


Abstract
In the central nervous system, HIV-1 has a defined tropism for brain macrophages and microglia. Nucleoside analog drugs such as zidovudine improve the clinical and neuropsychological functions in HIV-demented patients. Multiple carrier-mediated transport systems can play an important role in the membrane permeation of nucleosides and nucleoside analog drugs in a number of cells. The purpose of this project was to characterize the uptake properties of the pyrimidine nucleoside probe thymidine by a continuous rat microglia cell line (MLS-9) grown as a monolayer on an impermeable substratum. Approximately 50% of thymidine (10 μM) uptake by the monolayer cells was found to be Na+dependent. Kinetics of specific thymidine uptake showed a single saturation system (Km = 44 μM at 37°C) and a Na+/thymidine stoichiometry of 2:1. Pyrimidine and purine nucleoside probes (50 μM) exerted a competitive inhibitory effect on specific thymidine uptake withKi values of 40, 38, 45, and 39 μM for adenosine, uridine, guanosine, and cytidine, respectively. In addition, nucleoside analog drugs significantly decreased specific thymidine uptake, with IC50 values of 135.1 μM for abacavir and 0.6 μM for zidovudine, which inhibited in a noncompetitive manner. These results suggest that a Na+-dependent nucleoside transport system is present in rat microglia and that long-range interactions between antiretroviral nucleoside analog drugs and the nucleoside substrates may occur at the transporter sites.
HIV not only infects peripheral T lymphocytes and cells of the monocyte-macrophage lineage but also can spread to the central nervous system (CNS) and cause HIV-associated encephalopathy and dementia (Gendelman et al., 1994). The primary brain targets of HIV-1 are microglia and brain macrophages (Price et al., 1988; Watkins et al., 1990; Takahashi et al., 1996). Microglia, first described by Del Rio-Hortega (1932), form a dense network of immune-alert resident “macrophages” that are poised to continuously monitor the CNS microenvironment and respond to any subtle disturbance. Moreover, they are involved in tissue homeostasis, repair, and neuronal regeneration (Thomas et al., 1994; Streit and Kincaid-Colton, 1995; Gehrmann, 1996). Microglia can exhibit ramified, ameboid, or spheroid morphologies, which roughly correlate with their functional status (Streit and Kincaid-Colton, 1995; Gehrmann, 1996). In the healthy adult brain, most are highly ramified with multiple branching processes. However, in response to viral infection, brain injury, or inflammation, microglia become activated, a process that often includes changes in morphology and surface expression of immune-related molecules. In vitro, ameboid cells with short uropod-like processes are typical of mobile proliferating microglia, the spheroid morphology is typical of activated microglia that are primed for phagocytosis, and highly ramified cells are rare.
Nucleic acid (i.e., DNA, RNA) and nucleotide synthesis are intrinsic activities in the adult mammalian brain (Santos et al., 1968). The nucleoside triphosphates necessary for this synthesis may be formed from de novo synthesis by ribonucleotide reductase, or by salvage mechanisms that involve membrane transport of preformed nucleosides and subsequent intracellular phosphorylation. The membrane permeation of nucleosides and nucleoside analog drugs into mammalian cells involves both facilitated diffusive (equilibrative) and concentrative transport mechanisms (Plagemann et al., 1988). Two distinct types of equilibrative nucleoside transporters (NTs) have been identified and termed es, for equilibrative and sensitive to NBMPR [6-(4-nitrobenzyl)-thio-9-β-d-ribofuranosylpurine] andei, for equilibrative and insensitive to NBMPR (Paterson and Cass, 1986; Plagemann et al., 1988). Both systems exhibit broad substrate selectivity for purines and pyrimidines, have been cloned in rats and humans, and their cDNAs were functionally expressed inXenopus oocytes (designated rENT1/hENT1 and rENT2/hENT2 fores and ei, respectively) (Cass et al., 1998).
Concentrative transporters mediate the nucleoside flux against their concentration gradient, by Na+ or K+ cotransport mechanisms. The major types of Na+ nucleoside cotransport systems can be classified based on functional studies, including primarily substrate selectivity. The N1 (or cif) system is selective for purines, with guanosine and formycin B being the commonly used substrates. The N2 (or cit) transporter is selective for pyrimidines, and thymidine is the usual test substrate. Both types have been cloned from humans and rats (hCNT1/rCNT1 and hSPNT1/SPNT for N2 and N1, respectively) and when heterologously expressed, they display a 1:1 Na+/nucleoside-coupling ratio (Cass et al., 1998). The N3 (or cib) transporter has a 2:1 Na+/nucleoside stoichiometry and is broadly selective for both purines and pyrimidines. The fourth type, N4, has similar N2 stoichiometry and selectivity but guanosine and adenosine also are transported. The less characterized N5 (or cs) transporter is NBMPR sensitive and exhibits selectivity for adenosine and formycin B (Cass, 1995).
The widely expressed equilibrative NT systems appear to be less tissue specific than the concentrative systems that have been identified in macrophages (Plagemann and Aran, 1990), choroid plexus (Wu et al., 1992), leukemia cells (Crawford et al., 1990), splenocytes (Darnowski et al., 1987), intestinal cells (Vijayalakshmi and Belt, 1988), and renal and intestinal brush border membrane vesicles (Williams et al., 1989).
