


Abstract
Activation of G protein-coupled receptors (GPCRs) may bring about their disappearance from the cell membrane, and it is commonly accepted that G protein-coupled receptor kinases (GRKs) play a key function in this mechanism. Opioid receptors belong to the family of GPCRs and are substrates of GRKs. We examined the fate of δ- and μ-opioid receptors and GRK2 and 3 in living cells during the process of receptor sequestration, using laser scanning microscopy. For visualization purposes, receptors and kinases were tagged at their respective C terminus with a fluorophore. The opioid receptors were fused to enhanced green fluorescence protein (EGFP), and the GRKs were linked to red fluorescence protein (DsRed). The cDNAs of these constructs served for transfection of human embryonic kidney 293 cells and neuroblastoma-glioma hybrid cells (NG 108-15), respectively. We report that activation of δ-opioid-EGFP receptors triggers a rapid translocation of cytosolic GRK-DsRed toward the cell membrane, which in turn releases vesicles carrying both green fluorescent δ-receptors and red fluorescent GRKs. Phosducin, a Gβγ scavenger, completely prevents translocation of GRKs and the formation of vesicles. In analogous experiments with μ-opioid receptors fused to EGFP, we observed that receptor activation also discharges green fluorescent vesicles. In contrast to δ-receptors, μ-receptors failed to trigger accumulation of GRK2 or 3 at the membrane, and no cointernalization of μ-opioid receptors with GRK2 or 3 was observed. The results suggest that δ-opioid receptors, but not μ-receptors, cointernalize with GRK2 or 3.
G protein-coupled receptors (GPCRs) represent transmembrane proteins that mediate signals to the interior of cells. Activation of these receptors by extracellular ligands triggers their immediate phosphorylation, followed by uncoupling from intracellular G proteins and sequestration (Carman and Benovic, 1998). It is likely that the strength of receptor activation controls the intensity of receptor phosphorylation (Zhang et al., 1998) by enzymes termed G protein-coupled receptor kinases (GRKs). The GRKs consist of at least six related isoenzymes (GRK1–6), which transfer phosphate groups onto serine and threonine residues located close to the C terminus of receptor protein (Pitcher et al., 1998). GRKs mainly reside in the cytosol (Sterne-Marr and Benovic, 1995) and are suggested to translocate immediately upon GPCR-activation to the plasma membrane where they anchor at Gβγ subunits (Lefkowitz, 1993) to phosphorylate agonist-occupied receptors. Thereafter, GRKs are suggested to dissociate from Gβγ and redistribute to the cytosol. Arrestin will bind to the phosphorylated receptor to terminate signal transmission and to initiate receptor internalization (Carman and Benovic, 1998; Reiter et al., 2001).
Opioid receptors are members of the GPCR family (Cox, 1993). Their activation triggers the dissociation of inhibitory acting Gi/o proteins, resulting in attenuation or even termination of transmembrane signaling, a process defined as receptor desensitization (Ferguson, 2001). However, agonist-occupied receptors may undergo endocytosis followed either by down-regulation or their recycling to the cell membrane (Carman and Benovic, 1998). These mechanisms are differently affected both by the individual intrinsic activities of agonists (Yabaluri and Medzihradsky, 1997; Kovoor et al., 1998) and by the composition of GRKs with which the cells are equipped (Ferguson, 2001). Notably, the opiate agonist morphine fails to induce μ-opioid receptor internalization but will gain the capacity to sequester receptors as the cytosolic concentration of GRK2 is increased (Roettger et al., 1997; Zhang et al., 1998). Furthermore, the multiplicity of opioid receptors on the one hand and the at least six GRKs on the other suggests distinct interactions between these entities. Indeed, μ-opioid receptors are controlled by GRK2 (Zhang et al., 1998; Ferguson, 2001), GRK3 (Carman and Benovic, 1998; Hurle, 2001), and even by GRK6 (Hurle, 2001). δ-Opioid receptors are phosphorylated by GRK2 (Schulz et al., 1998b; Harrison et al., 1998), GRK3 (Carman and Benovic, 1998), and GRK5 (Harrison et al., 1998). Additional information links the function of GRKs to chronic opioid effects, as an increased abundance and activity of distinct GRKs was communicated for the state of opioid dependence and tolerance (Terwillinger et al., 1994; Kovoor et al., 1998; Ozaita et al., 1998;Hurle, 2001). Thus, our present knowledge favors the notion that GRKs account for the phosphorylation of activated opioid receptors, but there is no indication that the action of a distinct GRK is confined to a specific opioid receptor type.
Although our knowledge regarding phosphorylation and internalization of GPCRs has been derived mainly from experiments with cell fragments, the present study was designed to explore more closely the interrelation between activated opioid receptors and certain GRKs during the process of receptor sequestration in living cells. For this reason opioid receptors were fused with enhanced green fluorescence protein (EGFP;Heim et al., 1995), and GRKs were linked to red fluorescence protein (DsRed; Wall et al., 2000). The fusion proteins were expressed in cell lines (HEK 293, NG 108-15) and confocal laser microscopy was applied to track the fate of green fluorescent opioid receptors and red fluorescent GRKs in life cells. The major outcome of this study is the demonstration that activated δ-opioid receptors and receptor kinases 2 and 3 cointernalize. In contrast, internalization of μ-opioid receptors was not found accompanied by any GRK under investigation.
Materials and Methods
Chemicals.
The biochemical reagents were of analytical grade and purchased from Calbiochem (Deisenhofen, Germany) and Sigma (Taufkirchen, Germany). Opioid receptor ligands deltorphin II, [d-Ala2,N-Me-Phe4,glycinol5]-enkephalin (DAMGO), [d-Pen2,d-Pen5]-enkephalin (DPDPE), and [d-Ala2,d-Leu5]-enkephalin (DADLE) were from Bachem (Heidelberg, Germany), sufentanil was from Janssen Pharmaceuticals (Beerse, Belgium), and naloxone was from DuPont (Geneva, Switzerland). Guanosine-5′-O-(thio)-triphosphate (GTPγS) was from Sigma. The Taq-DNA polymerase was purchased from New England Biolabs (Schwalbach, Germany), G418 sulfate was from Calbiochem (Bad Soden, Germany), and the restriction enzymes were purchased from MBI Fermentas (St. Leon-Rot, Germany). The enzyme inhibitor Complete came from Roche Molecular Biochemicals (Mannheim, Germany).
