


Abstract
Increase in dopamine (DA) neurotransmission resulting from blockade of the DA transporter (DAT) after administration of cocaine is believed to play a major role in mediating its behavioral and reinforcing effects. Since it was hypothesized that drugs that block the DAT have cocaine-like behavioral effects, it was of interest to study in the present article the stimulant effects of cocaine on locomotor activity and on pattern of activation of DA neurotransmission in different DAergic terminal areas in rats and compare these effects with those of 4′-chloro-3α-(diphenylmethoxy)-tropane (4-Cl-BZT), a benztropine analog showing higher affinity for the DAT, but reduced behavioral effects compared with cocaine. Administration of cocaine resulted in a dose-dependent stimulation of locomotor activity and DA neurotransmission in the nucleus accumbens shell and core, dorsal caudate, and in the medial prefrontal cortex (PFCX) measured by microdialysis. At comparable doses, the effects of 4-Cl-BZT on DA levels in all brain areas except the PFCX were generally reduced compared with those of cocaine, as were the effects on locomotor activity. The differences in behavioral effects corresponded generally to differences between the drugs with regard to their stimulation of extracellular DA levels, although the mechanism(s) for the differences in extracellular DA may involve effects mediated by sites other than the DAT or differences in the efficiency of the two drugs in blocking DA uptake. Nonetheless, the present results suggest that the differences in behavioral effects between cocaine and 4-Cl-BZT are related to differences in their patterns of activation of DA transmission.
Blockade of the dopamine transporter (DAT), by cocaine, leads to increased dopamine (DA) neurotransmission resulting from accumulation of endogenous DA in the synaptic cleft and its increased availability for binding to DA receptors. DA neurotransmission is believed to directly mediate most of the behavioral and reinforcing effects of cocaine (Ritz et al., 1987; Kuhar et al., 1991; Callahan et al., 1997). Although other receptors/systems are involved in cocaine actions (e.g., norepinephrine and serotonin systems due to actions at their respective transporters; Ritz et al., 1990) and may contribute to its neurochemical and behavioral effects, DAT blockade is central to cocaine abuse (Ritz et al., 1987; Kuhar et al., 1991).
Among other novel molecules with affinity for the DAT, analogs of benztropine (BZT) possess a tropane ring, as does cocaine, and displace [3H]WIN 35,428 from its recognition site on the DAT, with many having higher affinity than cocaine (van der Zee et al., 1980; Newman et al., 1994). Despite high affinity for the DAT, BZT analogs are generally less effective than cocaine in stimulating locomotor activity in rodents and do not fully substitute for cocaine in rats trained to discriminate cocaine from saline (Katz et al., 1999, 2001; Newman and Kulkarni, 2002). Other studies have indicated that BZT analogs are less effective than cocaine in maintaining responding in self-administration procedures utilizing nonhuman primates (Woolverton et al., 2000, 2001). Thus under a variety of behavioral procedures, among different species, BZT analogs show a profile of behavioral effects that differs from that of cocaine, despite high affinity for the DAT. In these actions, BZT analogs differ from other compounds, for example, GBR 12909, bupropion, nomifensine, and mazindol, which show high affinity for the DAT (Baker et al., 1993). The reduced behavioral effects of BZT analogs compared with cocaine might be due to their different ability to increase DA neurotransmission compared with cocaine. For example, Tolliver et al. (1999) showed that the BZT analog 4′-chloro-3α-(diphenylmethoxy)-tropane (4-Cl-BZT) (Newman et al., 1994) and cocaine differently increase DA levels measured by microdialysis in the nucleus accumbens (NAC) in rats.
The mesolimbic dopaminergic pathway, consisting of ventral tegmental area neurons projecting to dopaminergic terminal fields in the NAC, is involved in many aspects of drug abuse (Di Chiara et al., 1999; Koob, 1999). Although less sharply defined boundaries have been found in primates (Haber and McFarland, 1999), in rodents the NAC has been divided into two anatomically and functionally well distinguished regions: 1) the shell, a ventromedial portion which belongs to the so-called extended amygdala and is suggested to play a prominent role in the reinforcing effects of drugs abused by humans; and 2) the core, a dorsolateral portion which is an integral part of the extrapyramidal motor system, mainly is involved in somatomotor and visceral functions (Koob, 1999; Zahm, 1999; but see Ikemoto, 2002; Sellings and Clarke, 2003). Differences in the sensitivity of activation of DA transmission in the shell and core subdivisions of the NAC after administration of drugs abused by humans (Pontieri et al., 1995, 1996; Tanda et al., 1997a) are thought to play an important role in the reinforcing effects of these drugs and in their abuse liability (Di Chiara et al., 1999; Koob, 1999).
Because in the study by Tolliver et al. (1999) microdialysis results for DA were collected from the NAC as a whole, it was of interest to examine the effects of 4-Cl-BZT on DA transmission in the shell and core compartments of the NAC. It was of further interest to compare the effects of cocaine and 4-Cl-BZT, which share their main pharmacological activity as inhibitors of DA uptake, but also possess other, nonoverlapping, pharmacological actions that might modulate their behavioral and neurochemical effects. For example, cocaine has similar affinities for DA and serotonin uptake sites (Katz et al., 2001), whereas 4-Cl-BZT has similar affinities for DAT sites and for muscarinic M1 receptors, but its affinity for serotonin uptake sites is about 150 times lower than that for DAT sites (Katz et al., 2001). DA transmission has been monitored in freely moving rats implanted with vertical microdialysis probes in four anatomically and functionally distinct brain DAergic terminal areas: 1) the NAC shell, 2) the NAC core, 3) the dorsal caudate, a major terminal site of the nigrostriatal DA system that plays an important role in the extrapyramidal motor functions, and 4) the medial prefrontal cortex (PFCX), the main terminal area of the mesocortical dopaminergic system which is involved in the control of environmental stimuli and stress or aversive-related behaviors.