The permeation of compounds into and out of the CNS is primarily controlled by the blood brain barrier (BBB) and the blood-cerebrospinal fluid (CSF) barrier. Results from in vivo studies measuring brain uptake suggest that several nucleosides (i.e., adenosine, guanosine, inosine, and uridine) are transported across the BBB by a saturable NT system (Cornford and Oldendorf, 1975). Uptake studies with isolated rat brain capillaries (Wu and Phillis, 1982), isolated bovine cortex capillaries (Stefanovich, 1983) and cerebral endothelial cells (Beck et al., 1983) have identified a carrier-mediated transport for adenosine and other purines. Experiments with thymidine as the probe have shown the presence of NBMPR-sensitive (es) and concentrative (cit/N2) NTs at the BBB of the guinea pig (Thomas and Segal, 1997).
At present, the existence and type of NTs in microglia, the primary target of HIV infection in the CNS, are not known. In the present study, we use radiolabeled thymidine to characterize the uptake kinetics, substrate specificity, and inhibition of saturable NT systems present in rat microglia (MLS-9) grown as a monolayer on an impermeable surface.
Materials and Methods
Cell Culture.
The MLS-9 cell line was prepared as described previously (Zhou et al., 1998). Briefly, essentially pure cultures of microglia were established from brain explants of 2- or 3-day-old Wistar rats with a modified version of our earlier protocol (Schlichter et al., 1996). This initial culture was >98% pure microglia as determined by labeling the microglia with isolectin B4 (Sigma Chemical Co., St. Louis, MO). Thereafter, the weekly feedings were supplemented with supernatant collected from a mouse fibroblast cell line that secretes large amounts of colony stimulating factor 1, a well known stimulus for microglia proliferation. After several weeks in culture, colonies arose which, when harvested, continued to grow in the absence of added growth factor. The MLS-9 cell line, which was established from one of these colonies, continued to display characteristic markers of microglia; i.e., 100% stained with isolectin B4, 98% with OX-42 antibody, and 99% with ED-1 antibody (Zhou et al., 1998). None of the cells were labeled with antibodies directed against glial fibrillary acidic protein or the fibroblast protein fibronectin under conditions that clearly stained astrocytes and fibroblasts in rat brain mixed cultures.
For the present study, MLS-9 cells (passages 25 to 39) were grown as a uniform monolayer on an impermeable substrate: 75-cm2 Falcon plastic tissue culture flasks or 24-well plates (Becton Dickinson & Co., Lincoln Park, NJ) at 37°C, 5% CO2, and 95% humidity. The culture medium (i.e., minimal essential medium, pH 7.2, containingl-glutamine, d-glucose, 5% fetal bovine serum, 5% horse serum, and 0.5% penicillin/streptomycin suspension, all obtained from Life Technologies Laboratories, Grand Island, NY) was changed every 2 days. When confluence was reached, the cells were subcultured by detaching them with a sodium citrate solution containing 130 mM NaCl, 15 mM sodium citrate, 10 mM glucose, and 10 mM HEPES, pH 7.4.
Cell Morphology.
In addition to conventional light microscopy, we examined the cells with scanning and transmission electron microscopy. For transmission electron microscopy studies, monolayers of MLS-9 cells were fixed in situ with 5 ml of 1% glutaraldehyde in 0.1 M PBS (10.4 mM Na2HPO4 · 7H2O, 10.4 mM Na2HPO4, 3.2 mM KH2PO4, and 123.2 mM NaCl) for at least 1 h. Cells were then detached from the Petri dish, dehydrated in graded ethanol, and embedded in Epon according to standard techniques. Ultrathin sections were cut, stained, and examined with a Philips 410 electron microscope. For scanning electron microscopy, standard dehydration and shadowing methods were applied.
Nucleoside Transport.
Nucleoside uptake measurements were performed on uniform, confluent monolayers of MLS-9 cells (passages 25 to 39). No changes in morphology and uptake measurements were observed among the different cell passages used. The confluency stage is observed 4 to 5 days after the cells had been subcultured in 2-cm2, 24-well plates at a cell density of ∼1 × 106 cells/ml. For nucleoside uptake studies, the cells were initially conditioned for 30 min with 0.5 ml of an Earle's balanced saline solution, containing 1.8 mM CaCl2, 5.4 mM KCl, 0.8 mM MgSO4, 138 mM NaCl, 1.0 mM Na2HPO4, 5.5 mMd-glucose, and 20 mM HEPES, with Trizma base added to bring the pH to 7.4. The uptake of [3H]thymidine (65 Ci/mmol; Moravek Biochemicals, Inc., Brea, CA) was determined at 37°C at specific time intervals, with or without cold thymidine. To test for nucleoside specificity, the cells were initially preincubated for 10 to 30 min with a potential inhibitor before adding the incubation media containing both the inhibitor and radiolabeled probe. At the completion of the incubation period, the medium was removed by aspiration and the uptake was terminated by adding an excess (2 ml) of ice-cold 0.16 M NaCl solution. The monolayer of cells was solubilized in 1 ml of 1 N NaOH for 30 min, then transferred to scintillation vials containing 0.5 ml of 2 N HCl. Cellular [3H]thymidine incorporation was measured by a Beckman liquid scintillation counter with automated quench correction. The distribution of the radiolabeled probe in the extracellular space as measured by [14C]d-mannitol (51.5 mCi/mmol; NEN Life Science Products, Boston, MA) was small (<5%); therefore, no correction was applied. The sample counts were corrected for “zero time” uptake and background radioactivity in each individual experiment. Nonspecific uptake was determined by measuring [3H]thymidine uptake in the presence of high concentrations (1 mM) of cold thymidine. Uptake data were standardized to the amount of cell protein (milligrams per milliliter) in each culture plate, as determined by the Bradford colorimetric method with BSA as the standard and BioRad reagent (BioRad, Mississauga, Ontario, Canada). The measured thymidine uptakes were expressed in picomoles per milligram protein per milliliter. The nucleoside analog drugs zidovudine (ZDV), lamivudine (3TC), and abacavir were provided by Glaxo Wellcome (Research Triangle Park, NC). Didanosine (ddI) and zalcitabine (ddC) were provided by Bristol-Myers Squibb (Princeton, NJ) and Hoffmann-La Roche Inc. (Nutley, NJ), respectively. Unless specified, all other chemicals were obtained from Sigma Chemical Co. (St. Louis, MO), and were of the highest purity available.