Radio-Labeled Tracers.
125I-cAMP (2000 Ci/mmol) was obtained from Amersham (Braunschweig, Germany) and [3H]diprenorphine (32 Ci/mmol) was purchased from PerkinElmer Life Sciences (Dreieich, Germany).
Cell Culture Reagents.
Chemicals were purchased from Invitrogen (Karlsruhe, Germany), fetal calf serum from PAN (Nürnberg, Germany), and cell culture material from NUNC GmbH & Co. KG (Wiesbaden, Germany).
Antibodies.
The anti-cAMP antiserum was from Bio-Yeda (Rehovot, Israel), the anti-DsRed antibody from CLONTECH (Heidelberg, Germany), and all other antibodies were delivered from Santa Cruz Biotechnology, Inc. (Heidelberg, Germany).
Cell Culture.
HEK 293 cells and neuroblastoma x glioma 108–15 cells were maintained in Dulbecco's modified Eagle's medium supplemented with 10% fetal calf serum as described (Ammer and Schulz, 1993). Experiments were conducted with cells at 60% confluency.
DNA Transfection.
Cells were transfected with Effectene Reagent (QIAGEN GmbH, Hilden, Germany) according to the manufacturer's recommended protocol and used for experiments after 48 h. Stable transfections of cells to express μ-opioid receptor-EGFP and δ-receptor-EGFP, respectively, were conducted according to standard procedures, using G418 selection.
Radioligand Binding.
Opioid receptor binding was conducted with freshly prepared cell membranes as detailed by Vachon et al. (1987). Membranes (100 μg of protein) were incubated for 30 min at 30°C with ligands (tritiated tracer) in the absence and presence of 1 μM cold opioid to define nonspecific binding.
Western Blots.
Protein bands were resolved as described (Ammer and Schulz, 1993). Samples were electrophoresed on 10% sodium dodecyl sulfate-polyacrylamide gels under reducing conditions. Markers provided size calibration, and proteins were electroblotted to nitrocellulose at 4°C. Membranes were washed with Tris-buffered saline containing 0.2% Tween 20, and incubated with the primary antibody overnight at 4°C. After appropriate secondary antibody steps (1 h, 25°C, 1:20,000) and rinsing, the blots were developed with enhanced chemiluminescence detection.
Protein Assay.
Protein was assayed by the method of Lowry et al. (1951), using bovine serum albumin as standard.
cAMP Assay.
A slightly modified method (Ammer and Schulz, 1997) was employed. Briefly, cells were seeded onto 96-well plates (4.8 × 104 cells/well), allowed to settle for 2 h at 37°C in supplemented Dulbecco's modified Eagle's medium. Cells were stimulated by 10 μM forskolin, and the assay was conducted in the presence of 0.25 mM 3-isobutyl-1-methylxanthine over 15 min at 37°C. Assays were conducted in duplicate.
Construction of Expression Vectors.
The general procedure for the construction of fusion proteins is described in a previous publication (Schulz et al., 1998b), using the CLONTECH vectors pEGFP, encoding the red-shifted variant of wt GFP (enhanced GFP, EGFP), and pDsRed. The cDNAs of individual opioid receptors and of GRKs were amplified by polymerase chain reaction, and after removal of the respective stop codon, they were inserted into the multiple cloning sites of pEGFP and pDsRed, respectively. The fusion constructs were sequenced (TopLab, Martinsried, Germany) to test for correctness and in frame cloning. The following expression constructs were generated: rat μ-opioid-EGFP receptor, mouse δ-opioid-EGFP receptor, human GRK6-DsRed, bovine GRK3-DsRed, and bovine GRK2-DsRed.
Confocal Microscopy.
Transfected cells were grown on coverslips (2.4 × 3.2 cm) for 2 days. For laser-guided confocal microscopy, cells were placed in a device (PeCon; Carl Zeiss GmbH, Jena, Germany), which allowed cell maintenance at 37°C and continuous CO2 flow to stabilize the pH at 7.4. The expression of fusion proteins varied between the transfected cells, that is the cells comprise a wide spectrum of fluorescence intensity. For screening of transfected cells we preferred clones of medium fluorescence intensity. Confocal images were collected with an inverted Zeiss LSM 510 microscope (63 × 1.3 oil-immersion Plan-Neofluar objective; Carl Zeiss GmbH). For excitation, the 488 nm argon ion laser was used, and the emission was collected with a band pass 505-530 filter that allowed simultaneous monitoring of EGFP-tagged receptors as well as DsRed-tagged GRKs (excitation 543 nm, emission long pass filter 560 nm) with no bleed through between channels. The generation of phosducin-EGFP fusion protein has been described (Schulz et al., 1998b). Images from EGFP and DsRed fluorescence patterns were processed as one-color images or merged as two-color overlays. The digitized images were prepared as graphics by use of Adobe Photoshop software (Adobe Systems, Mountain View, CA).
Results
The experiments conducted rely on the cellular expression of fluorescent opioid receptors and GRKs to enable their visibility in living cells. Receptors and GRKs were linked at their C terminus with rather large proteins (EGFP, DsRed), each consisting of about 30 kDa. It thus is an essential prerequisite to ensure the undisturbed biologic activity of these opioid receptors and GRKs fused with EGFP and DsRed, respectively.
