


Abstract
A turnover model for irreversible inhibition of gastric acid secretion by omeprazole in gastric fistula dogs was developed using data from studies with both short- and long-term measurement periods. In the short-term experiments, after stimulation of acid secretion with histamine, the dogs were infused i.v. with omeprazole and acid secretion was measured for 5 h. Dose and infusion times were varied to produce different concentration-time profiles and schedule dependence in the inhibitory effect of omeprazole was observed. In the long-term experiments, dogs were given single intraduodenal doses, which inhibited the acid secretion for several days. Combining the short-term and long-term data allowed the observation of a biphasic recovery of acid secretion that was described by the turnover model. Second order association rate constants (kome) for the covalent binding of omeprazole to H+,K+-ATPase were estimated to 11 and 3.0 l/μmol/h for the i.v. and intraduodenal experiments, respectively. The apparent turnover rate constant of the enzyme (kout) was 0.013 h−1 and the corresponding half-life of inhibition of acid secretory capacity was 54 h. The potency, calculated as koutover kome, was 4.3 and 1.2 nM for the intraduodenal and i.v. doses, respectively. Allometric scaling of the model resulted in trustworthy predictions for observations previously done in humans. The model predicted a good correlation between maximal inhibitory effect and exposure (area under the plasma concentration curve). The time dependence in this relation was also predicted by the model.
Omeprazole is a substituted benzimidazole with the capacity to inhibit gastric H+,K+-ATPase, the proton-transporting enzyme in the parietal cells that secrete hydrochloric acid into the stomach (Wallmark et al., 1983). The acidic condition in the parietal cell results in activation of omeprazole to a cationic sulfenamid, which binds irreversibly by forming disulfide link complexes with the enzyme (Keeling et al., 1987; Lorentzon et al., 1987). The gastric acid secretion is effectively controlled, resulting in a rapid healing rate for acid-related diseases.
The irreversible binding of omeprazole to the enzyme results in reduced acid secretion (response). Because of the irreversible inactivation of enzyme, the time course of the omeprazole effect is determined by the kinetics of enzyme synthesis, activation, and elimination and not the pharmacokinetics of omeprazole. Thus, even if omeprazole interacts directly with the enzyme there is no direct relationship between plasma concentration and effect and this is described as an indirect response. This has been clearly demonstrated both in animal studies and in patients. In the dog, the plasma half-life of omeprazole was about 1 h, whereas the duration of the effect was several days after single intraduodenal (i.d.) doses (Larsson et al., 1983). The maximal inhibition of gastric acid secretion measured in the dog was shown to correlate to the exposure (area under the plasma concentration curve, AUC) of omeprazole but not to the peak plasma concentration (Larsson et al., 1982). Similarly, the inhibitory effect of omeprazole on gastric acid secretion determined in patients correlated well with the AUC (Lind et al., 1983; Cederberg et al., 1985, 1992).
In the present report, the aim is to develop a turnover model for the inhibitory effect of omeprazole on acid secretion. Results from previously published experiments in the dog were used (Larsson et al., 1982, 1983). A set of four basic turnover models was previously reviewed and has been successfully applied to indirect drug effects (Dayneka et al., 1993; Jusko and Ko, 1994). This type of turnover model is now used and extended to describe the observed lack of temporal correlation between the plasma concentration and effect of omeprazole with the aim to separate drug-specific factors (pharmacokinetics and binding interaction) from system-specific factors (enzyme turnover and distribution). The system-specific parameters are scaled up from dogs to humans to compare the predictions of the response-time course with observations in humans.
Materials and Methods
Experimental Design
Short-Term Experiments.
