


Abstract
The ionic mechanism of actions of ruthenium red was examined in rat anterior pituitary GH3 cells. In whole-cell recording experiments, ruthenium red reversibly caused an inhibition of Ca2+-activated K+ current [IK(Ca)] in a dose-dependent manner. The IC50value of ruthenium red-induced inhibition of IK(Ca) was 15 μM. Neither carbonyl cyanide m-chlorophenyl hydrazone (CCCP; 10 μM), an uncoupler of oxidative phosphorylation in mitochondria, nor cyclosporin A (200 nM), an inhibitor of the mitochondrial permeability transition pore, affected the amplitude of IK(Ca). In inside-out configuration, application of ruthenium red (50 μM) into the bath medium did not change single-channel conductance but significantly suppressed the activity of large-conductance Ca2+-activated K+ channel (BKCa) channels. The ruthenium red-induced decrease in the channel activity of BKCa channels was reversed by an increase in intracellular Ca2+ concentration. Ruthenium red also shifted the activation curve of BKCa channels to positive membrane potentials. The change in the kinetic behavior of BKCa channels caused by ruthenium red in these cells is due to a decrease in mean open time and an increase in mean closed time. Ruthenium red (50 μM) did not affect the amplitude of voltage-dependent K+ current but produced a significant reduction of voltage-dependent L-type Ca2+ current. These results indicate that ruthenium red can directly suppress the activity of BKCa channels in GH3 cells. This effect is independent on the inhibition of Ca2+ release from internal stores or mitochondria.
Large-conductance Ca2+-activated K+(BKCa) channels are found in many tissues and may participate in a variety of cellular processes. These channels are gated open by both binding of intracellular Ca2+and by membrane depolarization (Tseng-Crank et al., 1994; Wei et al., 1994; Kaczorowski et al., 1996). Because of the large unitary conductance and dense distribution, the activity of these channels is partly responsible for spike repolarization and the early afterhyperpolarization that follows each action potential (Sah and McLachlan, 1991; Kaczorowski et al., 1996). Previous studies also showed that presynaptic Ca2+ signals and transmitter release from nerve terminals can be regulated by the activity of BKCa channels (Robitaille and Charlton, 1992). Cloning and expression of both α and β subunits of this channel will allow us to examine structural components that control these gating processes (Kaczorowski et al., 1996; Toro et al., 1998). To determine the structural components of channel gating and to study the role that these channels may play in different cellular processes, small organic modulators for the gating of these channels will be useful tools.
Ruthenium red, an inorganic polycationic dye, is known to antagonize capsaicin-induced response (Staszewska-Woolley and Woolley, 1991; Wang and Hakanson, 1993; Lo et al., 1997), to inhibit Ca2+ release from the mitochondria (Broekemeier et al., 1994; Gunter et al., 1998; Wood and Gillespie, 1998) or from endoplasmic reticulum (Cheek et al., 1991; Sah and McLachlan, 1991;Otun et al., 1996), and to induce neurodegeneration (Velasco and Tapia, 1997). This inorganic polycationic dye has been reported to block voltage-dependent Ca2+ current in various cell types, including chromaffin cells (Gomis et al., 1994), mouse sensory neurons (Duchen, 1992), and synaptosomes and neuromuscular preparations (Hamilton and Lundy, 1995; Tapia and Velasco, 1997). However, to date, the effect of ruthenium red on K+ currents still remains controversial. For instance, ruthenium red can inhibit Ca2+-activated K+ channels in neurons (Sah and McLachlan, 1991; Wann and Richards, 1994) and smooth myocytes (Duridanova et al., 1996; Hirano et al., 1998). Ruthenium red was also found to have no effect on voltage-dependent K+ current in chromaffin cells (Gomis et al., 1994) or bladder smooth myocytes (Hirano et al., 1998). In contrast, ruthenium red is recently reported to enhance both voltage-dependent and Ca2+-activated K+currents in mouse motor nerve terminals (Lin and Lin-Shiau, 1996).
In the present study, we sought to examine the effect of ruthenium red on Ca2+-activated K+currents in rat pituitary GH3 cells and to determine whether ruthenium red affects the activity of BKCa channels. These cells exhibit a wide variety of ionic conductances, including at least two types of Ca2+-activated K+conductance (Ritchie, 1987). These results show a ruthenium red-induced inhibition of BKCa channels in these cells.
