mucosa was removed using a microscope slide. The segment was cross-chopped at 350 μM using a McIlwain tissue chopper. The resulting colonic slices were immediately suspended in 37°C KRB gassed with O2/CO2 (19:1) and then washed with KRB (37°C) three times. The slices were allowed to incubate in KRB at 37°C gassed with O2/CO2 (19:1) for 30 min. In some experiments, segments of whole colon were first incubated with AF-DX 116 and 4-DAMP mustard for 2 h before slicing of the tissue. For these incubations, approximately 10 cm of colon was incubated in a final volume of 330 ml of KRB buffer containing AF-DX 116 (1 μM) and 4-DAMP mustard (40 nM). The tissue was washed extensively to remove AF-DX 116 and any unreacted 4-DAMP mustard, and the tissue was sliced as described above. To 10 ml of packed colonic slices, 8 ml of KRB buffer containing 100 μCi ofmyo-[3H]inositol was added, and the mixture was incubated for 90 min. The slices were gassed with O2/CO2 (19:1) every 30 min. After this incubation, the slices were washed with KRB (37°C) three times. The slices were allowed to settle before pipetting 100-μl aliquots of slices into tubes with 0.35 ml of KRB (37°C) gassed with O2/CO2 (19:1) containing 5 mM LiCl and various concentrations of oxotremorine-M without or with antagonists in concentrations identified in Results. The slices were incubated for 30 min in these tubes at 37°C. The incubation was stopped by adding 1.13 ml of chloroform/methanol (1:2, v/v). The accumulated inositol phosphates were extracted and separated according to the method of Berridge et al. (1982), as described briefly. Chloroform (0.37 ml) and water (0.37 ml) were added to the tubes to separate the organic and aqueous phases. After centrifugation, 1 ml of the aqueous phase was pipetted into tubes to which 2 ml of water was added. These tubes were centrifuged to sediment any chloroform, and 2.9 ml of the aqueous phase was applied to a Dowex AG 1-X8 (100–200 mesh, 1 ml) anion exchange column. The column was washed three times with 5 ml of water each to remove [3H]inositol and discarded. [3H]Inositol phosphates were eluted from the column with 2.5 ml of 1 M ammonium formate/0.1 M formic acid solution. The elutant was then counted to determine the amount of radioactivity using a scintillation counter. In some experiments and in all experiments in which pertussis toxin-treated colonic slices were used, 200 μl of the organic phase was removed, dried, and then counted to determine the amount of [3H]inositol incorporated into phospholipids. In these experiments, the amount of [3H]inositol phosphates formed is calculated as a percentage of the total amount of radioactivity in the organic phase plus the inositol phosphate fraction. This was done to determine whether there were differences in the uptake of [3H]inositol in tissues treated with pertussis toxin compared with tissues that were not treated with pertussis toxin.
In Vivo Pertussis Toxin Treatment.
Male Hartley guinea pigs (300–400 g) were injected i.p. with pertussis toxin (50–100 μg/kg b.wt.) 3 days before being euthanized for the in vitro experiments. For the contractile assays, a dose of 100 μg/kg b.wt. pertussis toxin was used, whereas in the phosphoinositide assays, a dose of 50 μg/kg b.wt. pertussis toxin was used. Initial experiments showed that these two doses yielded similar results in the phosphoinositide assay.
Data Analysis.
The concentration of oxotremorine-M eliciting half-maximal contraction (EC50) was estimated by nonlinear regression analysis according to an increasing logistic equation as described previously (Candell et al., 1990).
The dissociation constant (KB) of the antagonists AF-DX 116, pirenzepine, andpara-fluoro-hexahydro-sila-difenidol (p-F-HHSiD) were estimated from the shift that the antagonists caused in the oxotremorine-M concentration-response curve:KB = [B]/(CR − 1). In this equation, [B] represents the concentration of the antagonist, andCR corresponds to the concentration ratio (the ratio of the EC50 value of oxotremorine-M measured in the presence of the antagonist divided by that measured in the absence of the antagonist).
The amount of receptor inactivation and the pKA value were estimated using a modification of the method of Furchgott (1966) as described previously (Ehlert, 1987). Briefly, equieffective doses of oxotremorine-M were obtained from the concentration-response curves before and after treatment with 4-DAMP mustard. These estimates were fitted to the following equation by nonlinear regression analysis:
Significance values (p values) were calculated by using the paired t test and are reported where appropriate.
Mathematical Modeling.
In this investigation, we examined two mathematical models (model I and model II) for an interaction between two receptor subtypes (M2 and M3). In model I, activation of the M3 receptor by itself causes contraction. The M2 receptor has no effect by itself but can greatly potentiate the contraction elicited by the M3 receptor when activated. In model II, occupation of the M3 receptor initiates the activation of two parallel signaling pathways. One pathway causes a direct contraction and is referred to as component A. The other pathway has no effect by itself, but when activated in the presence of occupied M2 receptor, a contraction ensues (component B). The M2 receptor is incapable of eliciting contractions by itself. Component B of model II is similar to model I in that it represents an M2/M3 interaction. However, the contraction elicited by component B is conditional, occurring only when both M2 and M3receptors are activated. The mathematical derivation of these models is described in and illustrated in Fig.1.

Schematic representation of two models for interaction between M2 and M3 receptors. Solid lines indicate contractile signals. Dashed lines indicate silent signals that interact with another signal to elicit contraction.
Materials.