Nucleoside Metabolism.
Metabolism of thymidine by rat MLS-9 microglia cells was assessed by thin-layer chromatography as described previously (Williams and Jarvis, 1991). Monolayers of MLS-9 cells were incubated for 1 and 10 min with 10 μM [3H]thymidine (100 μCi/ml) at 37°C. The reaction was terminated as described above. Thymine and thymidine were used as the standards. The cell extracts were chromatographed for 4 h on 250-μm thick silica gel-coated plates (silica gel 60; Sigma Chemical Co.) impregnated with a fluorescent indicator. The solvent system consisted of butan-1-ol saturated with water. After drying the plate, the distinct zones bearing the standards (1 mM) were detected and delimited under UV light. The Rfvalues were 0.69, 0.68, and 0 for thymidine, thymine, and thymidine nucleotides, respectively. Radioactivity associated with these zones was determined by standard liquid scintillation counting methods.
Data Analysis.
Each experimental procedure was repeated at least twice in cells pertaining to different passages and each experimental data point in an individual experiment represents quadruplicate trials. The results are reported as means ± S.D. of the number of experiments specified in each figure legend. The kinetic parameters (Km andVmax) and the diffusion coefficientD for thymidine transport were determined by a nonlinear least-squares analysis with the computer program Enzyme Fitter 1.0 (Elsevier-Biosoft, Cambridge, UK). Other equations (seeResults) were fitted with Sigma Plot 4.0. In the stoichiometry experiments, the constantsKNa,Vmax, and n were obtained by least-squares fitting of the data to the hyperbolic Hill equation. To estimate the dissociation constants (Ki values) of the competitive nucleoside inhibitors, a least-squares regression analysis was applied to determine the linear correlation between V andV/[thymidine], the Eadie-Hofstee linear transformation. The IC50 values for inhibition of thymidine uptake by nucleosides and nucleoside analogs were calculated by fitting the data to a sigmoidal, four-parameter equation. The Student'st test for unpaired experimental values and/or the test of repeated measures of ANOVA were used to assess the significance of the inhibition, with P < .05 considered to be statistically significant.
Results
Cell Morphology.
The scanning and transmission electron micrographs (Fig. 1) of the MLS-9 cells show the characteristic surface and cytoplasmic features of microglia cells grown as monolayers on an impermeable surface in our laboratory. Two morphologies were prevalent. Within 2 days after plating, most cells had uropod- or lamellipod-like processes (Fig. 1A), whereas at confluence, most had retracted their processes and displayed spherical, granular cell bodies (Fig. 1B). This micrograph shows the distinctive short surface villi and cytoplasmic features of microglia at high cell density. Large cell body structures with scanty cytoplasm and defined cellular organelles (i.e., numerous mitochondria, vacuoles, enlarged nuclei) can be observed.
