


Abstract
The distribution of the binding sites for kaliotoxin (KTX), a blocker of voltage-dependent K+ channels, was studied with quantitative autoradiography in adult rat brain and during postnatal brain maturation. Iodinated KTX bound specifically to tissue sections with a high affinity (Kd = 82 pM) and a maximal binding capacity of 13.4 fmol/mg protein. The distribution of KTX binding sites within the central nervous system was heterogeneous. The highest densities were found in the neocortex, hypothalamus, dentate gyrus, bed nucleus of the stria terminalis, and parabrachial nuclei. The lowest level was observed in the white matter. From postnatal day 5 onward, KTX binding sites were detectable only in the hindbrain. The density of KTX binding sites in whole brain drastically increased after postnatal day 15 to achieve adult levels at postnatal day 60 in the whole brain. Bath application of KTX to Xenopus laevis oocytes blocked recombinant Kv1.3 and Kv1.1 channels potently and Kv1.2 channels less potently, with respectiveKd values of 0.1, 1.5, and 25 nM. KTX affinities for each of these channels expressed in mammalian cells were about 10-fold lower. A comparison of the distribution of KTX binding sites with that of Kv1 channel polypeptides, together with the pharmacology of KTX block, suggests that the principal targets for KTX in rat brain are K+ channels containing Kv1.1 and Kv1.3 α-subunits.
Excitable cells express voltage-gated potassium (Kv) channels. These membrane proteins regulate the resting membrane potential, control the firing pattern, and modulate the neurotransmitter release. This diversity of function is reflected in the heterogeneity of Kv channel genes and their sensitivities to various blockers. Based on their amino acid sequences, the α subunits of Kv channels have been classified into nine families, Kv1 to Kv9. Each family has several members, and the Shaker (Kv1) family includes nine cloned genes, Kv1.1 to Kv1.9 (Stühmer et al., 1989; Pongs, 1992; Chandy and Gutman, 1995). Kv channels expressed in vivo appear to be homotetramers or heterotetramers of α-subunits, sometimes but not always associated with regulatory β-subunits (Sheng et al.; 1993, Rhodes et al., 1997;Shamotienko et al., 1997). Such combinatorial associations increase greatly the Kv channel diversity.
As a consequence, our knowledge on the function of a given Kv channel in situ is presently restricted by several experimental limitations; these include the similarities in electrophysiological properties among some α subunits and difficulties in determination of in situ Kv channel subunit composition (Robertson, 1997). One approach to this problem is the autoradiographic detection of tissue binding sites of a radiolabeled toxin of well-defined Kv α-subunit specificity. For example, the distribution in mammalian brain of the binding sites of radiolabeled α-dendrotoxin (α-DTX) correlated with the localization of Kv1.1, Kv1.2, and Kv1.6 subunits (Shamotienko et al., 1997) and that of radiolabeled margatoxin (MgTX) with Kv1.2 and Kv1.3 subunits (Grissmer et al., 1994; Scott et al., 1994; Koch et al., 1997;Shamotienko et al., 1997).
Kaliotoxin (KTX) is a 38-amino-acid residue toxin purified from the venom of the Androctonus mauritanicus mauritanicus scorpion (Crest et al., 1992). KTX belongs to a toxin family that includes three agitoxins (Garcia et al., 1994), Buthus martensi toxin (Romi-Lebrun et al., 1997), and KTX2 (Laraba-Djebari et al., 1994). Agitoxin 2 blocks Kv1.1, Kv1.2, Kv1.3, and Kv1.6 channels, and B. martensi toxin blocks Kv1.3 channels. We have previously characterized KTX as a blocker of Ca2+-activated K+ channels of intermediate conductance (IKCa) in Helix neurons (Crest et al., 1992). Because Kv channels in these neurons were found to be insensitive to KTX, we proposed KTX as an IKCachannel blocker. In addition, because charybotoxin (CTX) had been shown to block both Helix IKCa channels (Hermann and Erxleben, 1987) and mammalian skeletal muscle high conductance Ca2+-activated K+ (BK) channels (Miller, 1995), we initially hypothesized that KTX might block BK-type channels.
Later studies demonstrated, however, that KTX competes with DTX and mast cell-degranulating (MCD) peptide in rat brain synaptosomes (Romi et al., 1993; Laraba-Djebari et al., 1994). Grissmer et al. (1994) then showed that KTX blocks Kv1.3 and Kv1.1 channels, stably expressed in mammalian cell lines. The KTX binding site on the exofacial surface of Kv1.3 was later mapped by thermodynamic coupling analysis with mutant toxins and channels (Aiyar et al., 1996). Although Grissmer et al. (1994) found Kv1.2 channels expressed in mammalian cells insensitive to KTX, Hopkins et al. (1996) reported that KTX blocks Kv1.2 channels expressed in Xenopus laevis oocytes at nanomolar concentrations.
The present study was undertaken to reevaluate the KTX sensitivity of selected mammalian Kv1 channels and then to identify KTX binding sites in adult rat brain. The results demonstrate that KTX potently blocks Kv1.1 and Kv1.3 channels but more weakly blocks Kv1.2. The distribution of KTX receptors in rat brain is heterogeneous, with high densities in neocortex, hypothalamus, and some hindbrain nuclei. KTX binding sites are detectable only after postnatal day 5 (P5) and increase to adult levels by P60.
Materials and Methods
Heterologous Expression of Kv Channels
Expression in Oocytes.