Materials and Methods
Cell Culture.
The clonal strain GH3cell line, originally derived from a rat anterior pituitary adenoma, was obtained from American Type Culture Collection [(CLL-82.1), Manassas, VA]. GH3 cells were grown in monolayer culture in 50-ml plastic culture flasks in a humidified environment of 5% CO2/95% air at 37°C. Cells were maintained at a density of 106/ml in 5 ml of Ham’s F-12 nutrient media (Life Technologies, Grand Island, NY) supplemented with 15% heat-inactivated horse serum (v/v), 2.5% fetal calf serum (v/v), and 2 mM l-glutamine (Life Technologies). Cells were subcultured once a week, and a new stock line was generated from frozen cells (frozen in 10% glycerol in medium plus serum) every 3 months. The experiments were performed after 5 or 6 days of subcultivation (60–80% confluence).
Electrophysiological Measurements.
Immediately before each experiment, GH3 cells were dissociated, and an aliquot of the cell suspension was transferred to a recording chamber positioned on the stage of an inverted phase-contrast microscope (Diaphot-200; Nikon, Tokyo, Japan). The microscope was coupled to a video camera system with magnification up to 1500× to continually monitor cell size during the experiments. Cells were bathed at room temperature (20–25°C) in normal Tyrode’s solution containing 1.8 mM CaCl2. The patch pipettes were prepared from Kimax capillary tubes (Vineland, NJ) using a vertical two-step electrode puller (PB-7; Narishige, Tokyo, Japan), and the tips were fire-polished with a microforge (MF-83; Narishige). The resistance of the patch pipette was 3 to 5 MΩ when it was immersed in normal Tyrode’s solution. Voltage pulses were digitally generated by a programmable stimulator (SMP-311; Biologic, Claix, France). Ionic currents were recorded with glass pipettes in whole-cell or inside-out configuration of patch-clamp technique, using an RK-400 patch amplifier (Biologic) (Hamill et al., 1981; Wu et al., 1998b). All potentials were corrected for liquid junction potential that developed at the tip of the pipette when the composition of pipette solution was different from that of bath. Tested drugs were applied by perfusion or added to the bath to obtain the final concentrations indicated.
Data Recording and Analysis.
The signals, consisting of voltage and current tracings, were monitored with a digital-storage oscilloscope (model 1602; Gould, Valley View, OH) and recorded on-line using a digital audio tape recorder (model 1204, Biologic). After the experiments, the stored data were then fed back and digitized at 5 to 10 kHz with a Digidata 1200 analog-to-digital device (Axon Instruments, Foster City, CA) interfaced to a Pentium-grade computer and pClamp 6.03 software package (Axon Instruments). Voltage-activated currents recorded during whole-cell experiments are stored digitally without leakage correction and analyzed using Clampfit subroutine (Axon Instruments) to establish a current-voltage relationship for ionic currents.
Single-channel currents were analyzed with Fetchan and Pstat subroutines in pClamp software (Axon instruments). Multi-Gaussian adjustments of the amplitude distributions between channels were used to determine unitary currents. The functional independence between channels was verified by comparing the observed stationary probabilities with the values calculated according to the binomial law. The number of active channels in the patch N was counted at the end of each experiment by perfusing a solution with 100 μM Ca2+ and then used to normalize opening probability at each potential. The opening probabilities were evaluated using an iterative process to minimize the χ2 calculated with a sufficiently large number of independent observations.
Open- or closed-lifetime distributions were fit with logarithmically scaled bin width by using the method of McManus et al. (1987). When the square root of the number of events in a bin was plotted against the open or closed lifetime, each component of the open- or closed-lifetime distribution appeared as a clear peak with the respective time constant falling in the vicinity of the distribution peak (Sigworth and Sine, 1987).
To calculate the percentage of inhibition of ruthenium red on Ca2+-activated K+ current [IK(Ca)], each cell was depolarized from 0 to +50 mV, and the current amplitudes during the application of ruthenium red was compared with the control value. The concentration of ruthenium red required to inhibit 50% of current amplitude was determined using a Hill function, y =Emax/{1 + (IC50n/[D]n)}, where [D] is the concentration of ruthenium red; IC50 and n are the half-maximal concentration of ruthenium red required to inhibit IK(Ca) and Hill coefficient, respectively; andEmax is ruthenium red-induced maximal inhibition of IK(Ca).