Materials and Methods
Subjects. Male Sprague-Dawley rats (Taconic Farms, Germantown, NY), experimentally naive at the start of the study and weighing 300 to 350 g, were double-housed and had free access to food and water. All rats were housed in a temperature- and humidity-controlled room and were maintained on a 12-h light/dark cycle (lights were on from 7:00 AM–7:00 PM). Experiments were conducted during the light phase. Subjects used in this study were maintained in facilities fully accredited by the American Association for the Accreditation of Laboratory Animal Care (AAALAC), and all experimentation was conducted in accordance with the guidelines of the Animal Care and Use Committee of the Intramural Research Program, National Institute on Drug Abuse, National Institutes of Health, and the Guide for Care and Use of Laboratory Animals (National Research Council, 1996).
Locomotor Activity. Subjects were placed in square (40 × 40 cm) acrylic chambers that were placed inside monitors (Omnitech Electronics, Columbus, OH) equipped with light-sensitive detectors (photocells) spaced 2.5 cm apart along two perpendicular walls. Mounted on the opposing walls were infrared light sources directed at the photocells. One count of horizontal activity was registered each time the photocell was activated by interruption of the light source. Subjects were allowed to habituate for 120 min before injections were administered. At that point, subjects were injected i.p. with either saline, cocaine (1.0 to 30.0 mg/kg), or 4-Cl-BZT (3.0 to 30.0 mg/kg) and immediately returned to the apparatus. Horizontal locomotor counts were tabulated every 10 min after the 120-min habituation period for 180 (cocaine) or 300 min (4-Cl-BZT, saline). Each dose was studied in six subjects, and subjects were used only once.
Surgery. Under a mixture of ketamine and xylazine anesthesia (60.0 and 12.0 mg/kg i.p., respectively), rats were placed in a stereotaxic apparatus where the skull was exposed and a small hole drilled to expose the dura. Rats were then randomly implanted in the right or left brain side with a concentric dialysis probe (see below) aimed at the NAC shell or core, dorsal caudate, and medial PFCX, as described previously (Tanda et al., 1997a; Fig. 1) according to the rat brain atlas by Paxinos and Watson (1987) [uncorrected coordinates: shell, A = +2.0, L = ±1.1, V = 7.9; core, A = +1.6, L = ±1.9, V = 7.7; dorsal caudate, A = +1.0, L = ±3.0, V = 6.0; PFCX, A = +3.5, L =±0.6, V = 5.0; anterior (A), millimeters from bregma; lateral (L) millimeters from bregma; vertical (V) millimeters from dura].

Forebrain sections, redrawn from Paxinos and Watson (1987) showing the limits of the positions of the dialyzing portions of the microdialysis probes (superimposed rectangles). On each section, the anterior coordinate (measured from Bregma) is indicated. CPu, caudate putamen; ca, anterior commissure; cc, corpus callosum; Co, nucleus accumbens core; Sh, nucleus accumbens shell.
In Vivo Microdialysis. Concentric dialysis probes were prepared with AN69 fibers (Hospal Dasco, Bologna, Italy). Briefly, two 4-cm pieces of silica-fused capillary tubes (the inlet and outlet tubing of the probes) were inserted into a 6-mm capillary dialyzing fiber (closed by a drop of glue on the other side), with the inlet tubing set at about 0.1 mm from the closed end of the fiber and the outlet set at 2.0 (NAC and caudate probes) or 3.0 mm (PFCX probes) from the inlet tip. The open end of the dialysis membrane was then glued and the protruding two silica-fused tubing were inserted and glued into a 22-gauge stainless steel needle (2.4-mm length). The needle was then clipped to a CMA/10 clip (CMA/Microdialysis AB, Solna, Sweden) and mounted in a stereotaxic holder. The exposed dialyzing surface of the fibers, i.e., not covered by glue, was limited to the lowest 2.0 mm (NAC and caudate) or 3.0 mm (PFCX) portion of the probes. After the surgery, rats were placed in hemispherical CMA-120 cages (CMA/Microdialysis AB) equipped with overhead fluid swivels (Instech Laboratories Inc., Plymouth Meeting, PA) for connections to the dialysis probes and allowed to recover overnight.
About 24 h after probe implant, experiments were performed on freely moving rats in the same hemispherical home cages in which they recovered overnight from surgery. Ringer's solution (147.0 mM NaCl, 2.2 mM CaCl2, and 4.0 mM KCl) was delivered by a 1.0-ml syringe, operated by a BAS Bee Syringe Pump Controller (BAS Bioanalytical Systems, West Lafayette, IN), through the dialysis probes at a constant flow rate of 1 μl/min. Collection of dialysate samples (10 μl) started after 30 min, and samples were taken every 10 min and immediately analyzed, as detailed below. After stable DA values (less than 10% variability) were obtained for at least three consecutive samples (typically after about 1 h), rats were treated with drug or saline. Samples were taken every 10 min during the first 2 h after treatment and every 20 min thereafter, but only 10 of 20 μl collected in these 20-min samples were analyzed. Rats were used only once and have received only one drug-dose treatment.
Analytical Procedure. Dialysate samples (10 μl) were injected without purification into a high-performance liquid chromatography apparatus equipped with a MD 150- × 3.2-mm column, particle size 3.0 μm (ESA, Chelmsford, MA) and a coulometric detector (5200a Coulochem II; ESA Inc., Chelmsford, MA) to quantify DA. The oxidation and reduction electrodes of the analytical cell (5014B; ESA Inc.) were set at +125 and –125 mV, respectively. The mobile phase, containing 100 mM NaH2PO4, 0.1 mM Na2EDTA, 0.5 mM noctyl sulfate, and 18% (v/v) methanol (pH adjusted to 5.5 with Na2HPO4) was pumped by an ESA 582 (ESA Inc.) solvent delivery module at 0.60 ml/min. Assay sensitivity for DA was 2 fmol per sample.
Histology. At the end of the experiment, rats were euthanized by pentobarbital overdose, and brains were removed and left to fix in 4% formaldehyde in saline solution. Brains were then cut on a vibratome in serial coronal slices oriented according to the atlas by Paxinos and Watson (1987) to identify the location of the probes. In all the experiments, the location of the probes was verified. Figure 1 schematically shows typical locations of the dialyzing portion of the probes implanted in the PFCX, dorsal caudate, shell, and core. Although the probes were randomly implanted in the right or left brain side, all probe placements have been schematically reported only on one brain side in the figure. The brain sections are redrawn from Paxinos and Watson (1987), and the anterior coordinates (measured from bregma) for each brain area have been indicated. Only the experiments in which the probes were located in these areas have been considered and used for the effects shown in the present study.