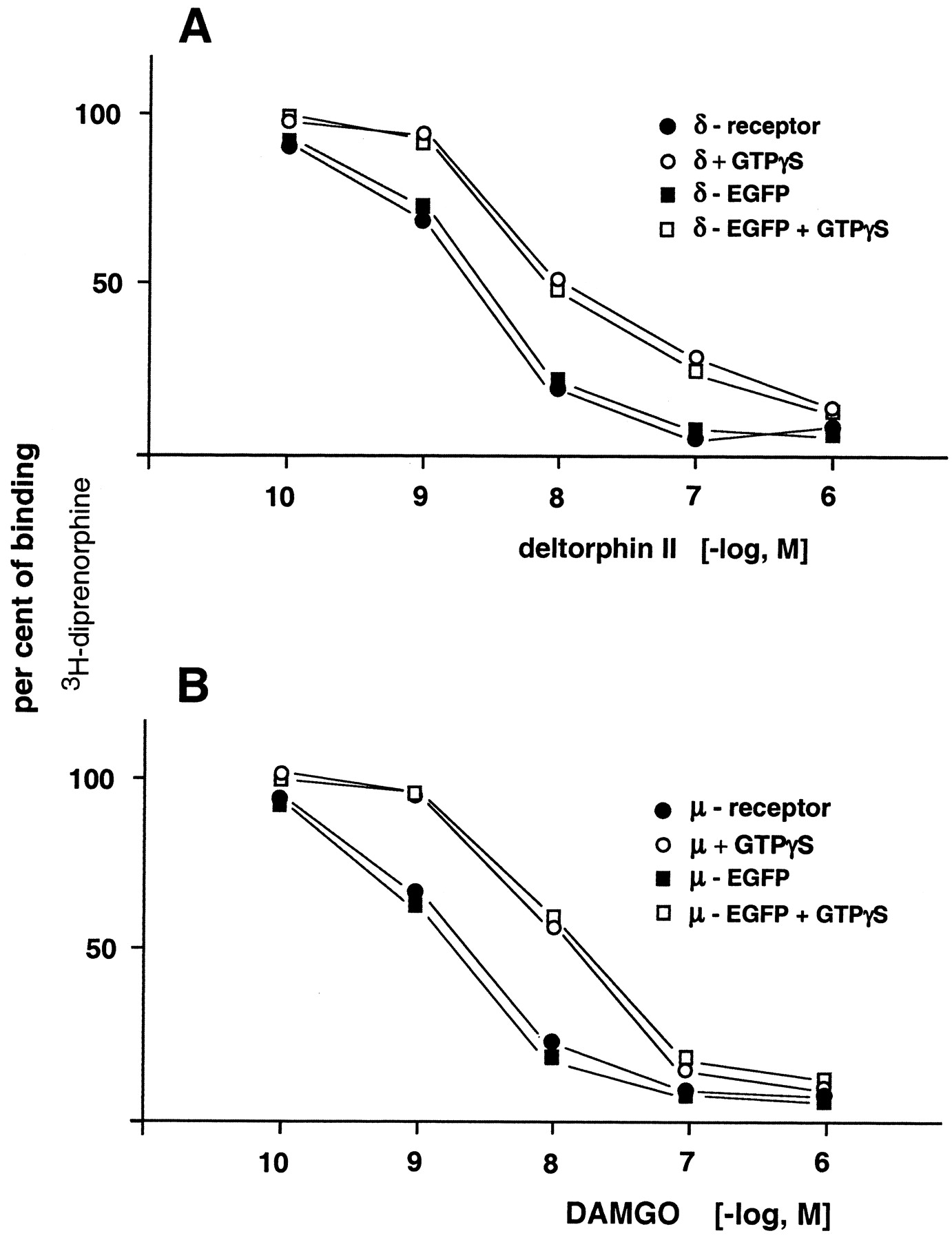
Since the receptor affinity of opioids is governed by G proteins (Cox, 1993), the effect of GTPγS on the potency of deltorphin II (δ-receptor agonist) and DAMGO (μ-receptor agonist) to displace [3H]diprenorphine from EGFP-labeled δ- and μ-opioid receptors, respectively, was assayed. Figure1A displays the effect of deltorphin II to displace the tritiated antagonist from δ-receptors and from δ-EGFP receptors in the absence and presence of GTPγS (10 μM). Regardless of the δ-receptor (wt or EGFP-tagged) examined, the stable GTP analog shifted the dose-response curve to the same degree. A similar outcome was observed with wt μ-receptors and μ-EGFP receptors, using DAMGO as a displacing ligand (Fig. 1B). The outcome of these experiments suggests that opioid receptor (δ- and μ-type) fused with EGFP remain fully controlled by their respective G proteins.

Displacement of [3H]diprenorphine by deltorphin II from δ-receptors and δ-EGFP receptors, respectively, over-expressed in HEK cells (A). Experiments were conducted with cell membranes in the absence (filled squares and circles) and presence (open symbols) of GTPγS (10 μM). Ordinate represents percentage of binding of tritiated tracer. The results represent means from three to four independent experiments. Standard errors of individual values were less than 10%. B, analogous experiments conducted with μ- and μ-EGFP-opioid receptors and GTPγS, using DAMGO as a displacing ligand. The data represent means from three independent experiments. Standard errors of individual values were less than 10%.
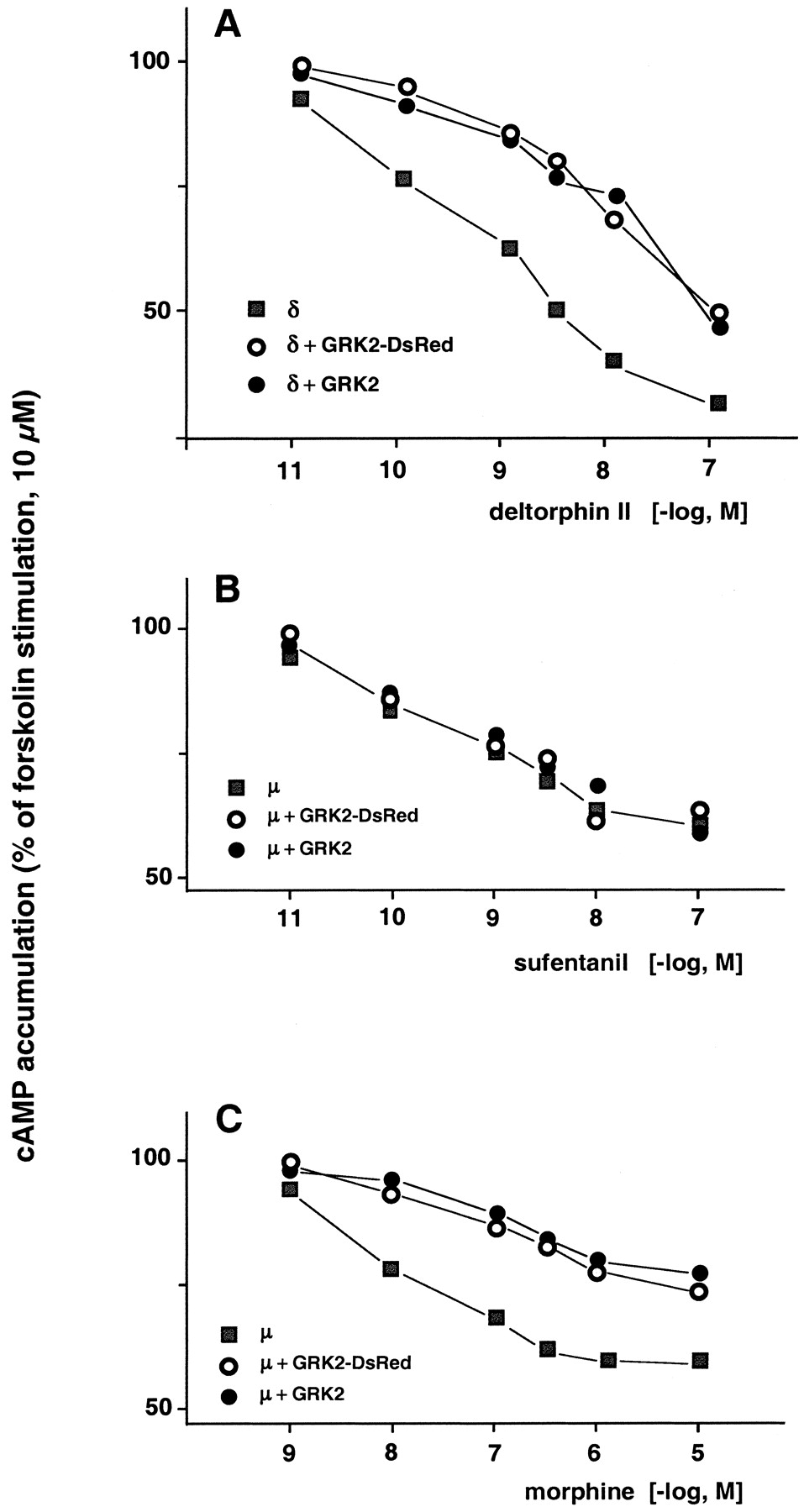
Tests for functionality of GRK-DsRed constructs made use of the finding that over-expression of GRK2 attenuates opioid-induced inhibition of adenylyl cyclase (Zhang et al., 1998). Figure2A displays the inhibitory potencies of deltorphin II on cAMP accumulation of forskolin-stimulated HEK cells transfected to permanently express wt δ-receptors only or δ-receptors together with GRK2 and GRK2-DsRed, respectively. Cells expressing δ-receptors only exhibit a half-maximal inhibition of forskolin-stimulated adenylyl cyclase at about 1 nM deltorphin II. Coexpression of opioid receptors and GRK2 attenuates the deltorphin II-induced inhibition of adenylyl cyclase activity. An identical decline of opioid-induced inhibition of cyclase activity is seen in cells over-expressing GRK2-DsRed. The experimental design was extended to examine the function of GRK3 and GRK3-DsRed in δ-receptors expressing HEK cells. The outcome (data not given) paralleled the findings with GRK2 and GRK2-DsRed that is the opioid exhibits a decreased inhibitory activity on adenylyl cyclase in these cells. Over-expression of GRK6 and GRK6-DsRed, respectively, in δ-receptor carrying HEK cells entirely failed to alter the potency of deltorphin II to inhibit forskolin-stimulated cAMP generation (data not given). The outcome of these experiments suggests that GRK2 and 3 as well as the corresponding DsRed fusion proteins own a similar potency to affect the function of δ-opioid receptors. The same approach was employed to test GRK2/3/6 and their DsRed fusion constructs on HEK cells permanently expressing μ-opioid receptors. We