Islet-activating protein (pertussis toxin) was obtained from LIST Biological Laboratories (Campbell, CA).myo-[3H]Inositol was from NEN (Boston, MA). AF-DX 116 was from Boehringer Ingelheim Pharmaceuticals (Ridgefield, CT). Pirenzepine, p-F-HHSiD, and oxotremorine-M were from Research Biochemicals Inc. (Natick, MA). 4-DAMP mustard was synthesized as described previously (Thomas et al., 1992). All remaining drugs and chemicals were from Sigma Chemical Co. (St. Louis, MO).
Results
Effect of 4-DAMP Mustard Treatment on Contraction Elicited by Oxotremorine-M Under Standard Conditions.
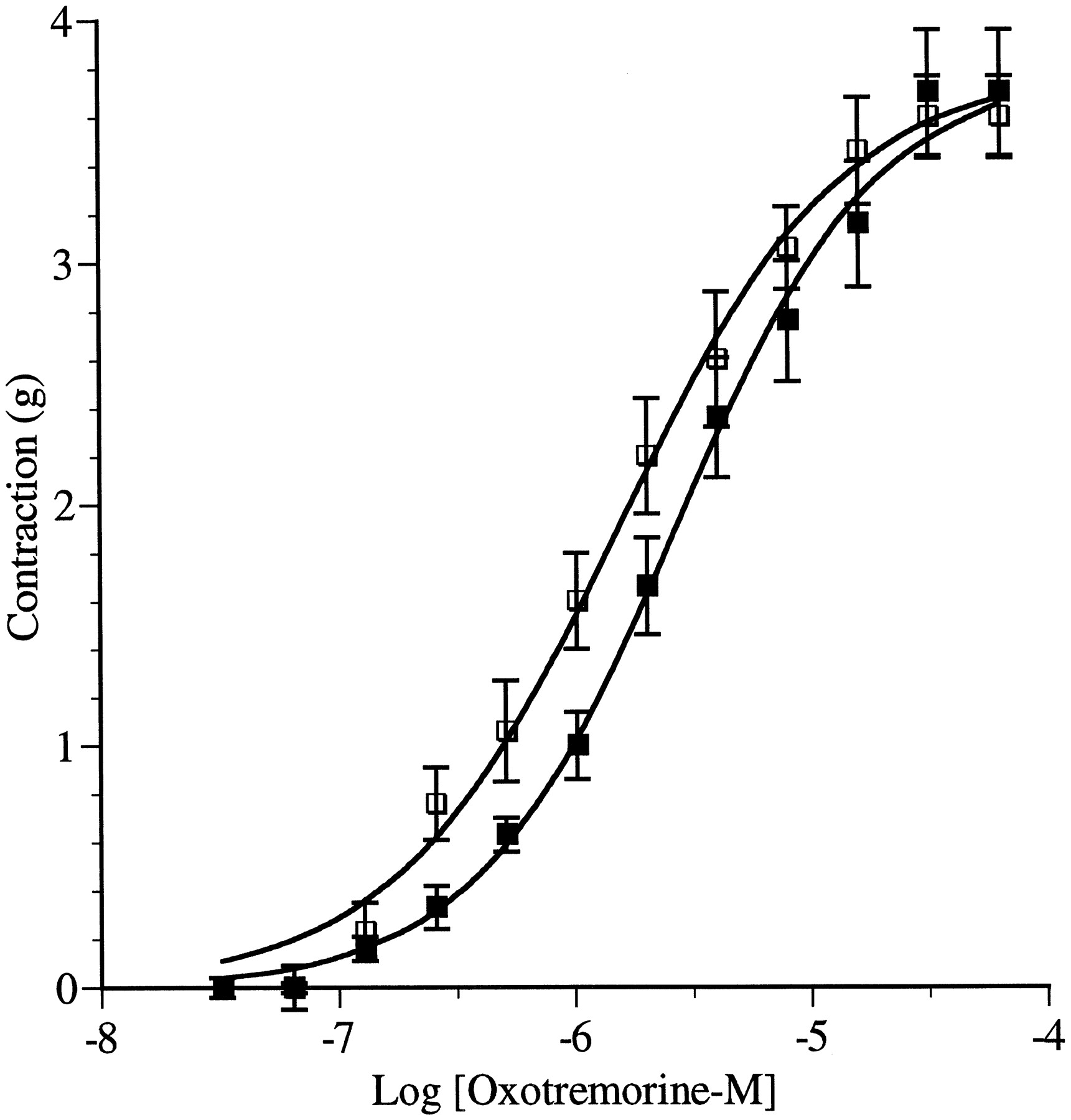
To investigate the possible contribution of the M2 receptor to the contractile response of the guinea pig colon, we treated colonic segments with 4-DAMP mustard to inactivate M3receptors. The isolated colon was incubated with the aziridinium ion of 4-DAMP mustard (40 nM) in the presence of AF-DX 116 (1 μM) for 1, 2, 3, or 4 h and then washed extensively. AF-DX 116 is used to protect M2 receptors from alkylation by 4-DAMP mustard. In the presence of AF-DX 116, the aziridinium ion of 4-DAMP mustard has been shown to be an irreversible, muscarinic antagonist that alkylates M3 receptors selectively over M2 receptors (Thomas et al., 1993). Incubation of the isolated colon with 4-DAMP mustard for 1, 2, 3, and 4 h caused a progressive shift to the right and decrease in the maximum of the concentration-response curve to oxotremorine-M (Fig.2 and Table1).

Effects of 4-DAMP mustard treatment on contractile response to increasing concentrations of oxotremorine-M under standard conditions. Contractile measurements were made in tissues before (▪) and following 4-DAMP mustard treatment (40 nM) for 1 h (♦), 2 h (▴), 3 h (●), and 4 h (▾). Each data point represents mean ± S.E.M. of four to eight experiments.
