


Abstract
Previous work has shown that the genes encoding each α1-adrenoceptor subtype are coexpressed throughout the peripheral vascular system. We have evaluated subtype-selective antibodies as tools to determine the extent of protein expression in arteries. The α1A-, α1B-, and α1D-adrenoceptors were detected in the medial layer of the aorta, caudal, femoral, iliac, renal, superior mesenteric, and mesenteric resistance arteries. In Rat1 fibroblasts expressing each subtype, immunoreactivity was noted both on the cell surface and in a perinuclear orientation. Intense α1B-adrenoceptor immunostaining was similarly localized in cultured femoral and renal vascular smooth muscle cells. Although the cellular localization appeared to be the same, immunoreactivity obtained with α1A- and α1D-adrenoceptors was much less intense than that with the α1B-adrenoceptor. The α1A-adrenoceptor selective agonist A-61603 was 22-fold more potent in activating renal artery contraction when compared with the femoral artery. The expression of each α1-adrenoceptor was significantly decreased by in vivo application of antisense oligonucleotides targeted against each subtype. Inhibition of the expression of only one, the α1A in renal and the α1D in femoral arteries, reduced the contractile response to naphazoline. The results show: 1) subtype-selective antibodies can be used in tissues and cell culture to localize the α1-adrenoceptor subtypes, 2) in addition to expression on the cell surface, the α1-adrenoceptors are expressed intracellularly, and 3) despite expression of all adrenoceptors, a single subtype mediates the contractile response in the femoral and renal arteries.
The α1-adrenoceptor (AR) family is a member of the G protein-coupled superfamily of receptors. These receptors have the familiar proposed structure of seven transmembrane-spanning domains connected by hydrophilic loops alternately exposed to the extracellular and intracellular environment (Minneman and Esbenshade, 1994; Strader et al., 1994; for reviews see Bylund et al., 1995; Graham et al., 1996;Guarino et al., 1996). Three genes encoding unique α1-AR subtypes (Cotecchia et al., 1988; Schwinn et al., 1990; Lomasney et al., 1991; Perez et al., 1991) have been discovered. Hieble et al. (1995) have proposed a nomenclature consisting of the α1A (formerly referred to as the α1c-)-, the α1B-, and the α1D (formerly the α1a- or α1a/d-)-AR subtypes.
The coupling of the α1-ARs to second messenger systems has been examined in detail. These receptors are coupled to the activation of phospholipase C (Schwinn et al., 1991; Perez et al., 1993) via pertussis toxin-insensitive G proteins of the Gq/11 family (Wu et al., 1992). In addition to mobilizing intracellular calcium (which would occur subsequent to activation of phospholipase), the α1-ARs have also been shown to activate calcium influx via both voltage-dependent and -independent calcium channels (Ljung and Kjellstedt, 1987; Han et al., 1992; Sayet et al., 1993; Minneman and Esbenshade, 1994; Lazou et al., 1994). Macrez-Leprêtre et al. (1997) used antisense oligonucleotides to show that α1-ARs utilize Gαq to activate phosphoinositide hydrolysis and Gα11 to induce the release of calcium from intracellular stores. Minneman and associates studied the coupling of the α1-AR subtypes in HEK-293 cell lines expressing each individual receptor (Theroux et al., 1996). These authors noted that there were marked differences in the coupling efficiency of the receptors with the α1A-AR being the most efficiently coupled and the α1D-AR the least efficiently coupled of the subtypes. This group also showed that there was only a small receptor reserve for each subtype in these cell lines (Minneman and Esbenshade, 1994) and that this reserve was similar for all receptors.
The α1-ARs are key effectors utilized by the sympathetic nervous system to regulate cardiovascular function. The genes encoding these receptors are widely expressed in the heart and peripheral arteries (Perez et al., 1994; Price et al., 1994; Piascik et al., 1995; Guarino et al., 1996). In addition, a variety of functional assays have demonstrated the presence of these receptors in cardiovascular tissues (Han et al., 1990; Elhawary et al., 1992; Bylund et al., 1995; Kenny et al., 1995; Piascik et al., 1995; Testa et al., 1995; Leech and Faber, 1996; Zhou and Varga, 1996).
Despite these reports, a systematic study of the expression of the α1-ARs and correlation to function has not been done. Recently we used a subtype-specific antibody to show that the α1B-AR can be detected in a series of peripheral blood vessels (Piascik et al., 1997). In spite of this widespread distribution, the receptor was not linked to the activation of contraction in a majority of the arteries in which it was expressed. This has led us to postulate that a receptor can be expressed but not participate in contractile regulation.
In this report we examined the feasibility of using commercially available antibodies against each of the α1-ARs to determine the distribution of all subtypes in peripheral arteries. We then used laser scanning confocal microscopy to characterize the cellular distribution of these receptors in cells cultured from femoral and renal arteries and Rat1 fibroblasts. The functional expression of these receptors was assessed in a series of contractile studies with selective agonists and antisense oligonucleotides.
Materials and Methods
Animals.