observed responses depending on the μ-ligand examined. In cells transiently over-expressing GRK2 and its DsRed fusion protein, respectively, the ability of sufentanil to control adenylyl cyclase activity was left unaffected (Fig. 2B). An identical result was obtained in cells expressing GRK3 and GRK6, respectively (data not given). In contrast, the potency of morphine (Fig. 2C) and etorphine as well as DAMGO (data not shown) was clearly attenuated to inhibit adenylyl cyclase activity in HEK cells over-expressing GRK2/3 or the DsRed fusion protein (data not given). Finally, GRK6 as well as the corresponding DsRed construct failed to control cAMP synthesis in μ-receptor carrying HEK cells challenged with morphine, etorphine, and DAMGO, respectively, over a tested range from 1 to 1000 nM (data not given).

Effect of opioids on inhibition of forskolin-stimulated cAMP accumulation in HEK cells transfected to permanently express μ- and δ-receptors, respectively. Ordinate reflects cAMP accumulation as percentage of forskolin stimulation. Abscissa: opioid concentration. A, inhibition of cAMP accumulation by deltorphin II in HEK cells expressing δ-receptor (squares) and of HEK cells over-expressing in addition GRK2-DsRed (circles) or GRK2 (filled circles). Individual values represent means of three independent experiments. Standard errors were less than 10%. B, experimental conditions as described for A, but HEK cells over-expressed μ-receptors. Sufentanil was employed to test for the generation of cAMP. Each dose-response curve was calculated from at least four individual experiments. C, experimental conditions as described for B, but cells were exposed to increasing concentrations of morphine (n = 3).
We further examined the expression of endogenous cytosolic GRKs in wt HEK cells using the Western blot technique (Fig.3). Lane 1 exhibits immunoreactivity located at 79 kDa, indicating the presence of cytosolic GRK2 in naive cells. Over-expression of GRK2 reveals strong immunoreactivity migrating at 79 kDa (lane 3). Expression of GRK2-DsRed results in the detection of two bands, endogenous GRK2 at 79 kDa and the expected fusion protein at 110 kDa (lane 2). Noteworthy, the GRK2 antibody recognized a nonidentified band at about 90 kDa. This immunoreactive band appears unique for HEK cells because we failed to detect a similar band in the cytosol of NG 108-15 or Chinese hamster ovary cells using the same antibody. GRK3 was not detected in the cytosol of wt HEK cells (lane 4), whereas over-expression of this kinase type is readily seen (lane 5). Interestingly, the anti-GRK3 antibody did not detect the fusion protein GRK3-DsRed (lane 6). We thus employed the anti-DsRed antibody, which clearly revealed an immunoreactive band at 110 kDa, indicating the presence of the over-expressed fusion protein (lane 7). A search for GRK6 immunoreactivity was unsuccessful in cytosol of HEK cells (lane 8) but easily detected in GRK6 over-expressing cells (lane 9). The DsRed-fusion protein was uncovered only by use of the anti-DsRed antibody (lane 11). Notably, cell membranes (5 μg of protein) prepared from wt HEK cells display GRK6 immunoreactivity (data not given), indicating an association of GRK6 with the plasma membrane.