The effect of omeprazole was studied in the dog with frequent measurements of gastric acid secretion during a relatively short period of 5 h post dose. The results of these experiments have partially been published previously (Larsson et al., 1982), but no attempts were made to model the data. Two dogs with a conventional gastric fistula for sampling of gastric juice were given omeprazole as i.v. infusions on six occasions separated by washout periods of 1 week. The infusion schedules are summarized in Table1. Food and water were withdrawn 18 h before each experiment. On each experimental occasion the dogs were examined for the presence of spontaneous gastric acid secretion. When the lack of basal secretion had been determined, the dogs were given histamine (150–175 nmol/kg/h) as a continuous i.v. infusion. After 2 h of histamine infusion, when the acid secretion was stimulated to a stable level of about 80% of maximal secretion, an i.v. infusion of omeprazole in 10% polyetyleneglycol or 10% polyetyleneglycol only (0.1 ml/min) was started. During the infusion period, gastric juice was collected by free flow from the cannula in consecutive 30-min intervals. An aliquot of each sample was titrated with 0.1 M NaOH to pH 7 using an automatic titrator and pH meter. The acid secretion (=response) was calculated as mmol H+/30 min. The baseline value of response, expressed as 100%, was determined by three placebo experiments in each dog (Fig. 1). The response decreased over time for the placebo dogs. The effect of omeprazole was calculated as the change in response in comparison with the placebo experiments at each corresponding time point. Samples of blood were taken at 0, 20, and 25 min and at 1, 2, 3, 4, and 5 h after the start of infusion with active compound and the plasma concentration of omeprazole was determined according to the method ofLagerström and Persson (1984).
Omeprazole infusion schedules, doses of the experiments, and estimates of pharmacokinetic parameters (CV%) when data for two dogs were pooled for each of six experiments

Baseline response versus time in dog 1 (solid lines) and dog 2 (dotted lines). Three placebo experiments were performed in each dog.
Long-Term Experiments.
Published data in the dog were used that referred to response every 24 h for 4 days after dosing with omeprazole (Larsson et al., 1983). The long measurement period in these experiments allowed quantification of the duration of the effect. Three dogs with a conventional gastric fistula for sampling of gastric juice and a duodenostomy for intraduodenal administration of omeprazole were used in the study. Gastric acid secretion was stimulated by an i.v. infusion of histamine (100–200 nmol/kg/h). The doses of histamine were chosen individually to give a stable acid secretory response (40–60% of maximal response) during the experiments. Each dog received single intraduodenal doses of omeprazole on four occasions for evaluation of gastric acid secretion. The doses studied were 0.5, 1.25, 2.5, and 5.0 μmol/kg. Mean estimates of response, expressed as a percentage of the acid secretion predose, for the three dogs were used in the modeling. The response was calculated as the change in response in comparison with placebo experiments performed in the same dogs. Pharmacokinetic data were available for one dose. A one-compartment model with first order absorption was fitted to the average plasma concentrations of omeprazole given intraduodenally (0.25 μmol/kg, n = 3). This dose was chosen because it corresponds approximately to the ED50 after i.d. administration regarding inhibition of acid secretion (Larsson et al., 1983). Blood samples for the determination of omeprazole concentration in plasma were taken before and 5, 15, 45, 105, 165, and 225 min after dose.
Data Analysis
The plasma concentrations of omeprazole and the response were modeled sequentially. Compartmental models were applied to describe the pharmacokinetics of omeprazole. The individual estimates of the pharmacokinetic parameters were then fixed and the model-predicted plasma concentrations were used as input to the pharmacodynamic models. Response data from the short-term experiments were used for development of the basic and back-flow turnover model, which were regressed simultaneously to response data from the six experiments in each dog. A combination of selected short-term and long-term experiments was used to allow discrimination between models and improve precision in the estimation of model parameters. For all modeling, ordinary least-squares in WinNonlin Professional (Pharsight Corporation, Mountain View, CA) was used.
Theoretical Basis for Turnover Models
Basic Turnover Model.
An important assumption of the model is that acid secretion (AS) is directly proportional to the concentration of active proton pumps (enzyme, E) (Fig.2A; eq. 1). The response (R) variable used in the modeling was the placebo-corrected gastric acid secretion according to eq. 1:

Schematic illustration of the basic turnover model (A) and the back-flow model (B) that were used to model the irreversible action of omeprazole. kome is the second order rate constant for the irreversible binding of omeprazole to active H+,K+-ATPase.
Back-Flow Model.