Effects of 4-DAMP mustard and pertussis toxin on the contractile response to oxotremorine-M
The effect of 4-DAMP mustard at 1, 2, 3, and 4 h corresponded to receptor inactivation values of 93.7%, 97.6%, 98.8%, and 98.7% (Table 1), respectively, as estimated according to the method of Furchgott (see Experimental Procedures) assuming that a single receptor type mediates the contractile response (Table 1). This large inhibitory effect of 4-DAMP mustard is consistent with the postulate that the M3 receptor mediates the contractile response under standard conditions. It also appears that the effect of 4-DAMP mustard reaches a limit of approximately 98% receptor inactivation after 2 h of treatment, with little additional alkylation at 4 h of treatment. These results suggest that some of the M3 receptors are inaccessible to 4-DAMP mustard or that a 4-DAMP mustard-insensitive receptor (i.e., M2) contributes to the contractile response after lengthy treatment with 4-DAMP mustard.
Effect of Pertussis Toxin on Contractile Response to Oxotremorine-M.
We examined the effect of pertussis toxin on the residual contractile response that persisted after 1- and 4-h treatment with 4-DAMP mustard to determine whether the M2receptor is contributing to the remaining contractile response. Pertussis toxin treatment had no inhibitory effect on the contractile response to oxotremorine-M when measured under standard conditions (Fig. 3). In fact, a small potentiation in contraction was observed at high concentrations of oxotremorine-M. After 1- and 4-h 4-DAMP mustard treatment, however, pertussis toxin caused a significant 25.4-fold (p = 3.1 × 10−4) and 72.5-fold (p = 8.5 × 10−6) increase in the EC50 value of oxotremorine-M, respectively (Fig.3, A and B). The sensitivity of the residual contraction to pertussis toxin suggests that the M2 receptor participates in the contractile response, when a substantial portion of M3 receptors are alkylated by 4-DAMP mustard. This participation by the M2 receptor can explain why the effects of 4-DAMP mustard appear to reach a limit (see prior discussion of Fig. 2).

Effects of pertussis toxin treatment (▪, ▴) on contractile response to oxotremorine-M in control (■, ▪) and 4-DAMP mustard-treated tissues (▵, ▴). Contractile responses were measured in 1-h (A) and 4-h (B) 4-DAMP mustard-treated tissue. Each data point represents mean ± S.E.M. of five to six experiments.
Effect of AF-DX 116 on Contractile Response After 4-DAMP Mustard Treatment.
If the M2 receptor is contributing to the contractile response after 4-DAMP mustard treatment, the remaining response may be antagonized by AF-DX 116 (M2- and M4-selective antagonist) in a manner consistent with an M2-mediated response. Isolated colonic segments were treated with 4-DAMP mustard (40 nM) in the presence of AF-DX 116 (1 μM) for 4 h and then washed extensively as described inExperimental Procedures. Treatment with 4-DAMP mustard caused a 16.3-fold increase in the EC50 value of oxotremorine-M (control EC50 = 94 nM) and a 60% reduction in the maximal contractile response (controlEmax = 9.5g). This suppression in maximal response was not as great as that observed in similar experiments shown in Figs. 2 and 3, which we attribute to experimental variation. The contractile response to oxotremorine-M was determined in the absence and presence of 1 μM AF-DX 116 after 4-h 4-DAMP mustard treatment (Fig. 4). AF-DX 116 caused a significant 1.8-fold increase (p = 3.9 × 10−3) in the EC50 value of the contractile response elicited to oxotremorine-M. The pKB value for AF-DX 116 estimated from this rightward shift (see Experimental Procedures) was 5.9 ± 0.11, which is in close agreement with the binding affinity (pKD = 6.10 ± 0.06) of AF-DX 116 at the human M3 receptor transfected into Chinese hamster ovary cells (Esqueda et al., 1996). Thus, the contractile response after 4-h 4-DAMP mustard treatment is enigmatic; it is M2-like in its sensitivity to pertussis toxin, yet it is M3-like in its weak antagonism by AF-DX 116. AF-DX 116 (1.0 μM) also weakly antagonized the contractile response to oxotremorine-M measured before 4-DAMP mustard treatment (pKB = 6.07; see Table 4).

Effects of AF-DX 116 (1 μM) on contractile response to oxotremorine-M in 4-h 4-DAMP mustard-treated tissues. Contractile measurements were made in 4-DAMP mustard-treated colon in the absence (■) and presence (▪) of AF-DX 116 (1.0 μM). Each data point represents mean ± S.E.M. of three experiments.
pKB values for antagonism of oxotremorine-M-induced phosphoinositide hydrolysis in guinea pig colon by AF-DX 116, pirenzepine, and p-F-HHSiD compared with pKB obtained from contractile studies in the guinea pig colon and binding affinities (pKD) at the human M2 and M3 receptors transfected into Chinese hamster ovary cells
Mathematical Modeling.
To explain the behavior of the muscarinic contractile response in the guinea pig colon after 4-DAMP mustard treatment, we investigated mathematical models for an interaction between two receptor subtypes (M2 and M3). We used a strategy based on the operational model of Black and Leff (1983) to calculate the theoretical concentration-response curves. We also calculated the effects of AF-DX 116 and pertussis toxin treatment on the theoretical responses. Our methods are described in .