Western blot analysis of cytosolic proteins from wt HEK cells and of cells transfected with DNAs encoding GRK2, GRK2-DsRed, GRK3, GRK3-DsRed, GRK6, and GRK6-DsRed, respectively. Five micrograms of protein was loaded on each lane, except for lanes 7 and 11 (10 μg). Lanes 1 to 3 represent immunoreactive bands detected by anti-GRK2 antibodies: lane 1 reveals endogenous GRK2 at about 79 kDA (marker), migrating with corresponding immunoreactivity of cells transfected with DNA coding for GRK2 (lane 3). The antibody recognizes also immunoreactivity in cells transfected to express GRK2-DsRed (about 110 kDa, lane 2). The anti-GRK2 antibody detects immunoreactivity of unknown origin at about 90 kDa (lanes 1–3). The results with anti-GRK3 antibodies are given in lanes 4 to 6. Endogenous GRK3 was not detected (lane 4), whereas over-expression of the kinase was documented (lane 5). In cells over-expressing GRK3-DsRed the antibody failed to recognize its presence (lane 6), although the fusion protein was detected by the anti-DsRed antibody (about 110 kDa, lane 7). Lanes 8 to 10 display the results with anti-GRK6 antibodies. Endogenous GRK6 (about 66 kDa) was not detected in cytosol (lane 8), but over-expression of the enzyme was clearly identified (lane 9). The anti-GRK6 antibody failed to detect GRK6-DsRed (lane 10), whereas the anti-DsRed antibody revealed the presence of the fusion protein (lane 11). The results given are representative of at least four independent experiments.
Confocal Microscopy.
Confocal microscopy images of HEK cells permanently expressing δ-EGFP receptors were transfected to transiently coexpress GRK2-DsRed and DsRed, respectively (Fig.4). Figure 4A reveals that fluorescent GRK2-DsRed is homogeneously distributed in the cytosol of cells, sparing the nucleus. Figure 4B exhibits the coexpression of GRK2-DsRed and δ-EGFP receptors (green fluorescence) of the same cell. An identical distribution was obtained for GRK3-DsRed and GRK6-DsRed (figures not displayed). Regardless of the fusion protein examined, we observed no enrichment of red fluorescence at the membrane of naive cells. In contrast to the GRK-DsRed constructs, cells transfected to express DsRed display fluorescence all over the cell, including the nucleus (Fig. 4C), and Fig. 4D displays the distribution of fluorescent DsRed and δ-EGFP receptors.

Confocal microscopy images of HEK cells transfected with cDNAs encoding for GRK2-DsRed and for the δ-opioid receptor tagged with EGFP (images A and B). Frames C and D originate from cells cotransfected with cDNAs coding for DsRed and for the δ-EGFP opioid receptor. GRK2-DsRed is homogeneously distributed within the cytosol, sparing the nucleus (image A). Image B (overlay) represents the location of δ-EGFP-receptors, which clearly separate from GRK2-DsRed fluorescence. Image C exhibits DsRed fluorescence dispersed throughout the cell, including the nucleus. Superimposition of confocal images (DsRed and δ-receptor-EGFP) of the same frame is given (D). Identical distribution patterns compared to GRK2-DsRed were found for GRK3-DsRed and GRK6-DsRed. Each cell depicted stands for at least 10 cells studied.
Internalization of δ-Opioid Receptors.
Experiments were designed to examine the fate of both green fluorescent δ-receptors and red-labeled kinases expressed in the same cell. Figure5, A through I display real-time confocal laser images of an NG 108-15 cell transiently expressing the δ-opioid receptor-EGFP construct and GRK2-DsRed. Prior to exposure of cells to the opioid (0 min), green fluorescence (δ-receptors) is primarily associated with the cell membrane, whereas very few green vesicles are present in the cytosol (Fig. 5, A and B). The GRK-DsRed construct is homogeneously distributed in the cytoplasm (Fig. 5, A and C). Stimulation of the cell with deltorphin II (5 nM) time dependently triggers redistribution of red and green fluorescence. As soon as 1 min after receptor activation, red fluorescence accumulates at the cell membrane (Fig. 5F; compare to Fig. 5C), suggesting translocation of cytosolic GRK2 toward agonist-occupied δ-receptors. GRKs remain visible at the cell membrane for several minutes (not shown). During this period no significant changes in distribution of receptors (green fluorescence) were observed (Fig. 5, D and E). However, after 15 min of exposure to deltorphin II, green fluorescence is associated with vesicles concentrating in the cytoplasm (Fig. 5H). This finding is typical for processes characterizing opioid receptor internalization (Chu et al., 1997; Schulz et al., 1999b). Very surprisingly, we noticed an association of red fluorescence (GRK2-DsRed) with migrating vesicles (Fig. 5I). Superimposing the “green” (H) and “red” (I) images, fluorescence turned yellow (G), indicating that green fluorescent receptors and the red fluorescent GRK2 exist in an extremely close vicinity within the vesicles. Preliminary investigations did not support the notion that the close colocalization of fluorescent entities, resulting in yellow fluorescence when images I and H were merged, bears any relationship to the mechanism underlying resonance energy transfer (FRET). Separate vesicles of either red or green fluorescence are difficult to detect. This finding suggests that exposure of cells to deltorphin II brings about cointernalization of δ-receptors and GRK2. In fact, the first vesicles separating from the plasma membrane were monitored 3 to 5 min subsequent to deltorphin II challenge, and they invariably carried red fluorescent GRK and green fluorescent receptors (images not given). The single cell-based assays proved that cointernalization of opioid receptors and GRK2/3-DsRed is brought about also by DADLE (10 nM) and DPDPE (5 nM). Identical results were demonstrated using HEK cells (data not shown).