All values are reported as means ± S.E. The paired or unpaired Student’s t test and one-way ANOVA with least-significant-difference method for multiple comparison were used for the statistical evaluation of differences among the means. Differences between the values were considered significant whenp < .05.
Drugs and Solutions.
Tetraethylammonium chloride, carbonyl cyanide m-chlorophenyl hydrazone (CCCP), cyclosporin A, neuraminidase, and tetrodotoxin were purchased from Sigma Chemical (St. Louis, MO). Ruthenium red (trans-tetradecaamine di-μ-oxotriruthenium6+ hexachloride) was obtained from Research Biochemicals (Natick, MA). All other chemicals were commercially available and of reagent grade. The composition of normal Tyrode’s solution was as follows: 136.5 mM NaCl, 5.4 mM KCl, 1.8 mM CaCl2, 0.53 mM MgCl2, 5.5 mM glucose, and 5.5 mM HEPES-NaOH buffer (pH 7.4). To record K+ currents, the patch pipette was filled with solution: 140 mM KCl, 1 mM MgCl2, 3 mM Na2ATP, 0.1 mM Na2GTP, 0.1 mM EGTA, and 5 mM HEPES-KOH buffer 5 (pH 7.2). To record Ca2+ current, KCl inside the pipette solution was replaced with equimolar CsCl, and pH was adjusted to 7.2 with CsOH. In single-channel recording, high K+ bathing solution contained 145 mM KCl, 0.53 mM MgCl2, and 5 mM HEPES-KOH 5 (pH 7.4). The pipette solution contained 145 mM KCl, 2 mM MgCl2, and 5 mM HEPES-KOH (pH 7.2).
Results
Effect of Ruthenium Red on Ca2+-Activated K+ Current in GH3 Cells.
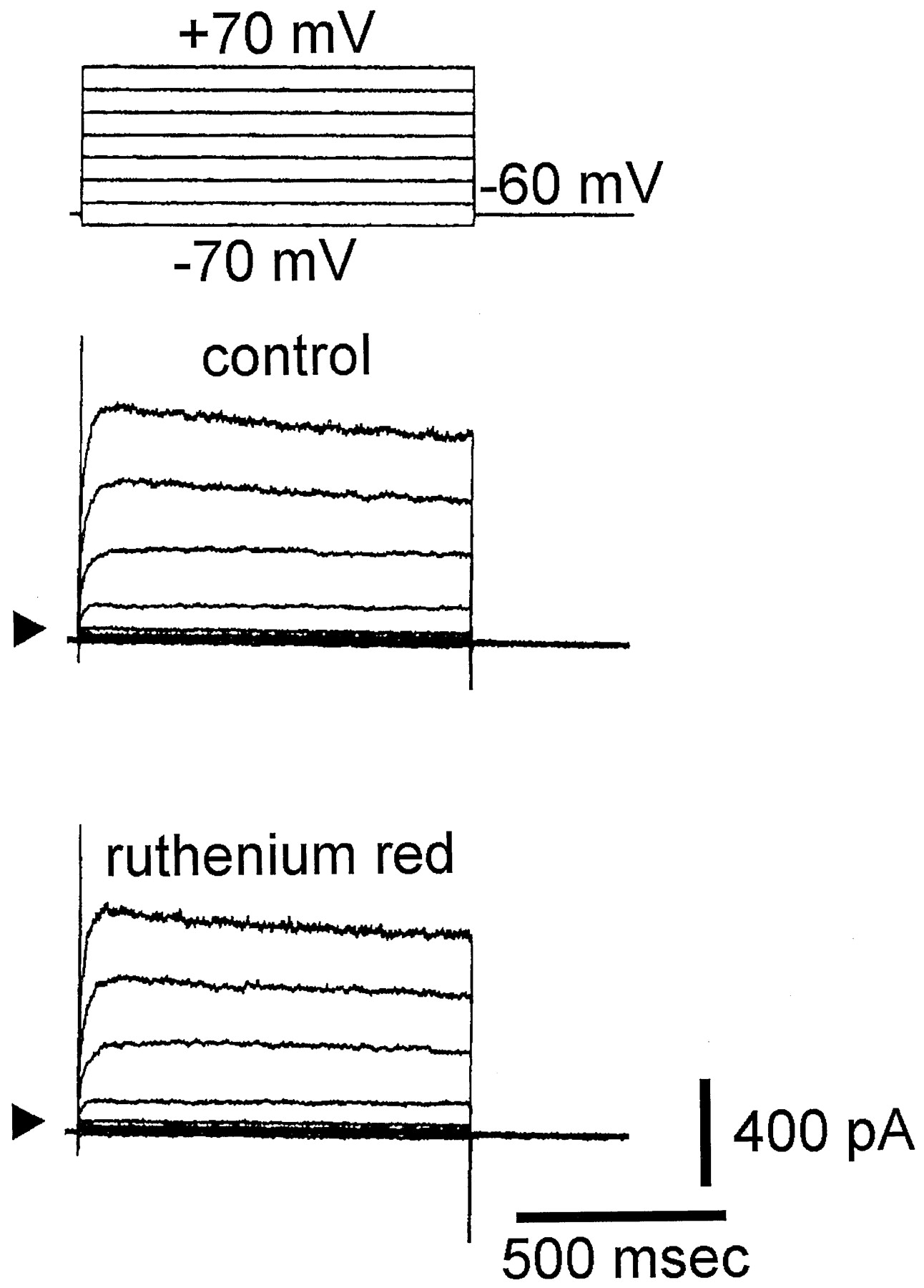
The whole-cell configuration of the patch-clamp technique was used to investigate the effect of ruthenium red on macroscopic ionic currents in these cells. In these experiments, the cells were bathed in normal Tyrode’s solution containing 1.8 mM CaCl2. To inactivate other voltage-dependent K+ currents, each cell was held at 0 mV. As shown in Fig. 1, when the cell was held at 0 mV, voltage pulses from −30 to +70 mV with 20-mV increments elicited a family of large, noisy, outward currents. The amplitudes of these currents were increased with greater depolarization. These outward currents are identified as Ca2+-activated K+ currents [IK(Ca)] (Olesen et al., 1994). Within 1 min of exposing the cells to ruthenium red (5 and 50 μM), the amplitude of outward currents was significantly decreased throughout the entire voltage-clamp