Low-dose and short-term exposure to omeprazole resulted in a rapid return of response to baseline. High-dose or extended exposure to omeprazole, on the other hand, resulted in a slow recovery of the response. The slow recovery is due to de novo synthesis of enzyme because of the irreversible action of omeprazole. To be able to describe both the fast initial and the slow terminal recovery process, the presence of a pool of enzyme in its inactive state was considered. In the modeling, the basic (Fig. 2A) turnover model was extended (Fig. 2B), where in the latter p represents the contribution of the enzyme pool. When active enzyme is inhibited by omeprazole, R will decrease. In the proposed model, this is governed by stimulation of loss of R. A low dose would produce a transient decline of active enzyme followed by a rapid replenishment of active enzyme from the pool of inactive enzymes, corresponding to replenishment of R from P. Consequently, a rapid recovery of R will be seen. A high dose or extensive exposure to omeprazole, on the other hand, would not only decrease active enzymes but also eventually inactive enzymes would be activated and subsequently eliminated, corresponding to a draining of both R and P. As a result, the recovery ofR would be slower because both compartments need to be refilled. The rate of change of P is described by eq. 5:
Model Scale-Up.
The allometric approach was used to scale the first order rate constants of the back-flow model (k1, k2, andkout). The first order rate constants for the enzyme turnover were scaled according to eq. 10:
Observed (symbols) and predicted (solid lines) plasma concentrations in dogs 1 (▪) and 2 (●) for each of six dosing schedules.
Results
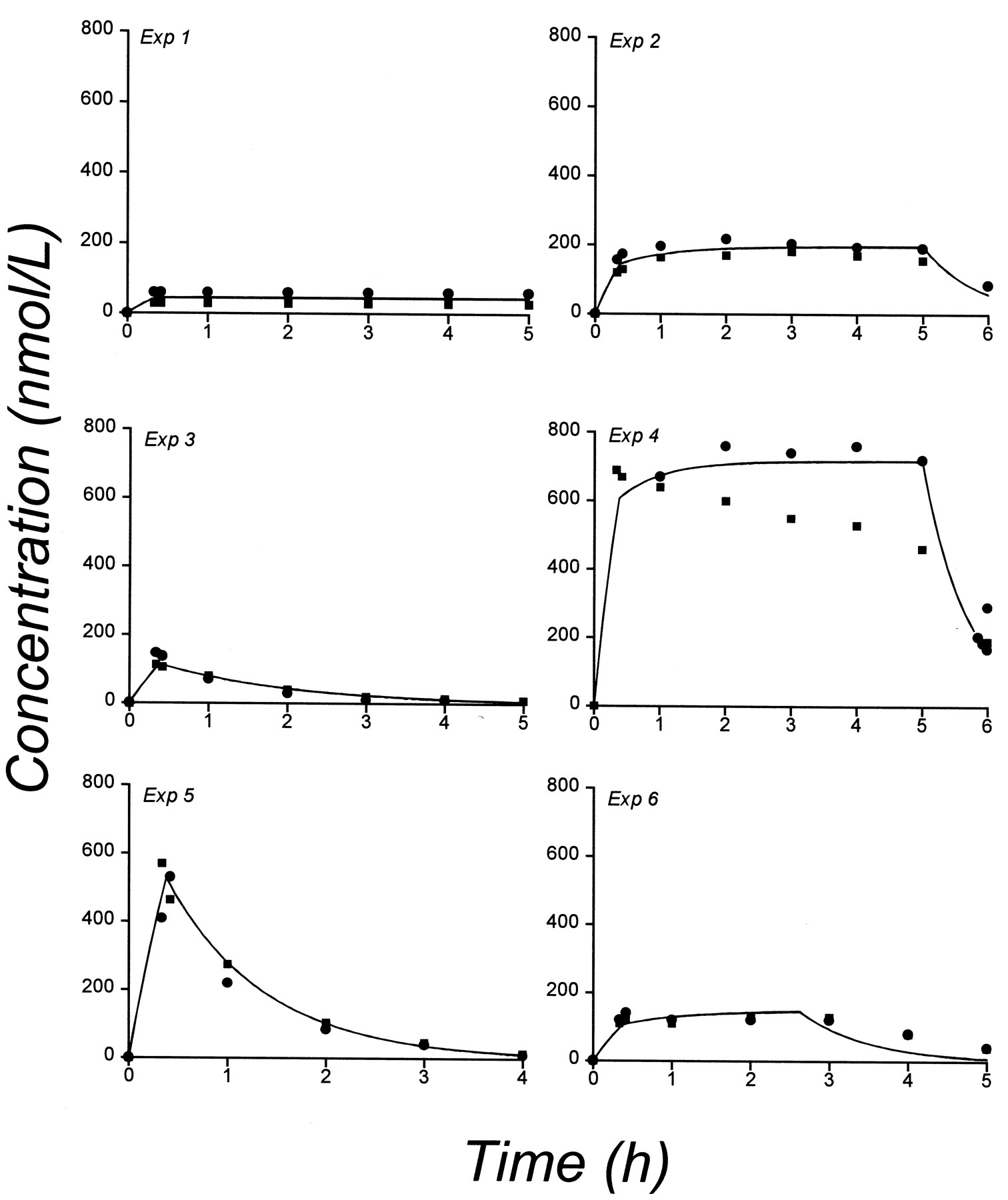
A one-compartment model was fitted to the plasma concentration data of the short-term experiments. Observed and predicted plasma concentrations in the two dogs for each of the six dosing schedules are shown in Fig. 3. Estimates of the pharmacokinetic parameters for the pooled data (i.e., V and k) are given in Table 1. Two enzyme turnover models, the basic and the back-flow model, were compared. The response data for the six short-term experiments were pooled in the model fitting. Parameter estimates for the two models are given in Table 2. For the basic model, the half-life of the apparent turnover rate constantt1/2 (kout) of the enzyme was estimated to 1.0 and 2.2 h for dog 1 and 2, respectively, and for the back-flow model the estimates were 1.4 and 4.2 h. Thus, the estimates of the turnover rate constant and the corresponding half-life were similar for the two models. The second order rate constant for omeprazole binding to the enzyme was 10-fold higher for the back-flow model compared with the basic model. Observed and model-predicted response curves are shown in Fig.4 for dog 1. As shown in the figure, the basic model appears to adequately describe the response in most of the short-term measurement experiments, and it is not possible to discriminate between the basic and the back-flow model using this data set. The systematic bias observed in some fits could be due to some change in parameter values occurring between experiments. To fit each experiment separately led to improved description of the data, but unacceptable uncertainty of the parameters. A schedule dependence in the exposure to omeprazole was observed. For example, in experiments 5 and 6 the same total dose was given with different rates and lengths of infusion, which resulted in different time course and duration of the effect. However, the maximum inhibition was similar.