Mouse Kv1.1, Kv1.3, and Kv3.1 cDNAs were gifts from K. G. Chandy (University of California, Irvine, CA). Kv1.1 was subcloned into pXT7, a modified Bluescript vector bracketing the insert with 5′- and 3′-noncoding sequences derived fromX. laevis β-globin. cRNA was transcribed with T7 RNA polymerase (Megascript Kit; Ambion, Austin, TX) fromPstI-linearized plasmid template. Kv1.3 was subcloned into pSP64T, another X. laevis β-globin noncoding region vector, and cRNA was transcribed with SP6 RNA polymerase fromEcoRI-linearized plasmid template. Kv3.1, subcloned into pBluescript vector with a poly(A)+ tail, was transcribed with T3 RNA polymerase from SacI-linearized plasmid template. Rat Kv1.2 cDNA (a gift from E. Peralta, Harvard University, Boston, MA) was transcribed with T3 RNA polymerase fromXbaI- or NotI-linearized plasmid template. cRNAs were stored at −20°C in water at 1 μg/ml. cRNAs were injected into X. laevis oocytes in 50-nl volumes at concentrations of 4 to 10 ng/ml, and currents were recorded 1 to 4 days after injection.
Expression in Mammalian Cells.
Human embryonic kidney cells stably transfected with human Kv1.1 and Kv1.2 cDNAs in pcDNA3 were kindly provided by O. Pongs (Zentrum für Molekulare Neurobiologie, Universität Hamburg, Hamburg, Germany). Cells were maintained in Dulbecco's modified Eagle's medium/Ham's F-12 (BioMedia, Boussens, France) supplemented with 10% FCS, 50 μg/ml streptomycin, 50 U/ml penicillin, and 4 mM l-glutamine (GIBCO BRL Life Technologies, Cergy-Pontoise, France).
Jurkat JH 6.2 T cells constitutively expressing Kv1.3 (kindly provided by E. Beraud-Juven, Laboratoire d'Immunologie, Faculté de Médecine, Marseille, France) were maintained in Dulbecco's modified Eagle's medium supplemented with 10% FCS and 100 μg/ml streptomycin, 100 U/ml penicillin (GIBCO BRL Life Technologies). Cells were plated onto Nunc (Roskilde, Denmark) dishes 3 days before use in electrophysiological experiments.
Electrophysiological Recordings
Voltage-Clamp Experiments in Oocytes.
Two-electrode voltage-clamp recordings were performed using a Gene Clamp 500 amplifier (Axon Instruments, Burlingame, CA). Oocytes were bathed in calcium-free saline containing 88 mM NaCl, 1 mM KCl, 0.8 mM MgSO4, 2.4 mM Na2CO3, and 10 mM HEPES, pH 7.4. Intracellular electrodes were filled with 3 M KCl. The holding potential was set at −80 mV. The perfusion system, designed to deliver a fast change in toxin concentration, was as previously described (Cotton et al., 1997). KTX was synthesized by J. van Rietschoten (CNRS UMR 6560, Marseille), and the steady-state effects of KTX were determined after 15 min of treatment.
Patch-Clamp Experiments in Mammalian Cells.
Whole-cell recordings were performed on cells bathed in 137 mM NaCl, 5.3 mM KCl, 0.8 mM MgSO4, 0.4 mM Na2HPO4, 1.8 mM CaCl2, and 10 mM HEPES, pH 7.4. Patch pipettes were pulled on a P87 Sutter puller (Novato, CA) from borosilicate glass capillaries (Clark Instruments, Pangbourne, UK) and filled with 130 mM KCl, 2 mM MgCl2, 10 mM EGTA, and 10 mM HEPES, pH 7.4. Holding potential was set at −80 mV. KTX was delivered to the bath via pneumatic picopump system (WPI, Aston, UK) by the application of pressure to a broken patch pipette.
Whole-Cell Current Analysis
Currents from voltage- and patch-clamp experiments were sampled at 2 kHz. Software for stimulation, acquisition, and analysis was custom-written by H. Chagneux (CNRS UPR 9024). Dose-response curves were determined by the successive addition of KTX at increasing concentrations. Each point was the mean ± S.E. of three to nine experiments. Experimental points were fitted to the theoretical hyperbolic curve: y = 1/[1 + ([T]/IC50)n], wherey is the fraction of unblocked current, IC50 is the concentration of toxin inducing 50% block, and n is the Hill coefficient corresponding to the number of molecules required to block one channel. Whenn = 1, the interaction KTX/channel is bimolecular, and IC50 values corresponds to the dissociation constant (Kd).
Animals and Tissue Preparation
Adult male Sprague-Dawley rats weighed 150 to 200 g. Postnatal rats were raised in litters of five pups of either sex, and the day of birth was designated P0. The minimum number of rats used in each experiment at each postnatal stage (P0, P3, P5, P7, P9, P15, P21, P60) was three infants from three different Sprague-Dawley females. They were given food and water ad libitum at constant room temperature under a 12-h light/dark cycle.
Anesthetized rats were sacrificed by decapitation, and their brains were immediately removed and frozen in isopentane at −40°C. Coronal or sagittal cryostat sections (15 μm) were collected, thaw-mounted onto cold chrom-alum/gelatin-coated glass slides, and stored at −60°C until used.