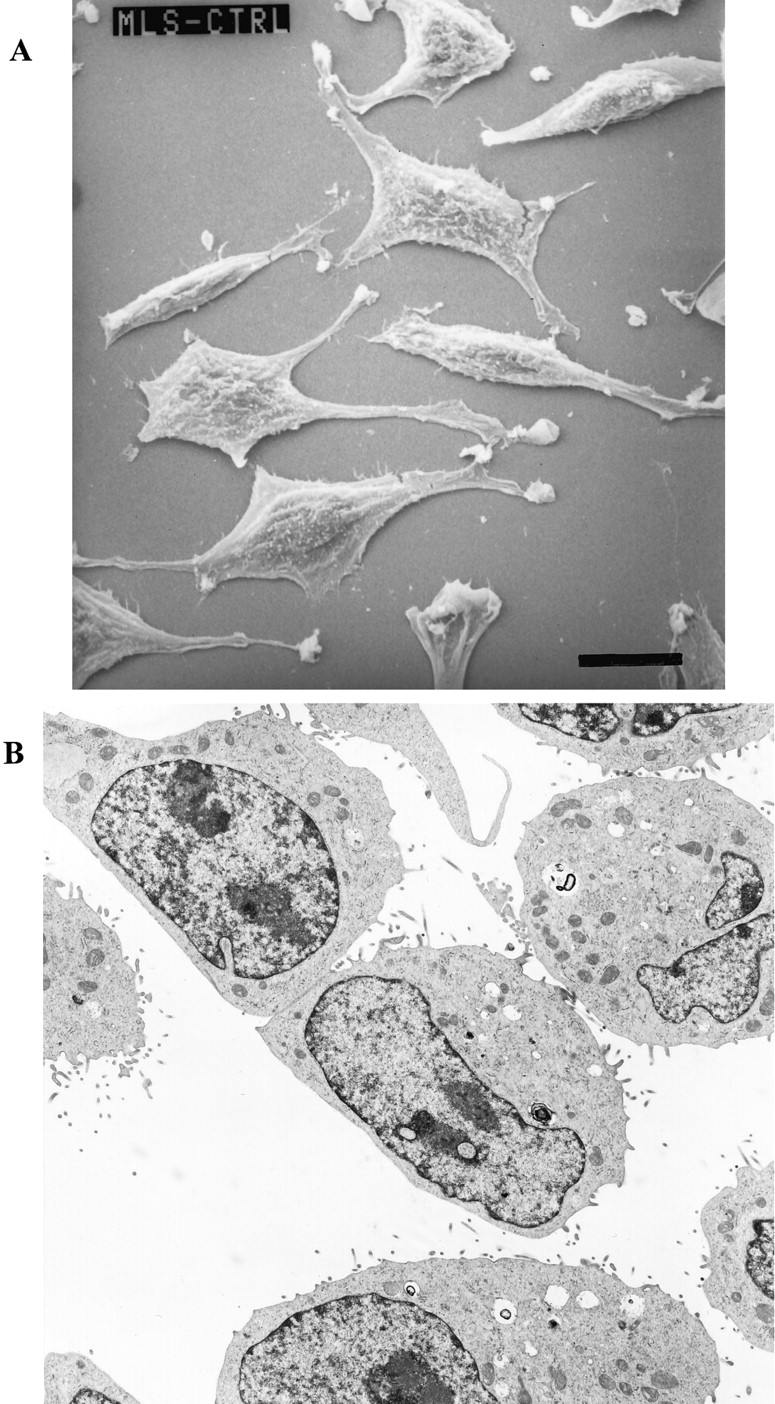
Morphology of MLS-9 cells. A, scanning electron micrograph of cells grown for 2 days. Most cells have an ameboid, elongated morphology with multiple uropods. The membranes show some ruffling and long protrusions, but no deep invaginations. Scale bar, 20 μm. B, transmission electron micrograph of cells grown for 7 days into a confluent monolayer. Most of the cells are rounded up at this time, and display relatively smooth membranes and several short villus-like processes. The cells exhibit large nuclei relative to cell volume, numerous mitochondria, and vacuoles. Original magnification, 5000×.
Thymidine Metabolism.
Intracellular metabolism of nucleosides often complicates and prevents characterization of their membrane transport mechanisms (Plagemann et al., 1988). Therefore, we investigated the extent of thymidine metabolism by microglia cells. From the thin-layer chromatography data, there was no significant metabolism of thymidine within the first minute of incubating MLS-9 cells with 10 μM [3H]thymidine in standard incubation medium. After a 1-min incubation period with thymidine, 88% of the intracellular radioactivity was recovered in the thymidine fraction, 10% cochromatographed with thymidine nucleotides, and the remaining 2% cochromatographed with thymine (data not shown). However, after a 10-min incubation ∼80% of the radioactivity comigrated with the thymidine nucleotides on the silica plate, indicating significant thymidine metabolism after the first 10 min.
Time Course and Sodium Dependence of Thymidine Uptake.
The time course of thymidine (10 μM) uptake by rat microglia MLS-9 cells (Fig. 2) shows that in the presence of Na+, uptake was a linear function of time for the first minute but began to plateau by ∼3 min. This linearity, taken together with the metabolism data, suggests that the uptake at 1 min reasonably well represents the initial unidirectional thymidine influx into the cells.

Time course of thymidine uptake. Uptake of thymidine (10 μM in standard incubation medium) was measured at 37°C. Inset shows the linearity of thymidine uptake at time points ≤1 min (three separate experiments; coefficient of correlation r= 0.99). The results are expressed as means ± S.D., with each data point in an individual experiment representing quadruplicate measurements.