Male Sprague-Dawley rats weighing between 250 to 350 g were used in all experiments. The aorta, caudal, femoral, iliac, mesenteric resistance, renal, and superior mesenteric arteries were removed and processed according to the experimental protocols described below. All protocols involving the use of animals were reviewed and approved by the Institutional Animal Care and Use Committee.
Antibodies.
Antibodies (raised in goats) against residues in the cytoplasmic tail of the α1-ARs were obtained from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA). The α1A-AR antibody was raised against the human sequence. This subtype epitope differs from the rat sequence by two amino acids. The α1B-AR antibody epitope, also that of the human sequence, differs from the rat sequence by one amino acid. These immunizing peptide sequences are as follows with the bold letters indicating where the rat and human sequences differ:
Immunohistochemistry in Arterial Sections.
The blood vessels were removed, placed in a cold, sterile physiological saline solution (PSS) of the following composition (in mM): NaCl, 130; KCl, 4.7; KH2PO4, 1.18; MgSO4-7 H2O, 1.17; CaCl2-2 H2O, 1.6; NaHCO3, 14.9; dextrose, 5.5; disodium EDTA, 0.03, and cleaned of extraneous tissue. The vessels were then cut into rings of approximately 2 mm in length, placed upright in Tissue Tek OCT Compound (Miles Inc., IN) mounting media and quick frozen (−50°C in isopentane cooled with dry ice.) Thirteen-micrometer sections were cut on a cryostat. Sections were applied to poly-l-lysine-subbed, circularly etched slides and stored at −80°C until use. The slides were thawed and fixed with 4% paraformaldehyde. A blocking solution [1% BSA and 0.1% Triton X-100 in PBS (0.88% NaCl, 0.1375% Na2HPO4, and 0.02% NaHPO4, pH 7.4)] with 10% donkey serum (Jackson Immunologicals, West Grove, PA) was placed on the sections and incubated at room temperature for 2 h. After removal of the blocking solution, the primary antibody (α1D-AR cat. no. s.c.-1475, α1B-AR cat. no. s.c.-1476, α1A-AR cat. no. s.c.-1477; Santa Cruz Biotechnologies) was applied at various dilutions (1:25–1:100) in blocking solution (without the donkey serum) and left overnight at 4°C in a humidity chamber. The slides were then rinsed and placed in blocking solution with 10% donkey serum for 30 min. The slides were washed in PBS followed by application of an affinity-purified fluorescein isothiocyanate (FITC)-conjugated donkey anti-goat IgG (1:200 dilution, Jackson Immunologicals) secondary antibody. After 1 h in the dark at room temperature, the secondary antibody was removed by washing with PBS. Vectashield mounting media (Vector Laboratories, Inc., Burlingame, CA) was placed on the slide and a 24- × 50-mm glass coverslip (Gold Seal) applied. The slides were then stored at 4°C in the dark until viewing. The tissue sections were examined using a Nikon Microphot fluorescence microscope with either a 40× dry (0.7) or a 60× oil immersion (1.4) lens.
Cultured Vascular Smooth Muscle Cells.
Our procedure is adapted from that of Gunther et al. (1982). The artery was removed and cleaned of fat and connective tissue. The vessel was then cut longitudinally, spread out flat, and the endothelial cells were removed by shearing with a cotton swab. The tissue was cut into smaller pieces and incubated in 0.5 ml of 0.1% collagenase (lot 46A034; Worthington Biochemical Corp., Freehold NJ), 0.125 mg/ml elastase (Worthington), and 2 mg/ml BSA in Hanks’ balanced salt solution (Gibco BRL, Gaithersburg, MD) for 90 min in a 37°C shaking water bath with intermittent trituration. The digestion mixture was centrifuged at 1600 rpm for 5 min. The excess collagenase solution was removed and the resulting pellet was washed with Hanks’ balanced salt solution. The pellet was resuspended in 10 ml of Dulbecco’s modified Eagle’s medium (DMEM; Gibco BRL) supplemented with 10% fetal bovine serum (Gibco BRL) and a 1% antibiotic/antimycotic mixture (Gibco BRL), transferred to a cell culture flask and maintained at 37°C in 5% CO2. The cells were fed every 2 to 3 days with DMEM supplemented as above. The cells were trypsinized every 5 to 7 days. Rat1 fibroblasts were also cultured in DMEM supplemented as above with the addition of 500 μg/ml geneticin (Gibco-BRL). After the cells were confluent they were plated out on sterile 20- × 20-mm glass coverslips and returned to the cell culture incubator for 48 h to allow attachment. Experiments were performed on cells that had been passaged twice and cells that had been passaged a minimum of six and maximum of nine times. These cells are referred to as greater than six passages in the figures.
Immunocytochemistry (ICC) and Laser Scanning Confocal Microscopy.