Confocal microscopy images of a single NG 108-15 cell (images A to I) and a single HEK cell (images J to R). The NG 108-15 cell was transfected with cDNAs coding for δ-opioid-EGFP receptors and for GRK2-DsRed, the HEK cell expresses green fluorescent δ-receptors and GRK3-DsRed. Images of living cells display the location of δ-receptor-associated fluorescence as well as GRK2/3-DsRed-associated fluorescence prior (0 min) and subsequent (1 min and 15 min) to exposure with 5 nM deltorphin II. The overlay images (A, D, G, J, M, and P) were obtained by merging the corresponding green fluorescent receptors and red fluorescent GRK. Confocal optical sections clearly demonstrate translocation of red fluorescence (GRK2/3-DsRed) toward the cell membrane within 1 min after exposure to the opioid (images F and O). After this period, no internalization of receptor-associated fluorescence was monitored (images D, E and M, N). Fifteen minutes after drug exposure, an accumulation of vesicles in the cytosol is apparent, carrying green fluorescence (δ-receptors, images H and Q) and red fluorescence (GRK2, images I and R). Merging images H and I as well as Q and R reveals the coincidence of δ-receptors and GRK2/3 (G and P). Under these conditions, fluorescence of vesicles turns yellow, indicating a very close colocalization (below the optical resolution limit) of the fluorescent entities. Arrowheads in G, H, and I indicate identical vesicles (also in P, Q, and R). The two cells monitored are representative of at least 15 experiments with an identical outcome.
Figure 5, J to R, displays the response of a HEK cell stably expressing the δ-receptor–EGFP construct and transiently GRK3-DsRed. The confocal images J to L demonstrate the cellular distribution of red (GRK3) and green (δ-receptor) fluorescence prior to deltorphin II (5 nM) challenge (0 min), images M to O reflect the situation after 1 min, and P to R after 15 min of continuous deltorphin II exposure. The results confirm that opioid receptor activation triggers rapid translocation of GRK3-DsRed to the cell membrane (Fig. 5O), which is followed by the generation of vesicles cointernalizing GRK3-DsRed and green fluorescent δ-receptors. Superimposing the images Q and R reveals the development of yellow vesicles, again indicating a close colocalization of the fluorescent fusion proteins.
To strengthen the notion that activated δ-opioid receptors and GRK2 cointernalize, HEK cells permanently expressing δ-receptors were transfected to express GRK2-DsRed (Fig.6A). Activation of unlabeled δ-receptors by deltorphin II (5 nM) is expected to translocate cytosolic GRK2-DsRed toward the cell membrane followed by an internalization of receptors together with GRK2-DsRed. The experimental results clearly meet the prediction as documented by images B (translocation of GRK2) and C (formation of red vesicles). The lower panel of Fig. 6 (d to F) reflects the outcome of the control experiment conducted with cells coexpressing the δ-EGFP receptor and DsRed (Fig. 6D). Deltorphin II (5 nM) triggers the internalization of green fluorescent δ-receptors as indicated by the appearance of green dots (vesicles), which increase in number over time (images E and F). No association of red fluorescence (DsRed) with vesicles was observed. In addition, the green fluorescent vesicles failed to turn yellow when superimposed with DsRed images.

Trafficking of GRK2-DsRed in δ-receptor expressing HEK cells. Confocal images were taken prior (0 min, image A) and subsequent to deltorphin II (5 nM). One minute after opioid challenge an accumulation of red fluorescence (GRK2-DsRed) is observed at the cell membrane (image B, arrowheads). After 15 min (image C, arrowheads) red vesicles accumulate in the cytosol, indicating cointernalization of GRK-DsRed and δ-opioid receptors. The confocal images D to F originate from a HEK cell coexpressing δ-EGFP-receptors and DsRed. Deltorphin II (5 nM) strongly induces the internalization of vesicles carrying the green fluorescent receptors (images E and F, arrowheads). DsRed does not cointernalize with δ-EGFP-receptors, as the vesicles remain “green”. Each cell is representative of three experiments.
Phosducin has been demonstrated to inhibit opioid receptor internalization (Schulz et al., 1999a), most likely by competing with GRKs for Gβγ subunits at the cell membrane (Schulz, 2001 and references therein). HEK cells permanently expressing δ-opioid receptors were thus cotransfected to express phosducin-EGFP and GRK2-DsRed. Figure 7 exhibits confocal images of a transfected cell prior (0 min) and subsequent to the exposure of 10 nM deltorphin II. Apparently, in the presence of phosducin, the opioid fails to induce an accumulation of red fluorescence at the cell membrane (2 min) and the formation of red vesicles (GRK2-DsRed) is completely prevented.

The effect of phosducin on GRK2-DsRed trafficking. HEK cells permanently expressing δ-receptors were transfected to transiently express phosducin-EGFP and GRK2-DsRed. Confocal microscopy images are presented prior (0 min) and subsequent to 5 nM deltorphin II exposure (2 and 20 min). Green fluorescent phosducin is expressed all over the cell, including the nucleus, whereas GRK2-DsRed is confined to the cytosol. Phosducin competes with GRK2 for cell membrane-associated Gβγ subunits. In the presence of phosducin, the opioid fails to induce accumulation of GRK2-DsRed at the cell membrane (2 min), and it prevents internalization of opioid receptors as is indicated by the absence of vesicles carrying GRK2-DsRed (“red” vesicles). The cells presented stand for the outcome of three experiments.
Experiments were conducted also to investigate the effect of GRK6-DsRed on internalization of δ-receptors expressed in HEK cells. Challenge of cells with deltorphin II (5 nM), DADLE (10 nM), and DPDPE (10 nM), respectively, revealed no hint of translocation of GRK6-DsRed or the formation of GRK6-DsRed-containing vesicles.
Internalization of μ-Opioid Receptors.
The demonstrated cointernalization of δ-receptors and GRK2/3 caused us to conduct analogous studies with the μ-opioid receptor. HEK cells were transfected to coexpress the μ-receptor-EGFP construct and GRK2-DsRed. As expected, green fluorescent receptors are located mainly in the cell membrane (Fig. 8, A and B), and the kinase was identified in the cytoplasma (images A and C; 0 min). Stimulation of cells with sufentanil (5 nM) accounts for μ-receptor sequestration within 20 min as indicated by the formation of numerous green vesicles in the cytoplasma (images G and H). In contrast to the findings with the δ-opioid receptor, sufentanil (2, 10, and 20 nM) failed to trigger an accumulation of red fluorescent GRK2 at the cell membrane (images D and F), and we never observed the formation of red vesicles (images G and I). The μ-receptor agonists DAMGO (1 μM), etorphine (10 and 100 nM), and endomorphin 1 (50 nM) brought about identical results. The same observations were made when NG 108-15 cells were transfected to coexpress the μ-receptor-EGFP and GRK2-DsRed (data not given).