Mean estimates (CV%) obtained with the basic turnover and back-flow model

Observed (symbols) and model-predicted (lines) acid secretion versus time in dog 1 for the basic turnover model (solid lines) and the back-flow model (dashed lines). Each model was fitted to all six experiments simultaneously.
Results of long-term experiments where the response was measured for several days were obtained from a previously published study (Larsson et al., 1983). Pharmacokinetic data were available for one dose (0.25 μmol/kg) and a one-compartment model with first order absorption was fitted to the average plasma concentration presented in Larsson et al. (1983) (Fig. 4). This resulted in CL/F = 0.33 l/h, V/F = 0.66 liter, and an absorption half-life = 2.3 min. At the higher doses (0.5, 1.25, 2.5, and 5 μmol/kg), the plasma concentration measured at 30 min was 0.54, 1.0, 2.11, and 4.12, respectively. The kinetics was therefore assumed to be linear and the predicted curves based on the parameters obtained after 0.25 μmol/kg are shown in Fig.5. To be able to discriminate between the two pharmacodynamic models, data for the long-term experiments were combined with two of the short-term experiments in dog 1 (experiments 4 and 5). For the combined data, the back-flow model was parameterized with two different rate constants for the binding of omeprazole [kome1 (i.v.) andkome2 (i.d.)]. The rate constants were estimated to 11 and 3.0 l/μmol/h forkome1 andkome2, respectively. The potency of omeprazole (kout/kome) was calculated to be 1.2 and 4.5 nM for the i.v. and i.d. doses, respectively. The effective half-life for inhibition of acid secretion [t1/2(kout)] was 54 h in the dog. Figure 6shows two representative curves for the observed and model-predicted response when short-term and long-term experiments were fitted simultaneously, using both the basic and the back-flow model. In experiment 5 a rapid recovery to baseline was observed, which was predicted by the back-flow model, but not by the basic model. The slow recovery observed in the i.d. experiments was predicted by both models. The onset of the effect for the basic model was slower because the basic model can only predict a monophasic recovery. The parameters obtained with the back-flow model was scaled to humans (k1 = 0.3 h−1,k2 = 1.52 h−1,kout = 0.0097 h−1). The predicted response in humans, based on these parameters, is shown in Fig. 7 superimposed on observations from Lind et al. (1983). Predicted response after repeated administration of 10, 40, and 400 mg of omeprazole is shown in Fig.8A. The Hill equation was used in an attempt to correlate the maximum effect and AUC (Fig. 8B) because such a correlation has been reported previously for omeprazole (Larsson et al., 1982; Lind et al., 1983; Cederberg et al., 1985, 1992). The obtained parameters were AUC50(day 1) = 51.5 μM/h, AUC50(day 6) = 20.2 μM/h,N(day 1) = 1.11, andN(day 6) = 1.15.