The coverslips with cells attached were washed in PBS and fixed in 1.5 ml 3.7% formaldehyde in PBS for 10 min. All washes were in PBS containing 0.05% BSA and 0.1% saponin to permeabilize the cells. After fixation, the cells were washed again and 100 μl of blocking solution (10% donkey serum, 1% BSA in PBS) was placed on the coverslips and incubated at room temperature for 1 h. After washing, 100 μl of the primary antibody (diluted in PBS with 1% BSA and 1% saponin) was placed on the coverslips and incubated at room temperature for 2 h. At this time the coverslips were washed and the FITC-labeled secondary antibody (in PBS) was applied to the coverslips for 1 h. After a wash in PBS, the coverslips were mounted onto glass slides with Vectashield and stored at 4°C in the dark until viewing. The cells were imaged using the Nikon RCM 8000 laser-scanning confocal microscope, which is equipped with an NCF Fluor 40 X Water Immersion objective. The argon ion laser emits at 488 nm to excite FITC. FITC absorbs at 488 nm and the fluorescence collected was emitted at a wavelength no greater than 545 nm. Images were stored on an optical disk recorder as eight-bit, 512 × 483 pixel tagged image file format files. The files were transferred to an 80486 microcomputer for off-line analysis using the program Metamorph (Universal Imaging, West Chester, PA). In certain studies confocal microscopy was used to assess the effect of antisense oligonucleotide application on α1-AR expression in femoral or renal artery sections. Fluorescence measurements were confined to areas between the highly autofluorescent elastin bands. The mean fluorescence in these defined regions was calculated for antisense-treated and -untreated controls. Values from three sections per slide were averaged and are reported as mean relative fluorescent units ± the S.E.M (n represents the number of slides immunostained from a single surgery). Differences in these mean fluorescent values were determined by a t test for unpaired data.
Experiments with Immunizing Peptides.
In certain studies the antibodies were preabsorbed to their respective immunizing peptides (see above) before use in ICC or immunohistochemistry protocols. Peptides, in a 5-fold excess for cultured cells and a 10-fold excess for blood vessel sections, were incubated with antibodies overnight at 4°C and used in immunostaining protocols as described above.
In Vitro Assessment of Contractile Function.
Isolated blood vessel segments were prepared by techniques routinely used in our laboratory (Piascik et al., 1994, 1995, 1997). Arterial segments were removed and placed in cold PSS. Three-millimeter ring segments of the femoral or renal arteries were cut and cleaned of surrounding fat and connective tissue. Stainless steel or platinum wires were threaded through the lumen of each vessel. One wire was connected to a fixed base and the other to a micrometer clamp to adjust passive force on the tissues. The tissues were mounted in water-jacketed muscle baths containing PSS maintained at 37°C under constant oxygenation (95% O2, 5% CO2; pH 7.4). A passive force of 1 g was then placed on the vessels. Previous studies have shown that this passive force gives optimal agonist responses. Changes in force generation were recorded using Grass FT.03 force transducers connected to a Grass model 7 polygraph.
In Vivo Application of Antisense Oligonucleotides.
Oligonucleotides were obtained from Life Technologies. The following sequences, antisense to the translational start site, were used: α1A-AR- AGA GAG AAG CAC CAT, α1B-AR-CAG ATC GGG ATT CAT, α1D-AR-GTC TCG GAA AGT CAT. These oligonucleotides were applied to the femoral and renal arteries in a pluronic gel medium as we have reported previously (Piascik et al., 1997). A 40%, w/v, solution of pluronic F-127 gel (Sigma Chemical Co., St. Louis, MO) was prepared in water at 4°C by mixing on a platform shaker overnight in a cold room. F-127 pluronic gel solutions are liquid at 4°C and solidify at room temperature (Schmolka, 1972). Sufficient oligonucleotide was added to the cold gel solution to give the desired concentration (see below). Addition of oligonucleotide reduces the pluronic gel concentration to 30%. Care was taken to ensure that all pipet tips and storage tubes were kept at 4°C. Animals were anesthetized with a mixture of ketamine/acepromazine. After a surgical incision, the femoral or renal artery was located and gently cleaned of adhering tissue. Forty microliters of the pluronic gel/oligonucleotide solution was applied around the artery. After the gel solidified, the artery was gently placed back into its original position and the wound was closed with surgical staples. The animal was then allowed to recover for 24 h before the femoral or renal arteries were removed for experimentation.
Agonist Dose-Response Curves.
A-61603, an α1A-AR-selective agonist, was obtained from Dr. Arthur A. Hancock (Abbott Laboratories, Abbott Park, IL) and prepared in 0.1% ascorbic acid. Naphazoline was also prepared in 0.1% ascorbic acid. Cumulative dose-response curves were prepared for each agonist. Before beginning the dose-response curve, the muscles were assessed for viability by sequentially challenging with 80 mM KCl and 1 μM phenylephrine.
Statistical Analysis.
Concentration response curves represent the mean of a minimum of four experiments on individual blood vessel segments, each of which was from a different animal. Only one concentration response curve was run on any segment. ED50 and 95% confidence limits were calculated. A two-way ANOVA followed by Student-Newman-Kuels (SNK) analysis was used to determine where statistically significant differences existed between the various treatment conditions. In all figures the data are presented as the mean ± S.E.M. Stars indicate statistically significant differences at the p < .05 level.