Binding and Autoradiographic Procedures
The binding of KTX to Kv channel proteins was performed on tissue sections using a highly radioactive KTX synthesized and radiolabeled using 125I as previously described (Romi et al., 1993). The brain sections were incubated with 1 to 200 pM125I-KTX (for binding studies) or with 20 pM125I-KTX (for autoradiographic procedures), at 4°C in a 20 mM Tris-Cl buffer, pH 8, containing 50 mM NaCl and 1% BSA. The nonspecific binding component was measured by the addition of a large excess of native KTX (0.2 mM) 30 min before the addition of125I-KTX. After a 60-min incubation, the sections were rinsed four times (with each wash lasting 10 s) in a 20 mM Tris-Cl buffer, pH 8, containing 150 mM NaCl and 1% BSA and then rinsed once in the same buffer containing 0.1% BSA.
For biochemical studies, a portion of the labeled brain sections was removed. Radioactivity was measured with a Packard spectrometer (Crystal II multidetector system). Protein was determined according toBradford (1976). Equilibrium binding constants were determined by Scatchard plot analysis, and linear regression was performed to obtain equilibrium dissociation constant (Kd) and maximal receptor concentration (Bmax).Kd and Bmaxvalues (mean ± S.E.) were obtained from three independent experiments.
For autoradiographic investigations, the slices were dried with a stream of cold air and exposed to Kodak BioMax MR film. After a 7-day exposure, the films were processed in Kodak Industrex developer at room temperature for 2 min, fixed, and then washed. Azur II-stained sections were used for reference. The autoradiograms were analyzed and quantified using NIH Image software. Plastic standards (Amersham Corp., Paisley, UK) were used to calibrate 125I concentrations. Receptor densities were expressed in fmol/mg protein. A mean receptor density value for each nucleus was calculated from six to eight bilateral measurements in three animals. The specific binding value was determined as the difference between total and nonspecific binding components for a given area. Rat brain regions were identified and named according to the rat brain atlas of Paxinos and Watson (1986).
Results
Kv1.1, Kv1.2, and Kv1.3 Channels Are Blocked by KTX.
Figure1 shows the reversible inhibitory effect of various concentrations of KTX (1–50 nM) on currents in X. laevis oocytes expressing either Kv1.1, Kv1.2, and Kv1.3. Block by KTX was voltage independent. Even at 1 μM, KTX had no effect on Kv3.1 channel (not illustrated). The relative blocking potency was determined by measurement of current remaining after stepwise increases in KTX concentration (Fig. 2). Experimental points (mean ± S.E.) of three to nine experiments were fitted to hyperbolic curves. For Kv1.3 channel, we found an IC50 value of 0.1 ± 0.04 nM and a Hill coefficient value of 1.08 ± 0.1. The corresponding values for Kv1.1 and Kv1.2 channels were 1.1 ± 0.2 and 25 ± 1.5 nM for IC50 and 1.03 ± 0.4 and 0.9 ± 0.1 for the Hill coefficient. Because the Hill coefficient was very close to 1, these results confirm that KTX blocks Kv channels via a bimolecular mechanism. Therefore, the IC50 values were considered dissociation constants (Kd).

KTX blockade of Kv1.1, Kv1.2, and Kv1.3 channels expressed in X. laevis oocytes. Superimposed Kv1.1, Kv1.2, and Kv1.3 currents induced by 400-ms depolarizing pulses at the indicated voltages from a holding potential of −80 mV. Leak and capacitative currents were subtracted using a scaled −50 mV hyperpolarizing pulse. The currents were reduced by 76, 82, and 86% after the application of 5, 50, and 1 nM KTX, respectively. Current block by KTX was reversible and voltage independent.