step. For instance, when the voltage step from 0 to +70 mV was evoked, ruthenium red (50 μM) significantly decreased the current amplitude from 1609 ± 108 to 538 ± 48 pA (n = 7). This inhibitory effect was readily reversed after the removal of ruthenium red (Fig. 1). The averaged current-voltage (I-V) relations for these currents in the absence and presence of ruthenium red (5 and 50 μM) were shown in Fig. 1B. Figure1C shows the relationships between the concentration of ruthenium red and the percentage of inhibition of IK(Ca). Ruthenium red (5–200 μM) inhibited the amplitude of IK(Ca) in a concentration-dependent manner. The half-maximal concentration required for the inhibitory effect of ruthenium red on IK(Ca) was 15 μM, and 200 μM ruthenium red almost completely suppressed the amplitude of IK(Ca). These results demonstrate that ruthenium red significantly reduces the action on IK(Ca) in these cells.

Inhibitory effect of ruthenium red on IK(Ca) in GH3 cells. A, superimposed current traces in control, during the exposure to ruthenium red (5 and 50 μM) and after washout of ruthenium red. The cells, bathed in normal Tyrode’s solution containing 1.8 mM CaCl2, were held at 0 mV, and voltage pulses from −30 to +70 mV in 20-mV increments were then applied. The leftmost side in A indicates the voltage protocol. Arrows denote the 0-current level. B, the averaged current-voltage relations of outward currents measured at the end of voltage pulses in control (●), during exposure to 5 μM (▪), and 50 μM ruthenium red (■), and washout of ruthenium red (○) (mean ± S.E.;n = 6–10 for each point). C, concentration-dependent inhibition of IK(Ca) by ruthenium red. The relation between the percentage of inhibition of IK(Ca) and the concentration of ruthenium red was illustrated. Each cell was depolarized from 0 to +50 mV with a duration of 300 ms. Various concentrations of ruthenium red (5–200 μM) were applied. The amplitude of IK(Ca) during the application of ruthenium red was compared to the control value, i.e., in the absence of ruthenium red (mean ± S.E.; n = 6–10 for each point). The smooth line represents the best fit to the Hill equation). The values of IC50 and maximally inhibited percentage of IK(Ca) in the presence of ruthenium red were 15 μM and 98%, respectively. The Hill coefficient was 1.2.