Drugs. The drugs tested were 4-Cl-BZT, synthesized in our laboratories (Newman et al., 1994, 1995) and (–)-cocaine HCl (Sigma-Aldrich, St. Louis, MO). Drugs were dissolved in saline (0.9% NaCl) and were injected i.p. in a volume of 2.0 ml/kg. Injections of saline (2 ml/kg i.p.) served as vehicle controls.
Data Analysis. For the locomotor activity study, results were expressed as horizontal counts (instances of photocell activation). Two-way repeated measures analyses of variance (ANOVA) were performed on each test drug such that the effects of dose and time were assessed. Tukey's honestly significant difference post hoc tests provided pair-wise comparison information.
In the microdialysis study, results were expressed as a percentage of basal DA values. Basal DA values were calculated as the mean of three consecutive samples (differing no more than 10%) immediately preceding the first drug or vehicle injection. All results are presented as group means (±S.E.M.). Differences in basal levels of DA between different experimental groups within the same brain area, or between different brain areas, were analyzed by one-way ANOVA. Statistical analysis of those data were carried out using a two-way ANOVA (drug dose and time as factors) for repeated measures over time, with results from treatments showing overall changes subjected to post hoc Tukey's test.
The effects of both 4-Cl-BZT and cocaine (expressed in micromoles per kilogram) during the 30-min period of maximum stimulation of DA levels (0–30 and 30–60 min for cocaine and 4-Cl-BZT, respectively) were further analyzed by ANOVA and linear regression techniques to determine the doses and 95% confidence limits effective in producing either an increase in extracellular DA levels to 300% of basal values or an increase in horizontal activity by 100 counts (Snedecor and Cochran, 1967). The dose-effect data were further analyzed by standard parallel line bioassay techniques (Finney, 1964) to determine relative potencies of the two drugs. This analysis involves a one-way ANOVA which determines whether the slopes of the two dose-response curves are significantly different from parallel and fits a common slope to the two dose-response curves. It then compares the ratio of doses for a given effect to provide a value for relative potency. This value represents the dose of the 4-Cl-BZT in micromoles per kilogram equal to 1 μmol/kg cocaine. A significant relative potency difference is indicated when the 95% confidence limits for that ratio did not include 1.0. A significant effect of preparations in this analysis represents a difference in the mean response (weighted by N) for the two drugs, and as presently applied is reflective of differences in efficacy of the drugs. Changes were considered to be significant when p < 0.05.
Results
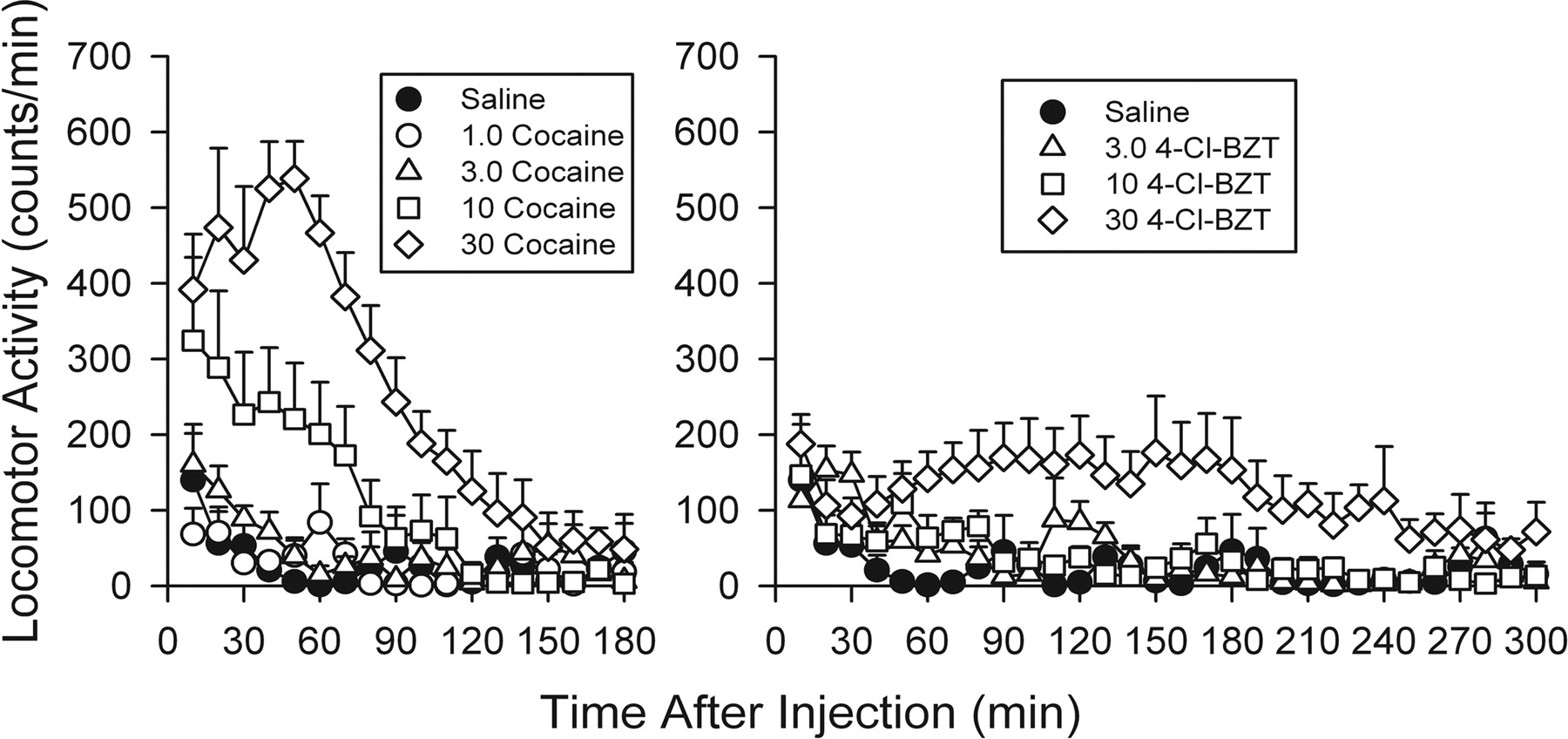
Effects of Administration of Cocaine or 4-Cl-BZT on Locomotor Activity. As has been demonstrated previously, cocaine increased ambulatory activity. At 10 mg/kg the maximum increase was obtained within the first 10 min after injection, whereas at the 30-mg/kg dose the maximum was observed at 40 to 50 min after injection (Fig. 2, left). Locomotor activity returned to control levels within 90 or 150 min of injection of 10 or 30 mg/kg, respectively. ANOVA indicated that the effects of both cocaine dose (F4,408 = 18.4; p < 0.001) and time after injection (F17,408 = 22.6; p < 0.001) were significant.