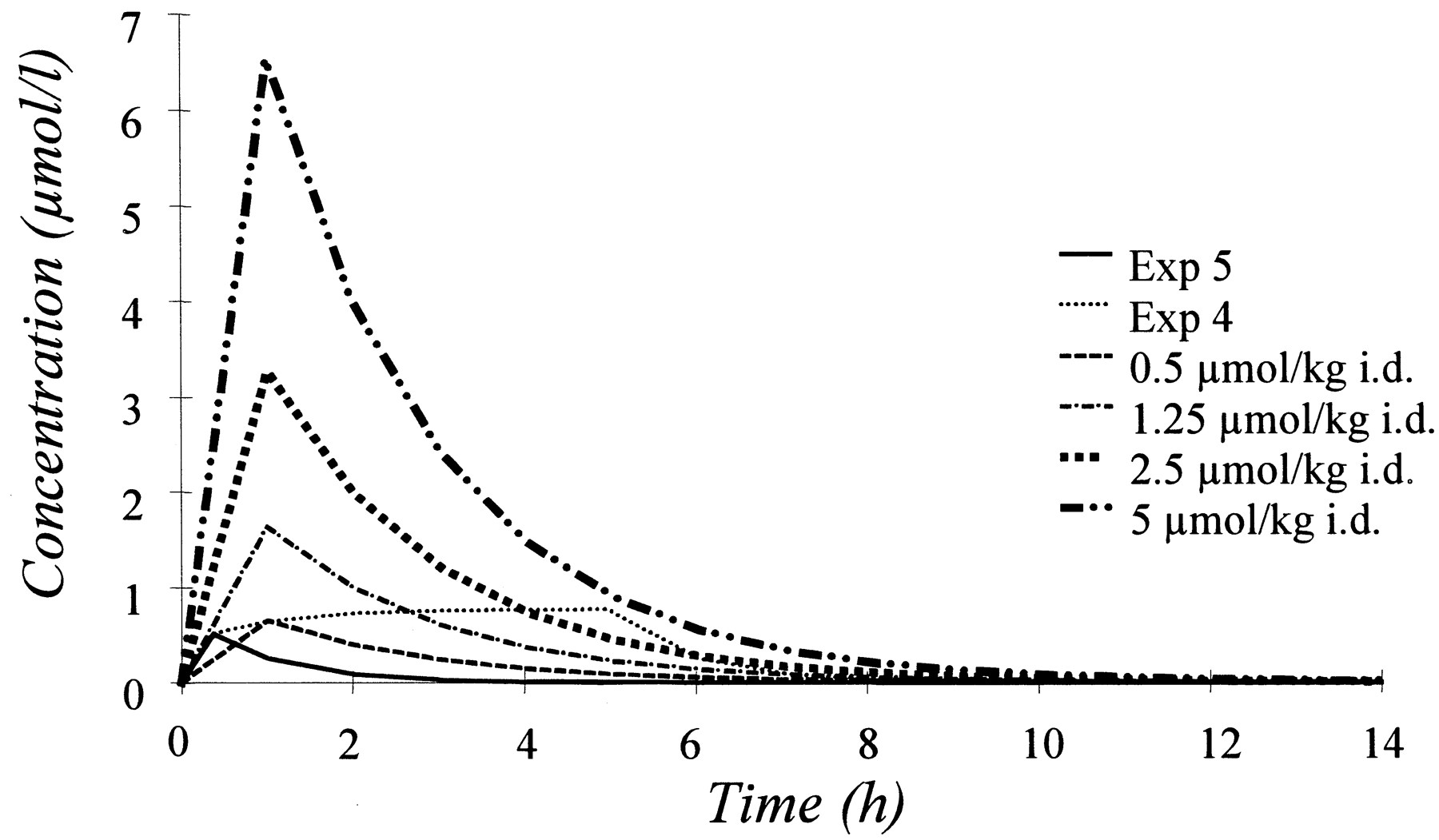
Simulated plasma concentration-time curves for the two short-term experiments (experiments 4 and 5) and the four long-term experiments that were used in the model fitting.