Real-time imaging of a single HEK cell coexpressing μ-opioid receptor (EGFP-labeled) and GRK2-DsRed. Confocal images were taken prior to (0 min) and after exposure to sufentanil (5 nM). The localization of green fluorescent μ-receptors (B) and of red fluorescent GRK2 (C) is given prior to sufentanil challenge. Images A, D, and G were obtained by merging the corresponding images. Two minutes after sufentanil exposure neither translocation of GRK2-DsRed toward the cell membrane (image F) nor changes in the distribution of μ-receptors were observed (E). Monitoring the cell after 20 min of drug exposure reveals a strong internalization of receptors (enrichment of green vesicles; H), but absolutely no formation of vesicles carrying red fluorescence (GRK2-DsRed) was seen (I). Superimposition of images H and I uncover that vesicles remain green (G), confirming the absence of closely associated red fluorescent entities (GRK2-DsRed) and green fluorescence (μ-EGFP-receptor). The bright red spot in the center of the images is believed to represent the Golgi apparatus. The experimental outcome stands for at least 30 observations with sufentanil and a similar number of experiments conducted with etorphine and DAMGO both in HEK and NG cells.
Further experiments were conducted with HEK cells coexpressing GRK3-DsRed and the μ-receptor-EGFP construct. Exposure of cells to sufentanil (1, 5, and 10 nM) strongly internalized the μ-receptor construct but failed to translocate detectable amounts of GRK3-DsRed toward the membrane or to trigger the formation of vesicles containing red fluorescence. Experiments with etorphine (10 and 100 nM), DAMGO (1 μM), and endomorphin 1 (0.5 μM) strongly internalized receptors when expressed in HEK cells, but translocation or even internalization (red vesicles) of GRK2-DsRed or GRK3-DsRed was not observed.
HEK cells coexpressing μ-receptor-EGFP and GRK6-DsRed were also examined. Exposure of cells to etorphine (1 μM) or DAMGO (10 μM) failed to uncover any interaction between the opioid receptor and the kinase during the process of internalization.
Morphine-Stimulated Internalization of μ-Opioid Receptors.
Activation of μ-receptors may trigger their sequestration but this receptor type fails to cointernalize with GRK2/3-DsRed. It is necessary to rule out that this failure is caused by an inability of GRK-DsRed to combine with the μ-receptor fused to EGFP. We thus made use of the finding that morphine fails to internalize μ-receptors but will do so upon over-expression of GRK2. Figure 9demonstrates that HEK cells expressing the μ-receptor-EGFP construct (confocal image A) fail to internalize these receptors when challenged with morphine (10 μM; Fig. 9B). In cells expressing GRK2-DsRed (images C, E, and G), morphine exposure (10 μM) efficiently triggered an internalization of μ-receptors (Fig. 9, F and H). Again, we never observed translocation of GRK2-DsRed during the course of morphine exposure, and the internalized vesicles did not exhibit red fluorescence (Fig. 9, D and H). The experiment supports the concept that over-expression of GRK2 (here GRK2-DsRed) clearly affects the mechanisms responsible for μ-receptor-EGFP internalization. Experiments conducted with HEK cells expressing GRK3-DsRed instead of GRK2-DsRed brought about identical results when exposed to morphine (data not given).

Confocal microscopy of HEK cells expressing μ-opioid receptors tagged with EGFP. Image A uncovers the localization of opioid receptors mainly at the cell membrane. Exposure of the cell to morphine (10 μM) largely fails to affect receptor distribution (B). A different HEK cell was transfected to express μ-EGFP-receptors as well as GRK2-DsRed (C to H). The location of receptors and GRK2 is given in images C and E prior to morphine challenge (superimposing C and E results in image G). Fifteen minutes after morphine (10 μM) the cell displays a strong internalization of μ-receptors (green vesicles; F, arrowheads), but no vesicles carrying red fluorescence (D and H) were observed. Moreover, the green vesicles (F, arrowheads) do not turn yellow (H) when images were merged. The experimental outcome stands for more than 15 cells examined.
Discussion
Activation of GPCRs, including μ- and δ- opioid receptors, triggers the dissociation of heterotrimeric G proteins in Gα- and Gβγ subunits (Lefkowitz, 1993; Carman and Benovic, 1998). The liberated Gβγ subunits associate with the cell membrane, where they serve as an anchor for receptor phosphorylating kinases (Ferguson, 2001). Subsequent to phosphorylation, the GRKs are believed to dissociate from their substrate (Reiter et al., 2001; Ferguson, 2001), and the phosphorylated receptor eventually cointernalizes with arrestin (Carman and Benovic, 1998). So far, the fate of GRKs in this ongoing process is unknown. The results reported here may add to our knowledge regarding the fate of GRKs as well as of GPCR internalization, specifically of δ- and μ-opioid receptors. We found that activation of δ-opioid receptors triggers translocation of GRK2 and 3 toward the cell membrane, which is followed by cointernalization of δ-opioid receptors and GRKs. On the other hand, stimulation of μ-opioid receptors does not result in accumulation of GRKs at the plasma membrane and no cointernalization of μ-receptors with GRK2 or 3 was observed. In addition, we found no hint for GRK6 to affect the function of δ- or μ-opioid receptors.