The first model (model I) is based on the assumption that oxotremorine-M elicits a contractile response through an interaction between M2 and M3receptors. Activation of the M3 receptor causes smooth muscle contraction, whereas activation of the M2 receptor has no effect by itself but greatly potentiates the contraction elicited by the M3receptor. Figure 5 shows the results of these calculations. In this model, the affinity of the agonist is the same for both the M2 and M3receptor; however, the sensitivities of the signals elicited by the two receptors are varied, allowing us to modulate the contribution of either receptor to the response (see ). We therefore describe three cases within model I: in case 1, the M2 signal is less sensitive than the M3 signal (Fig. 5A); in case 2, the sensitivities of both the M2 and M3receptor are the same (Fig. 5B); and in case 3, the M3 signal is less sensitive than the M2 signal (Fig. 5C). The effects of progressive M3 receptor inactivation on this model are shown in Fig. 5. The closed symbols show a response mediated by the M2/M3 interaction, whereas the open symbols show the response mediated solely by the M3 receptor.

Simulation of a response mediated through interaction of an agonist with two receptors (i.e., M2 and M3 receptors; model I). Response mediated by M2/M3 interaction (filled symbols) was compared with a response mediated solely by M3 receptor (open symbols). Curves are shown before (▪, ■) and after 90% (●, ○), 99% (▴, ▵), and 99.8% (♦, ⋄) M3 receptor inactivation. A, response of M2 receptor was less sensitive than that of M3 receptor (KM2 = 0.1; KM3 = 0.01). B, response of M2 and M3 receptors were equisensitive (KM2 = 0.01; KM3= 0.01). C, response of M3 receptor was less sensitive than that of M2 receptor (KM2 = 0.001; KM3 = 0.01). Theoretical points for M3 response and M2/M3 interaction were calculated using eqs. 7 and 8, respectively, as described in the.
When the M2 signal is less sensitive to agonist than the M3 signal (Fig. 5A), the M2 receptor exhibits no potentiating effect on the M3 response before receptor inactivation. However, with progressive inactivation of M3receptors, the potentiating effect of the M2receptor becomes apparent (Fig. 5A and Table2). Because pertussis toxin eliminates signaling through the M2 receptor, the responses mediated solely by the M3 receptor can be likened to the consequence of pertussis toxin treatment on the M2/M3 interaction. Thus, pertussis toxin treatment has no effect before receptor inactivation but greatly inhibits the response measured after M3 receptor inactivation (Fig. 5A and Table 2).
Effects of M3 receptor inactivation, AF-DX 116 (1 μM), and pertussis toxin on the response predicted by model I
Cases 2 (Fig. 5B) and 3 (Fig. 5C) yield calculated responses that are inhibited by pertussis toxin treatment before and after M3 receptor inactivation. These calculated responses were inconsistent with the contractile response observed in the guinea pig colon. The results are summarized in Table 2.
Figure 6 shows the effects of AF-DX 116 (1.0 μM) on the M2/M3interaction calculated in case 1 before and after varying degrees of M3 receptor inactivation. It can be seen that the dextral shift in the concentration-response curve caused by AF-DX 116 is only 2.3-fold before M3 receptor inactivation. As the degree of receptor inactivation increases to 99%, the dextral shift increases to 5.5-fold. However, with further M3 receptor inactivation (99.8%), the shift declines to 3.6-fold. These results are summarized in Table 2 along with the results from cases 2 and 3.

Simulation of effects of AF-DX 116 and M3receptor inactivation on response to an agonist mediated through interaction of an agonist with two receptors (i.e., M2 and M3 receptors; model I). Curves are shown before (▪, ■) and after 90% (●, ○), 99% (▴, ▵), and 99.8% (♦, ⋄) M3 receptor inactivation in the absence (filled symbols) and presence (open symbols) of AF-DX 116 1 μM. Response of M2 receptor was less sensitive than that of M3receptor (KM2 = 0.1;KM3 = 0.01). Each point was calculated using eq. 8 as described in the .
The results shown in Figs. 5A and 6 are similar to the behavior of the guinea pig colon in that the contractile response is sensitive only to pertussis toxin after 4-DAMP mustard treatment, not before. In addition, the response is relatively insensitive to AF-DX 116 both before and after extensive M3 receptor inactivation.
We also considered an alternative model (model II) for M2/M3 receptor interactions consisting of two components (A and B). Stimulation of M3 receptors leads to the activation of two parallel signaling pathways. One pathway causes a direct contraction of smooth muscle (component A). The other pathway has no effect by itself, but it is required for M2 receptor-mediated contractions (component B). By itself, the M2receptor is incapable of eliciting contractions. Thus, component B represents an M2/M3interaction, with the response being conditional on stimulation of both receptors. We show the behavior of component B in Fig. 8. In Figs. 9and 10, we show the behavior of the complete model obtained by combining both components (A plus B).

Simulation of effects of AF-DX 116 and M3receptor inactivation on response mediated through interaction of an agonist with two receptors (i.e., M2 and M3; component B, model II). Curves are shown before (▪, ■) and after 90% (●, ○), 99% (▴, ▵), and 99.8% (♦, ⋄) M3receptor inactivation in the absence (filled symbols) and presence (open symbols) of AF-DX 116 (1 μM). A, response of M2receptor was less sensitive than that of M3 receptor (KM2 = 0.03; KM3= 0.01). B, response of M3 receptor was less sensitive than that of M2 receptor (KM2 = 0.003; KM3 = 0.01). C, response of M2 and M3 receptors were equisensitive (KM2 = 0.01; KM3= 0.01). Each point was calculated using eq. 10 as described in the.