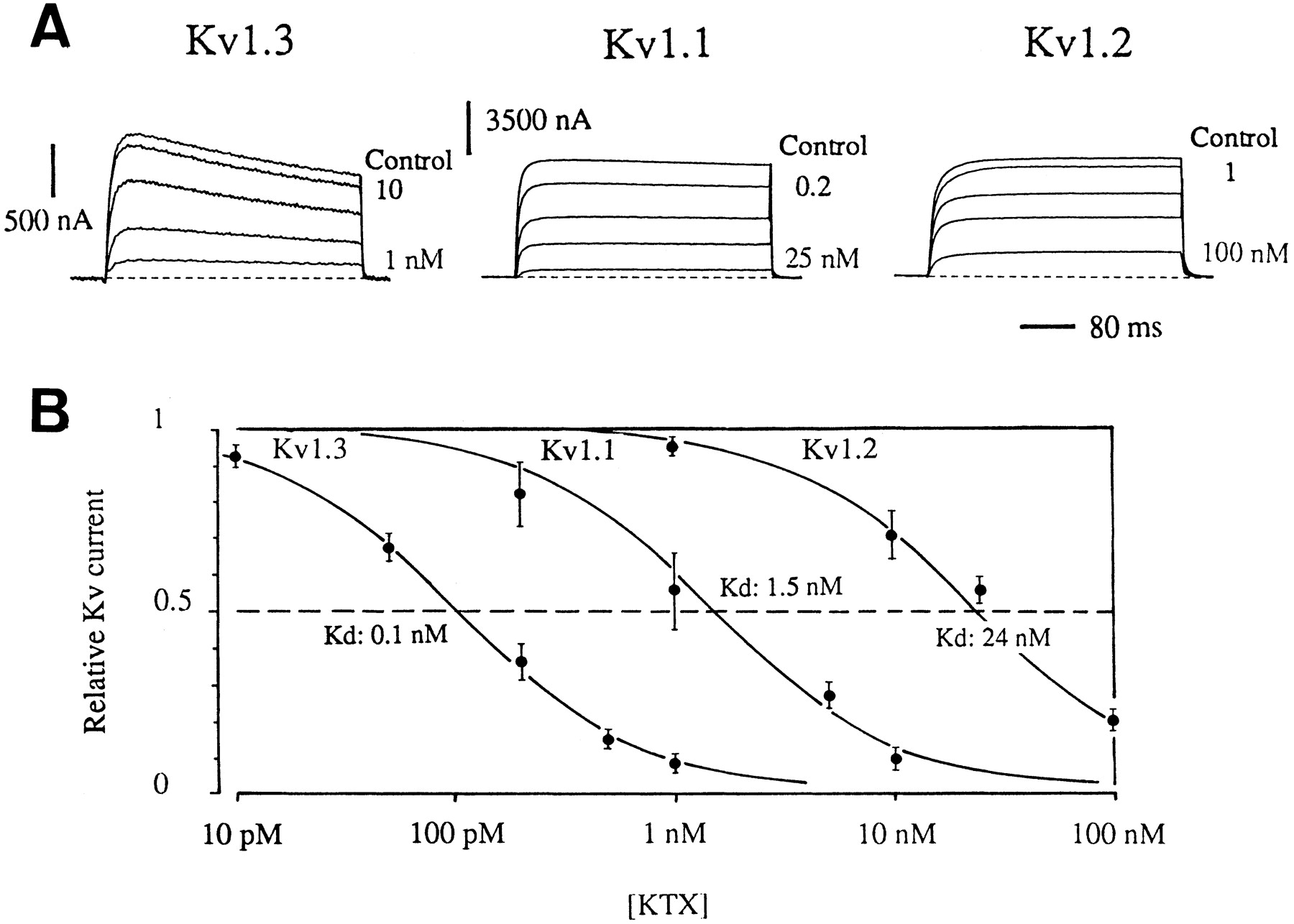
Dose-response of KTX blockade in X. laevis oocytes expressing Kv1.1, Kv1.2, and Kv1.3 channels. A, cumulative current block induced by increasing KTX concentrations. Current was elicited at a test potential of +10 mV from a holding potential of −80 mV. Each trace shows the current recorded under steady-state condition. B, KTX dose-response curves. Points represent mean ± S.E. of four to nine separate experiments. The continuous curves are best fits to Hill functions, with a Hill coefficient of 1.08 ± 0.1 for Kv1.3, 1.03 ± 0.4 for Kv1.1, and 0.9 ± 0.1 for Kv1.2.
Several previous studies have concluded that toxin affinity determined with electrophysiological methods depends on the system in which the ionic channels are expressed (for a review, see Robertson et al., 1997). To estimate the Kd value of KTX for Kv1.1, Kv1.2, and Kv1.3 channels expressed in mammalian cells, we used human embryonic kidney 293 cells stably expressing human Kv1.1 and Kv1.2 genes and Jurkat cells constitutively expressing the Kv1.3 channel. Figure 3A shows the percentage of Kv1.3, Kv1.1, and Kv1.2 currents blocked at various voltages by 2, 100, and 400 nM KTX, respectively. To estimate precisely any shift in affinity compared with the oocyte data, we performed dose-response experiments with various KTX concentrations (Fig. 3B). TheKd values measured were 2, 14, and 270 nM for Kv1.3, Kv1.1, and Kv1.2, respectively, and the Hill coefficient for each was very close to 1. These results indicate that KTX affinity for Kv1.1, Kv1.2, and Kv1.3 channels expressed in mammalian cells was 10- to 20-fold lower than that in X. laevis oocytes.

Dose-response of KTX blockade in mammalian cells expressing Kv1.1, Kv1.2, and Kv1.3 channels. A, Kv1.3 currents were recorded in Jurkat cells. Kv1.1 and Kv1.2 currents were recorded in human embryonic kidney 293 cells stably transfected with the corresponding human cDNAs. Shown are superimposed currents resulting from 80-ms depolarizing pulses from −40 to 30 mV (in 10-mV incremental steps for Kv1.3 and Kv1.2 and in 20-mV incremental steps for Kv1.1). Current block was induced by adding 2 nM (Kv1.3), 100 nM (Kv1.1), and 400 nM KTX (Kv1.2). B, KTX dose-response curves. Points are mean ± S.E. of three to seven separate experiments. The continuous curves are best fits to Hill functions, with Hill coefficients of 1.01 ± 0.2 for Kv1.3, 1.03 ± 0.2 for Kv1.1, and 0.93 ± 0.1 for Kv1.2.
To determine the consequence of the KTX iodination, we tested the blocking potency of 125I-KTX on Kv1.1, Kv1.2, and Kv1.3 channels expressed in X. laevis oocytes. TheKd value decreased 4-fold for Kv1.1 and Kv1.3 and 3-fold for Kv1.2 (not shown). Similar reductions in potency on erythroid Ca2+-activated K+ channel activity have been reported as a consequence of radioiodination of CTX (Brugnara et al., 1993).
Binding Assays of KTX in Adult Rat Brain Sections.
The binding data are presented in Fig. 4. Tissue sections were incubated in Tris-Cl buffer with increasing concentration of 125I-KTX until equilibrium was achieved. The labeled toxin bound in a concentration-dependent manner. The specific125I-KTX binding was found to be saturable and of high affinity, in the range of studied concentrations. Scatchard plot analysis was consistent with the presence of a single class of noninteracting binding sites, with a Kdvalue for the 125I-KTX-receptor complex of 82 ± 17 pM and a Bmax value of 13.4 ± 1.3 fmol/mg protein.