Time course of effects of cocaine (left panel) and 4-Cl-BZT (right panel) on locomotor activity in rats. Ordinates: horizontal locomotor activity counts after drug or vehicle administration in counts per minute. Abscissae: time since injection. Effects of several doses are shown (see symbol keys on each panel). Each point represents the average effect determined in five to six rats. Vertical bars represent S.E.M. Points represent average total counts for successive 10-min time periods up to 300 min (5 h) after injection.
A stimulation of locomotor activity was also obtained with 4-Cl-BZT. The increase was generally at its maximum between 90 and 170 min after injection (Fig. 2, right). Locomotor activity slowly returned and approached control levels by the end of the observation period. ANOVA indicated that the effects of both 4-Cl-BZT dose (F3,551 = 6.93; p = 0.002) and time (F29,551 = 5.94; p < 0.001) were significant. The locomotor stimulant effects of 4-Cl-BZT were significantly reduced compared with those of cocaine. Maximal stimulation of locomotor activity above vehicle levels was obtained 140 to 150 min after injection of 30 mg/kg, and the effect was approximately 176 (±74.8) counts per min (Fig. 2, right), which was less than the maximal value of 538 (±49.3) counts per min obtained 40 to 50 min after cocaine injection. There was a significant effect of preparations in this comparison of dose effects (F1,24 = 8.92; p = 0.006) indicating a difference in efficacy between the two drugs.
Effects of Administration of Cocaine or 4-Cl-BZT on Extracellular Levels of DA. Basal levels of extracellular DA (expressed as femtomole/10-μl sample ± S.E.M.) obtained in dialysates from four brain areas under investigation in the present study were: 47.3 ± 2.66 in dialysates from the accumbens shell (n = 42), 57.4 ± 3.24 in dialysates from the accumbens core (n = 36), 75.1 ± 3.89 in dialysates from the dorsal caudate (n = 38), and 9.76 ± 0.48 in dialysates from the PFCX (n = 39). No significant differences in basal levels of DA have been found among different experimental groups within each brain area. Significant differences have been found in basal extracellular levels of DA between PFCX and all of the other brain areas and between dorsal caudate and all other brain areas.
Saline administration did not significantly modify extracellular DA levels in dialysates from the NAC shell (data not shown). Cocaine administration (1, 3, 10, and 30 mg/kg) produced a dose-dependent and significant increases in DA levels in the NAC shell (two-way ANOVA, main effect dose, F4,23 = 83.6, p < 0.001; main effect time, F18,414 = 29.7, p < 0.001; time × dose interaction, F72,414 = 12.6, p < 0.001) (Fig. 3, left). Cocaine produced a rapid and relatively transient increase in DA levels at all doses tested. DA values reached maximum at about 20 min after cocaine administration, with the maximum increase reaching about 900% of basal values at the highest dose. DA levels returned to basal values after about 260 min from administration of the 30-mg/kg dose of cocaine (Fig. 3; left).

Time course of effects of systemic administration of cocaine (left panel) or 4-Cl-BZT (right panel) on extracellular levels of DA in dialysates from the NAC shell. Results are means with vertical bars representing S.E.M. of the amount of DA in 10-min dialysate samples, expressed as percentage of basal values, uncorrected for probe recovery. The N was 6, 5, 7, and 6 for cocaine doses 1, 3, 10, and 30 mg/kg, respectively. The N was 5, 5, and 4 for 4-Cl-BZT doses of 3, 10, and 30 mg/kg, respectively. Filled symbols = p < 0.05 compared with basal DA values.
Administration of 4-Cl-BZT (3, 10, and 30 mg/kg) produced a significant, dose-dependent increase of DA levels in dialysates from the NAC shell (two-way ANOVA, main effect dose, F3,14 = 64.0, p < 0.001; main effect time, F18,252 = 21.4, p < 0.001; time × dose interaction, F54,252 = 9.040 p < 0.001; Fig. 3, right). Administration of the lowest dose of 4-Cl-BZT (3 mg/kg) did not significantly modify basal values of DA (p = 0.94, N.S.). Administration of higher doses of 4-Cl-BZT (10 and 30 mg/kg) produced significant and prolonged (>5 h) increases in DA levels (Fig. 3; right). DA values reached maximum at about 60 to 90 min after 4-Cl-BZT administration, with maximum increases of 245 and 420% of basal values for 10 and 30 mg/kg, respectively. Over the course of the 300-min observation period there was a minimal decrease in DA levels from the obtained maxima.
Saline administration did not significantly modify extracellular DA levels in dialysates from the NAC core (data not shown), whereas cocaine (1, 3, 10, and 30 mg/kg) produced dose-related increases (two-way ANOVA, main effect dose, F4,16 = 98.4, p < 0.001; main effect time, F18,288 = 114, p < 0.001; time × dose interaction, F72,288 = 43.4, p < 0.001) (Fig. 4, left). The increase was rapid and transient and was obtained with all but the lowest dose (1 mg/kg) (post hoc, p = 0.97, N.S.). DA levels reached maximum at about 20 min after cocaine administration, with the maximum increase at about 600% of basal values at the highest (30 mg/kg) dose, which was two-thirds of that produced in the NAC shell (Fig. 4, left). DA levels returned to basal values after about 180 min from administration of the 30-mg/kg dose of cocaine.