Simulation of effects of AF-DX 116 and M3receptor inactivation on combined response of model II (i.e., combination of components A and B). Curves are shown before (▪, ■) and after 90% (●, ○), 99% (▴, ▵), and 99.8% (♦, ⋄) M3 receptor inactivation in the absence (filled symbols) and presence (open symbols) of AF-DX 116 (1 μM). A, response of M2 receptor was less sensitive than that of M3receptor (KM2 = 0.03;KM3 = 0.01). B, response of M3receptor was less sensitive than that of M2 receptor (KM2 = 0.003; KM3= 0.01). C, response of M2 and M3 receptors were equisensitive (KM2 = 0.01;KM3 = 0.01). Each point was calculated using eq. 13 as described in the .

Simulation of effects of pertussis toxin treatment and M3 receptor inactivation on combined response of model II (i.e., combination of components A and B). Curves are shown before (▪, ■) and after 90% (●, ○), 99% (▴, ▵), and 99.8% (♦, ⋄) M3 receptor inactivation before (filled symbols) and after (open symbols) pertussis toxin treatment. A, response of M2 receptor was less sensitive than that of M3 receptor (KM2 = 0.03; KM3= 0.01). B, response of M3 receptor was less sensitive than that of M2 receptor (KM2 = 0.003; KM3 = 0.01). C, response of M2 and M3 receptors were equisensitive (KM2 = 0.01; KM3 = 0.01). Theoretical points before and after pertussis toxin treatment were calculated using eqs. 13 and 7 as described in the .
Figure 7 illustrates the effects of AF-DX 116 (1 μM) and M3 receptor inactivation on the response of component A of model II. This response is highly sensitive, so progressive receptor inactivation (90%, 99%, and 99.8%) causes a sizable shift to the right in the concentration-response curve, followed by a depression in its maximum (Table3). With greater than 99.8% receptor inactivation, the response is essentially eliminated. Assuming a pKD value of 6.1 for the AF-DX 116-M3 receptor complex, it can be shown that AF-DX 116 should cause a 2.3-fold shift to the right in the concentration-response curve before and after receptor inactivation.

Simulation of effects of AF-DX 116 and M3receptor inactivation on response to an agonist mediated through a single receptor site (model II, component A). Curves are shown before (▪, ■) and after 90% (●, ○), 99% (▴, ▵), and 99.8% (♦, ⋄) receptor inactivation in the absence (filled symbols) and presence (open symbols) of AF-DX 116 (1 μM). Each point was calculated using eq. 7 as described in the .
Effects of M3 receptor inactivation and AF-DX 116 (1 μM) on the responses predicted by model II
Component B of model II is illustrated in Fig.8. In component B, the affinity of the agonist is the same for both the M2 and M3 receptor; however, the sensitivities of the signals elicited by the two receptors are varied, allowing us to modulate the contribution of either receptor to the response (see ). We therefore describe three cases within component B: in case B1, the M2 signal is less sensitive than the M3 signal (Fig. 8A); in case B2, the M3 signal is less sensitive than the M2 signal (Fig. 8B); and in case B3, the sensitivities of both the M2 and M3 receptor are the same (Fig. 8C). Figure 8shows the effects of AF-DX 116 (1 μM) on the contractile-response curves of the model before and after selective inactivation of M3 receptors.
When the signal elicited by the M2 receptor is less sensitive to the agonist than that elicited by the M3 receptor (Fig. 8A), 90% M3 receptor inactivation has little effect on the concentration-response curve of component B1. Moreover, AF-DX 116 (1 μM) caused a large 20-fold dextral shift in the concentration-response curve both before and after 90% M3 receptor inactivation. However, as the degree of M3 receptor inactivation increased to 99% and 99.8%, the concentration-response curve shifts to the right and shows a decrease in maximum (Table 3). Under these conditions, the dextral shift caused by AF-DX 116 declined to 6.1- and 2.3-fold at 99% and 99.8% receptor inactivation, respectively. Thus, when the M2 signal is less sensitive to the agonist than the M3 signal, component B1 initially behaves like an M2 response with high sensitivity to AF-DX 116. However, as the degree of M3 receptor inactivation increases, component B1 behaves more like an M3 response with low sensitivity to AF-DX 116.
Figure 8B illustrates the response of component B2 when the M2 signal is more sensitive to agonist than the M3 signal. Under these conditions, the response is highly sensitive to M3 receptor inactivation. There was a progressive shift to the right followed by a decrease in maximal response as the degree of M3 receptor inactivation increased (Table 3). AF-DX 116 (1 μM) caused a 6.3-fold increase in the EC50 value before M3 receptor inactivation and 2.3-fold increases after 90%, 99%, and 99.8% M3 receptor inactivation (Table 3). Thus, when the M2 signal is more sensitive to agonist then the M3 signal, component B2 behaves more like an M3 response (i.e., compared with Fig. 8A) both before and after M3 receptor inactivation.
When the sensitivities of the M2 and M3 receptors are identical (Fig. 8C), the effects of M3 receptor inactivation were similar to those observed in Fig. 8B (Table 3). AF-DX 116 (1 μM) caused a large 12.6-fold dextral shift in the concentration-response curve before M3 receptor inactivation. After M3 receptor inactivation, AF-DX 116 caused 2.3- to 3.3-fold increases in the EC50 value (Table3).