Internalization of GPCRs is linked to the formation of vesicles formed from rather large fragments of the cell membrane (Carman and Benovic, 1998). The components involved in transmembrane signaling, e.g., opioid receptor types, G proteins, and Gβγ-associated GRKs, represent functional units of close spatial link. The membrane-located proteins may comprise Gβγ subunits of heterogeneous composition (Mueller and Lohse, 1995; Mueller et al., 1997) as they originate from different inhibitory acting G proteins. Indeed, multiple opioid receptors are known to interact with multiple G proteins (Cox, 1993), resulting in the release of distinct combinations of Gβγ dimers. In addition, δ-opioid receptors combine with different G proteins, but Gi1 is more efficiently activated than Go1 (Moon et al., 2001). Thus, depending on the type of GPCR activated by the ligand, specific compositions of Gβγ subunits are released, and these subunits are likely to bind to the cell membrane (Daaka et al., 1997; Krupnick and Benovic, 1998). The distinct Gβγ subunits bound to the membrane are faced by GRKs translocated from the cytosol toward the cell membrane upon receptor stimulation. We have demonstrated here that activation of δ-opioid receptors triggers within seconds translocation of cytosolic GRKs to the cell membrane. They are believed to bind to specific Gβγ units and remain accumulated there for several minutes. During this period, larger areas of the cell membrane form vesicles equipped with green fluorescent receptors, arrestin (Carman and Benovic, 1998), and probably with Gβγ subunits. It is highly suggestive that the association of vesicles with red fluorescent GRKs is the consequence of a strong interaction of the kinase with Gβγ (Mueller and Lohse, 1995). This notion is supported by the experimental outcome with phosducin-EGFP. There is considerable evidence documenting that phosducin competes with GRK2/3 for Gβγ (Schulz, 2001), preventing both the accumulation of GRK2/3-DsRed at the cell membrane and the formation of vesicles. The concept implicates that GRK2/3 tightly bind to their membrane-associated anchor (Gβγ) during the entire process of sequestration. We demonstrated the appearance of yellow fluorescence, when red fluorescent GRKs and green fluorescent receptors were merged. This observation suggests that GRK/Gβγ and δ-receptors are separate entities but very closely located during the process of internalization with distances below the limit of optical resolution.
Cointernalization of GRK2/3 and δ-receptors implicates that fractions of the kinases become neutralized upon receptor activation and during trafficking of vesicles. If so, reconstitution of kinase activities is expected when recycled from the vesicles. Whether this transient loss of availability of functional GRKs will affect overall function of the δ-receptor population probably depends on the concentration of GRKs in the cytosol, as documented here for the effect of morphine on μ-receptor internalization (Fig. 9). It is hypothesized that prolonged exposure of δ-receptors to an agonist will result in a reduced disposition of functional GRK2/3, which in turn may result in an improved δ-receptor signaling. Similar to our findings with opioid receptors, Ruiz-Gomez and Mayor (1997) reported cointernalization of GRK2 and excitatory acting β2-adrenergic receptors in HEK cells. A functional significance of their observation was not communicated.
Documentation of an unimpaired activity of GRK2/3 fused with DsRed was achieved when coexpressed with δ- or μ-opioid receptors controlling the generation of cAMP. In HEK cells carrying δ-receptors, over-expression of the tagged kinase clearly reduced the ability of deltorphin II to inhibit cAMP synthesis. Identical results were obtained when wt GRK2/3 were expressed in these cells. Analogous experiments conducted with μ-receptors revealed that morphine, etorphine, and DAMGO lost potency in cells over-expressing GRK2/3 or the corresponding DsRed-tagged kinases to inhibit forskolin-stimulated adenylyl cyclase activity. We included sufentanil in these studies as this opiate strongly internalizes μ-receptors. Interestingly, the potency of sufentanil to inhibit cAMP generation was not affected by over-expression of any of the kinases tested. Whether this unexpected observation relates to the extreme potency of the opiate, suggesting a full pharmacologic effect at a very low fractional occupation of opiate receptors, remains to be seen.
In contrast to the observation with δ-receptors, μ-receptor activation fails to accumulate GRK2/3-DsRed at the cell membrane, and LSM failed to detect cointernalization of EGFP-tagged μ-receptors with GRK2/3-DsRed. Our data are not in conflict with the notion that internalization of μ-receptors requires their phosphorylation (Zhang et al., 1998), and even morphine does so in cells over-expressing GRK2 (Roettger et al., 1997) or 3. Thus, the failure to observe accumulation of GRKs at the cell membrane upon μ-receptor activation, and the absence of GRK internalization cannot be due to an inability of GRKs to phosphorylate μ-receptors. It has been suggested that μ-receptors interact with different inhibitory G proteins compared with δ-receptors (Carman and Benovic, 1998). This finding may imply that activation of δ-receptors generates different Gβγ subunits than those released by μ-receptor activation. These different Gβγ subunits may own different affinities to the individual GRKs. Thus, δ-receptor-related Gβγ subunits may tightly bind to GRK2/3, accounting for an accumulation of kinases 2/3 at the cell membrane and their subsequent cointernalization. In contrast, Gβγ subunits associated with μ-receptors less tightly and rather transiently bind with GRK2/3 to bring about receptor phosphorylation, causing the kinases to rapidly redistribute to the cytosol. This hypothesis could explain the failure of LSM to identify GRK2/3-DsRed at the membranes, and the lack of GRK2/3 and μ-receptors to cointernalize. This notion would also imply that the endogenous level of functional GRK2/3 remains unchanged during the course of μ-receptor internalization. This working hypothesis is not in conflict with suggestions that GRK2 displays distinct preferences to phosphorylate specific opioid receptor types (Zhang et al., 1999) and that GRK2 mediates desensitization of μ-opioid receptors (Li and Wang, 2001).
The experiments designed to examine whether GRK6 affects μ- and δ-opioid receptors failed to provide data in support of this notion. Although GRK6 was found to be associated with wt HEK cell membranes (unpublished observation; Lazari et al., 1999), even over-expression of this kinase did not affect μ- and δ-opioid receptor activity as judged by their ability to control adenylyl cyclase or internalization of opioid receptors.
Acknowledgments
We are grateful to the following persons for providing cDNAs: Drs. G. I. Bell (mouse δ-opioid receptor, Chicago, IL), J. L. Benovic (bovine GRK2 and 3, Philadelphia, PA), B. Moepps (GRK6, National Institutes of Health, accession no. 002082, Ulm, Germany), and L. Yu (μ-opioid receptor, Cincinnati, OH).
Footnotes
- Abbreviations:
- GPCR
- G protein-coupled receptor
- DADLE
- [d-Ala2,d-Leu5]-enkephalin
- DAMGO
- [d-Ala2,N-Me-Phe4,glycinol5]-enkephalin
- DPDPE
- [d-Pen2,d-Pen5]-enkephalin
- EGFP
- enhanced green fluorescence protein
- DsRed
- red fluorescence protein
- GRK
- G protein-coupled receptor kinase
- GTPγS
- guanosine-5′-O-(thio)triphosphate
- HEK
- human embryonic kidney
- LSM
- laser scanning microscopy
- wt
- wild-type
- Received August 30, 2001.
- Accepted October 31, 2001.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}