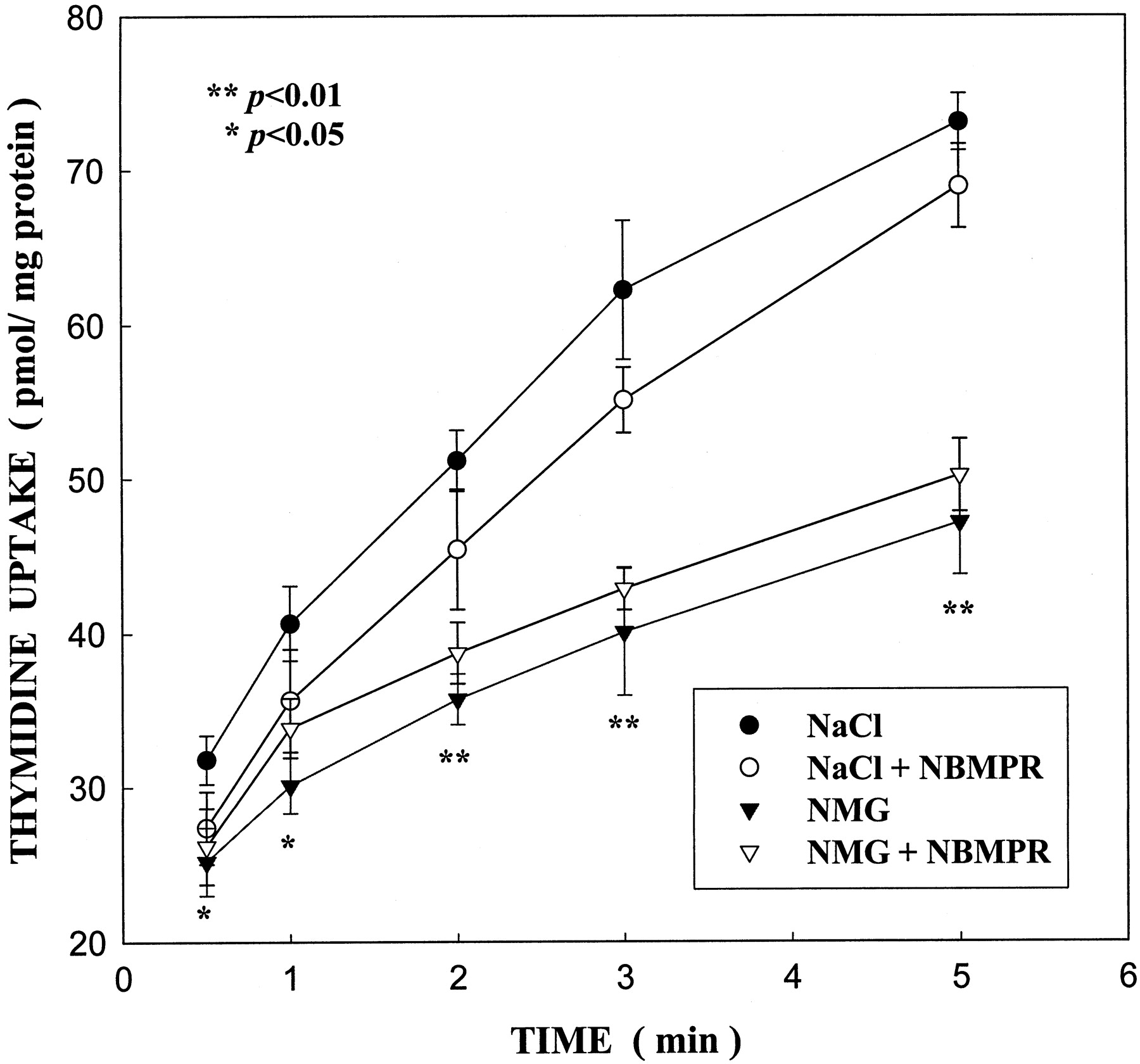
Thymidine (10 μM) uptake by MLS-9 cells at 37°C was found to be in part dependent on a sodium gradient (concentration outside greater than inside; Fig. 3). Intracellular accumulation of thymidine at these early time points (0.5–5 min) was significantly inhibited by substituting sodium by the impermeant cation, N-methyl-d-glucamine (NMG). For instance, thymidine uptake at 1 min was reduced by ∼50%. This suggests the involvement of both a Na+-dependent membrane transporter and possibly a Na+-independent system (i.e., equilibrative). Both transport mechanisms showed insensitivity to NBMPR. Further examination of the Na+-independent component was undertaken. A concentration-dependence study of thymidine (10 μM to 3 mM) in the absence of sodium showed that the thymidine Na+-independent component was linear (data not shown) and similar to the diffusion component (i.e., nonspecific uptake) obtained in the thymidine kinetic experiment (see below). This suggests that in the absence of sodium, thymidine uptake does not involve a carrier-mediated process. Our studies focused then primarily on the characterization of thymidine uptake by microglia cells in the presence of sodium. Correction for the nonspecific uptake, as measured by uptake values in the presence of 1 mM thymidine, was applied.

Effect of Na+ and NBMPR on thymidine uptake. The uptake of 10 μM thymidine at 37°C was determined at the indicated times in standard NaCl incubation medium (●) or in 138 mM NMG medium (no sodium; ▾) in the absence or presence of 1 μM NBMPR. Data are expressed as means ± S.D. of two separate experiments; each experimental point represents four replicates. Values marked with asterisks are significantly different from control (no NBMPR) in standard NaCl medium.
Na+ Concentration Dependence.
Specific thymidine uptake (10 μM) as a function of extracellular Na+ concentrations increased in a hyperbolic manner (Fig. 4), which was well described by the Hill equation:

Na+ concentration dependence of thymidine uptake. MLS-9 cells were incubated for 1 min at 37°C in standard incubation medium containing 10 μM thymidine and 5 to 150 mM Na+ (substituted by equimolar NMG). A least-squares fit of the data was used to estimate KNa,Vmax, and n, the Hill coefficient. The 1 mM thymidine uptake value was used to correct for the nonspecific uptake (see Materials and Methods). Results are expressed as means ± S.D. of three separate experiments; each experimental point represents quadruplicate measurements.
Kinetics of Thymidine Uptake.
Thymidine uptake during the first minute was measured as a function of thymidine concentration (10–750 μM) at 37°C (Fig. 5). Nonlinear least-squares regression analysis was applied. The total rate of thymidine uptake can be described by the following equation:

Kinetics of Na+-driven thymidine uptake by MLS-9 monolayer cells. The thymidine uptake rate was measured at various thymidine concentrations (10–750 μM) in the standard NaCl medium at 1 min, 37°C. The specific rate was calculated by subtracting the nonspecific diffusion component from the total rate. The kinetic constants (Km,Vmax) and the diffusion coefficientD were determined by a nonlinear least-squares fit of the Michaelis-Menten equation to the data from five separate experiments. Results are expressed as means ± S.D.; each experimental point represents quadruplicate measurements.