Results
Commercially available subtype-selective antibodies were used to examine the vascular distribution of the α1-ARs. To demonstrate the specificity and the lack of cross-reactivity between the antibodies, immunostaining was performed on Rat1 fibroblasts that had been stably transfected with each of the α1-ARs. Using laser scanning confocal microscopy, intense immunostaining was obtained with each antibody when assessed in fibroblasts expressing the corresponding receptor (see Fig. 1). Immunoreactivity was detected along the boundary of the cell indicating a cell membrane localization. There was also perinuclear and cytoplasmic immunostaining, which indicates the possibility that the receptors are located in intracellular compartments. Little immunofluorescence was obtained when the antibodies were used in cells that did not express the receptor against which the antibodies were raised (see Fig. 1). To further demonstrate the lack of nonspecific attachment to cellular components, the antibodies were incubated with their respective immunizing peptides before use in experimental protocols with fibroblasts. Under these conditions, the immunostaining intensity was significantly reduced (see Fig. 2). In another series of studies, the permeabilizing agent saponin was omitted from the buffers. In these experiments, little immunoreactivity was noted (see Fig. 2). The epitopes are on the cytoplasmic tail of the receptor, and the lack of staining in the absence of an agent that would make the cell permeable to the antibody also argues that the immunoreactivity is specific. When experiments with immunizing peptides or without saponin were performed in blood vessel sections or cultured vascular smooth muscle cells, the immunostain was significantly reduced (data not shown). In aggregate, these data argue that the α1-AR antibodies possess the necessary specificity to be used in these studies.

Immunostaining of Rat1 fibroblasts expressing each of the α1-AR subtypes. Each cell line was treated with the indicated receptor-specific antibody as described in Materials and Methods.
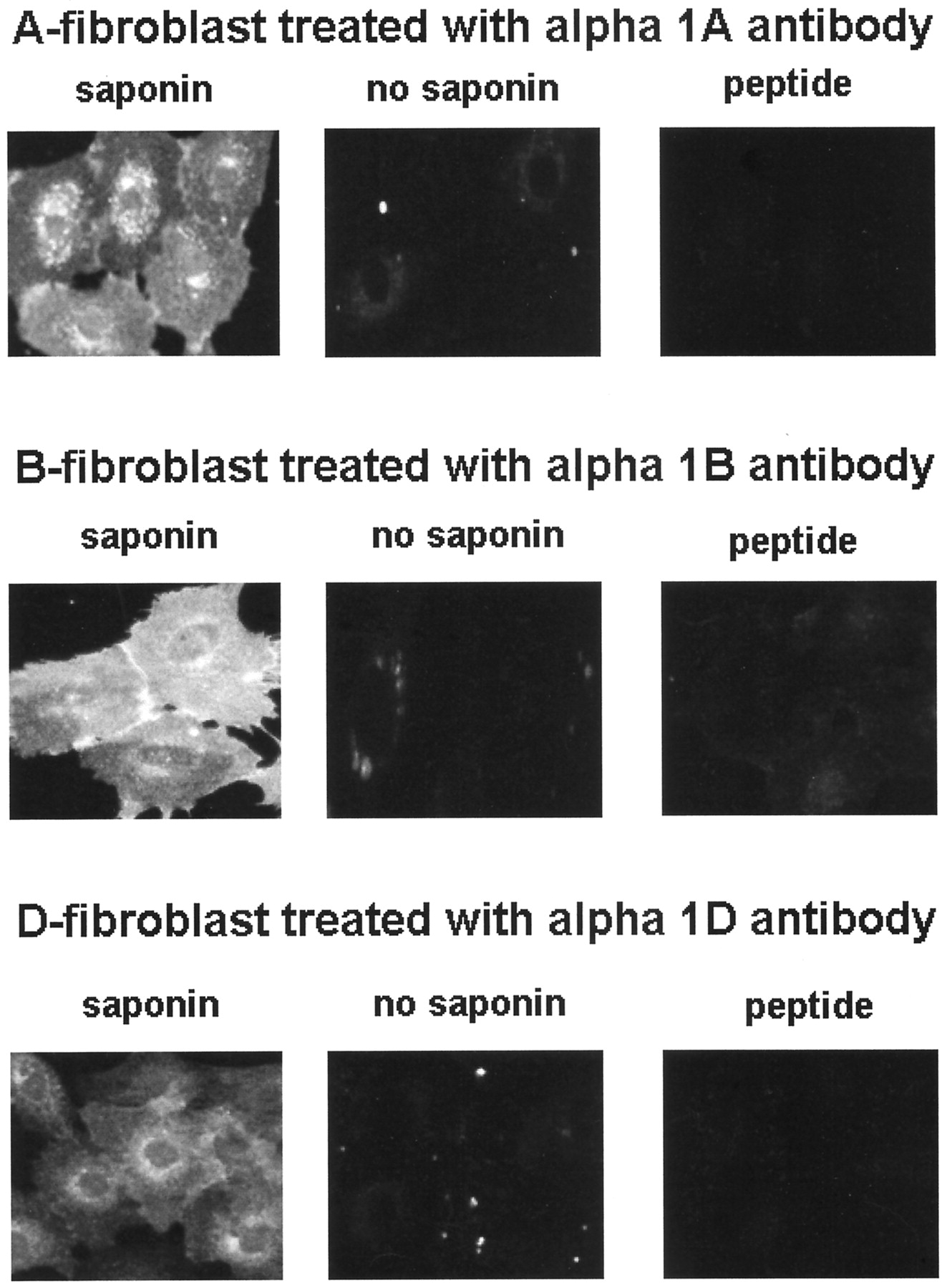
Immunostaining of Rat1 fibroblasts expressing each of the α1-AR subtypes. Conditions were permeabilization in the presence of saponin (control staining, “saponin”), the removal of saponin from the incubation buffers (“no saponin”), or preincubating the α1-AR antibody with its respective immunizing peptide (“peptide”). Experiments were carried out as described in Materials and Methods.