Equilibrium binding of 125I-KTX to rat brain sections. Serial sections were incubated in the presence of increasing concentrations (1–200 pM) of 125I-KTX. Nonspecific binding was determined in the presence of 0.2 μM unlabeled KTX. Specific binding (▪) was assessed from the difference between total (•) and nonspecific binding (▴). Inset, Scatchard plot of the data. A Kd value (mean ± S.E.) of 82 ± 17 pM and aBmax value of 13.4 ± 1.3 fmol/mg protein were obtained from three similar experiments.
Distribution of KTX Binding Sites in Adult Rat.
To minimize nonspecific binding, the concentration of 125I-KTX used in autoradiographic experiments was 20 pM. The nonspecific component of binding was homogeneously distributed in brain regions, and no difference was detectable between gray and white matter (Fig.5J).

Distribution of KTX binding sites in rat brain. Coronal sections were incubated with 125I-KTX (20 pM) in the absence (A–I) or presence (J) of excess unlabeled KTX. The dark areas indicate high grain density, representing high binding site density. Bar, 2 mm. AO, anterior olfactory nucleus; Acb, accumbens nucleus; BST, bed nucleus of the stria terminalis; CPu, caudate putamen; CG, central gray; DG, dentate gyrus; DM, dorsomedial hypothalamic nucleus; DR, dorsal raphe nucleus; Fr, frontal cortex; gr, granular layer of the cerebellar cortex; IC, inferior colliculus; IGr, internal granular layer of the olfactory bulb; IP, interpeduncular nucleus; LPO, lateral preoptic area; Ls, lateral septal nucleus; Me, medial amygdaloid nucleus; mol, molecular layer of the cerebellar cortex; PnC, pontine reticular nucleus, caudal part; Pu, Purkinje cell layer; SNC, substantia nigra, compacta part; SuG, superior colliculus; Tg, tegmental area; Ve, vestibular nuclei.
The values of specific binding presented in Table1 corresponded to about 65% of the total binding in each brain region. Figure 5 shows that the distribution of the 125I-KTX binding sites is heterogeneous, because a 3.5-fold difference was observed between the highest and lowest receptor levels. Nevertheless, 71% of the studied structures exhibited values of specific 125I-KTX binding within ±20% of the mean value for whole rat brain (2.64 fmol/mg protein). These receptor levels were considered as “intermediate” densities, and “high” and “low” levels were defined in relation to the intermediate range.
Distribution of KTX binding sites in rat brain
In the rhinencephalon, intermediate levels of KTX binding sites were observed in the internal granular layer of the olfactory bulb and anterior olfactory nucleus (Fig. 5A). The external plexiform layer presented the lowest density of receptors in brain (Fig. 5B).
In the telencephalon, the neocortex contained only intermediate densities of KTX binding sites. However, the labeling was not homogeneous, with a higher density of receptors in frontal cortex than in cingulate and parietal cortices (Fig. 5, D and E). No marked difference was found among the layers of cortex. In the septal region, the bed nuclei of the stria terminalis showed high concentrations of KTX receptors. The septum and the nuclei of the horizontal and vertical limbs of the diagonal band contained intermediate levels of binding sites (Fig. 5C). The nuclei of basal ganglia presented intermediate to low levels of KTX sites, but in the caudate putamen, the labeling was associated with neuron clusters and not with fiber bundles. In this region, the global densitometric evaluation likely underestimated the densities of neuronal receptors. Most of the amygdaloid nuclei showed high densities of binding sites, particularly in the amygdalohippocampal area. The hippocampal formation presented a heterogeneous distribution of KTX sites. The superior molecular layer of dentate gyrus was the most heavily labeled, and the CA1 field was least heavily labeled. In the CA3 field, a narrow band of high labeling was found in the stratum lucidum, close to the pyramidal layer. No density variation was observed among the different strati of Ammon's horn fields.
In the diencephalon, high levels of KTX receptors were observed in the hypothalamus, particularly in the ventromedial hypothalamic nucleus and the medial preoptic area (Fig. 5E). In contrast, the thalamic nuclei presented low to intermediate binding site densities except in the central medial thalamic nucleus, which contained a high density of KTX binding sites.
KTX binding sites were focally and discreetly distributed in the mesencephalon or midbrain (Fig. 5). A very high level of receptors was found in the central gray. The ventral tegmental area and the interpeduncular nucleus also presented high receptor densities. The superior colliculi showed higher concentrations of KTX binding sites than did the inferior colliculi. The pretectal areas, substantia nigra, and mammillary, mesencephalic, and raphe nuclei contained intermediate densities of binding sites.
In the metencephalon, the parabrachial nuclei and tegmental areas showed very high labeling (Fig. 5, G and H). In contrast, low levels of KTX receptors were found in the pontine and vestibular nuclei. The superior olive and the facial nucleus also exhibited low concentrations. In the cerebellum, the Purkinje layer contained very high densities of binding sites, whereas intermediate levels of KTX binding sites were present in the granular and molecular layers. Low levels were found in cerebellar nuclei. No difference was observed among the cerebellar lobes.
Postnatal Ontogenesis of KTX Binding Sites in Rat Brain.
Table2 presents the mean density of KTX binding sites during the maturation of the rat central nervous system. At birth and up to P3, whole brain was almost devoid of high-affinity KTX receptors. From P5 onward, a distinct increase in KTX binding site density was observed in the midbrain, particularly in the central gray, tegmental nuclei, and hindbrain. From P5 to P7, the binding increased in mid and hindbrain, whereas it was still low in forebrain. From P9 to P13, most binding sites in forebrain increased slowly and then more dramatically by P15 (Fig. 6). At P15, significant contrast remained between elevated labeling of midbrain and lower levels of labeling in forebrain and hindbrain. In many structures, a slight decrease in KTX receptor densities was observed out to P21. Nevertheless, the heterogeneous distribution characteristically observed in adult animals was already present at P21 (Fig. 6).