Observed (●) and model-predicted (lines) response for the basic turnover model (solid lines) and the back-flow model (dashed lines). Each model was simultaneously fitted to experiments 4 and 5 (short-term experiments) and to four different single intraduodenal doses: 0.5, 1.25, 2.5, and 5.0 μmol/kg (long-term experiments). Left, one representative long-term experiment and right, one of the short-term experiments.

Predicted response-time course (solid line) of the scaled-up human model superimposed on observations (●) from Lind et al. (1983). A, different kome values were estimated for the i.v. and i.d. doses. The dashed lines represent a 100% increase and 50% decrease in kome. These simulations were done to predict the impact of potential differences in protein binding of omeprazole in dogs and humans. B, different k1 values estimated for the i.v. and i.d. doses are shown. The dashed lines represent a 100% increase and 50% decrease in k1. These simulations were done because the degree of stimulation of acid secretion may vary between dogs and humans.
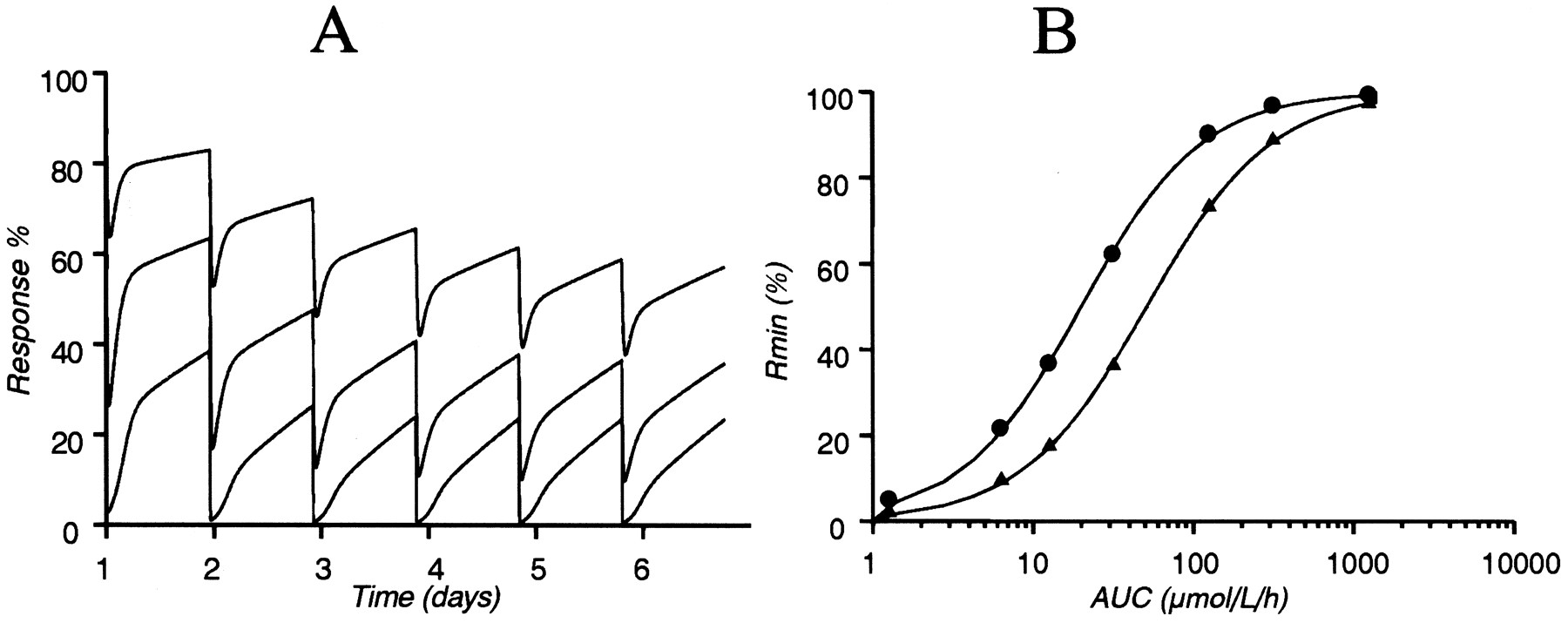
Predicted response after repeated administration of 10, 40, and 400 mg of omeprazole is shown (A). Correlation between the AUCs and maximum inhibition of acid secretion (Rmin) for omeprazole on day 1 (▴) and day 6 (●) (B). The Hill equation was used to describe the correlation between Rminand AUC. AUC50 (day 1) and AUC50 (day 6) were estimated to 51.5 and 20.2 μmol/l/h, respectively. Simulations were done for human responses.
