


Abstract
The theoretical basis for analyzing the effects of an allosteric modulator on the response to an agonist is described. The effects of an allosteric modulator on the concentration-response curve to an agonist can be attributed to changes in the observed dissociation constant and intrinsic efficacy of the agonist-receptor complex. Each of these two changes can be represented by a coefficient or factor. It is possible to estimate the ratio of the coefficient of change in agonist efficacy divided by that for the agonist dissociation constant. This ratio is designated as the relative activity (RA) of the agonist in the presence of the allosteric modulator. The RA value can be estimated for each concentration of allosteric modulator by nonlinear regression analysis, regardless of the shape of the concentration-response curve. Regression analysis of the RA values against the concentration of allosteric modulator yields estimates of the dissociation constant (KA) of the allosteric modulator and the maximal RA value. If the concentration-response curve to the agonist obeys a logistic function and the allosteric modulator influences the maximal response, it is possible to distinguish between the maximal change in affinity from that of efficacy. If the agonist concentration-response curve obeys a logistic equation with a Hill slope of 1, the RA values can be estimated easily from the agonist EC50 and Emax values. This analysis illustrates the utility of the RA value in quantifying allosteric effects.
Drugs that act allosterically to modify receptor function have unique advantages over those that bind directly to the primary recognition site of a receptor. The latter agents produce a continuous effect, whereas allosteric drugs only tune up or tune down signaling when the endogenous neurotransmitter is released (Burgen, 1981; Ehlert, 1986; Christopoulos, 2002). Thus, allosteric drugs preserve the temporal pattern of signaling across the synapse or neuroeffector junction. In addition, with overdose, high concentrations of a directly acting agonist or antagonist can cause excessive receptor activation or inhibition, whereas the maximal effect of an allosteric drug has a ceiling depending upon the nature of its allosteric effect. Thus, protection against overdose can be achieved with allosteric drugs.
There are three pharmacodynamic properties that determine the effects of an allosteric drug on the action of an agonist or endogenous neurotransmitter. These properties are 1) the affinity (KD) of the allosteric drug for its site on the receptor, 2) its modulatory effect on the affinity of the agonist-receptor complex (cooperativity), and 3) its modulatory effect on the intrinsic efficacy of the agonist-receptor complex. Several investigators have described methods for measuring the affinities and cooperative effects of allosteric drugs in radioligand binding assays (Stockton et al., 1983; Ehlert, 1988a; Lazareno and Birdsall, 1995). However, to date, there have been few attempts to discriminate between allosteric effects on agonist affinity and intrinsic efficacy in functional assays. Moreover, it is often assumed that the methods presently available for investigating allosterism in functional assays are valid only if the allosteric agent is without effect on the intrinsic efficacy of the agonist-receptor complex. However, a null method (Ehlert, 1988a) was described quite some time ago for estimating allosteric ligand affinities in functional assays under conditions where the allosteric agent modulates either the affinity or intrinsic efficacy of the agonist or both. The method was based on the assumption that the intrinsic efficacy of the agonist-receptor-allosteric drug complex (XRA) could be different from that of the agonist-receptor complex (XR). By deriving the mathematical equation for the combined stimulus elicited by both receptor complexes (i.e., XR and XRA), it was possible rearrange the latter equation to derive a null equation for comparing equivalent tissue responses in the presence and absence of an allosteric drug (Ehlert, 1988a). In the previous report, simple methods for applying the null equation to evaluate allosteric changes in agonist affinity and efficacy were described for highly efficacious agonists exhibiting a large receptor reserve or for situations with little receptor reserve only involving a change in affinity. In principle, it should be possible to apply this mathematics to all conditions using nonlinear regression analysis.
In the present report, the theory for investigating allosteric interactions in functional assays is described. This theory is applied to predict the behavior of the concentration-response curve of an agonist in the presence of allosteric drugs that modify agonist affinity and efficacy. Three mathematical methods are described and applied to analyze the theoretical data. These are 1) a null method, which requires no assumption regarding the nature of the stimulus-response relationship; 2) a method based on a logistic relationship between the stimulus and response (i.e., operational model); and 3) a special case of the former method, in which the Hill slope of the concentration-response curve equals 1. Finally, methods based on the operational model are used to analyze experimental data from the literature. The results show that it is possible to estimate the affinity of an allosteric drug in functional assays regardless of whether it influences the affinity and intrinsic efficacy of the agonist. Under all conditions, it is possible to measure the product of the allosteric changes in efficacy and affinity (i.e., reciprocal of the dissociation constant). If the allosteric agent influences the maximal response of the agonist, it is possible to distinguish the allosteric change in affinity from the change in efficacy.
Materials and Methods
Theory
Allosterism. An allosteric drug binds at a secondary allosteric site on the receptor, distinct from the site at which the agonist binds. The scheme showing the interaction of an allosteric drug (A) with an agonist (X) at a receptor (R) is as follows:
 in which KX denotes the dissociation constant of the agonist-receptor complex, KA denotes the dissociation constant of the allosteric drug-receptor complex, and αKA and αKX denote the corresponding dissociation constants when the receptor is occupied by both ligands. Receptor theory posits that the response to an agonist is a function (f) of the stimulus, with the stimulus being equivalent to the product of receptor occupancy and intrinsic efficacy (Stephenson, 1956; Furchgott, 1966). If it is assumed that the RA complex is inactive, then the stimulus (S) of an agonist in the presence of an allosteric drug is given by the following (Ehlert, 1988a):
in which KX denotes the dissociation constant of the agonist-receptor complex, KA denotes the dissociation constant of the allosteric drug-receptor complex, and αKA and αKX denote the corresponding dissociation constants when the receptor is occupied by both ligands. Receptor theory posits that the response to an agonist is a function (f) of the stimulus, with the stimulus being equivalent to the product of receptor occupancy and intrinsic efficacy (Stephenson, 1956; Furchgott, 1966). If it is assumed that the RA complex is inactive, then the stimulus (S) of an agonist in the presence of an allosteric drug is given by the following (Ehlert, 1988a):  in which ϵ and ϵ′ denote the intrinsic efficacies of the XR and XRA complexes, respectively. As described previously (Ehlert, 1988a), making the appropriate substitutions yields the following:
in which ϵ and ϵ′ denote the intrinsic efficacies of the XR and XRA complexes, respectively. As described previously (Ehlert, 1988a), making the appropriate substitutions yields the following:  in which RT denotes the total amount of receptors. This equation simplifies to the following:
in which RT denotes the total amount of receptors. This equation simplifies to the following:  in which
in which 

 An equivalent form of eq. 3 has been described by Christopoulos (see eq. 1.21.8; Christopoulos, 2000). In addition, the activation state model of Hall (2000) for allosteric interactions is entirely consistent with eq. 3 under the condition where little receptor is active in the absence of agonist and the RA complex is inactive. Equation 3 shows that, in the absence of the allosteric modulator (i.e., A = 0 and therefore p = 1 and q = 1), the stimulus function resembles a one-site model with a maximum proportional to ϵRT and the concentration of agonist eliciting a half-maximal stimulus equivalent to KX. The effect of the allosteric modulator is to cause a concentration-dependent modification in either the observed dissociation constant or the maximal stimulus or both by the coefficients p and q, respectively. If it were possible to measure the stimulus directly—such as in voltage-clamp experiments on a ligand-gated ion channel with a simple one-site occupancy curve— one could use regression analysis to fit eqs. 4 and 5 to the estimates of the relative change in potency and maximal current, respectively, to obtain estimates of KX, α, and β. Similarly, in enzyme assays, where the activity of the enzyme is measured directly, one could use regression analysis to fit eqs. 4 and 5 to the estimates of the allosteric change in the maximum velocity (Vmax) and Michaelis constant (Km) to obtain the allosteric parameters KX, α, and β. However, in most pharmacological assays of function, it is impossible to measure the stimulus directly. Rather, a consequent, downstream response is usually measured. In this situation, the response can be described by the following equation:
An equivalent form of eq. 3 has been described by Christopoulos (see eq. 1.21.8; Christopoulos, 2000). In addition, the activation state model of Hall (2000) for allosteric interactions is entirely consistent with eq. 3 under the condition where little receptor is active in the absence of agonist and the RA complex is inactive. Equation 3 shows that, in the absence of the allosteric modulator (i.e., A = 0 and therefore p = 1 and q = 1), the stimulus function resembles a one-site model with a maximum proportional to ϵRT and the concentration of agonist eliciting a half-maximal stimulus equivalent to KX. The effect of the allosteric modulator is to cause a concentration-dependent modification in either the observed dissociation constant or the maximal stimulus or both by the coefficients p and q, respectively. If it were possible to measure the stimulus directly—such as in voltage-clamp experiments on a ligand-gated ion channel with a simple one-site occupancy curve— one could use regression analysis to fit eqs. 4 and 5 to the estimates of the relative change in potency and maximal current, respectively, to obtain estimates of KX, α, and β. Similarly, in enzyme assays, where the activity of the enzyme is measured directly, one could use regression analysis to fit eqs. 4 and 5 to the estimates of the allosteric change in the maximum velocity (Vmax) and Michaelis constant (Km) to obtain the allosteric parameters KX, α, and β. However, in most pharmacological assays of function, it is impossible to measure the stimulus directly. Rather, a consequent, downstream response is usually measured. In this situation, the response can be described by the following equation:  in which f denotes the unknown stimulus-response function. Below, three different approaches for estimating the influence of an allosteric modulator on the affinity and efficacy of the agonist-receptor complex are described. The first involves eliminating the stimulus-response function from the analysis through the use of a null method. The second involves the use of the operational model to describe the stimulus-response function, and the third involves a special case of the operational model in which the Hill slope of the concentration-response curve equals 1.