Distribution of KTX binding sites during the ontogenesis of rat brain
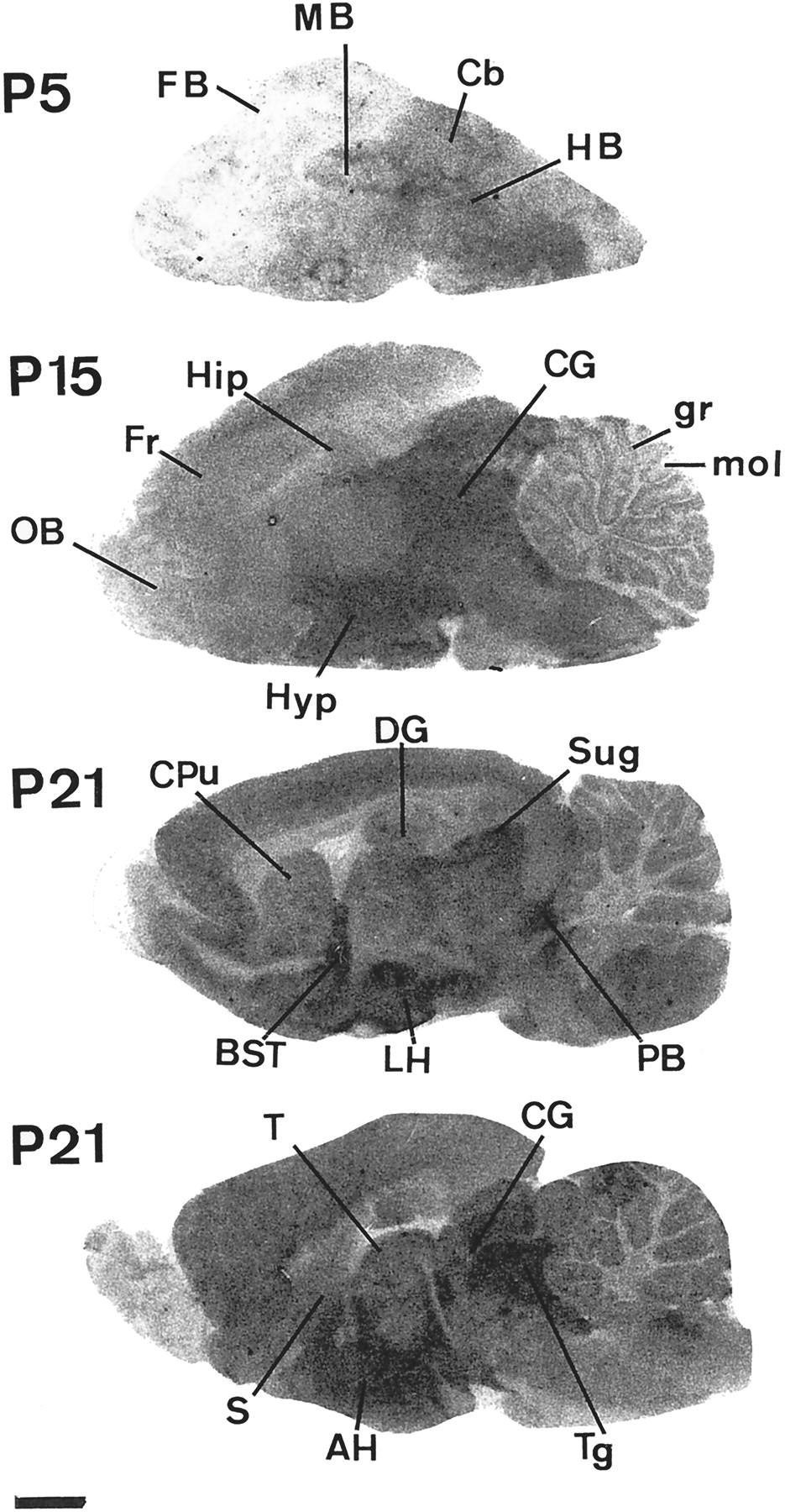
Ontogenesis of KTX binding sites in rat brain. The autoradiograms were obtained under the experimental conditions described in Fig. 5. Bar, 1.6 mm. AH, anterior hypothalamic nucleus; BST, bed nucleus of the stria terminalis; Cb, cerebellum; CG, central gray; CPu, caudate putamen; DG, dentate gyrus; FB, forebrain; Fr, frontal cortex; gr, granular layer of the cerebellar cortex; HB, hindbrain; HIP, hippocampus; Hyp, hypothalamus; LH, lateral hypothalamic nucleus; MB, midbrain; mol, molecular layer of the cerebellar cortex; OB, olfactory bulb; SuG, superior colliculus; PB, parabrachial nucleus; S, septal area; T, thalamus; Tg, tegmental area.
The different portions of the neocortex and the hippocampus presented a similar and regularly progressing evolution of KTX binding site densities extending to the adult stage (Table 2). In the cerebellar cortex, the KTX receptors of the granular layer were detectable from P9 onward. Their concentration increased very quickly between P13 and P15 to reach the adult level by P15. In contrast, the density of KTX binding sites in the molecular layer of the cerebellum steadily increased to the adult stage (Table 2). In most of the brain regions studied, the level of KTX receptors continued to increase until P60 (i.e., adult stage).