Substrate Specificity.
To explore the substrate selectivity of the transporter, thymidine uptake was measured (1 min at 37°C) in the presence of a variety of nucleosides: adenosine, guanosine, cytidine, and uridine. All purine and pyrimidine nucleosides (50 μM) tested significantly inhibited (by 40–48%) specific thymidine uptake by MLS-9 cells. The uptake of radiolabeled thymidine obtained in the presence of 1 mM thymidine was measured to account for the nonspecific transport shown in Figs. 4 and 6 to 11, and the total uptakes were corrected accordingly. The IC50 values were determined by fitting the data to a sigmoidal equation:

Dose dependence of pyrimidine (A) and purine (B) nucleosides in inhibiting thymidine uptake by MLS-9 cells. Specific thymidine (8.3 nM) uptake was determined at 1 min, 37°C, and in the presence of increasing concentrations of the nucleosides. The data were fit to a sigmoidal inhibition model (see Materials and Methods). Nonspecific thymidine uptake was estimated as in Fig.4. Results are expressed as percentage of the control uptake ± S.D. of three separate experiments; each experimental point represents quadruplicate measurements. Some error bars are smaller than the symbols.

Eadie-Hofstee plots of ZDV inhibition of thymidine uptake. Thymidine (1, 10, 50, and 100 μM) uptake was measured in standard NaCl medium at 1 min, 37°C, in the absence (control) or presence of various concentrations of ZDV (1, 10, and 30 μM). A least-squares regression analysis was used to determine the Eadie-Hofstee linear correlations. The data were corrected for the nonspecific uptake as described in Fig. 4. Results are expressed as means ± S.D. of two separate experiments; each experimental point represents quadruplicate measurements.
In addition, all the nucleoside probes (purines, pyrimidines) inhibited thymidine uptake in a competitive manner. TheKi values were obtained from the competitive inhibition equation:

Eadie-Hofstee plots of pyrimidine nucleoside inhibition of thymidine uptake. Thymidine uptake was measured at 1 min, 37°C, in standard NaCl medium containing various thymidine concentrations (1, 10, 50, and 1 μM), in the absence (control) or presence of a test nucleoside; uridine (A) or cytidine (B) at 10, 30, and 50 μM. The Ki values were estimated from the apparent Km values with a least-squares regression of the data from two separate experiments. Nonspecific thymidine uptake was estimated as in Fig. 4. Results are expressed as means ± S.D.; each experimental point represents quadruplicate measurements.

Eadie-Hofstee plots of purine nucleoside inhibition of thymidine uptake. Thymidine (1, 10, 50, and 100 μM) uptake was measured in standard NaCl medium at 1 min, 37°C, in the absence (control) or presence of various concentrations of a test nucleoside; guanosine (A) or adenosine (B) at 10, 30, and 50 μM. TheKi values were estimated as in Fig. 7 with two separate experiments. Nonspecific thymidine uptake was estimated as in Fig. 4. Results are expressed as means ± S.D.; each experimental point represents quadruplicate measurements.
Nucleoside Analog Drugs Inhibit Thymidine Uptake.
Because multiple carrier systems, including NTs, may be involved in the membrane permeation of nucleoside analog drugs, we investigated the effect of 3TC, ddI, ddC, abacavir, and ZDV on specific thymidine uptake by MLS-9 cells (Fig. 9). At 1 mM, the drugs that showed the greatest inhibition were ZDV (86%) and abacavir (71%). The IC50 values for these nucleoside analog drugs were estimated as 0.6 and 135.1 μM for ZDV and abacavir, respectively (Fig. 10). ZDV inhibited thymidine uptake in a noncompetitive manner, i.e, a least-squares regression analysis yielded Eadie-Hofstee linear correlations, wherein the Km (slope) remained approximately constant but Vmax (they-intercept) decreased with increasing concentrations of the inhibitor (Fig. 11). These data suggest that some nucleoside analog drugs (i.e., ZDV) are strong inhibitors of thymidine uptake by microglia cells and may modulate the accumulation of the natural nucleoside substrates into the cells.

Effect of nucleoside analog drugs (3TC, ddI, ddC, abacavir, and ZDV) on specific thymidine uptake. Specific thymidine uptake (8.3 nM) was measured in standard NaCl medium at 1 min, 37°C, in the presence of 1 mM various nucleoside analogs. The data were corrected for the nonspecific uptake as in Fig. 4. Results are expressed as means ± S.D. of three different experiments; each experimental point represents quadruplicate measurements. Asterisks indicate values that are significantly different from the control value.

Dose dependence of nucleoside analogs in inhibiting thymidine uptake. Uptake of thymidine (8.3 nM) was measured in standard NaCl medium at 1 min, 37°C, in the presence of various concentrations (0.25 μM-1 mM) of ZDV or abacavir. The 50% inhibition values calculated from the sigmoidal model of inhibition were 0.6 and 135.1 μM for ZDV and abacavir, respectively. The data were corrected for the nonspecific uptake as described in Fig. 4. Results are expressed as percentages of the control ± S.D. of two separate experiments; each experimental point represents quadruplicate measurements.