in which f denotes the unknown stimulus-response function. Below, three different approaches for estimating the influence of an allosteric modulator on the affinity and efficacy of the agonist-receptor complex are described. The first involves eliminating the stimulus-response function from the analysis through the use of a null method. The second involves the use of the operational model to describe the stimulus-response function, and the third involves a special case of the operational model in which the Hill slope of the concentration-response curve equals 1.
Null Method. One approach to estimate allosteric effects in functional assays is to compare equivalent tissue responses in the presence and absence of the allosteric drug (Ehlert 1988a) so that the unknown relationship between the stimulus and response is eliminated. Using this approach, the relationship describing equivalent tissue responses in the absence and presence of an allosteric drug is given by the following:  in which X and X′ denote the equiactive concentrations of agonist in the absence and presence of the allosteric drug, respectively. This equation simplifies to the following:
in which X and X′ denote the equiactive concentrations of agonist in the absence and presence of the allosteric drug, respectively. This equation simplifies to the following:  Equation 9 can also be rearranged in the form
Equation 9 can also be rearranged in the form  It is useful to define the term relative activity of the agonist in the presence of the allosteric modulator (RA), which represents the ratio of the allosteric change in intrinsic efficacy (q) divided by the allosteric change in the dissociation constant of the agonist (p):
It is useful to define the term relative activity of the agonist in the presence of the allosteric modulator (RA), which represents the ratio of the allosteric change in intrinsic efficacy (q) divided by the allosteric change in the dissociation constant of the agonist (p):  Making the appropriate substitutions for p (eq. 4) and q (eq. 5) yields the following:
Making the appropriate substitutions for p (eq. 4) and q (eq. 5) yields the following:  in which
in which  The method for using eqs. 9, 10, 11, and 12 for the analysis of allosteric effects in functional assays is described below under Results.
The method for using eqs. 9, 10, 11, and 12 for the analysis of allosteric effects in functional assays is described below under Results.
Operational Model. Although the null method just described has the advantage of being applicable for any type of stimulus-response function (i.e., any shape of concentration-response curve), it is widely observed that agonists usually exhibit logistic concentration-response curves. Indeed, a curve-fitting procedure based on the following logistic equation is the most common computational method that investigators use to estimate the maximal response (Emax) and concentration of agonist eliciting half-maximal response (EC50):  in which n denotes the Hill slope. Several investigators have shown that if the input to the stimulus-response function (f) is the stimulus (i.e., product of receptor occupancy and intrinsic efficacy) and the output obeys the logistic function just described, then the stimulus-response function must be the following (Furchgott, 1966; Mackay, 1981; Kenakin and Beek, 1982; Black and Leff, 1983):
in which n denotes the Hill slope. Several investigators have shown that if the input to the stimulus-response function (f) is the stimulus (i.e., product of receptor occupancy and intrinsic efficacy) and the output obeys the logistic function just described, then the stimulus-response function must be the following (Furchgott, 1966; Mackay, 1981; Kenakin and Beek, 1982; Black and Leff, 1983):  in which Msys denotes the maximum response of the system, KE denotes the sensitivity of the stimulus-response function, and m denotes the transducer slope factor. This exponent is related to, but not identical to, the Hill slope of the agonist concentration-response curve. Substitution of eq. 3 for S in eq. 15 above followed by simplification yields the following:
in which Msys denotes the maximum response of the system, KE denotes the sensitivity of the stimulus-response function, and m denotes the transducer slope factor. This exponent is related to, but not identical to, the Hill slope of the agonist concentration-response curve. Substitution of eq. 3 for S in eq. 15 above followed by simplification yields the following:  in which
in which 

 It is useful to define the term relative activity (RA) of the agonist in the presence of the allosteric drug as follows:
It is useful to define the term relative activity (RA) of the agonist in the presence of the allosteric drug as follows:  in which τobs′ and Kobs′ denote the τobs and Kobs values of the agonist in the absence of the allosteric drug. By making the appropriate substitutions for Kobs and τobs (eqs. 17 and 18, respectively), it can be shown that eq. 20 simplifies to eq. 12. In addition, it can be shown by substitution of eq. 4 for p in eq. 17 that the observed dissociation constant of the agonist (Kobs) in the presence of the allosteric drug divided by that measured in its absence (Kobs′) is equivalent to the following:
in which τobs′ and Kobs′ denote the τobs and Kobs values of the agonist in the absence of the allosteric drug. By making the appropriate substitutions for Kobs and τobs (eqs. 17 and 18, respectively), it can be shown that eq. 20 simplifies to eq. 12. In addition, it can be shown by substitution of eq. 4 for p in eq. 17 that the observed dissociation constant of the agonist (Kobs) in the presence of the allosteric drug divided by that measured in its absence (Kobs′) is equivalent to the following:  In situations where the exponent m in eq. 16 equals 1, the Hill slope is also equal to 1 (n = 1), and eq. 16 reduces to the following:
In situations where the exponent m in eq. 16 equals 1, the Hill slope is also equal to 1 (n = 1), and eq. 16 reduces to the following: 

 When the Hill slope is equal to 1, it is possible to estimate the RA of the agonist in the presence of the allosteric drug as follows:
When the Hill slope is equal to 1, it is possible to estimate the RA of the agonist in the presence of the allosteric drug as follows:  in which EC50′ and Emax′ denote the EC50 and Emax values of the agonist in the absence of the allosteric drug, and EC50 and Emax denote those measured in the presence of the allosteric drug. By making the appropriate substitutions for the various EC50 and Emax values (eqs. 23 and 24, respectively), it can be shown that eq. 25, like eq. 20, also simplifies to eq. 12.
in which EC50′ and Emax′ denote the EC50 and Emax values of the agonist in the absence of the allosteric drug, and EC50 and Emax denote those measured in the presence of the allosteric drug. By making the appropriate substitutions for the various EC50 and Emax values (eqs. 23 and 24, respectively), it can be shown that eq. 25, like eq. 20, also simplifies to eq. 12.