Discussion
The pharmacological identification of Kv channel subtypes expressed in rat brain requires the use of blockers with unambiguously defined activity profiles. Toward this goal, we determined that 1) KTX blocks the Kv1.3 and Kv1.1 channels with a high affinity and blocks the Kv1.2 channel with a low affinity, 2) KTX binds to specific receptor sites in the rat brain with a maximal capacity of 13.4 fmol/mg protein, 3) the distribution of KTX binding sites in rat brain is heterogeneous, and 4) KTX receptors appeared during brain ontogenesis only at P5.
KTX Blocks Kv1 Channels in Mammals.
KTX blocks Kv1.1, Kv1.2, and Kv1.3 channels expressed in X. laevis oocytes with subnanomolar to nanomolar affinity. When these channels are expressed in mammalian cell lines, KTX affinity was shifted to the right by one order of magnitude. This result suggests that the binding potency of KTX for Kv channels expressed in situ in rat brain is weak for the channels containing Kv1.2 α-subunits and higher for channels containing Kv1.3 and KV1.1 α-subunits. According to the cell system used to express Kv channels, similar differences in KTX affinity have been reported (Grissmer et al., 1994; Hopkins, 1996). However, the origin of such differences remains undetermined and may result from different levels of channel glycosylation or phosphorylation, inducing, in turn, variability in voltage sensitivity, activation threshold, and gating kinetics (Robertson, 1997). For various Kv1 channels, differences in affinity for ligands like DTX, MCD peptide, CTX, and 4-aminopyridine have also been observed in different heterologous expression systems (Stühmer et al., 1989; Grissmer et al., 1994). In addition, KTX blocks very weakly skeletal muscle BK channel (Kd = 13 μM; not illustrated).
KTX Binding Sites in Rat Brain.
Our results reveal a relatively heterogeneous distribution of KTX binding sites in adult rat brain. The highest levels of KTX receptors were found in the neocortex, bed nuclei of the stria terminalis, most of the hypothalamus, dentate gyrus, central gray, and parabrachial nuclei. The white matter contained the lowest level of KTX binding. The autoradiographic data are in good agreement with biochemical studies with synaptosome preparations. Indeed, for a 20 nM concentration of125I-KTX, the average value for KTX binding site densities determined by autoradiographic procedures was similar to the biochemical values of KTX binding sites measured in brain homogenates (present data) and in P2 synaptosomal membranes (17–22 fmol/mg protein; Martin-Eauclaire, personal communication).
The histological resolution of the autoradiograms does not allow the identification of KTX binding sites as exclusively neuronal. Indeed, these receptors can also be present in glial, endothelial, and arterial smooth muscle cells. With the use of electrophysiological recording, various glial Kv currents have been characterized in tissue slices from the central nervous system (Akopian et al., 1997; Bordey and Sontheimer, 1997). Nevertheless, autoradiographic analysis indicated a very low density of KTX receptors in adult white matter, suggesting very weak KTX binding in oligodendrocytes. However, a contribution to KTX binding from astrocytes cannot be excluded.
Several facts favor the attribution of the measured KTX binding sites to neurons: 1) no published immunological or in situ hybridization study has mentioned the contribution of glial cells to the labeling of Kv channels in brain (Kues and Wunder, 1992; Sheng et al., 1994; Wang et al., 1994; Veh et al., 1995); 2) the Kv1.1 channel is only weakly detected in mature astrocytes, and the Kv1.3 channel was not found in glial cells (Smart et al., 1997; Hallows and Tempel, 1998); 3) the expression of Kv channels is reduced in spinal cord astrocytes cocultured with dorsal root neurons (Thio et al., 1993); and 4) Kv currents decrease 3-fold between P5 and P20 in astrocytes of hippocampal slices (Bordey and Sontheimer, 1997), whereas KTX binding site density increases concomitantly in the same region during brain maturation.
Maximal KTX binding capacity (13.4 fmol/mg protein of brain tissue) is very low compared with values determined for DTX (500–1100 fmol/mg protein; Bidard et al., 1989), MgTX (800–900 fmol/mg protein; Knaus et al., 1995; Koch et al., 1997), and MCD peptide (160 fmol/mg protein;Bidard et al., 1989). Studies of mutual inhibition of125I-KTX binding by DTX and of125I-DTX binding by KTX reveals that KTX recognizes only a small fraction of the DTX binding sites (Harvey et al., 1995).
Electrophysiological data show that DTX blocks Kv1.2, Kv1.1, and Kv1.6, and, with a lower affinity, and Kv1.3 channels. MgTX blocks Kv1.2 and Kv1.3 channels, and MCD peptide blocks Kv1.1, Kv1.6, and, less potently, Kv1.2 (Stühmer et al., 1989; Grissmer et al., 1994). These data suggest that the low maximal binding capacity of KTX may result from its very low affinity for the Kv1.2 channel.
In agreement with this hypothesis, the distribution of KTX binding sites in rat brain was found to be heterogeneous, whereas the density of DTX and MCD peptide binding sites was evenly distributed in gray matter except in hippocampus and cerebellum (Bidard et al., 1987, 1989;Mourre et al., 1988; Awan and Dolly, 1991).
In the hippocampus, the highest levels of DTX binding sites were found in the CA3 field and the dentate gyrus (Bidard et al., 1989), and the highest density of MCD peptide binding sites was found in the stratum lucidum of the CA3 field. The stratum lacunosum moleculare of the CA1 field was poor in density of MCD peptide binding. The KTX binding sites were particularly located in the outer molecular layer of the dentate gyrus and in the stratum lucidum.