Comparison between the Effect of Ruthenium Red and Those of CCCP, Cyclosporin A, 4-Aminopyridine, and Tetraethylammonium Chloride.
Effects of CCCP, cyclosporin A, 4-aminopyridine, tetraethylammonium chloride, and ruthenium red on IK(Ca) in GH3 cells were examined and compared. CCCP is an uncoupler of oxidative phosphorylation (Hehl et al., 1996), whereas cyclosporin A is considered to be a specific inhibitor of mitochondrial permeability transition (Shinohara et al., 1998). As depicted in Fig.2, CCCP (10 μM), cyclosporin A (200 nM), or 4-aminopyridine (1 mM) did not produce a significant reduction of the amplitude of IK(Ca). However, tetraethylammonium chloride (10 mM) and ruthenium red (50 μM) suppressed IK(Ca) significantly. Thus, ruthenium red-induced inhibition of IK(Ca) in GH3 cells appears to be unrelated to its inhibition of mitochondrial function. However, at this point, it is still not easily excluded that the reduction of IK(Ca) by ruthenium red may be due to a decrease in intracellular Ca2+ concentration.

Comparison of the effect of ruthenium red and CCCP, cyclosporin A, 4-aminopyridine, and tetraethylammonium chloride on IK(Ca). The cells were held at 0 mV, and voltage pulses to +50 mV (300 ms in duration) were applied. The amplitude of IK(Ca) in the control was considered to be 1.0, and the relative amplitude of IK(Ca) after application of each agent was then plotted. The parentheses denote the number of cells examined. Mean ± S.E. CCCP (10 μM); Cyc, cyclosporin A (200 nM); 4-AP, 4-aminopyridine (1 mM); TEA, tetraethylammonium chloride (10 mM); Ruth, ruthenium red (50 μM). ∗, significantly different from controls.
Inhibitory Effect of Ruthenium Red on the Activity of Large-Conductance Ca2+-Activated K+(BKCa) Channels in Inside-Out Patches.
To characterize the effect of ruthenium red on ionic currents, we also examined the effect of ruthenium red on BKCa channels. In an inside-out configuration, when ruthenium red (50 μM) was applied to the bath containing 1 μM Ca2+, the channel activity was profoundly reduced. A representative recording of BKCa channels after the application of ruthenium red is shown in Fig. 3. The opening probability of BKCa channels in control at the level of +80 mV was found to be 0.75 ± 0.06 (n = 12). One minute after the application of ruthenium red into the bath, the opening probability was significantly decreased to 0.08 ± 0.01 (n = 12). The further addition of 10 μM Ca2+ into the bath rapidly increased the channel activity to 0.55 ± 0.06 (n = 5). However, when ruthenium red (50 μM) was included in the pipette solution, the channel activity was unaffected (data not shown). Figure4A illustrates current-voltage relations of BKCa channels in the absence and presence of ruthenium red (50 μM). Fitting current amplitudes with a linear regression revealed a single-channel conductance of BKCa channels in control of 151 ± 6 pS (n = 14). This value was not significantly different from that (150 ± 5 pS, n = 10) measured in the presence of ruthenium red (50 μM). These results clearly indicate that ruthenium red causes no change in the single-channel conductance but inhibits the opening probability of BKCachannels in GH3 cells.

Effect of ruthenium red on BKCa channels in GH3 cells. The single-channel experiments in an excised inside-out membrane patch were conducted with symmetrical K+ concentration (145 mM). The bath solution contained 1 μM Ca2+. The membrane potential was held at +100 mV. A, the original current trace showing the change in the activity of BKCa channels after addition of ruthenium red. Channel openings are shown as upward deflection. The lower parts in A show the current traces obtained in expanded time scale. Note that the channel shows only a few, brief openings in the presence of ruthenium red (50 μM) and the further addition of 10 μM Ca2+ into the bath rapidly increases the channel activity. B, open probability (NPo) for the activity of BKCa channels shown in A plotted against time of recording. Bin width is 0.3 sec. Horizontal bars shown in the panel indicate the application of ruthenium red (50 μM) and Ca2+ (10 μM).