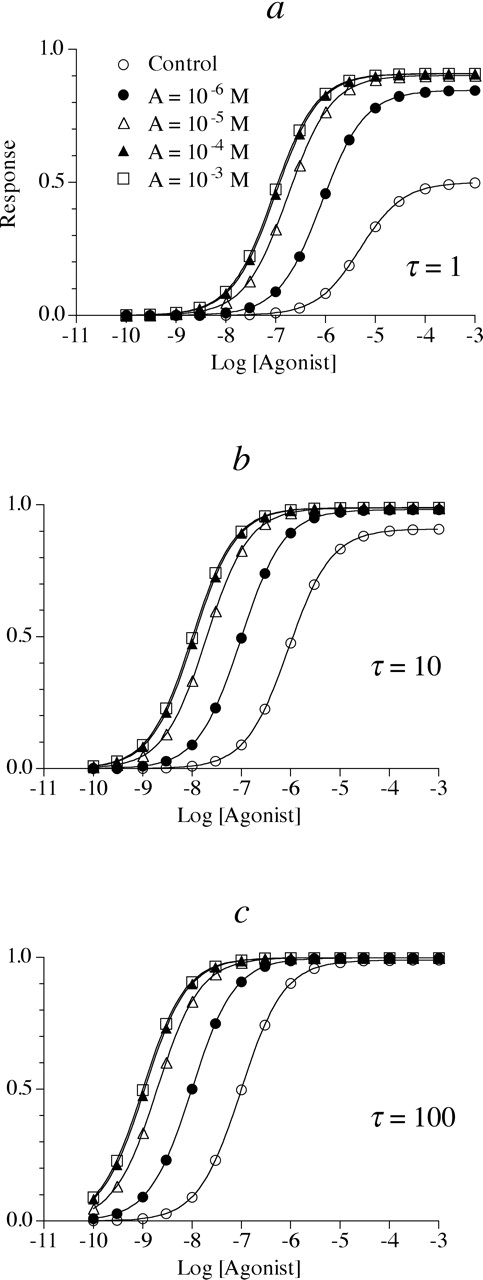
Simulation of the effects of an allosteric modulator inhibiting both agonist affinity and intrinsic efficacy on the concentration-response curve to an agonist having a Hill slope (n) of 1 (i.e., m = n = 1). In these examples, the allosteric modulator causes both a maximal 10-fold reduction in the affinity (α = 10) and intrinsic efficacy (β = 0.1) of the agonist-receptor complex. Small (τ = 1) (a), intermediate (τ = 10) (b), and large (τ = 100) (c) receptor reserves were simulated. The dissociation constant of the allosteric inhibitor (KA) was 10-5 M and Msys = 1. The concentrations of allosteric modulator are indicated in a for the entire figure.
The use of eqs. 20, 21, and 25 in the analysis of allosteric effects in functional assays is described under Results.
Results
Simulation of Allosterism in Functional Assays
The influence of allosteric modulators on the agonist concentration-response curve has been simulated by Kenakin (1997) and Christopoulos (2000) for a variety of conditions. These simulations and the theory for allosterism (Ehlert, 1988a) show that an allosteric modulator causes a concentration-dependent change in the agonist concentration-response curve. The magnitude of this change approaches a limit with high concentrations of the modulator. Allosteric changes in affinity are manifest as a parallel shift in the agonist concentration-response curve along the abscissa, whereas a modulation in efficacy is usually manifest as a shift or both a shift and change in the maximal response, depending upon the receptor reserve. Below, four simulations are shown to provide examples for the analysis of allosteric interactions.
Allosteric Modulator Causing a Decrease in Both Agonist Affinity and Efficacy. The behavior of allosterism in functional assays can be appreciated by using eq. 16 to generate agonist concentration-response curves in the presence of an allosteric drug having differential effects on agonist affinity and intrinsic efficacy. Figure 1, a to c, shows the effect of an allosteric drug causing a maximal 90% reduction in intrinsic efficacy (β = 0.1) and observed affinity (10-fold increase in Kobs; α = 10) for a response exhibiting a Hill slope (n) of 1 (m = n = 1). When there is little receptor reserve, increasing concentrations of the allosteric inhibitor cause a shift to the right and a decrease in the maximal response of the agonist concentration-response curve (Fig. 1a). However, with an intermediate receptor reserve (Fig. 1b) there is less of a decline in Emax and a greater shift to the right in the agonist concentration-response curve. With a large receptor reserve (Fig. 1c), there is practically no decrease in Emax and a maximal 100-fold shift to the right in the agonist concentration-response curve. The EC50 and Emax values of the agonist in the presence of different concentrations of the allosteric inhibitor were estimated by nonlinear regression analysis of the data according to eq. 14, and these values are listed in Table 1 for the data shown in Fig. 1, a to c.
Estimates of EC50, Emax, Hill slope, Kobs, and τobs for the simulated data in Fig. 1, a to c In this example, the allosteric modulator causes a 10-fold reduction in both the affinity (α = 10) and intrinsic efficacy (β = 0.1) of the agonist-receptor complex.
To investigate the influence of the Hill slope on the behavior of allosterism, the effects of an allosteric drug essentially identical to that described for Fig. 1, a to c, were simulated, but with the response of the agonist exhibiting a Hill slope (n) greater than 1. This change was accomplished by setting the transducer slope faction in the operational model (m) equal to 2. As shown in Fig. 2, a to c, behavior generally consistent with that observed in Fig. 1, a to c, was observed. The effects of the allosteric inhibitor on the EC50 and Emax values of the agonist concentration-response curves shown in Fig. 2 are listed in Table 2.
Estimates of EC50, Emax, Hill slope, Kobs, and τobs for the simulated data in Fig. 2, a to c In this example, the allosteric modulator causes a 10-fold reduction in both the affinity (α = 10) and intrinsic efficacy (β = 0.1) of the agonist-receptor complex.

Simulation of the effects of an allosteric inhibitor on the concentration-response curve to an agonist having a transducer slope factor in the operational model of two (m = 2). In this example, the allosteric modulator causes maximal 10-fold reductions in both affinity and intrinsic efficacy (α = 10; β = 0.1) of the agonist-receptor complex. Small (τ = 1) (a), intermediate (τ = 10) (b), and large (τ = 100) (c) receptor reserves were simulated. The dissociation constant of the allosteric inhibitor (KA) was 10-5 M and Msys = 1. The concentrations of allosteric modulator are indicated in a for the entire figure.