In the cerebellum, the KTX receptor levels were similar in the granular and molecular layers with a higher density in the Purkinje layer, whereas the granular and molecular layers, respectively, contained high and moderate levels of DTX and MCD peptide binding sites.
Comparison of Distribution of KTX Binding Sites and Distribution of Kv1.1 and Kv1.3 α Subunits.
In most of the structures analyzed, and particularly in olfactory bulb, hippocampal CA3 subfield, and cerebellum, a comparison of the distribution of Kv1.1 and Kv1.3 α-subunits with the localization of 125I-KTX binding sites suggests that KTX recognizes channels containing these α subunits.
In the olfactory bulb, KTX binding sites and Kv1.1 and Kv1.3 α-subunit polypeptides are strongly expressed in the internal plexiform layer, granular layer, and mitral soma layer, and all are moderately present in the glomerular layer. Moreover, the outer plexiform layer, which is weakly labeled by KTX, shows a strong Kv1.2 immunoreactivity (Veh et al., 1995). In situ hybridization experiments also indicate that high levels of Kv1.3 mRNA are found only in the internal granule layer (Kues and Wunder, 1992).
In the hippocampus, the dentate gyrus contains high densities of Kv1.1 and Kv1.3 mRNAs, and the CA3 field contains a high density of Kv1.1 mRNA. These structures contain high or intermediate densities of KTX binding sites. In both regions, the distribution of immunoreactivity to Kv1.1 and Kv1.3 α-subunits overlaps the distribution of KTX binding sites. However, surprisingly, in CA1 subfield, the density of KTX receptors is low when Kv1.1 immunoreactivity is intense (Wang et al., 1994; Veh et al., 1995; Rhodes et al., 1997).
In cerebellum, as in neocortex, thalamic nuclei, and in many areas of the hindbrain and medulla, Kv1.1 mRNA and Kv1.1 α subunit polypeptide are detected in high or intermediate density (Kues and Wunder, 1992;Wang et al., 1994; Veh et al., 1995). These structures also contain high or intermediate density of KTX binding sites.
These data suggest that in every structure containing intermediate or high densities of KTX binding sites, Kv1.1 or Kv1.3 mRNA (or protein) have been detected. In contrast, in CA1 subfield where the Kv1.1 α-subunit was intensively detected, the KTX binding sites density was weak. It has been demonstrated that the binding of KTX to Kv1 channel requires at least two KTX-sensitive α-subunits (Aiyar et al., 1994). Therefore, one Kv1.1 (or Kv1.3) α-subunit detected with immunohistochemistry may be included in an insensitive KTX channel.
Because in bovine brain Kv1.1 and Kv1.3 have not been described as partners in heterotetrameric Kv channels (Scott et al., 1994;Shamotienko et al., 1997), we hypothesize that KTX receptors are channels containing at least two Kv1.1 or Kv1.3 α-subunits.
Postnatal Development of KTX Binding Sites in Rat Brain.
Very few studies has been performed on the expression and distribution of Kv1 α-subunits during the maturation of rat brain. Hallows and Tempel (1998) reported that Kv1.1 expression in mouse brain is low at birth and increases dramatically at the end of the first postnatal week. These data are consistent with the measured development of KTX binding sites.
In the hippocampal formation, the development of KTX binding sites was different, because the increment in specific KTX autoradiographic labeling was delayed until about P9. Kues and Wunder (1992) showed that Kv1.1 mRNA in this region was present at nearly constant level from P1 to the adult stage. In contrast, Kv1.3 mRNA expression was maximal at P8 and, surprisingly, down-regulated in the CA1 and CA3 fields by P15 to undetectable levels, while remaining constant in the dentate gyrus. We observed that the density of KTX receptors is much higher in dentate gyrus than in CA1 and CA3 fields.
Previous developmental studies with Kv toxins have been performed only with MCD peptide (Mourre et al., 1988). The main difference with KTX is that the MCD peptide receptors are initially present in the forebrain, whereas the KTX binding sites appear first in the hindbrain.
In conclusion, the pharmacological specificity of KTX determined with recombinant Kv channels and the autoradiographically determined distribution of KTX receptors in rat brain should be useful tools for future investigations of channels containing Kv1.3 and Kv1.1 α-subunits. KTX can be used to purify these channels, to discern their roles in shaping neuronal excitability, and to define their involvement in cerebral functions.
Acknowledgments
We thank Michèle André and Hélène Chagneux for technical assistance, Raymond Fayolle for manufacturing the bilayer amplifier, and Régine Romi for help in KTX synthesis.
Footnotes
-
Send reprint requests to: Dr. C. Mourre, Laboratoire de Neurobiologie Intégrative et Adaptative, UMR 6562, CNRS-Université de Provence, Escadrille Normandie-Niemen, Case 351, 13397, Marseille, Cedex 20, France. E-mail:mourre{at}newsup.univ-mrs.fr
-
↵1 This work was supported by the Centre National de la Recherche Scientifique and by the Association Française contre les Myopathies (Grant 5030). M.N. and S.L.A. were supported by NIH Grants HL15157 (Boston Sickle Cell Center) and DK34854 (Harvard Digestive Diseases Center). S.L.A. is an Established Investigator of the American Heart Association.
- Abbreviations:
- Kv
- voltage-gated potassium
- CTX
- charybdotoxin
- DTX
- dendrotoxin
- KTX
- kaliotoxin
- MCD
- mast cell-degranulating
- IKCa
- intermediate conductance Ca2+-activated K+
- MgTX
- margatoxin
- BK
- high conductance Ca2+-activated K+
- Received April 20, 1999.
- Accepted July 27, 1999.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}