Time course of effects of systemic administration of cocaine (left panel) or 4-Cl-BZT (right panel) on DA extracellular levels in dialysates from the NAC core. The N was 4, 4, 5, and 4 for cocaine doses of 1, 3, 10, and 30 mg/kg, respectively. The N was 5 for all doses of 4-Cl-BZT. All other details are as in Fig. 3.
Two-way ANOVA showed a dose-dependent effect over time on DA levels in the core after 4-Cl-BZT administration (3, 10, and 30 mg/kg) (two-way ANOVA, main effect dose, F3,15 = 142, p < 0.001; main effect time, F18,270 = 33.2, p < 0.001; time × dose interaction, F54,1068 = 26.7, p < 0.001) (Fig. 4, right). However, only the 30-mg/kg dose of 4-Cl-BZT significantly (post hoc, p < 0.05) increased DA levels in the core of the NAC. At this dose, 4-Cl-BZT produced a prolonged (>5 h) and significant increase in DA levels, which reached maximum at about 30 min after administration. The maximum increase was about 450% of basal values. There was a relatively constant rate of decrease from the maximum DA level obtained over the remaining 270-min course of the observation period (Fig. 4, right).
Saline administration did not significantly modify extracellular DA levels in dialysates from the dorsal caudate (data not shown), whereas the effects of cocaine were generally similar to those already shown for the shell and core of the NAC (Fig. 5, left). Cocaine (1, 3, 10, and 30 mg/kg) produced a graded dose-dependent increase in DA levels (two-way ANOVA, main effect dose, F4,19 = 34.2, p < 0.001; main effect time, F18,342 = 59.7, p < 0.001; time × dose interaction, F72,342 = 17.0, p < 0.001) that decreased over time. The lowest dose of cocaine (1 mg/kg) did not significantly modify DA levels (p = 0.98, N.S.), whereas transient increases in DA levels were obtained at each of the higher doses (Fig. 5, left). DA values reached maximum at 20 min after cocaine administration with the maximum increase approximately 400% of basal values at the 30-mg/kg dose. DA levels returned to basal values after about 100 min from administration of this dose of cocaine (Fig. 5, left).
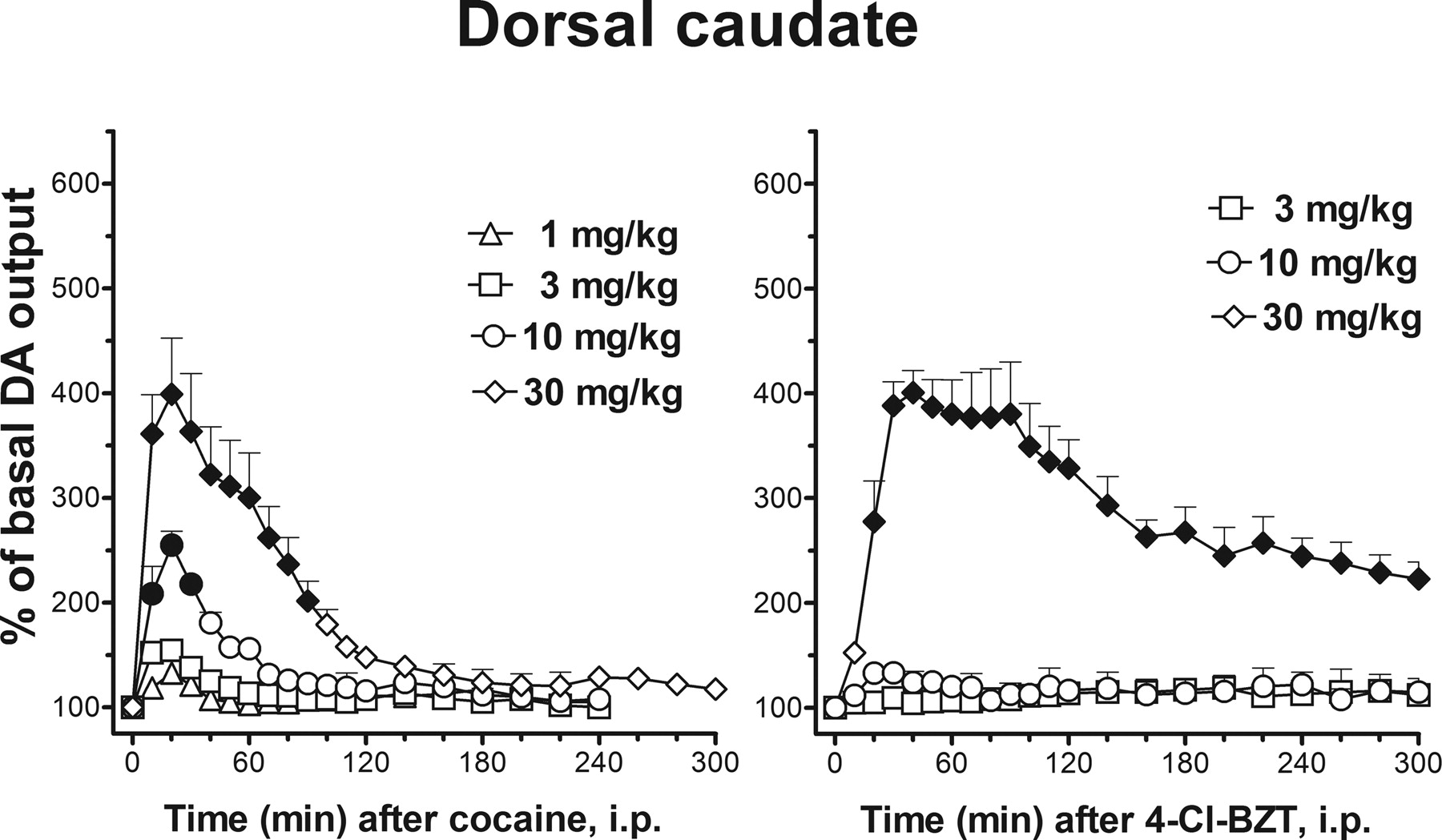
Time course of effects of systemic administration of cocaine (left panel) or 4-Cl-BZT (right panel) on DA extracellular levels in dialysates from the dorsal caudate. The N was 5, 6, 5, and 4 for cocaine doses of 1, 3, 10, and 30 mg/kg, respectively. The N was 6, 4, and 4 for 4-Cl-BZT doses of 3, 10, and 30 mg/kg, respectively. All other details are as in Fig. 3.