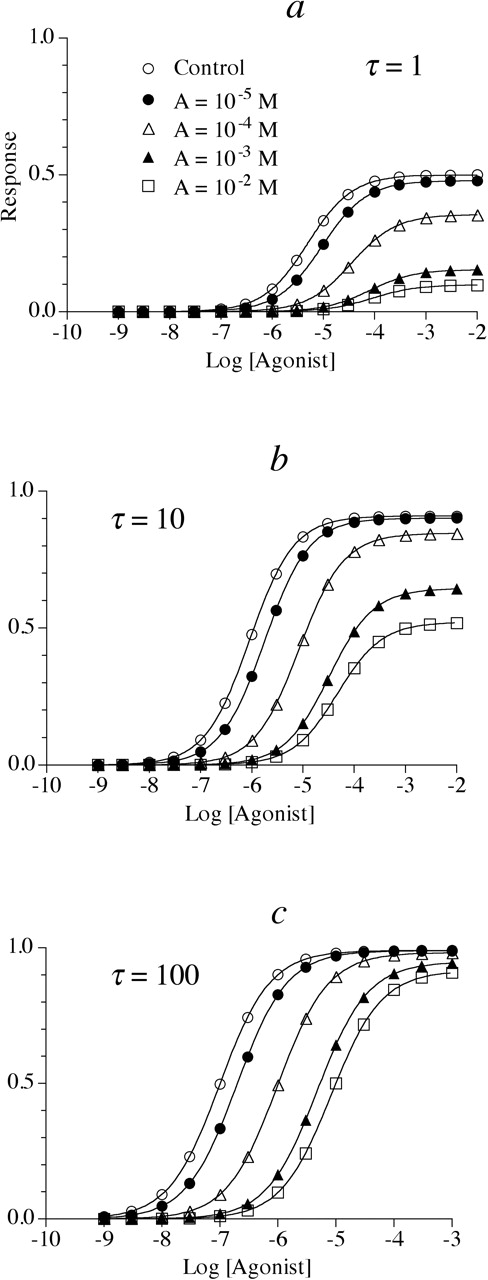
Allosteric Modulator Causing an Increase in Both Agonist Affinity and Efficacy. The effects of an allosteric modulator causing both an increase in affinity and intrinsic efficacy of the agonist are shown in Fig. 3, a to c. In these simulations the maximal increases in affinity and efficacy were both 10-fold (i.e., α = 0.1 and β = 10), and the response exhibited a Hill slope (n)of1(m = n = 1). When there is little receptor reserve, increasing concentrations of the allosteric enhancer cause a shift to the left and an increase in the maximal response of the agonist concentration-response curve (Fig. 3a). However, with an intermediate receptor reserve (Fig. 3b) there is a very small increase in Emax and a greater increase in potency. With a large receptor reserve (Fig. 3c), there is essentially no change in Emax and a maximal 100-fold shift to the left in the agonist concentration-response curve. The effects of the allosteric enhancers on the EC50 and Emax values for all of the curves show in Fig. 3, a to c, are listed in Table 3.
Estimates of EC50, Emax, and Hill slope for the simulated data in Fig. 3, a to c In this example, the allosteric modulator causes a 10-fold enhancement in both the affinity (α = 0.1) and intrinsic efficacy (β = 10) of the agonist-receptor complex.
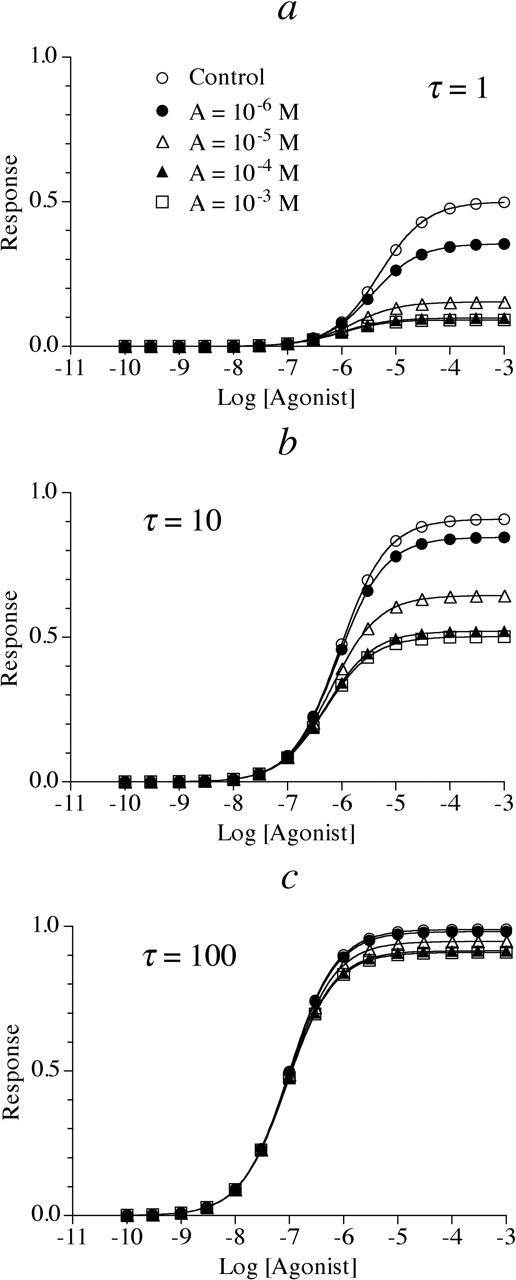
Allosteric Modulator Having Opposite Effects on Agonist Affinity and Efficacy. The effects of an allosteric modulator causing equal and opposite modulation of agonist affinity and intrinsic efficacy were simulated. Figure 4, a to c, shows the effects of an allosteric modulator causing a maximal 10-fold increase in agonist affinity (α = 0.1) and a maximal 90% reduction in agonist efficacy (β = 0.1) with the response exhibiting a Hill slope (n) of 1 (m = n = 1). When the receptor reserve is low (Fig. 4a), the allosteric modulator causes a concentration-dependent decrease in both the Emax and EC50 values, so that the concentration-response curve shifts to the left with a lowered maximum. Similar effects are observed when the receptor reserve is intermediate. However, when the receptor reserve is great, the allosteric inhibitor has little effect on the concentration-response curve because the reduction in efficacy is offset by an equivalent increase in affinity. The effect of the allosteric modulator on the EC50 and Emax values of the data simulated in Fig. 4 is listed in Table 4.
Estimates of EC50, Emax, Hill slope, Kobs, and τobs for the simulated data in Fig. 4, a to c In this example, the allosteric modulator causes 10-fold increase in affinity (α = 0.1) and a 10-fold reduction in intrinsic efficacy (β = 0.1) of the agonist-receptor complex.
Methods for the Analysis of Allosterism in Functional Assays
The methods for the analysis of allosterism presented here can be divided into two main categories, based on the application of the operational model and the null method described under Materials and Methods. These approaches are described below in sections 1 and 2, respectively, where they are applied in the analysis of the simulated data in Figs. 1, 2, 3, 4. In section 3, a set of experimental data from the literature is analyzed.
Section 1: Operational Model. The application of the operational model to the analysis of allosterism in functional assays involves three steps: 1) estimation of the RA values of the agonist in the presence of different concentrations of the allosteric modulator, 2) estimation of the Kobs/Kobs′ ratios in the presence of different concentrations of the allosteric modulator, and 3) nonlinear regression analysis of the foregoing RA values and Kobs/Kobs′ ratios according to eqs. 12 and 21, respectively. This analysis ultimately yields estimates of KA and γ (eq. 12) as well as α (eq. 21), and hence β, since β = γ/α. In the case where the Hill slopes of the agonist concentration-response curves do not differ significantly from 1, it is possible to estimate the RA values from the Emax and EC50 values of the concentration-response curves. Thus, in this instance, the concentration-response curves can be analyzed independently, using a nonlinear regression program (Prism; Graph-Pad Software Inc., San Diego, CA) to estimate the EC50 and Emax values of each curve by fitting the standard equation for a sigmoidal dose-response curve having a Hill slope of 1 (eq. 22) to the data. In situations where the Hill slopes differ from 1, the RA values are estimated from the τobs and Kobs values. These latter values can only be estimated by global nonlinear regression analysis according to eq. 16, which involves analyzing the all of the curves measured at different amounts of allosteric modulator simultaneously, sharing estimates of Msys and m among the curves. Similarly, estimation of the cooperativity factor α always involves global nonlinear regression analysis of all of the concentration response curves, regardless of the estimate of the Hill slope.