Discussion
The major target cells for HIV-1 infection in the brain appear to be macrophages and microglia cells (Price et al., 1988; Watkins et al., 1990; Takahashi et al., 1996). These cells act as a viral reservoir, possibly releasing the virus, viral proteins, and other factors that lead to neurotoxicity. The entry of the virus into the CNS may occur by: 1) the normal physiological repopulation of the CNS by infected peripheral monocytes/macrophages, 2) the latent infection of cells comprising the choroid plexus and BBB, and/or 3) the increased permeability of the BBB during disease progression (Dickson et al., 1991). Approximately half of the children and one-third of adults with AIDS develop neurological complications, including cognitive-motor dysfunction, sensory impairment, and frank dementia (Price et al., 1988; Gendelman et al., 1994). After microglial infection, several of the neurotoxins that appear to be prime mediators of the adverse effects on neurons (platelet activating factor; nitric oxides; superoxides; tumor necrosis factor-α; and interleukins 1, 6, and 1β) are produced by activated microglia (Gendelman et al., 1994). In AIDS, as in a variety of neurodegenerative diseases, an excessive rise in neuronal intracellular Ca2+ is thought to contribute to neurotoxic events leading to free radical formation, cell necrosis, and/or apoptosis (Lipton, 1994).
Because the CNS infection by HIV-1, like the infection in the peripheral immune system, may be accessible to control by nucleoside analog drugs, it is important to understand how these drugs are transported into the CNS, and particularly, into the brain parenchyma cells that HIV-1 infects, i.e., the microglia. Numerous studies have investigated the permeation of nucleosides and nucleoside analog drugs across the brain barriers (i.e., blood/CSF barrier and BBB); however, at present, no information is available on their accumulation, distribution, and metabolism within the brain parenchyma. The purpose of the present study was to characterize the uptake of the nucleoside pyrimidine probe thymidine and its inhibition by nucleoside analogs by using an in vitro model of microglia cells.
The cell culture model used in the present study consisted of a rat microglia cell line called MLS-9. Both the phenotype and physiology of these cells have been examined (Zhou et al., 1998; L. C. Schlichter, F. S. Cayabyab, R. Khanna, and T. Mertens, unpublished data; see Materials and Methods). The cells stain for several specific microglial markers, i.e., OX-42 antibody (labels membrane C3 complement receptors), ED-1 antibody (labels a lysosomal membrane antigen), isolectin B4 (labels membrane α-d-galactose residues), and diI-acetylated low density lipoprotein and lucifer yellow, which are taken up by pinocytosis. When grown to confluence, MLS-9 cells exhibit several of the morphological and functional properties of activated microglia. That is, the cells are rounded up (like phagocytic microglia), they express high levels of complement C3 receptors and lysosomes (as judged by OX-42 and ED-1 staining), and they can produce large amounts of nitric oxide (C. A. Colton and L. C. Schlichter, unpublished data). Distinct clinical features associated with HIV-specific neuropathology include astrocytosis, perivascular cluster of macrophages, multinucleated giant cells, and microglial nodules [extensive aggregation of pleomorphic (majority rounded up) microglia with sparse cytoplasm and short processes (Kure et al., 1990)]. Thus, it is appropriate to use activated microglia to investigate nucleoside transport because this reflects the clinical stage found during the HIV-1 infection in the brain (Dickson et al., 1991), a state at which there is enhanced microglial expression of activation antigens and proliferation (Gehrmann, 1996).
Our data show that intracellular thymidine accumulation by MLS-9 microglia is in part dependent on extracellular sodium, exhibits a hyperbolic response to increasing external Na+, and is significantly reduced by abolishing the normal inwardly directed sodium gradient. This suggests that thymidine influx occurs in part via an active Na+-coupled transporter. These results are consistent with a kinetic model in which the cosubstrate, Na+, binds and modulates the carrier-substrate interactions. The Na+/nucleoside-coupling stoichiometry was found to be 2:1. As previously shown for a number of Na+-dependent concentrative nucleoside transport systems (N1-N4) and the equilibrative system (ei) (Cass, 1995; Wang et al., 1997), in MLS-9 cells the standard inhibitor (1 μM) for the equilibrative NT, NBMPR, had no effect on thymidine accumulation.
Kinetic analysis of the Na+-dependent component of thymidine uptake revealed a saturable, single process obeying Michaelis-Menten kinetics. The Km (44 μM) and Vmax (273 pmol/mg/min) values for MLS-9 cells are higher than those estimated for the concentrative (N3 type) thymidine transporter in rabbit choroid plexus (13 μM and 62 pmol/mg/min; Wu et al., 1992). However, they are similar to the range reported for Na+-dependent NTs in bovine renal (8 μM and 362 pmol/mg/min; Williams and Jarvis, 1991) and human renal brush border membrane vesicles (27 μM and 325 pmol/mg/min; Gutierrez and Giacomini, 1993).
Our studies show that purine and pyrimidine nucleosides (50 μM) inhibit 1-min thymidine specific uptake in a concentration-dependent (IC50 = 30–40 μM) and competitive (Ki = 38–45 μM) manner, suggesting that these compounds permeate microglia cells by a common transporter. Because the substrate and the inhibitor appear to bind reversibly to the same active site, our results are consistent with the broad substrate selectivity of a cib (N3) type of concentrative NT (Cass, 1995; Wang et al., 1997). Similar broad specificity, Na+-dependent N3 transporters for purines and pyrimidines have been identified in a human colon cell line (CaCo2), myeloid cell lines (HL60 and V937), blast cells from patients with acute myelogenous leukemia (Belt, 1983), and rat intestinal epithelial cells (Huang et al., 1993).