Having described the individual components of model II (A and B), we calculated the complete model by combining the two components. Figure9 shows the results of these calculations. Figure 9A shows the composite response resulting from the summation of components A and B1. It can be seen that increasing the degree of M3 receptor inactivation causes a progressive shift to the right and a decrease in the maximal response. However, the effects of M3 receptor inactivation are not as great as those observed with a homogeneous M3 receptor-mediated response, like component A shown in Fig. 7. Thus, the summation of component A with component B1 yields a composite response that exhibits a refractoriness to M3 receptor inactivation, like that observed in the guinea pig colon (see Fig. 2). Moreover, this composite response is relatively insensitive to AF-DX 116. At 1 μM, AF-DX 116 caused only 2.3- to 3.7-fold shifts in the concentration response curves both before and after varying degrees of M3 receptor inactivation. These results are similar to those observed in the guinea pig colon.
Figure 9B shows the composite response derived from a combination of components A and B2. Increasing amounts of M3receptor inactivation caused a progressive dextral shift of the concentration-response curve and a decrease in the maximum (Table 3). After 99.8% M3 receptor inactivation, the response was essentially eliminated. AF-DX 116 (1 μM) caused 2.3- to 2.6-fold increases in the EC50 value before and after varying degrees of M3 receptor inactivation (Table 3). Similar results were obtained for the composite response derived from a combination of components A and B3 (Fig. 9C). Compared with the model shown in Fig. 9A, the models of Figs. 9B and C were more sensitive to M3 receptor inactivation.
Figure 10A show the effects of pertussis toxin on the composite response resulting from a combination of components A and B1. It can be seen that the response is initially insensitive to pertussis toxin; however, after progressive M3 inactivation, the response becomes pertussis toxin sensitive. These results are similar to those observed in the guinea pig colon (see Fig. 3). The effects of pertussis toxin were modeled by assuming that the toxin prevented M2receptor signaling (see ). The effects of pertussis toxin on the composite responses derived by a combination of components A and B2 and a combination of components A and B3 are shown in Fig. 10, B and C. It can be seen that these composite responses remained fairly insensitive to pertussis toxin both before and after M3 receptor inactivation.
Phosphoinositide Hydrolysis.
When transfected into cells in high abundance, M2 receptors have been shown to mediate a weak phosphoinositide response through a pertussis toxin-sensitive G protein. Consequently, we investigated whether a pertussis toxin-sensitive phosphoinositide response could be detected in the guinea pig colon after inactivation of most of the M3 receptors. Such a mechanism could account for a direct, pertussis toxin-sensitive contractile response mediated by the M2 receptor after inactivation of most of the M3 receptors with 4-DAMP mustard. However, such a response should be potently antagonized by AF-DX 116, which was not observed in the guinea pig colon (see Fig. 4). Nevertheless, we wished to explore the possible coupling of M2 receptors to phospholipase Cβ via a pertussis toxin-sensitive G protein. First, we characterized the phosphoinositide response by measuring the ability of the subtype-selective antagonists AF-DX 116, p-F-HHSiD, and pirenzepine to shift the concentration-response curve for oxotremorine-M-induced phosphoinositide hydrolysis to the right (Fig.11). We estimated theKB values of the antagonists from these rightward shifts as described in Experimental Procedures. These KB values are listed in Table 4 together with the binding affinities (KD values) of the same antagonists measured in Chinese hamster ovary cells transfected with the M2 and M3 subtypes of the muscarinic receptor (Esqueda et al., 1996; Ehlert et al., 1997). Also listed in Table 4 are the pKB for the antagonists determined in contractile studies in the guinea pig colon (Sawyer and Ehlert, 1998). The binding assays for theseKD estimates were conducted in a HEPES-buffered KRB solution similar to that used in our contractile and phosphoinositide hydrolysis assays. It can be seen that theKB values of AF-DX 116,p-F-HHSiD, and pirenzepine are in agreement with their respective KD values for the M3 receptor but not with those of the M2 receptor (Table 4). These values also were in close agreement with the values obtained previously from contractile studies conducted in the guinea pig colon (Table 4). There also was a lack of agreement between the antagonistKB values of AF-DX 116,p-F-HHSiD, and pirenzepine and their respectiveKD values for the M1 (6.24, 7.08, 7.77), M4(6.96, 7.08, 7.23), and M5 (5.29, 6.26, 6.55) subtypes, as reported by Esqueda et al. (1996) and Ehlert et al. (1997). Therefore, we conclude that the M3receptor mediates phosphoinositide turnover in the guinea pig colon under standard conditions.

Competitive antagonism of phosphoinositide response to oxotremorine-M in guinea pig colon. Phosphoinositide response to oxotremorine-M in the absence (■) and presence (▪) of p-F-HHSiD (0.1 μM) (A), AF-DX 116 (1 μM) (B), and pirenzepine (1 μM) (C).
Effect of 4-DAMP Mustard Treatment and Pertussis Toxin on Phosphoinositide Hydrolysis Elicited by Oxotremorine-M.
We measured the pertussis toxin sensitivity of the phosphoinositide response both before and after 4-DAMP mustard treatment to investigate the possible contribution of the M2 receptor to the response. Colonic slices were incubated with 4-DAMP mustard (40 nM) in the presence of AF-DX 116 (1 μM) for 2 h and washed extensively. Incubation for 2 h with 4-DAMP mustard caused a significant 9.34-fold (p = 1.5 × 10−5) increase in the EC50value and a 60% reduction in the maximal response to oxotremorine-M (Fig. 12). As in the functional assays, pertussis toxin did not affect the response to oxotremorine-M; in fact, there was a slight increase in the hydrolysis of phosphoinositides (Fig. 12). Unlike the contractile experiments, however, pertussis toxin had no inhibitory effect on phosphoinositide hydrolysis after 2-h 4-DAMP mustard treatment (Fig. 12). In fact, there was a slight potentiation of the response as well. Consequently, the M2 receptor does not appear to couple with a phosphoinositide signaling pathway to rescue the contractile response after 4-DAMP mustard treatment.