Estimation of KA and γ when the Hill slope is equivalent to 1. Since the agonist concentration-response curves shown in Fig. 1, a to c, all exhibit Hill slopes of 1, accurate estimates of the RA of the agonist can be estimated from the Emax and EC50 values using eq. 25. These RA values are listed in Table 1 for the different concentrations of allosteric modulator. Figure 5a shows the log RA values plotted against the log of the concentration of allosteric modulator for the data from Fig. 1a. In the presence of maximally effective concentrations of the allosteric modulator, the plot approaches an asymptote corresponding to the maximal combined change in observed affinity (α) and intrinsic efficacy (β) of the agonist. This combined effect on affinity and efficacy is denoted as γ and is equivalent to β/α (eq. 13). The log form of eq. 12 was fitted to the data by nonlinear regression analysis, which yielded estimates of 10-5 M and 0.01 for the KA and γ, respectively. These estimates are essentially identical to those used in simulating the data in the first place, demonstrating that the nonlinear regression method provides accurate estimates of the allosteric parameters of the modulator. The same procedure was used to analyze the simulated data in Fig. 1, b and c, and identical results were obtained. This result was expected since the only difference among the plots in Fig. 1, a to c, is the receptor reserve (τ), and not the parameters of the allosteric modulator.
The functional data in Fig. 3 for the allosteric modulator causing an increase in agonist affinity and efficacy can be analyzed in a manner analogous to that described above for the allosteric inhibitor in Fig. 1 because both sets of data exhibit Hill slopes of 1. Figure 5c shows a plot of the RA values (Table 3) of the agonist plotted against the log of the allosteric modulator concentration for the data in Fig. 3, a to c. Regression analysis of the data according to eq. 12 yielded estimates of 10-5 M and 100 for KA and γ, respectively. These values are identical to those used in the simulation of the data.
The functional data in Fig. 4, a to c, can also be analyzed in a manner similar to that described for Fig. 1, a to c. Since the Hill slopes of the curves are equal to 1, the RA values can be calculated according to eq. 25, and these values are listed in Table 4. Figure 5e shows that the plot of log RA against log A yield a horizontal line at RA = 1. Thus, it is impossible to estimate KA from this plot, since any value of KA will yield a best fit of eq. 12 to the data as long as γ = 1. In this case where α/β = 1, the KA value can be estimated through the analysis of the Kobs values as described in the next section.
Resolution of γ into its α and β components when the Hill slope is equivalent to 1. To discriminate between the modulatory effects of the allosteric agent on the observed dissociation constant and intrinsic efficacy of the agonist receptor complex, it is necessary to fit the operational model (eq. 16) to all of the agonist concentration-response curves with varying allosteric modulator simultaneously. With regard to the data in Fig. 1, regression analysis is done sharing the estimate of Msys among all of the curves and estimating unique values of Kobs and τobs for each curve. Since all of the curves have Hill slopes of 1, the exponent m can be removed from eq. 16 or constrained at a constant value of 1. Global nonlinear regression analysis of the data in Fig. 1a yielded an estimate of 1.0 for Msys, essentially the same as that used in the simulation of the data. The values of τobs and Kobs for the various concentrations of allosteric modulator are listed in Table 1. The RA values of the agonist can also be estimated from the τobs and Kobs values for each concentration of allosteric modulator using eq. 20, and these are also listed in Table 1. These RA values are essentially identical to those estimated from the Emax and EC50 values of the agonist (compare the last two columns in Table 1). Figure 5b shows a plot of the log ratio (Kobs/Kobs′) against the log concentration of the allosteric modulator. It can be seen that this ratio reaches a plateau corresponding to α at high concentrations of allosteric modulator. Nonlinear regression analysis was used to fit the log form of eq. 21 to the data, which yielded estimates of 10-5 M and 10 for KA and α, respectively. This estimate of KA is identical to that estimated from the RA values shown in Fig. 5a and that used in the simulations, and the estimate of α is equivalent to that used in the simulations. Knowing γ and α, it is possible to estimate β by rearrangement of eq. 13 (γ = β/α), which yields an estimate of 0.1 for β. This value is identical to that used in the simulations.
Simulation of the effects of an allosteric modulator enhancing both agonist affinity and intrinsic efficacy on the concentration-response curve to an agonist having a Hill slope (n) of one (m = n = 1). In these examples, the allosteric modulator causes both a maximal 10-fold increase in the affinity (α = 0.1) and intrinsic efficacy (β = 10) of the agonist-receptor complex. Small (τ = 1) (a), intermediate (τ = 10) (b), and large (τ = 100) (c) receptor reserves were simulated. The dissociation constant of the allosteric inhibitor (KA) was 10-5 M and Msys = 1. The concentrations of the allosteric modulator are indicated in a for the entire figure.
A similar analysis was done to estimate the τobs and Kobs values for the concentration-response curves shown in Fig. 1b, and these parameters are also listed in Table 1. The corresponding RA values were estimated (Table 1) and found to be identical to those estimated from the curves in Fig. 1a. Consequently, plots identical to those shown in Fig. 5, a and b, were derived for the curves in Fig. 1b, and the same values for KA, α, β, and γ were estimated.
Under normal circumstances, where actual experimental data are analyzed, it would be difficult to obtain unique values of τobs and Kobs for each curve shown in Fig. 1c because all of the Emax values of the curves are approximately the same. However, with the simulated data in Fig. 1c, it is possible to obtain these estimates because the data have no error, and there are actually small differences in the Emax values. Regardless, in the situation of little change in Emax,it is always possible to obtain accurate estimates of the ratio of the two parameters (i.e., τobs/Kobs), even though estimation of the individual values may be impossible. The nonlinear regression method for obtaining this ratio is described below in connection with Fig. 6. As described in the next section, it is possible to estimate the RA values from these ratios, and these estimates are listed in Table 1. The estimates of τobs and Kobs for the data in Fig. 1c have been omitted from Table 1 because they would be difficult to estimate in experiments with data having moderate experimental error.
The approach described for the estimation of α from the data in Fig. 1 was applied to the data in Figs. 3 and 4, and the results are summarized in Tables 3 and 4. Figure 5d shows the ratio of Kobs values plotted against the log of the allosteric enhancer concentration for the data in Fig. 3, a and b. Regression analysis (eq. 21) yielded estimates of 10-5 M and 0.1 for KA and α, respectively. Knowing γ and α, it is possible to estimate a value of 10 for β using eq. 13. Figure 5f shows a plot of the log ratio (Kobs/Kobs′) against log A for the data in Fig. 4, a and b. Regression analysis according to eq. 21 yielded estimates of 10-5 M and 0.1 for KA and α, respectively. Knowing that the horizontal plot in Fig. 5e with RA = 1 implies that γ = 1, it is possible to estimate that β = 0.1 by rearrangement of eq. 13. It is impossible to estimate any allosteric parameters from the data in Fig. 4c because the allosteric modulator has little or no effect on the concentration-response curve. This situation occurs because the allosteric effects on affinity and intrinsic efficacy are equal and opposite. When there is a large receptor reserve, the 10-fold reduction in intrinsic efficacy is manifest as a 10-fold shift to the right in the concentration-response curve, whereas the 10-fold increase in affinity causes the opposite effect, resulting in no net effect.
Estimation of allosteric parameters when the Hill slope differs from 1 and there is a modulation of Emax. To analyze the data shown in Fig. 2 according to the operational model, it is inaccurate to estimate agonist RA from their EC50 and Emax values according to eq. 25 because the Hill slopes of the concentration-response curve differ from 1. Consequently, eq. 16 was fitted to all of the concentration response-curves simultaneously, sharing the estimate of Msys and the exponent m among the curves and estimating unique values of τobs and Kobs for each curve. The global estimates of Msys and m from regression analysis were 1.0 and 2.0 for Msys and m, respectively. These values are identical to those used in the simulations. The estimates of τobs and Kobs were used to calculate the agonist RA values (eq. 20). These RA estimates are listed in Table 2 and are essentially the same as those listed in Table 1 for the curves shown in Fig. 1, a and b. This result was expected because the only difference between the parameters for the data in Figs. 1, a to c, and 2, a to c, is the value of m, but not any of the other parameters, including the RA values. Consequently, analysis of the RA values for Fig. 2, a and b, yields plots essentially identical to those shown in Fig. 5, a and b, and the KA, α, and β estimates for the allosteric drug were 10-5 M, 10, and 0.1, respectively.