Receptor antibodies were then used to detect α1-AR expression in frozen sections of a series of peripheral arteries. A yellow autofluorescence corresponding to the elastin layers and endothelial lining was always present in these vessel sections. When receptor antibodies were applied, intense green immunoreactivity was obtained throughout the medial layer, indicating that each subtype is expressed on each blood vessel examined (see Fig.3).

Immunohistochemical localization of the three α1-AR subtypes in frozen sections from the aorta, caudal, femoral, iliac, mesenteric resistance, renal, and superior mesenteric arteries. Immunostaining was viewed by fluorescence microscopy. Experiments were carried out as described inMaterials and Methods.
Although these data demonstrate receptor expression, they do not give the same resolution as is obtainable with the optical sectioning that can be performed with confocal microscopy. To this end we cultured smooth muscle cells from the femoral and renal arteries. These cells stained intensely with smooth muscle specific actin, indicating a smooth muscle phenotype (data not shown). In contrast with the intense immunostaining seen with all antibodies in blood vessel sections, the immunostaining pattern seen in culture was dependent on the cell line, the antibody, and the time in culture (see Fig.4). After two passages, faint immunostaining was obtained with the α1-AR antibodies in femoral artery cells. The immunoreactivity increased significantly after six passages in culture. Of the three receptor antibodies, the α1B-AR gave the most intense staining pattern. After six passages, there was a vibrant immunostaining detected along the margin of the cell, indicating a cell surface locale for the α1B-AR. In addition, immunostaining was detected in a perinuclear orientation. Immunostaining obtained with the α1A- and α1D-AR antibodies was not as intense nor as clearly defined as that obtained with the α1B-AR. Although in certain instances we could note a clear cell membrane or perinuclear localization, the majority of the immunostaining patterns obtained were faint, diffuse, and not of sufficient intensity to allow us to define the cellular localization of the signal. Nonetheless, this immunoreactivity did decrease in experiments in which the α1A- and α1D-AR antibodies were preincubated with their respective immunizing peptides (data not shown). This result would argue that the immunostaining that we did observe was due to a specific antigen-antibody reaction.

Immunocytochemical localization of the α1-AR subtypes in vascular smooth muscle cells cultured from femoral or renal arteries. Immunostaining was viewed by confocal microscopy. Experiments were carried out as described inMaterials and Methods.
Renal artery cells were stained with all antibodies at both two and greater than six passages. As in the femoral artery, cell staining in the renal artery was most intense with the α1B-AR antibody. Immunoreactivity was detected along the boundary of the renal artery cells and in a perinuclear orientation. The quality of the α1A- and α1D-AR immmunolabeling in renal artery cells was similar to that seen in femoral artery cells. Thus, although we can present evidence of receptor expression in renal artery cells, we are unable to make definitive conclusions regarding the cellular localization of the signal of the α1A- and the α1D-ARs.
The coupling of these receptors to the activation of smooth muscle contraction was assessed in functional studies with the α1A-AR selective agonist A-61603 and the partial agonist naphazoline. A-61603 stimulated the contraction of the renal artery with an ED50 and 95% confidence limits of 1.5 nM (0.963–2.4; see Fig.5). This ligand was much less potent in activating femoral artery contraction, with an ED50 and 95% confidence limits of 33.4 nM (18.4–52.7; Fig. 5). These data indicate that the α1A-AR is coupled to the activation of contraction in the renal artery whereas another subtype(s) modulates the femoral artery response.

Cumulative log dose-response curves for the α1A-AR specific agonist, A-61603, in the femoral and renal arteries. Data are the mean of determinations on four ring segments, each from a different artery. ED50 and 95% confidence limits were calculated for each dose-response curve.