The specificity of the NT system was further investigated by measuring thymidine uptake in the presence of nucleoside analog drugs, compounds structurally modified on the ribose ring that are important for the treatment of HIV infection. Most importantly, we found that two of these drugs, ZDV and abacavir, profoundly reduced thymidine uptake by MLS-9 cells, and ZDV, with an IC50 value of 0.6 μM, is a very potent, noncompetitive inhibitor, rather than a substrate for the NT. The IC50 value for ZDV is within the therapeutic concentration (1 μM) found in the human CSF (Klecker et al., 1987).
It is thought that the physiological role of concentrative NTs is to salvage nucleosides required for several cellular processes, thus avoiding the high-energy cost of de novo synthesis. In situ brain perfusion studies in guinea pigs have demonstrated that thymidine is transported across the BBB by facilitated diffusion (ei andes) systems and/or an active N2 (cit) transporter (Thomas and Segal, 1997). However, single-pass studies with i.v. infusion methods have shown that thymidine does not readily cross the BBB (Cornford and Oldendorf, 1975; Spector and Berlinger, 1982), suggesting as the primary route of entry, the choroid plexus. At the CSF-blood barrier, an N3 concentrative transporter plays an important role in purine and pyrimidine transport into the CNS and, thus, in the homeostatic function of the choroid plexus (Spector and Berlinger, 1982). In rabbit choroid plexus, both es and eiequilibrative transport systems have been reported and in vivo both systems transport nucleosides from blood to CSF (Spector, 1982). The presence of an active NT in activated microglia may enhance the capacity of the cells to salvage nucleosides from the low levels found in the brain circulation. The Km for thymidine uptake is substantially higher than the presumed endogenous nucleoside plasma levels, e.g., in rabbit the estimated plasma and CSF concentrations are 0.6 ± 0.1 and 0.6 ± 0.0 μM, respectively (Eells and Spector, 1983a). In humans, much lower endogenous thymidine concentrations have been reported in plasma (0.2 ± 0.0 μM) and lumbar CSF (0.06 ± 0.00 μM; Eells and Spector, 1983b). Thus, if a similar NT is functionally expressed in humans, this transport system would be undersaturated and would function as an efficient pathway for nucleoside influx into microglia. It is also important that these low plasma thymidine concentrations should not inhibit the uptake of other nucleosides that share the same transporter.
The present study is, to our knowledge, the first report of a sodium-driven, NT system in microglia cells. Moreover, we do not know if the expression of NTs in microglia is regulated, i.e., depending on cell morphology and activation status, which are well known to correlate with changes in a wide range of surface molecules. It is interesting to note that expression of concentrative NTs is up-regulated in proliferating (activated) lymphocytes and thymocytes (Meckling-Gill et al., 1993). Although similar changes in expression of equilibrative systems have not been observed (Plagemann et al., 1988), this warrants further investigation. In the future, it will be valuable to compare the functional expression of these transporters in resting versus activated microglia to determine whether there is differential expression, and if so, the regulation pathways that control these transport systems in brain immune cells. Most importantly, NTs may play an important role in the entry of some antiretroviral drugs into microglia cells, the primary HIV-1 target in the brain.
Acknowledgments
We thank Dr. Pennefather for helpful comments on the manuscript.
Footnotes
-
Send reprint requests to: Dr. Reina Bendayan. Department of Pharmaceutical Sciences, Faculty of Pharmacy, University of Toronto, 19 Russell St., Toronto, Ontario M5S 2S2, Canada. E-mail:r.bendayan{at}utoronto.ca
-
↵1 This work is supported by a grant from the Ontario HIV Treatment Network, the Canadian Foundation for AIDS Research, and the Medical Research Council (MT-13657). M. Hong is a recipient of an Ontario HIV Treatment Network Studentship Award.
-
↵2 Current address: Department of Pharmaceutical Sciences, Faculty of Pharmacy, University of Toronto, Toronto, Ontario M5S 2S2, Canada.
-
↵3 M. Hong, P. Pennefather, L. Schlichter, and R. Bendayan. Transport properties of thymidine by a rat microglia cell line. Abstract selected for presentation at the 100th Annual Meeting of the American Society for Clinical Pharmacology and Therapeutics, San Antonio, TX, March 1999.
-
↵4 Current address: The Neuroscience Institute, University Health Network, Toronto, Ontario M5T 2S8 and Department of Physiology, University of Toronto, Toronto, Ontario M5S 1A1, Canada.
- Abbreviations:
- CNS
- central nervous system
- NT
- nucleoside transporter
- es
- equilibrative, sensitive to NBMPR inhibition
- NBMPR
- 6-(4-nitrobenzyl)-thio-9-β-d-ribofuranosylpurine
- ei
- equilibrative, insensitive to NBMPR
- cif or N1
- concentrative, NBMPR insensitive, accepts formycin B as a permeant
- cit or N2
- concentrative, NBMPR insensitive, common permeant: thymidine
- cib or N3/N4
- concentrative, NBMPR insensitive, broad specificity
- cs or N5
- concentrative, NBMPR sensitive
- BBB
- blood brain barrier
- CSF
- cerebrospinal fluid
- ZDV
- zidovudine
- 3TC
- lamivudine
- ddI
- didanosine
- ddC
- zalcitabine
- NMG
- N-methyl-d-glucamine
- Received June 10, 1999.
- Accepted September 29, 1999.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}