The dose-dependent increases in DA levels produced by 4-Cl-BZT (3, 10, and 30 mg/kg) in the dorsal caudate resembled those of rats implanted in the NAC core (Fig. 5, right; two-way ANOVA, main effect dose, F3,14 = 62.1, p < 0.001; main effect time, F18,252 = 19.9, p < 0.001; time × dose interaction, F54,252 = 18.2, p < 0.001). As in the NAC core, only the highest dose of 4-Cl-BZT (30 mg/kg) significantly increased (post hoc, p < 0.05) DA levels in dialysates from the dorsal caudate (Fig. 5; right). Administration of this dose produced a prolonged (>5 h) and significant increase in DA levels. The maximum increase of about 400% of basal values was reached at 40 min and was sustained for about 100 min after 4-Cl-BZT administration.
In the PFCX, as in the other areas, saline administration was without significant effects (data not shown), whereas cocaine administration (1, 3, 10, and 30 mg/kg) produced dose-dependent and significant increases in DA levels (two-way ANOVA, main effect dose, F4,20 = 34.1, p < 0.001; main effect time, F18,360 = 73.8, p < 0.001; time × dose interaction, F72,360 = 10.1, p < 0.001) (Fig. 6, left). The increases in DA levels produced by cocaine were rapid and relatively transient at all doses and reached maximum at about 10 to 20 min after injection (Fig. 6; left). At the 30-mg/kg dose, the maximum increase in DA levels was about 435% of basal values. DA levels returned to basal values after about 180 min from injection of this dose of cocaine (Fig. 6; left).

Time course of effects of systemic administration of cocaine (left panel) or 4-Cl-BZT (right panel) on DA extracellular levels in dialysates from the PFCX. The N was 4, 6, 6, and 4 for cocaine doses of 1, 3, 10, and 30 mg/kg, respectively. The N was 5, 5, and 4 for 4-Cl-BZT doses of 3, 10, and 30 mg/kg, respectively. All other details are as in Fig. 3.
Administration of 4-Cl-BZT (3, 10, and 30 mg/kg) produced a dose-dependent and significant increase in DA levels in the PFCX (two-way ANOVA, main effect dose, F3,14 = 192, p < 0.001; main effect time, F18,252 = 51.5, p < 0.001; time × dose interaction, F54,252 = 26.5, p < 0.001) (Fig. 6, right). Administration of the highest dose of 4-Cl-BZT (30 mg/kg) induced a prolonged (>5 h) and significant increase in DA levels, which reached maximum at about 40 to 60 min after administration (Fig. 6; right). The maximum increase was about 550% of basal values. Lower doses of 4-Cl-BZT (3 and 10 mg/kg) produced a reduced but still significant (post hoc, p < 0.05) effect.
The effects of each of the drugs (in micromoles per kilogram) on extracellular DA levels in the different areas, during the 30-min period in which maximal stimulation of DA transmission was observed, are compared in Fig. 7 (0 to 30 and 30 to 60 min for cocaine and 4-Cl-BZT, respectively). Across the range of doses examined, cocaine was more effective than was 4-Cl-BZT in the shell and core of the NAC (significant effect of preparations; F values >10.8, p < 0.003). The two compounds were about equally effective in caudate (F1,25 = 3.11, p = 0.09), and 4-Cl-BZT was more effective than cocaine in PFCX (F1,26 = 7.55, p = 0.011). Cocaine was more potent in the shell of the NAC than it was in the PFCX, NAC core, and caudate. In contrast, 4-Cl-BZT was slightly more potent in PFCX than it was in shell (Table 1). In general, cocaine was more potent than 4-Cl-BZT in each brain region examined (Fig. 7, Table 1), with relative potency values ranging from 1.88- to 2.88-fold more potent than 4-Cl-BZT.

Dose-dependent maximal effects of cocaine and 4-Cl-BZT on extracellular levels of DA. Ordinates: extracellular DA levels as a percentage of baseline during the 30-min period of time after drug administration in which maximal stimulation of DA transmission was observed. Abscissae: dose of drug in micromoles per kilogram, log scale. Each point represents the average effect determined in four to seven rats.
Effective doses and relative potencies of cocaine and 4-Cl-BZT in each of the brain regions studied
Effective doses are expressed as the doses producing an increase in extracellular DA levels to 300% of basal values, or an increase in horizontal locomotor activity of 100 counts, determined by linear regression. Relative potency is the dose of 4-Cl-BZT in micromoles per kilogram equal to 1 μmol/kg cocaine.