Effects of pertussis toxin treatment on phosphoinositide response to oxotremorine-M in control and 2-h 4-DAMP mustard-treated colon. Phosphoinositide hydrolysis was measured in 2-h 4-DAMP mustard-treated (▵, ▴) and untreated (■, ▪) colon from pertussis toxin-treated (▪, ▴) and untreated (■, ▵) guinea pigs. Each data point represents mean ± S.E.M. of four experiments.
Discussion
There is an abundance of data on smooth muscle demonstrating that it is primarily the M3 receptor that mediates contraction to muscarinic agonists in the absence of other contractile and relaxant agents (Eglen, 1996; Ehlert et al., 1997). This condition occurs in variety of smooth muscle types, including the circular and longitudinal muscles of the colon (Zhang and Buxton, 1991; Sawyer and Ehlert, 1998). The data of the present report are also consistent with this hypothesis; the contractile response was sensitive to 4-DAMP mustard and insensitive to pertussis toxin and the M2-selective antagonist AF-DX 116.
However, a different picture emerges when we consider the residual contractile response that persisted after selective inactivation of most of the M3 receptors with 4-DAMP mustard. This response was highly sensitive to pertussis toxin and relatively insensitive to 4-DAMP mustard. Both of these characteristics suggest an M2 receptor mechanism; but the contractile response was relatively insensitive to the M2-selective antagonist AF-DX 116. To explain this enigma, we suggest that the response may represent an interaction between M2 and M3receptors. Accordingly, the M2 receptor may mediate contractions through some mechanism that depends on a low level of signaling through the M3 receptor. By itself, this low level of M3 signaling is insufficient to generate a sizable contraction, yet it enables activated M2 receptors to elicit contraction. The results of our mathematical analysis are not inconsistent with this hypothesis.
We considered two models for the interaction between M2 and M3 receptors. In both models, it was assumed that activation of M2receptors has no effect by itself, but M2receptors can interact with M3 receptors to trigger a contraction. In the first model (model I), it was assumed that the M2 receptor simply potentiated the contractile response of the M3 receptor. This model yielded results similar to the behavior of the colon when it was assumed that oxotremorine-M elicits contraction through the M3 receptor with greater potency than that with which it activates the potentiating M2 signal (case 1). Under this condition, oxotremorine-M can elicit a maximal contraction through the M3 receptor at concentrations that are insufficient to activate the M2 pathway. Thus, the M2receptor does not contribute to the contraction in untreated tissue, and consequently, the response is pertussis toxin insensitive. However, after inactivation of most of the M3 receptors, the M2 receptor contributes to the contraction, rendering it sensitive to pertussis toxin. Model I, case 1, also predicts that AF-DX 116 should cause only a 2.3- to 3.6-fold shift to the right in the concentration-response curve both before and after extensive M3 receptor inactivation. Such results were observed in the colon. After a moderate degree of M3 receptor inactivation, model I, case 1, predicts that AF-DX 116 (1.0 μM) should cause a 4.2- to 5.5-fold dextral shift in the concentration-response curve. The largest shift that we observed for AF-DX 116 after a moderate degree of receptor inactivation was 4.3-fold (Sawyer and Ehlert, 1998).
We also investigated an additional model (model II) that provided a somewhat better approximation of our data in the colon. Model II is based on the assumption that the M3 receptor signals through two parallel pathways. The first pathway gives rise to a simple M3 receptor-mediated contraction (component A), and the second pathway is silent by itself but interacts with a silent M2 receptor to generate a contraction (component B) (see Fig. 8). Component A is more sensitive, so when contractile responses to oxotremorine-M are measured in untreated tissue, the contractions are due to activation of M3 receptors. However, after selective inactivation of M3 receptors, the sensitivity of component A is greatly reduced so component B becomes the more sensitive component.
Our analysis of component B provides an understanding of the competitive antagonism of a response to an agonist that is mediated through an interaction between two receptors. We have shown that there is a tendency for the antagonism to resemble that predicted for the less sensitive receptor mechanism. Consider an example where receptor X interacts with receptor Y to trigger a response. If the signal elicited by receptor X is less sensitive than that elicited by receptor Y, then the nature of the competitive antagonism of the interaction will tend to resemble that predicted for receptor X. If a highly Y-selective competitive antagonist is used, its calculated pKB value will be less than that of the Y receptor and may approach that of the X receptor. On the other hand, if an X-selective antagonist is used, its calculated pKB value will be equivalent to that of the X receptor.
Model II agreed best with the contractile data when it was assumed that the M3 signal of component B was more sensitive than the M2 signal (i.e., the conditions of model II, B1). Under these conditions, the model predicts that the competitive antagonism of component B1 by AF-DX 116 (1.0 μM) should resemble that expected for the less sensitive receptor signaling pathway: M2. This antagonism was manifest as a relatively large AF-DX 116-induced shift (19.6-fold) in the concentration-response curve of component B1 (see Fig. 8A). Nevertheless, the complete response obtained by combining components A and B1 exhibited relatively low sensitivity to AF-DX 116 (1 μM) (see Fig. 9A). This behavior can be explained by the greater sensitivity of component A, which is capable of mediating a maximum contraction at agonist concentrations that are insufficient to elicit a response through component B1. It is only after M3receptors are inactivated that component B1 dominates the contractile response. However, under these conditions, the M3signal of component B1 is less sensitive than the M2 signal, and consequently, the competitive antagonism of component B1 resembles that expected for an M3 response, with low sensitivity to AF-DX 116. As a result, the complete response (i.e., the combination of components A and B1) exhibited relatively low sensitivity to AF-DX 116 both before and after varying degrees of M3 receptor inactivation. However, careful inspection of the theoretical curves shows that the shift in the concentration-response curved caused by AF-DX 116 (1.0 μM) increased from 2.3-fold before receptor inactivation to 3.7-fold after 90% inactivation of M3 receptors. With further receptor inactivation, the AF-DX 116-induced shift declined to 2.3-fold. Interestingly, we observed a similar phenomenon in the guinea pig colon. We found that the shift in the concentration response curve caused by AF-DX 116 increased from 2.2-fold in untreated tissue to 4.3-fold after 2-h 4-DAMP mustard treatment (Sawyer and Ehlert, 1998). After 4-h treatment, the AF-DX 116-induced shift returned to a low value of 1.8-fold (see Fig. 4).