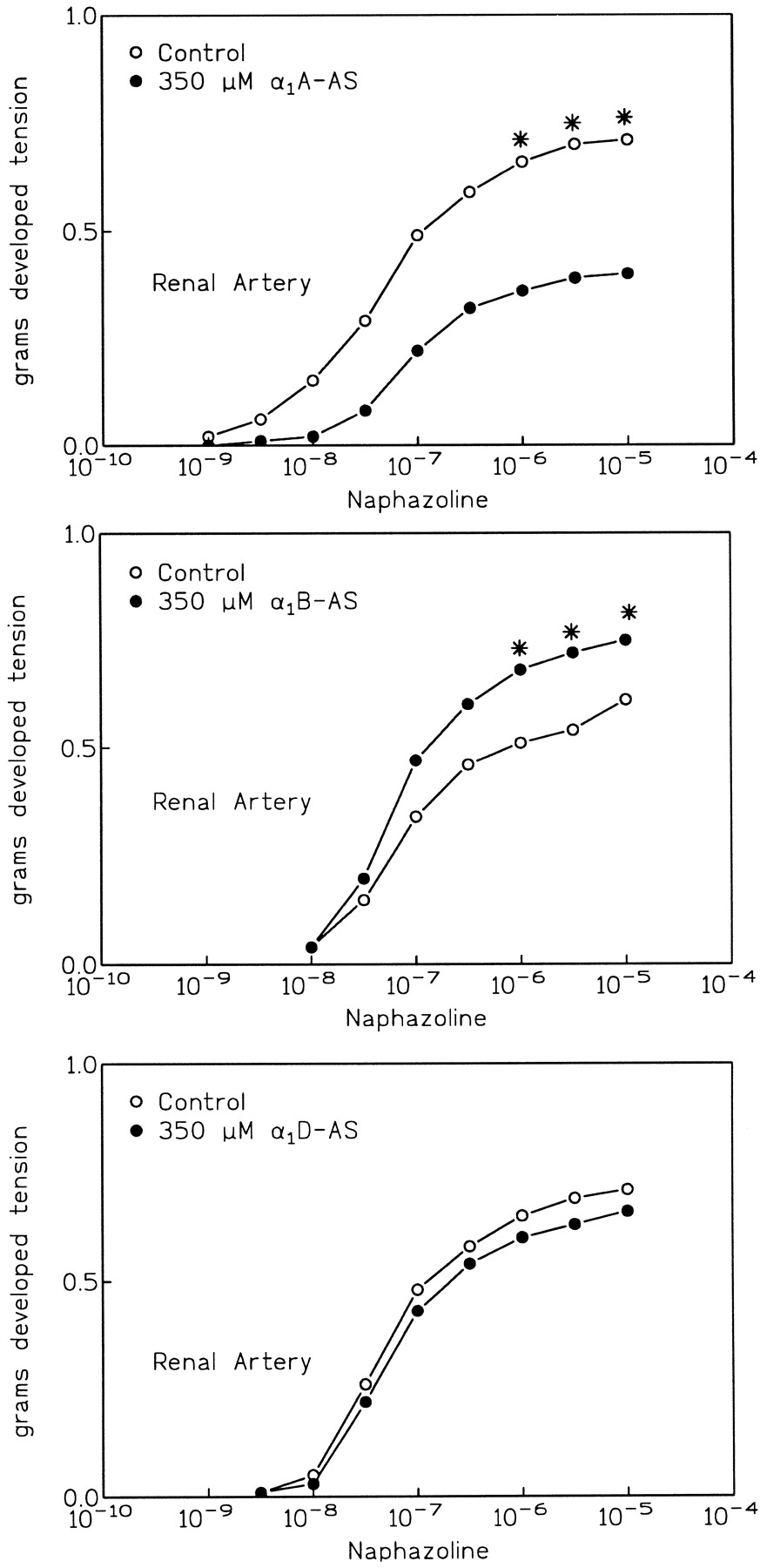
Antisense oligonucleotides were used to inhibit the expression of each of the α1-ARs in the femoral and renal arteries. Antisense was applied using a pluronic gel delivery matrix as we have described previously (Piascik et al., 1997). In both the femoral and renal arteries, this treatment reduced the immunostaining of the α1-AR against which the oligonucleotide was targeted (Fig. 6). These images were viewed by confocal microscopy followed by computerized image analysis. In all cases the intensity of the fluorescent staining was significantly less after antisense oligonucleotide application (see Table1). Antisense treatment had no effect on the immunoreactivity of those receptors not targeted by the oligonucleotide (data not shown). In the renal artery, antisense against the α1A-AR inhibited the response to naphazoline. The oligonucleotide against the α1D-AR was without effect (see Fig.7). Surprisingly, antisense against the α1B-AR actually potentiated the naphazoline response [compare the control (open circles) to treated (closed circles)]. This potentiation was statistically significant.


A. Immunohistochemistry on femoral arterial sections. Left, control antibody staining. Right, effect of treatment with antisense oligonucleotides targeted against each receptor. Antisense application and immunohistochemistry were performed as described inMaterials and Methods. B. Immunohistochemistry on renal arterial sections. Left, control antibody staining. Right, effect of treatment with antisense oligonucleotides targeted against each receptor. Antisense application and immunohistochemistry were performed as described inMaterials and Methods.