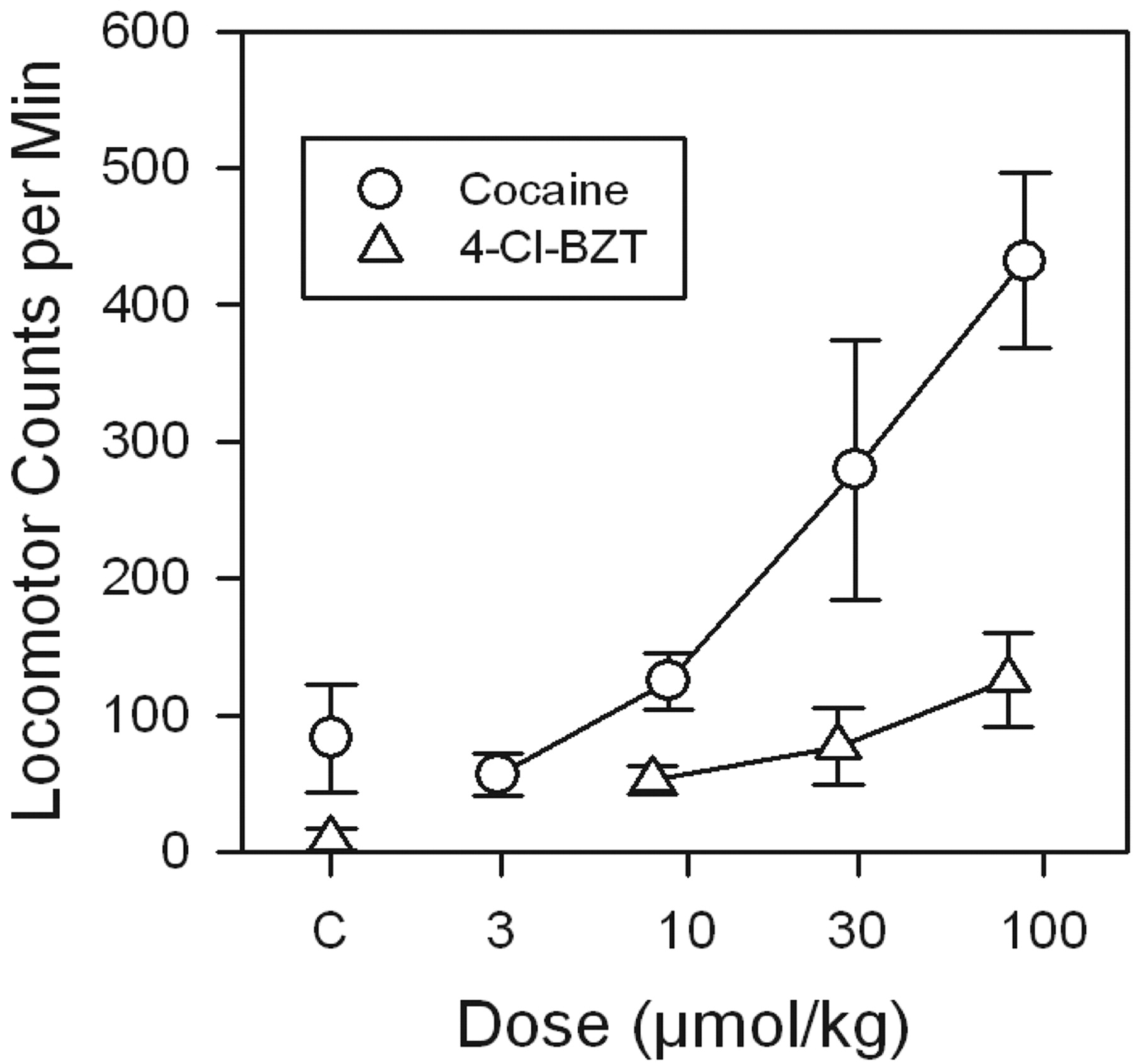
Cocaine was more potent and more effective than was 4-Cl-BZT as a stimulant of locomotor activity at the times during which each of the drugs had their maximal effects on extracellular DA (Fig. 8). The cocaine dose producing a 100-count increase in horizontal activity was about 7.5-fold lower than that for 4-Cl-BZT, consistent with the greater potency observed for cocaine (Table 1).

Dose-dependent maximal effects of 4-Cl-BZT and cocaine on locomotor activity in rats. Ordinates: horizontal locomotor activity counts during the 30 min after drug administration in which maximal effects on extracellular DA were obtained. Abscissae: dose of drug in micromoles per kilogram, log scale. Each point represents the average effect determined in five to six rats.
Discussion
It has been hypothesized that inhibition of DA uptake through actions at the DAT confers behavioral effects like those of cocaine (Kuhar et al., 1991). However, analogs of BZT bind to the DAT and inhibit DA uptake in vitro, but have behavioral effects that differ from those of cocaine (Katz et al., 1999, 2004; Tolliver et al., 1999; Woolverton et al., 2000). The present study was initiated to compare the effects of a BZT analog and cocaine on DA levels in vivo, to obtain a better appreciation of how those effects might account for differences in behavioral effects.
The present results indicate that 4-Cl-BZT and cocaine induced different dose-dependent patterns of activation of DA in selected dopaminergic terminal areas. The increase in DA levels produced by cocaine was graded in response to increments in dose in all areas. In contrast, the response to 4-Cl-BZT dose was graded in NAC shell and PFCX, whereas only the highest dose produced a substantial and significant effect in dorsal caudate and NAC core. Cocaine was most potent in the shell of the NAC compared with the other regions, whereas 4-Cl-BZT was slightly more potent in PFCX than it was in NAC shell. Moreover, cocaine was much more effective than 4-Cl-BZT in increasing levels of DA in the shell of the NAC.
The study by Tolliver et al. (1999) also showed a difference in the effectiveness of cocaine and 4-Cl-BZT on DA in the NAC, which presently was more notable in the shell than core. Activation of DA transmission in the NAC shell and core have both been implicated in stimulation of locomotor activity (Ikemoto, 2002; Sellings and Clarke, 2003). In the present study, differences in the efficacies of 4-Cl-BZT and cocaine in elevating DA levels in the shell corresponded to their differences in efficacies in stimulating locomotor activity, as well as cocaine discrimination results described in the Introduction. The relatively similar efficacies of the drugs on DA in other regions suggests that these other regions are less involved with the behavioral effects of the drugs than is the NAC shell and that the differences between BZT analogs and cocaine in their effects on DA in this region may account for their different effects on behavior.
However, the effects on extracellular DA levels in NAC shell do not alone account for all of the observed behavioral differences between the drugs. For example, cocaine was at the greatest about 3-fold more potent than 4-Cl-BZT in producing increases in extracellular DA, whereas it was about 7.5-fold more potent in increasing locomotor activity. Drug discrimination studies have shown, along with efficacy differences, an approximate 3-fold potency difference between the drugs (Katz et al., 1999), indicating a better concordance among these effects and microdialysis than with locomotor activity.
None of these potency differences are in line with the relative affinities of the two drugs at the DAT, with Ki values of 30 and 189 nM for 4-Cl-BZT and cocaine, respectively (Izenwasser et al., 1994; Katz et al., 1999). In contrast to 4-Cl-BZT, the binding of cocaine often fits a two-site model yielding a Ki value of 32 nM for the high-affinity component (Katz et al., 1997), which is more in line with the present findings, as well as drug discrimination studies (Katz et al., 2000), and about equal to that of 4-Cl-BZT. However, neither the 7.5-fold differences in potency in stimulating locomotor activity nor the 3-fold differences in potency for increasing extracellular DA levels can be accounted for solely on the basis of any of the binding affinities.