Effect of ruthenium red on the current-voltage relation of BKCa channels in GH3 cells. The cells were bathed in symmetrical K+ solution (145 mM), and the experiments were conducted under inside-out configuration. A, examples of BKCa channels in the absence (left) and presence (right) of ruthenium red (50 μM) measured from cells at various membrane potentials. Ruthenium red was applied to the bathing solution. The arrows and numbers at the beginning of each current trace mark the 0 current and voltage applied to the patch pipette, respectively. Upward deflections are the opening events of the channel. Currents were filtered at 1 kHz. B, the current-voltage relations of BKCa channels in the absence (●) and presence (○) of ruthenium red (50 μM). Of note, the single-channel conductance in the absence and presence of ruthenium red is nearly identical. C, the relationship between opening probability of BKCa channels and membrane potential in the absence (●) and presence (○) of 5 μM ruthenium red. The smooth lines represent the best fit to the Boltzmann equation.
Effect of Ruthenium Red on the Activation Curve of BKCaChannels.
Figure 4C shows the activation curve of BKCa channels in the absence and presence of ruthenium red. The relationships between membrane potentials and opening probability of BKCa channels with or without the application of ruthenium red (5 and 50 μM) were plotted and filled by the Boltzmann equation using a nonlinear regression analysis
Effect of Ruthenium Red on Kinetic Behavior of BKCaChannels in GH3 Cells.
The effect of ruthenium red on open and closed time of BKCa channels was examined and analyzed during recordings from patches showing only single-channel openings. As shown in Fig.5, in control cells (i.e., in the absence of ruthenium red), the open-time histogram of BKCa channels at +40 mV can be fitted by a single-exponential curve with a mean open time of 3.3 ± 0.4 ms (n = 6), whereas the closed-time histogram was fitted by a two-exponential curve with a mean closed time of 2.9 ± 0.3 and 38.1 ± 1.4 msec (n = 6). The presence of ruthenium red decreased the lifetime of the open state to 1.6 ± 0.2 ms (n = 5) and increased the mean closed time to 7.3 ± 0.2 and 72.2 ± 8.2 msec (n = 5). Thus, it is clear that the inhibitory effect of ruthenium red on BKCa channel activity is due to both a decrease in open time and an increase in closed time.

Effect of ruthenium red on mean open (left) and closed time (right) of BKCa channel in GH3cells. Under symmetrical K+ condition, the cells were held at +40 mV in inside-out configuration. Open-time histogram in control was fitted by a single exponential function with a mean open time of 3.3 ms, whereas closed-time histogram in control was fitted by a sum of two-exponential function with a mean closed time of 2.9 and 38.1 ms, respectively. The open time (upper) and closed-time (lower) histograms after the addition of ruthenium red (10 μM) into the bath are shown in the lower part. The mean open time was decreased to 1.6 ms, whereas the mean closed time was increased to 7.3 and 72.2 ms. Of note, the abscissa and ordinate show the logarithm of the open or closed time (msec) and the square root of the number of events (n1/2), respectively. The solid curve indicates single- or two-exponential fitting using the least-squares method. In the control, data were obtained from a measurement of 984 channel openings with a total record time of 30 s. In the presence of ruthenium red, data were measured from 895 channel openings with a total record time of 2 min. Bin width for the open- and closed-time histograms is 0.5 and 3 ms, respectively.
Lack of Effect of Ruthenium Red on Voltage-Dependent K+Outward Current (IK) in GH3 Cells.
To determine whether ruthenium red also affects the amplitude of IK in these cells, the experiments were conducted in cells bathed in Ca2+-free Tyrode’s solution containing 1 μM tetrodotoxin and 0.5 mM CdCl2. When the cell was held at −60 mV and various potentials ranging from −70 to +70 mV were applied, no significant difference in current amplitudes at each voltage step tested between the absence and presence of ruthenium red (50 μM) can be demonstrated (Fig.6). Similar results were obtained in seven different cells. Thus, these results indicate that ruthenium red did not affect the amplitude of IK in GH3 cells.

Lack of effect of ruthenium red on voltage-dependent K+ outward currents in GH3 cells. The cells, bathed in Ca2+-free Tyrode’s solution containing tetrodotoxin (1 μM) and CdCl2 (0.5 mM), were held at the level of −60 mV, and voltage pulses from −70 to +70 mV in 20-mV increments were applied. Superimposed current traces shown in the upper part are control, and those in lower part were recorded 2 min after addition of ruthenium red (50 μM). Voltage protocols are shown in uppermost part. Arrows indicate 0 current level.
Inhibitory Effect of Ruthenium Red on Voltage-Dependent L-Type Ca2+ Current (ICa,L) in GH3Cells.