Mean fluorescence values obtained from laser scanning confocal microscopy of α1-AR immunostaining in arterial sections

The effect of application of antisense oligonucleotides on the naphazoline cumulative log-dose-response curve in the femoral artery. Antisense was applied as described inMaterials and Methods. Each point represents the mean of determinations on at least seven individual segments. A two-way ANOVA followed by SNK analysis was used to assess statistically significant differences. (∗), statistical significance atp < .05.
Despite inducing a reduction in α1A-AR immunoreactivity in the femoral artery (see Fig. 6 and Table 1) the α1A-AR antisense had no effect on the femoral response to naphazoline (see Fig. 8). In this artery it was the α1D-AR oligonucleotide that inhibited naphazoline action. Inhibition of expression of the α1B-AR had no effect on naphazoline response in the femoral artery.

The effect of application of antisense oligonucleotides on the naphazoline cumulative log-dose-response curve in the renal artery. Antisense was applied as described inMaterials and Methods. Each point represents the mean of determinations on at least eight individual segments. A two-way ANOVA followed by SNK analysis was used to assess statistically significant differences. (∗), statistical significance at p< .05.
Discussion
Subtype-selective antibodies were used to examine the vascular distribution of the α1-AR subtypes. In Rat1 fibroblasts stably transfected with each of the α1-ARs, the antibodies stained only those cells that expressed the receptor against which they were raised. In fibroblasts, cultured smooth muscle cells, and frozen blood vessel sections, prior incubation of the antibody with the immunizing peptide significantly reduced the intensity of the immunostaining. Removal of the permeabilizing agent saponin, which prevents antibody access to the receptor epitope, also reduced the immunoreactivity. Finally, the use of antisense oligonucleotides to decrease α1-AR expression in arteries also resulted in a decrease in immunoreactivity. These data are good evidence that the antibodies are specific, have little cross-reactivity, and are thus suitable for use in detecting receptor expression.
In our previous work we showed that the genes encoding each of the α1-ARs were coexpressed throughout the peripheral vascular system (Piascik et al., 1995; Guarino et al., 1996). We now show that all α1-ARs are expressed as protein in these same arteries. We previously used a different antibody to map the distribution of the α1B-AR (Piascik et al., 1997). This antibody was raised to a 10-amino acid region of the C terminus of the hamster α1B-AR sequence. The α1B-AR antibody used in this study was also raised against 18 amino acids in the C terminus but from the human sequence (see Materials and Methods for details). The results obtained with the different α1B-AR antibodies regarding receptor distribution agree very well.
Although data showing coexpression of all three α1-ARs in the medial smooth layer are new, they do not address the interesting question as to why the vasculature would express three regulatory receptors, each of which could potentially activate contraction. Recent work on the localization of the α2-AR has shown that there is a differential cellular localization of these receptors (von Zastrow et al., 1993). In transfected fibroblasts, the α2C-AR was localized to the cell membrane and intracellular compartments whereas the α2A-AR was found exclusively on the cell membrane. In COS7 cells transiently transfected with green fluorescent protein-labeled α1-AR constructs, the α1A-AR was localized in a perinuclear fashion whereas the α1B-AR was detected throughout the entire border of the cell (Hirasawa et al., 1997).
Our data obtained in fibroblasts expressing each of the receptors support the idea that the α1-ARs are expressed not only on the cell surface but also in intracellular compartments. In these fibroblasts, immunostaining was noted on the cell surface and also in a perinuclear orientation. To determine if differences in receptor localization could modulate α1-AR responsiveness in vascular smooth muscle, we examined the distribution of the α1-ARs in cells cultured from the femoral and renal arteries. We used cells that had been passaged twice and up to a maximum of nine times. All cells stained intensely with smooth muscle actin, indicating that they retain the appropriate phenotype in culture. We observed differences in the immunostaining pattern that were dependent on the receptor, cell type, and the time in culture. This indicates that the expression of the α1-ARs can be affected by culture conditions. These data also suggest caution in using cultured smooth muscle cells to delineate α1-AR function. Little receptor immunostaining was observed with any antibody in early passage femoral artery cells. The α1B-AR gave the most vibrant immunostaining in later passage femoral artery cells and all renal artery cells examined. Receptor immunoreactivity was noted on the cell membrane and in a perinuclear orientation. The α1B-AR immunostaining pattern in cultured vascular smooth muscle cells agrees with that observed in Rat1 fibroblasts expressing the α1B-AR. Therefore these data show that in addition to membrane expression, the α1B-AR can also be detected in intracellular compartments. The antibodies against the α1A- and α1D-ARs gave good immunostaining in fibroblasts expressing each of these receptors. However, the results in cultured smooth muscle cells were equivocal. We did observe immunostaining for these receptors both on the cell membrane and in intracellular compartments. However, in most experiments α1A- and α1D-AR antibodies did not give the clear localization pattern that we observed with the α1B-AR, and definitive resolution could not be obtained with antibodies against these receptors. This could indicate that the expression of these receptors is low. However, we do obtain good immunostaining in frozen sections from intact blood vessels. Therefore, the culturing conditions may affect the expression of the α1A- and the α1D-AR such that their expression is depressed in culture. It is also possible that these commercial antibodies are not sufficiently sensitive to ICC in smooth muscle cells. Although never the subject of a systematic study, the quality of these antibodies has been questioned by workers in the field. Clearly the quality of the α1A- and α1D-AR immunoreactivity in smooth muscle cells could be much better. Yet we show specific and intense immunostaining with these antibodies in Rat1 fibroblasts. Furthermore, we show with a variety of control conditions that the immunostaining we obtain in smooth muscle cells is not due to nonspecific interactions or cross-reactivity. Therefore, the lack of good immunostaining of smooth muscle cells may represent a phenomenon related to maintaining the cells in culture.