Discussion
A turnover model was developed that describes the kinetics of the effect of omeprazole on acid secretion in the dog. The aim was to develop a model that accounts for the action of omeprazole on the turnover of the gastric proton pumps and separates drug-specific factors from enzyme disposition.
A biphasic recovery of the response was observed by combining experiments with short- and long-term measurement periods. A short-term exposure to omeprazole resulted in a rapid recovery of response, whereas long-term exposure resulted in a slower return of response to baseline. This complexity of the enzyme turnover is supported by in vitro studies (Ekblad, 1989; Licko and Ekblad, 1992). Studies on acid secretory mechanisms performed in isolated frog gastric mucosa showed that the amount of the acid inhibited depends on omeprazole exposure, i.e., it was shown that the amount of suppressed acid increases proportionally with exposure at low exposure but disproportionally with the exposure at high exposure. These observations may be explained by the turnover of H+,K+-ATPase, i.e., at extended exposure the available enzyme is completely depleted and de novo synthesis is required to restore the acid secretion. However, after limited exposure to omeprazole, the depleted amount of enzyme will be replenished from an intracellular pool of enzyme. The basic turnover model was not able to describe the biphasic recovery profile, as shown in Fig. 6. The basic model was therefore extended to the back-flow model, which adequately could describe the combined data for short- and long-term experiments. In the back-flow model, a pool compartment is included that represents a reserve of inactive proton pumps that becomes partially activated upon stimulation of the acid secretion. Elimination of enzyme takes place from both compartments. Omeprazole can only inhibit active ion pumps located at the surface of the secretory membrane and in contact with the K+, Cl− conductances (Hersey and Sachs, 1995). A low dose of omeprazole will remove a minor fraction of the proton pumps in its active form. Then, the loss of active pumps will be replenished from the pool of inactive pumps located in the tubulovesicles in the cytosol of the parietal cell. When the stimulus (histamine) of acid secretion is withdrawn, the pool of inactive enzyme is restored as the acid pumps return to the cytoplasmic tubules. If the dose of omeprazole is high, on the other hand, synthesis of new ion pumps is also required.
According to the obtained parameters with the back-flow model it seems as if histamine only activates a small fraction of the total amount of enzyme, even at maximal stimulation. The pool size in relation to the response compartment size was calculated using the estimated parameters and it was found that only 20% of enzyme was active at maximal stimulation. This is supported by a study by Helander and Hirschowitz (1974) where the surface of the secretory and tubulovesicular membranes in the parietal cells was measured in the dog under various conditions. If it is assumed that the membrane surface is equivalent to amount of active enzyme, about 40% of the pool would be activated at maximal stimulation with histamine according to that study. This suggests that a single dose of omeprazole to a maximally stimulated dog would be able to cause 100% acute inhibition even though only a fraction of the pumps is active.
In vitro studies have shown that the recovery of acid secretion may be more rapid than predicted if recovery would depend on de novo synthesis of enzyme only (Im et al., 1985; Gedda et al., 1995). It was shown that the half-life of the enzyme (protein) is 50 h in the rat and that the half-life of recovery of the enzyme was 15 h, suggesting that the recovery from inhibition by omeprazole depends on both synthesis of new enzymes and on the reversal of inhibition (Gedda et al., 1995). It was suggested that enzyme may be regenerated from the drug-enzyme complex, possibly due to exposure to glutathione in vivo (Fujisaki et al., 1991). If this is the case in humans, recovery of acid secretion after omeprazole treatment does not allow calculation of the rate of turnover of the actual pump protein in humans but rather the half-life of recovery of acid secretory capacity, as pointed out by Gedda et al. (1995). Repeated oral doses of 20 mg of omeprazole resulted in a maximal inhibition of acid secretion of 70% after about 3 days, which was consistent with the effective half-life of inhibition (48 h) (Lind et al., 1983).