Estimation of allosteric parameters when the Hill slope differs from 1 and there is no modulation of Emax. Since the receptor reserve is great for Fig. 2c and all of the agonist concentration-response curves exhibit nearly the same Emax, numerous combinations of τobs and Kobs all yield the same least-squares fit. Nevertheless, it is possible to estimate the ratio of τobs/Kobs for each curve shown in Fig. 2c. A least-squares fit can be obtained when the value of Kobs is equal to or greater than the larger of the two constants, KX or αKX. Thus, the best fit can be obtained by constraining Kobs as a constant at any arbitrary value equal to or greater than the true value of αKX (10-4 M) and estimating unique values of τobs for each of the curves. A summary of this analysis is illustrated in Fig. 6, which shows the best fitting parameter estimates for various values of Kobs between 10-8 and 10-1 M as well as the residual sum of squares (RSS). The ordinate scale on the left corresponds to the various parameter estimates, whereas that on the right corresponds to RSS. The data in Fig. 6 are from the concentration-response curve simulated in the presence of 10-4 M A; analogous plots can be made for the data simulated at the other concentrations of A. The estimate of RSS in the figure is based on all of the curves, not just the one simulated at A = 10-4 M. As the constrained value of Kobs increases to 10-4, RSS approaches a minimum plateau and remains at this best fitting level with a further increase in Kobs no matter how large. Thus, it can be seen that a least-squares fit is obtained once the value of Kobs is equal to or greater than αKX (10-4). As the value of Kobs increases, the estimate of τobs increases in a proportional manner so that the ratio of the two estimates (τobs/Kobs) is constant (Fig. 6). Thus, the best fitting estimate of the ratio τobs/Kobs can be obtained by constraining Kobs to an arbitrarily high constant value and estimating the other parameters that minimize RSS. Using this approach, the ratio of τobs/Kobs was estimated for each of the curves shown in Fig. 2c. Alternatively, it is possible to set τobs as a constant and estimate the value of Kobs that gives the least-squares fit (analysis not shown).
Knowing the ratio of τobs/Kobs for each of the curves shown in Fig. 2c, it is possible to estimate their respective RA values by dividing the ratio estimated in the presence of a given concentration of A by the corresponding ratio obtained for the control curve. In other words, if the τobs/Kobs ratio of one curve is divided by the corresponding ratio for the control curve (τobs′/Kobs′), then the following ratio (τobs′Kobs′/τobs′Kobs) is obtained, which is equivalent to the RA value (eq. 20). The RA values for the data in Fig. 2c were estimated in this manner and are listed in Table 2. These values are identical to those estimated for the data in Figs. 1, a to c, and 2, a and b; consequently, regression analysis of these data according to eq. 20 yields a plot identical to that shown in Fig. 5a with KA and γ estimates of 10-5 M and 0.01, respectively. Since it was impossible to estimate unique Kobs values for each of the curves shown in Fig. 2c, it was impossible to separate γ into its α and β components. As described above, it is impossible to make this distinction when the allosteric inhibitor has no influence on the Emax of the concentration-response curve.
It is informative to estimate the RA values of the data shown in Fig. 2 from the Emax and EC50 values according to eq. 25. As described above, this equation is only appropriate for conditions when the Hill slopes of the agonist concentration-response curve are equal to 1. When these estimate were made, and the data analyzed according to the plots shown in Fig. 5, a and b, estimates of 5.5 × 10-6 M and 0.00073 (Fig. 2a) and 9.21 × 10-6 M and 0.0037 (Fig. 2b) were made for KA and γ, respectively. Thus, there is substantial error in these parameters estimates, which should equal 10-5 M and 0.01. However, when this method was used to analyze the data in Fig. 2c, values of 10-5 M and 0.0090 were estimated for KA and γ, respectively. Note that the latter value of KA is correct, whereas the estimate of γ is only off by 10%. Thus, when the allosteric inhibitor has no influence on Emax it is possible to estimate the RA values according to eq. 25, even if the Hill slopes differ from 1. This conclusion seems reasonable because the two different methods for calculating the RA value (eqs. 20 and 25) yield similar estimates for the data in Fig. 2c but not for those of Fig. 2, a and b (compare the last two columns of Table 2). Note that when the allosteric inhibitor has no effect on the Emax, the estimation of RA from eq. 25 can be simplified to the following:  Figure 7 shows the ratio of EC50 values of the agonist in the presence of the allosteric drug divided by that measured in its absence plotted against the log of the allosteric drug concentration for the data from Fig. 2c. Nonlinear regression analysis of the data according to the log form of eq. 12 yielded estimates of 10-5 M and 0.009 for KA and γ, respectively, as described above.
Figure 7 shows the ratio of EC50 values of the agonist in the presence of the allosteric drug divided by that measured in its absence plotted against the log of the allosteric drug concentration for the data from Fig. 2c. Nonlinear regression analysis of the data according to the log form of eq. 12 yielded estimates of 10-5 M and 0.009 for KA and γ, respectively, as described above.

Simulation of the effects of an allosteric modulator on the concentration-response curve to an agonist having a Hill slope (n) of 1 (m = n = 1). In this example, the allosteric modulator causes both a maximal 10-fold increase in affinity and a 10-fold decrease in intrinsic efficacy (α = 0.1; β = 0.1) of the agonist-receptor complex. Small (τ = 1) (a), intermediate (τ = 10) (b), and large (τ = 100) (c) receptor reserves were simulated. The dissociation constant of the allosteric inhibitor (KA) was 10-5 M and Msys = 1. The concentrations of the allosteric modulator are indicated in a for the entire figure.
In summary, it can be concluded that whenever an allosteric modulator is without effect on Emax, only KA and γ can be estimated. Under these conditions, one can estimate the RA values from eq. 25 or 26 even if the Hill slope differs from 1. Equation 12 can be fitted to the data by nonlinear regression analysis to obtain estimates of KA and γ. It is unnecessary to perform the complicated regression analysis summarized in Fig. 6 to estimate the RA values in this situation. Nevertheless, this analysis has been presented here to illustrate the relationship among the various parameter estimates in this situation.
Section 2: Null Method. An advantage of the null method for the analysis of allosteric interactions is that it can be applied to agonist concentration-response curves that deviate from logistic behavior. Nevertheless, the method will be used here to analyze the logistic curves shown in Figs. 1 and 3. The null method involves three steps: 1) estimation of equiactive concentrations of agonist in the absence (X) and presence (X′) of the various concentrations of the allosteric modulator; 2) global nonlinear regression analysis of the log equiactive agonist concentrations to obtain estimates of the ratio q/p (RA values; see eq. 11) of the agonist in the presence of different concentrations of the allosteric modulator; and 3) nonlinear regression analysis of the foregoing RA values according to eq. 12. This analysis yields estimates of KA and γ.