Substantial differences between time courses of the two drugs obtained in the present study suggest that differences in the central nervous system permeability of the drugs may also contribute to the differences between their behavioral effects. Pharmacokinetic studies in rats have shown that maximal brain-to-plasma ratios for several BZT analogs were observed later than for cocaine, indicating differences in their central nervous system permeability (Raje et al., 2003). However, the BZT analogs were themselves highly permeable and detectable in brain within 10 min after injection. Thus, if the pharmacokinetic profile of 4-Cl-BZT resembles those of the previously studied BZT analogs, 4-Cl-BZT is in the brain in sufficient concentrations and with an affinity sufficient to produce an effect comparable with that of cocaine.
Like most drugs of abuse, cocaine preferentially increases DA levels in the NAC shell compared with core (Pontieri et al., 1995; present report), an effect that has been suggested to be the result of blockade of the DAT coupled with differences in the neuropharmacology and/or neurophysiology of the NAC subregions (Kuczenski et al., 1991; David et al., 1998; Wu et al., 2001). 4-Cl-BZT showed selective increases in DA levels in the NAC shell compared with the core, although only at 10 mg/kg, suggesting that at least this dose might have reinforcing effects. However, the slower onset of effects compared with cocaine and long duration of action of this drug (Figs. 3, 4, 5, 6) may have minimized reinforcing effects shown in primate self-administration studies (Woolverton et al., 2000, 2001). Because the temporal contingency between the behavior and onset of effects produced by drug injections can be a critical feature in self-administration (Balster and Schuster, 1973), it is possible that other procedures might reveal reinforcing effects of 4-Cl-BZT.
The selective effect on DA levels in the shell compared with the core and dorsal caudate produced by 10 mg/kg 4-Cl-BZT might not be solely the result of actions at the DAT. The affinity of 4-Cl-BZT for muscarinic M1 receptors is higher than its affinity for the DAT (Katz et al., 2001), and antagonists of M1 receptors potentiate behavioral effects of cocaine in rats (Katz et al., 1999) and selectively potentiate cocaine-induced elevations of DA in the NAC shell compared with the core in rats (Tanda et al., 2004). Thus, the selective effect of 4-Cl-BZT on DA transmission in the shell compared with core and dorsal caudate might be the result of blockade of both the DAT and M1 receptors.
4-Cl-BZT had greater efficacy on DA levels in the PFCX compared with striatal areas. The ability of cocaine to increase DA transmission in the PFCX is mainly related to its actions at the norepinephrine transporter (NET) (Di Chiara et al., 1992; Tanda et al., 1997b; Moron et al., 2002; Williams and Steketee, 2004). Indeed, GBR 12909, administered at doses selective for DAT blockade did not increase DA transmission in the PFCX to the same extent as cocaine (Tanda et al., 1997b). However, the affinity of 4-Cl-BZT for the NET is about 50-fold lower than its affinity for the DAT (Katz et al., 2001). Thus, the increase in DA levels in PFCX produced by 4-Cl-BZT depends, at least partly, on mechanisms other than DAT and NET blockade.
Mesolimbic DA plays a critical role in the discriminative-stimulus effects of cocaine (e.g., Wood and Emmett-Oglesby, 1989; Callahan et al., 1997; De La Garza et al., 1998; Filip et al., 2000). In rodents, those effects usually have been investigated in subjects trained with 10 mg/kg cocaine. Only the highest dose of 4-Cl-BZT (30 mg/kg) induced a stimulation of DA similar to that produced by 10 mg/kg cocaine in the caudate, PFCX, and NAC core, whereas in the shell 10 mg/kg 4-Cl-BZT produced an increase in DA levels comparable with that produced by the same dose of cocaine. Thus, if the NAC shell is primarily mediating the discriminative effects of cocaine, it is not clear why 4-Cl-BZT does not fully substitute for cocaine. Although the dose effects of cocaine and 4-Cl-BZT in the NAC shell were similar up to about 30 μmol/kg, the different effects of 4-Cl-BZT in the NAC core and caudate and the putative different mechanisms in the PFCX suggest a basis for differences in the subjective effects of the two drugs.
The results shown in the present study indicate that 4-Cl-BZT, although blocking the DAT with higher affinity than cocaine, does not produce a cocaine-like stimulation of locomotor activity and a cocaine-like pattern of activation of DA neurotransmission. Our results, in agreement with others (Katz et al., 1999, 2004; Tolliver et al., 1999; Woolverton et al., 2000), suggest that a simple blockade of the DAT does not account for all the behavioral and reinforcing effects of cocaine. Nonetheless, the relative effects of the two drugs on DA levels showed similarities to relative effects on behavior, most notably the reduced potency and, in most areas, reduced efficacy of 4-Cl-BZT compared with cocaine. However, the mechanism(s) for the differences between these drugs with respect to their effects on extracellular DA remain unclear. Effects mediated by mechanisms in addition to DA uptake inhibition may contribute to the differences between the drugs on DA levels. Alternatively, the differences could represent primary differences between the drugs with regard to their efficiency in blocking DA uptake. Whatever mechanism is ultimately revealed, the present results suggest that the pattern of activation of DA transmission produced by 4-Cl-BZT is not qualitatively or quantitatively sufficient to completely reproduce cocaine-like effects.
Acknowledgments
We thank Bettye Campbell for technical support and Patty Ballerstadt for administrative and clerical support.
Footnotes
-
Portions of this paper were presented at a meeting on Monitoring Molecules in Neuroscience, Stockholm, Sweden, June 2003 and the 33rd Annual Meeting of the Society for Neuroscience, 2003.
-
doi:10.1124/jpet.104.080465.
-
ABBREVIATIONS: DAT, dopamine transporter; DA, dopamine; BZT, benztropine; WIN 35,428, 2β-carbomethoxy-3β-4-(fluorophenyl)-tropane; GBR 12909, 1-[2-[bis(4-fluorophenyl)methoxy]ethyl]-4-(3-phenylpropyl)piperazine dihydrochloride; 4-Cl-BZT, 4′-chloro-3α-(diphenylmethoxy)-tropane; NAC, nucleus accumbens; PFCX, medial prefrontal cortex; ANOVA, analyses of variance; NET, norepinephrine transporter.
- Received November 9, 2004.
- Accepted January 25, 2005.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}