So far, we have no direct evidence on the possible signaling mechanisms involved in the interaction between M2 and M3 receptors. Nevertheless, it is interesting to note that muscarinic agonists trigger a nonselective cation conductance in smooth muscle (Benham et al., 1985; Inoue and Insenberg, 1990a). This conductance is pertussis toxin sensitive, and it is enhanced by calcium, particularly in the colon, where the response is completely dependent on calcium (Inoue and Insenberg, 1990b; Lee et al., 1993). The pertussis toxin sensitivity and the calcium requirement suggest a mechanism involving both M2 and M3 receptors. Bolton and Zholos (1997) have shown that the competitive antagonism of the muscarinic-induced cation conductance is consistent with a response mediated by both the M2 and M3 receptors. Perhaps this conductance is involved in the interaction between M2 and M3 receptors.
The low sensitivity of the postulated interaction between M2 and M3 receptors raises the question of its physiological significance. What function could the interaction have if muscarinic agonists are capable of eliciting contraction through a more potent M3 mechanism? This question is particularly relevant if the nature of the interaction simply involves an M2 receptor-mediated potentiation of M3 receptor-mediated contractions (model I). However, if the interaction involves an M2 potentiation of an alternate M3 signal that is incapable of eliciting contraction by itself (model II), then this mechanism could function in isolation of the standard M3 receptor-mediated contraction. Moreover, it could have a pattern of distribution distinct from that of the standard M3 receptor mechanism, making it the dominant mechanism at some neuroeffector junctions.
Acknowledgments
We thank Regina Bailon for her excellent technical assistance.
Appendix
This appendix describes our calculations for generating the theoretical concentration-response curves for the two models of M2 and M3 receptor interactions. Some general calculations that apply to both models are described first. Later, we describe the specific details of each model in Model I and Model II.
In our analysis, we assumed that occupancy obeys the law of mass-action and that the concentration-response curve of the agonist obeys a logistic equation. These assumptions are valid because 1) the intact cell contains high concentrations of GTP and the binding of agonists to M2 and M3 muscarinic receptors obeys mass-action behavior when measured in the presence of guanine nucleotides (Ehlert and Yamamura, 1995) and 2) the concentration-response curves of muscarinic agonists in the guinea pig colon and ileum typically obey a logistic equation with Hill coefficients of approximately 2. Black and Leff (1983) have shown that if occupancy obeys mass-action behavior and the concentration-response curve obeys logistic behavior, then the relationship between occupancy and response must also be logistic. Consequently, we calculated the response (R) using the following logistic equation:
Model I.
Model I is based on the assumption that activation of M2 receptors yields no response by itself; however, activation of the M2 receptor potentiates the stimulus elicited by the M3receptor by a factor of 5.0. To describe this interaction, we begin by calculating the response to the M3 receptor (RM3) using eq. 2:
Model II.
Model II is based on the assumption that the M3 receptor signals through two parallel pathways to trigger a contractile response. The first pathway, component A, represents an M3 receptor-mediated contractile response that exhibits no interaction with the M2receptor. This component is described by eq. 7. The second, alternative signaling pathway of the M3 receptor is inactive by itself, but it is required for agonist elicitation of a contractile response through the M2 receptor. This M3 signal is designated as the conditional M3 response (RM3-cond). By itself, the M2 receptor is also inactive, and its signal is designated as the conditional M2 response (RM2-cond). The interaction between these two signals is equivalent to the product of the two conditional responses of the M2(RM2-cond) and M3 (RM3-cond) receptors. This product was multiplied by 0.3 because we observed that the pertussis toxin-sensitive contractile response that persisted after 4-DAMP mustard treatment had a maximal response of approximately 30% that of the maximal response before 4-DAMP mustard treatment. This interaction is defined as the response of component B (RB):

Relationship between combined response of model II and summation of components A and B. Curve was derived from eq. 13 in the . Dashed line shows simple additive relationship between components A and B.
Footnotes
-
Send reprint requests to: Frederick J. Ehlert, Ph.D., Department of Pharmacology, College of Medicine, University of California, Irvine, Irvine, CA 92697-4625.
-
↵1 This work was supported by National Institutes of Health Grant NS30882.
- Abbreviations:
- AF-DX 116
- [[2-[(diethylamino)methyl]-1-piperidinyl]acetyl]-5,11-dihydro-6H-pyrido[2,3b][1,4]benzodiazepine-6-one
- 4-DAMP mustard
- N-(2-chloroethyl)-4-piperidinyl diphenylacetate
- KRB
- Krebs-Ringer-bicarbonate
- p-F-HHSiD
- para-fluoro-hexahydro-sila-diphenidol
- Received June 23, 1998.
- Accepted November 9, 1998.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}