Allosteric modulator causes a reduction in Emax. Figure 8 shows the application of this method for the data in Fig. 1a. Since the allosteric modulator causes a reduction in the Emax of the agonist, the agonist concentrations (X) are interpolated from the control concentration-response curve that yield responses equivalent to those elicited by the agonist concentrations (X′) used in the presence of the allosteric modulator (see Fig. 8a for the data simulated at A = 10-3 M). These pairs of equiactive agonist concentrations are then plotted on a log scale as shown in Fig. 8b. Each curve in Fig. 8b corresponds to a different allosteric modulator concentration. These curves were analyzed simultaneously by global nonlinear regression analysis of the data according to the log form of eq. 9, with the estimate of KX shared among all the curves and unique values of p and q estimated for each curve. However, it is possible to estimate numerous combinations of KX, p, and q that yield the same least-squares fit for each curve. Using an approach analogous to that described above in connection with Fig. 6, it is possible to estimate the ratio q/p (RA; see eq. 11), by constraining KX to a constant and determining the values of the other parameters that minimize RSS. Figure 8c shows the results of regression analysis for the data simulated at A = 10-3 M. Over the range of KX values shown (10-9–10-1), RSS was insignificant and depended only on the criteria used for convergence of the nonlinear regression algorithm and not on KX. Analogous behavior was observed for the data simulated at the other concentrations of A. Using this approach, various values of RA can be estimated at various the concentrations of the allosteric modulator (A). The resulting log RA values are plotted against log A and analyzed by nonlinear regression analysis according to eq. 12. The results yielded a plot identical to that shown in Fig. 5a, and values of 10-5 M and 0.01 were estimated for KA and γ, respectively. These values are identical to those used in the simulation of the data in Fig. 1a. The data in Fig. 1b were analyzed in a similar manner and the same estimates of γ and KA were obtained (i.e., 0.01 and 10-5 M, respectively).
It is also possible to use the same strategy for the simulated data in Fig. 1c; however, since the data exhibit little or no change in Emax with allosteric modulation, it is necessary to constrain both KX and p as constants and analyze all of the data simultaneously by global nonlinear regression analysis (eq. 9) to obtain q values for each of the curves. By dividing the estimates of q value by the constant value of p, it is possible to obtain the RA estimates for each curve, and hence the values of γ and KA by regression analysis according to eq. 12. When this was done, a plot identical to that shown in Fig. 5a was obtained as well as the same estimates of γ and KA (0.01 and 10-5 M, respectively). Regardless, this complicated analysis is probably unnecessary because the RA values can be estimated accurately from EC50 values according to eq. 26 when there is little change in Emax with the allosteric modulator. Presumably, this situation would apply to nonlogistic agonist concentration-response curves.
Allosteric modulator causes an increase in Emax. In situations where the allosteric modulator causes an increase in the Emax of the agonist, it is possible to analyze the data according to null eq. 10. However, in this instance, where the concentration-response curve in the presence of the allosteric modulator has a greater Emax than that measured in its absence, the X′ values are interpolated from the curve measured in the presence of the allosteric inhibitor that yield responses equivalent to those generated by the concentrations (X) of the control concentration-response curve. Using this approach and an overall strategy analogous to that just described in the preceding paragraph, it is possible to estimate the γ and KA values for the allosteric enhancer shown in Fig. 3, a to c. However, an important difference is the range of values over which the parameter KX can be constrained as a constant for global nonlinear regression analysis. This range was found to be KX ≤ 10-5 for the data in Fig. 3. When this analysis was done, plots identical to that shown in Fig. 5c were obtained, and values of 100 and 10-5 M were estimated for γ and KA, respectively, which are equivalent to those used in simulating the data.
Using the null approach, it is also possible to estimate the γ and KA values of the other allosteric modulators simulated above, with the exception of the data in Fig. 4. In this case, the plot of RA against log A yields a horizontal line at RA = 1 as shown in Fig. 5e. As described above, it is impossible to estimate KA from this figure, only γ. In summary, if the allosteric modulator causes a decrease in Emax, the approach describe in connection with Fig. 8 is used, whereas if an increase in Emax is observed, the approach described in the preceding paragraph is used.
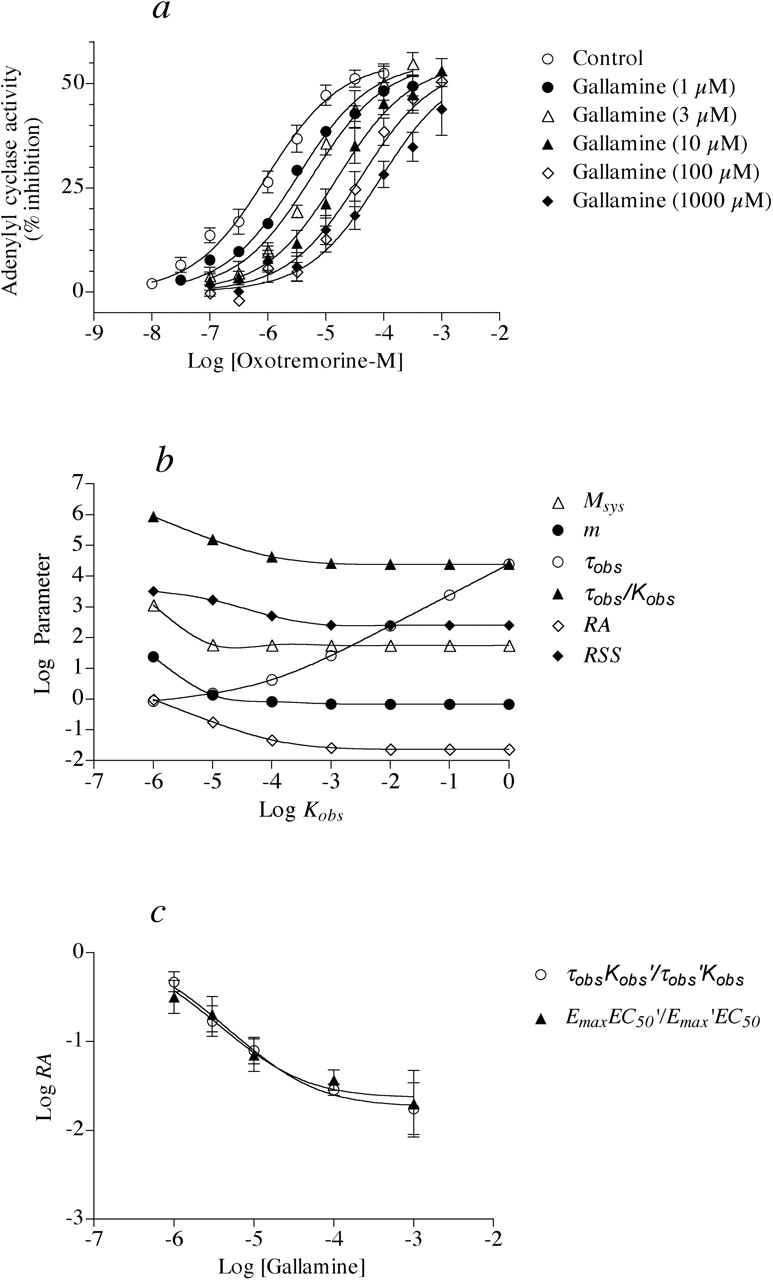
Section 3: Experimental Data. The methods based on the operational model were applied to experimental data to determine the impact of experimental error on the estimates of allosteric parameters. Figure 9a shows the results of a prior study in which the influence of gallamine on muscarinic receptor mediated inhibition of adenylyl cyclase activity was measured in homogenates of the rat myocardium (Ehlert, 1988b). This preparation is known to contain a relatively homogeneous population of M2 muscarinic receptors (Hammer et al., 1986). Gallamine is a neuromuscular blocking agent long known to cause sinus tachycardia (Walton, 1950) through a mechanism involving an inhibition of postjunctional muscarinic receptors (Riker and Wescoe, 1951). The mechanism was later shown to be allosteric in functional studies on isolated beating hearts (Clark and Mitchelson, 1976) and in muscarinic receptor binding assays (Stockton et al., 1983).