Although we report that all α1-ARs are present in the vessels we examined, our results with antisense oligonucleotides would argue that expression is not sufficient to link the receptor to contraction. We show that treatment of the femoral and renal arteries with antisense oligonucleotides decreases the expression of each of the α1-ARs targeted by the antisense. However, this decrease was not always associated with a reduction in functional responsiveness. Only in the renal artery were decreases in α1A-AR expression associated with a decrement in naphazoline responsiveness. In the femoral artery the same α1A-AR antisense oligonucleotide was without effect. The only oligonucleotide that caused a decrease in contraction in the femoral artery was that directed against the α1D-AR. These data are supported by experiments that showed that the α1A-AR agonist A-61603 selectively activated renal artery contraction when compared to the femoral. These results are consistent with the idea that a receptor can be expressed but not function in contractile regulation. It is unlikely that the inhibitory effects we observe represent nonspecific oligonucleotide effects. If this were the case, application of any oligonucleotide would be expected to depress contractile function, which did not occur. In a previous study we showed that antisense treatment is not associated with a decrease in the response of smooth muscle to KCl depolarization or the actions of serotonin (Piascik et al., 1997), which further argues against nonspecific effects. Furthermore, inhibition of α1B-AR expression actually enhances naphazoline responsiveness in the renal artery. We previously made the same observation in studies with phenylephrine as the agonist (Piascik et al., 1997). These data suggest that there is a reciprocal relationship between the expression of the α1-ARs and contractile regulation in the renal artery. However, this same phenomenon was not observed in the femoral artery.
The present study goes beyond our previous work in that it demonstrates that the oligonucleotide treatment actually decreases receptor expression. In the past, we used phenylephrine as the agonist. The fact that we did not see an inhibitory response on the actions of this compound with a given antisense oligonucleotide could be due to the presence of spare receptors. The antisense constructs may not have decreased receptor expression below a critical level to see a decrement in contractile function. According to receptor theory, a partial agonist must occupy all receptors to produce a maximal effect and does not have a reserve. Therefore, using naphazoline eliminates the problem of spare receptors. Naphazoline responsiveness is unaffected by decreasing the expression of two of the α1-ARs in each test artery (α1A and α1B in femoral and α1Band α1D in renal). This is evidence that those receptors do not play a regulatory role in the contraction of those arteries.
The mechanisms controlling the linkage between receptor expression and the regulation of smooth muscle contraction are not known. In smooth muscle cells different subtypes could be expressed predominantly in intracellular compartments and be sequestered from the cell surface. In this fashion a receptor could be expressed but not localized to the cell membrane where it can be activated by hydrophilic agonists.
We show all receptors can be expressed within the cell indicating the possibility that expression may regulate function. However, we show an abundance of α1B-AR staining on the cell border, which we assume is associated with the cell membrane. Yet in the femoral and renal arteries we have no evidence, from this or a variety of other work, that the α1B-AR has any role in contractile regulation. This would indicate that regulation of cellular localization alone is not a control point in modulating α1B-AR signaling or that the α1B-AR exists on the cell surface to regulate other cellular processes.
Acknowledgments
We thank Glaxo-Wellcome Pharmaceuticals for their gift to Dr. Perez of Rat1 fibroblasts expressing each of the α1-ARs.
Footnotes
-
Send reprint requests to: Dr. Michael T. Piascik, Ph.D., Vascular Biology Research Group, Department of Pharmacology, The University of Kentucky College of Medicine, 800 Rose Street, Lexington, Kentucky 40536-0084. E-mail: mtp{at}pop.uky.edu
-
↵1 This work was supported by National Institutes of Health Grants HL-38120 (M.T.P.), HL-56910 (R.W.H.), and HL-52544 (D.M.P.), and a grant-in-aid (M.T.P.) and an Established Investigator Award (D.M.P.) from the American Heart Association.
- Abbreviations:
- AR
- adrenoceptor
- DMEM
- Dulbecco’s modified Eagle’s medium
- FITC
- fluorescein isothiocyanate
- ICC
- immunocytochemistry
- PSS
- physiological saline solution
- SNK
- Student-Newman-Kuel’s
- Received September 30, 1998.
- Accepted March 1, 1999.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}