The effect of ruthenium red on ICa,Lwas also examined. The experiments were conducted with the Cs+-containing pipette solution. As shown in Fig.7A, the cell was held at −50 mV, and the depolarizing pulses (300 ms in duration) to 0 mV were delivered at 0.1 Hz. Ruthenium red can inhibit the amplitude of ICa,L in a concentration-dependent manner. When the cells were depolarized from −50 to 0 mV, the amplitude of ICa,L was significantly decreased by ruthenium red (10 μM) to 52 ± 5 pA from a control value of 227 ± 12 pA (n = 8). In addition, the current-voltage relation of ICa,L was unaffected by ruthenium red (Fig.7B). These results indicate that ruthenium red also suppressed the amplitude of ICa,L.

Effect of ruthenium red on ICa,L in GH3 cells. The patch pipette was filled with Cs+-containing solution, and cells were bathed in normal Tyrode’s solution containing 1.8 mM CaCl2, 1 μM tetrodotoxin, and 10 mM tetraethylammonium chloride. A, superimposed current traces when the cell was depolarized from −50 to 0 mV with a duration of 300 ms. a, control. b, c, and d were obtained 1 min after the application of 3, 10, and 30 μM ruthenium red, respectively. Arrow indicates the 0 current level. B, the averaged current-voltage relations for ICa,L in the absence (●) and presence (○) of ruthenium red (10 μM) (mean ± S.E.;n = 8 for each point).
Discussion
The major findings of this study were as follows: 1) in rat pituitary GH3 cells, ruthenium red, an organometallic dye, suppressed IK(Ca); 2) ruthenium red decreases the activity of BKCachannels but does not change single-channel conductance; 3) ruthenium red shifts the activation curve of BKCa channels to more positive potentials; 4) ruthenium red-induced block in BKCa channels can be reversed by an increase in intracellular Ca2+ concentration; and 5) ruthenium red inhibits voltage-dependent ICa,L. These findings suggest that ruthenium red-induced decrease in the activity of BKCachannels and that inhibition of ICa,L may contribute to its modulatory effects on neuronal or neuroendocrine function.
Previous reports have demonstrated that ruthenium red can affect Ca2+ release from mitochondria (Broekemeier et al., 1994; Gunter et al., 1998; Wood and Gillespie, 1998). However, the present study shows that in GH3 cells, neither CCCP (10 μM), an uncoupler of oxidative phosphorylation (Hehl et al., 1996), nor cyclosporin A, an inhibitor of mitochondrial permeability transition pore (Shinohara et al., 1998), can produce any effect on the amplitude of IK(Ca). Additionally, our results from single-channel experiments with an excised membrane patch led to the interpretation that the inhibitory effect of ruthenium red on BKCa channels is mainly due to the result of direct binding to the inner surface of channel. Thus, these results make it unlikely that the effect of ruthenium red on mitochondrial function in these cells is directly responsible for the reduced activity of BKCa channels, although the two agents used to differentiate among potential sites of action of ruthenium red are not specific.
The IC50 value for ruthenium red-induced inhibition of BKCa channels in GH3 cells was 15 μM. This value is higher than that which produced the inhibition of BKCachannels in bladder smooth myocytes, with an IC50value of 5 μM (Hirano et al., 1998), but similar to those required to inhibit mitochondrial Ca2+-induced Ca2+ release (Broekemeier et al., 1994; Gunter et al., 1998; Wood and Gillespie, 1998), to block vanilloid receptor (Staszewska-Woolley and Woolley, 1991; Lo et al., 1997), and to inhibit Ca2+ release from endoplasmic reticulum (Cheek et al., 1991; Sah and McLachlan, 1991; Otun et al., 1996). Therefore, BKCa channels are likely to be a relevant “target” for the action of ruthenium red, although it remains to be determined whether this effect occurs in other types of cells.
Ruthenium red had no effect on single-channel conductance of BKCa channels in GH3 cells (Fig. 4). Thus, the reduction in the conductance of whole-cell current shown in Fig. 1 must be due to a decrease in channel-open probability. In addition, ruthenium red also caused a shift in the midpoint for voltage-dependent opening. Thus, it is clear that ruthenium red can suppress the activity of BKCa channels in a voltage-dependent fashion.