The ability to predict the effect in humans was tested by using previously published data (Lind et al., 1983). The turnover rate constants of the enzyme in humans were calculated from the values estimated in the dog when the combined data set of short- and long-term experiments was used (Table 2, back-flow B.). The predictedt1/2(kout) in humans based on the allometric scaling is 71.7 h. The predicted response in humans is shown in Fig. 7 superimposed on the observations. Figure 8 shows simulations of the scaled-up model for different doses of omeprazole (0.4, 2, 4, 10, 40, 100, and 400 mg) given once daily for 6 days. The maximal inhibition is correlated to the daily AUC. The maximal inhibition is predicted to increase over the 6 days even though there is no accumulation of omeprazole in plasma. After 6 days of treatment the relationship between maximal inhibition and AUC is shifted such that a more pronounced inhibition is predicted. The AUC required to give 50% inhibition is approximately twice as high on day 1 as on day 6. A dose of 40 mg of omeprazole is predicted to result in a maximal inhibition of 90% at steady state.
A response model for the effect of omeprazole on gastric acid secretion has previously been reported and successfully applied to rat and human data (Katashima et al., 1995, 1998). The model was built on the principle of reversible action of omeprazole on gastric acid secretion and no pool of inactive enzymes was taken into consideration. The apparent half-life of the monophasic recovery of acid secretory capacity was estimated to 27.5 h for omeprazole. We have attempted to develop a model based on the mechanism of action and time course of restoration of acid secretion, namely, irreversible inhibition of activated enzymes, a pool of inactive enzymes not susceptible to omeprazole binding but in slow equilibration with activated enzymes. Different estimates of the apparent half-life of the inhibitory effect were obtained with the two models. However, due to the differences in the models, these estimates are not comparable and the results from the simulations are more relevant for comparison of the models than the actual estimate.
It is generally difficult to pool results from different study occasions and in this case the routes of administration and activation of the baseline response were different in the two studies. This was accounted for by allowing the rate constants for the irreversible binding of omeprazole to be different in the two studies (kome1 andkome2), assuming that the availability for the drug to the enzyme may be different, depending on route of administration. The plasma protein binding of omeprazole is 90 and 95% in the dog and human, respectively (Regårdh et al., 1985). Being a weak base, omeprazole accumulates in the acidic interior of the parietal cell, i.e., there is a high extraction of omeprazole into the parietal cell. The 2-fold difference in free fraction was therefore not corrected for and total plasma concentrations were used in all modeling, but simulations were done to study the impact of potential differences in protein binding on the response by varyingkome (Fig. 7A). In addition, in the long-term experiments the histamine doses were chosen to give a stable acid secretory response that was 40 to 60% of the maximal response. In the short-term experiments the acid secretion was stimulated to a stable level of 80% of maximal secretion. This may result in differences in k1 and/ork2. When the back-flow model was parameterized with different sets of k1 andk2 for the two studies and with one commonkome, a better fit to the data was obtained. However, the precision of the k1and k2 parameters for the i.d. experiments was low. In Fig. 7B. the predicted response-time course of the scaled-up human model when different k1values were estimated for the i.v. and i.d. doses is shown superimposed on observations from Lind et al. (1983). Different stimulation of the acid secretion was simulated (Fig. 7B) by changingk1 because the degree of stimulation of acid secretion may vary between dogs and humans.
In conclusion, by combining pharmacodynamic data for omeprazole from several sources, it was possible to discriminate between drug characteristics and turnover characteristics for the enzyme. The model was able to describe both a rapid onset of inhibition and a slow recovery, as observed after administration of high doses or extended exposure to omeprazole, and a rapid recovery as observed after low doses. The model predicted the previous observations that the maximum inhibitory effect correlates with AUC, and that the apparent AUC50 value is lower on day 6 compared with on day 1. To validate the model a long-term exposure with response measured also during the washout would be optimal.
Acknowledgment
We thank Björn Holstein for helpful discussions regarding the pharmacologic action of omeprazole.
Footnotes
-
Send reprint requests to: Angela Äbelö, AstraZeneca R&D Mölndal, S-431 83 Mölndal, Sweden. E-mail:angela.abelo{at}astrazeneca.com
- Abbreviations:
- i.d.
- intraduodenal
- AUC
- area under the plasma concentration curve
- V
- volume of distribution
- F
- bioavailability
- CL
- clearance
- Received March 17, 2000.
- Accepted August 3, 2000.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}