Previous reports showed that ruthenium red can interact with the negative charges of the sialic acid moieties, thus disturbing synaptic transmission in rat hippocampal slices (Wieraszko, 1986). The reflex effects caused by capsaicin can be reversed by treatment with ruthenium red (Wang and Hakanson, 1993; Lo et al., 1997) or neuraminidase, an enzyme that cleaves sialic acid residues from glycosides and sialoglycoproteins (Staszewska-Woolley and Woolley, 1991). However, in our study, the inhibition of IK(Ca) caused by ruthenium red was not affected by pretreatment of cells with neuraminidase (1 U/ml) (data not shown). Thus, it is unlikely that the blockade of BKCa channels by ruthenium red is due to its binding to sialic acid residues. In fact, a recent study showing that ruthenium red-induced neurotoxicity cannot be prevented by treatment with neuraminidase (Velasco and Tapia, 1997) appears to be compatible with our findings. Furthermore, the inhibitory effects of ruthenium red on BKCa channels cannot be due to its antagonistic action on vanilloid receptors because GH3 cells do not express these receptors (Wu et al., 1998a).
The present results appear to be inconsistent with a previous report in which it was found that ruthenium red can increase both IK and BKCa at mouse motor nerve terminals (Lin and Lin-Shiau, 1996). However, several studies, including the present results, showed that ruthenium red can suppress the activity of BKCa channels in hippocampal neurons (Wann and Richards, 1994) and smooth myocytes (Duridanova et al., 1996; Hirano et al., 1998). Alternatively, the activity of BKCa channels was reported to affect presynaptic Ca2+ signals and transmitter release from nerve terminals (Robitaille and Charlton, 1992). This discrepancy is currently unexplained, but it may be related to different channel isoforms in various tissues as reported (Tseng-Crank et al., 1994). However, our work showed that ruthenium red did not affect the amplitude of voltage-dependent K+ current, but did inhibit voltage-dependent L-type Ca2+ current. The inhibitory effect of ruthenium red on Ca2+ current is consistent with previous reports observed in chromaffin cells (Gomis et al., 1994), mouse sensory neurons (Duchen, 1992), and synaptosomes and neuromuscular preparations (Hamilton and Lundy, 1995; Tapia and Velasco, 1997).
One interesting finding is the antagonism by an increase in internal Ca2+ of ruthenium red-mediated inhibition of BKCa channels. This property distinguishes ruthenium red from some other types of channel blockers that act as open channel or use-dependent blockers (Wu et al., 1997). For such drugs, block is increased as the channel opening is promoted. The functional antagonism between the effect of internal Ca2+ and ruthenium red on BKCa channels may be interpreted to mean that these two molecules interact to bind allosterically. This notion is based on the findings showing the rapid relief of ruthenium red block by elevated Ca2+ (Fig. 3), as compared with the slow decrease in the channel activity during the exposure to ruthenium red and slow recovery from the inhibition of ruthenium red during washout.
In summary, the results of our study provide substantial evidence that in addition to suppressing L-type Ca2+ current, ruthenium red can also suppress the activity of BKCa channels in GH3 cells. Ruthenium red may prove to be a useful tool for learning about BKCa channels at both molecular and macroscopic levels. These compounds are small molecules that can bind to BKCa channels at sites that are important for channel gating and are coupled to Ca2+ binding sites. They may also give insights into the role of BKCa channels in cell and tissue function.
Footnotes
-
Send reprint requests to: Dr. Sheng-Nan Wu, Department of Medical Education and Research, Veterans General Hospital-Kaohsiung, 386, Ta-Chung 1st Rd., Kaohsiung City, Taiwan, Republic of China. E-mail: snwu{at}isca.vghks.gov.tw
-
↵1 This study was aided by grants from the National Science Council (NSC-87-2341-B075B-013) and Veterans General Hospital-Kaohsiung (VGHNSU-87-06 and VGHKS-88-31), Taiwan, ROC.
- Abbreviations:
- BKCa channel
- large-conductance Ca2+-activated K+ channel
- IK(Ca)
- Ca2+-activated K+ current
- ICa,L
- voltage-dependent L-type Ca2+ current
- IK
- voltage-dependent K+ current
- CCCP
- carbonyl cyanidem-chlorophenyl hydrazone
- ruthenium red
- trans-tetradecaamine di-μ-oxotriruthenium6+ hexachloride
- Received February 8, 1999.
- Accepted May 3, 1999.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}