Effect of allosteric modulators on the RA (a, c, and e) and Kobs (b, d, and f) values of the agonist. The plots in a and b represent an analysis of the data shown in Figs. 1, a to c, and 2, a to c, and the estimates of RA and Kobs are from Tables 1 and 2. The plots in c and d represent an analysis of the data shown in Fig. 3, a to c, and the estimates of RA and Kobs are from Table 3. The plots in e and f represent an analysis of the data shown in Fig. 4, a to c, and the estimates of RA and Kobs are from Table 4. The dashed lines in a and c denote the maximal effect of the allosteric modulator on the RA value (γ). The dashed lines in b, d, and f denote the maximal shift (α) of the allosteric modulator on the observed affinity of the agonist.
As shown in Fig. 9a, gallamine caused a concentration-dependent shift to the right in the agonist concentration-response curve with little change in Emax. Regression analysis of each curve according to logistic eq. 14 yielded the estimates of EC50, Emax, and n (Hill slope), and these values are listed in Table 5. The results show that the agonist concentration-response curves exhibit low Hill slopes (approximately 0.7–1.0), and that there is little change in Emax. In this situation, it is only possible to estimate KA and γ; it is impossible to resolve γ into its α and β components from these data alone (see analysis of the data in Figs. 1c and 3c). Accordingly, the RA value of the agonist can be estimated easily by calculating the ratio of EC50 values measured in the presence of gallamine divided by that measured in its absence as described above in eq. 26. Nevertheless, the RA values were estimated using the operational model so that the influence of experimental error on these estimates might be appreciated. Normally, this method would only be required in situations where the allosteric modulator influences the Emax.
Estimates of EC50, Emax, Hill slope, and RA for the experimental data in Fig. 9a The mean negative logarithm ± S.E.M. of each estimate of EC50 is indicated in parentheses beneath each EC50 estimate; the mean logarithm ± S.E.M. of each estimate of RA is indicated in parentheses beneath each RA estimate.

Summary of global nonlinear regression analysis of the data in Fig. 2c at A = 10-4 M according to eq. 16. All curves in Fig. 2c were fitted simultaneously sharing the estimate of m and Msys among the curves and constraining the value of Kobs as a constant at various values between 10-5 and 10-1. Regression analysis was used to estimate the values of the various parameters in eq. 16 that yielded the least-squares fit to the data. The plot shows how the estimate of τobs for the data simulated at A = 10-4 M is proportional to Kobs such that their ratio (τobs/Kobs) remains constant at approximately 106.

Influence of an allosteric inhibitor on the EC50 value of an agonist when the receptor reserve is great (τ = 100) and the transducer slope factor in the operational model is equal to 2 (m = 2). The ratio of the control EC50 value divided by that measured in the presence of the allosteric modulator is plotted on a log scale against the concentration of the allosteric modulator. The allosteric modulator causes a 10-fold reduction in the affinity (α = 10) and intrinsic efficacy (β = 0.1) of the agonist receptor complex. The plot represents an analysis of the data shown in Fig. 2c. The EC50 values are from Table 2.
The data were analyzed by global nonlinear regression analysis according to eq. 16 using the procedure described above in connection with Fig. 6 for the estimation of RA values. Regression analysis was done sharing the estimates of Msys and m among the curves and constraining Kobs as a constant so that the ratio τobs/Kobs could be estimated. A summary of the analysis of the data at 10-4 M gallamine is shown in Fig. 9b where the logarithm of the parameter estimates are plotted against Kobs, which was constrained to various fixed values during regression analysis. A least-squares fit was obtained over the range Kobs > 10-3 as shown by the minimum value for log RSS. Over this range the estimates of Msys, m, and the ratio τobs/Kobs were constant. This process was repeated for each curve in Fig. 9a, and a least-squares fit was always obtained over the domain Kobs > 10-3. The estimates ± S.E. of Msys and m were 55 ± 1.4% inhibition and 0.68 ± 0.037, respectively. The RA values were calculated from the τobs/Kobs ratios estimated with Kobs constrained to an arbitrarily high value (i.e., 10-2), and these estimates are also listed in Table 5 and plotted in Fig. 9b. Regression analysis of the data according to eq. 12 (Fig. 9c) yielded estimates of KA and γ of 0.62 μM and 0.019, respectively (mean ± S.E.M.; pKA = 6.21 ± 0.15; log γ = -1.72 ± 0.16).
For heuristic purposes, the RA values were also estimated using eq. 25. Generally, the use of eq. 25 is restricted to data with Hill slopes of 1, but as described above, it is applicable in situations where there is little change in Emax. These estimates were made for each experiment and the mean values of four experiments ± S.E.M. are listed in Table 5. The corresponding log RA values are also plotted against the log of the gallamine concentration in Fig. 9c. Regression analysis of the data according to eq. 12 yielded estimates of KA and γ of 0.56 μM and 0.025, respectively (mean ± S.E.M.; pKA = 6.25 ± 0.21; log γ = -1.60 ± 0.21).
Discussion
The study of allosterism in functional assays provides potential information about the allosteric modulation of the intrinsic efficacy of the agonist-receptor complex. This information is important because the therapeutic effects of allosteric modulators depend on their modulation of both the affinity and intrinsic efficacy of the endogenous ligand. The theoretical basis for the present analysis was described in a previous report (Ehlert, 1988a), where it was shown that simple graphical techniques can be used to estimate the affinity of the allosteric modulator and its combined effects on the affinity and intrinsic efficacy (γ) of the agonist-receptor complex when the receptor reserve is great. Given the widespread availability of software for nonlinear regression analysis, the time seemed appropriate to reinvestigate this method and to apply it to all conditions. To this end, the concept of relative activity of the agonist (RA) is developed here and shown to be a robust measure of allosteric effects.
The present report shows that it is always possible to estimate the influence of the allosteric modulator on the RA value of the agonist. Regression analysis of the RA values according to the logarithmic form of eq. 12:  provides estimates of the dissociation constant (KA) of the allosteric modulator as well as the product (γ) of its modulatory effects on the intrinsic efficacy (β) and affinity (1/α) of the agonist-receptor complex. In situations where the Hill slopes of the agonist concentration-response curves are equal to 1, the RA values can be estimated easily from the EC50 and Emax values (eq. 25). If the allosteric modulator has no influence on the Emax of the agonist, it is impossible to determine the individual components of γ (α and β) unless an independent estimate of α is made, perhaps through ligand binding analysis (Lazareno and Birdsall, 1995). If the allosteric modulator does influence the Emax of the agonist, then it is possible to estimate the individual components α and β as described under Results. The method involves global nonlinear regression analysis with the operational model (eq. 16) to obtain individual estimates of τobs and Kobs.
provides estimates of the dissociation constant (KA) of the allosteric modulator as well as the product (γ) of its modulatory effects on the intrinsic efficacy (β) and affinity (1/α) of the agonist-receptor complex. In situations where the Hill slopes of the agonist concentration-response curves are equal to 1, the RA values can be estimated easily from the EC50 and Emax values (eq. 25). If the allosteric modulator has no influence on the Emax of the agonist, it is impossible to determine the individual components of γ (α and β) unless an independent estimate of α is made, perhaps through ligand binding analysis (Lazareno and Birdsall, 1995). If the allosteric modulator does influence the Emax of the agonist, then it is possible to estimate the individual components α and β as described under Results. The method involves global nonlinear regression analysis with the operational model (eq. 16) to obtain individual estimates of τobs and Kobs.

Estimation of the modulatory effects of an allosteric inhibitor using a null method. The data are from Fig. 1a. a, pairs of equiactive agonist concentrations in the absence and presence of the allosteric modulator are estimated. This plot is for the data simulated for A = 10-3 M; a similar analysis was done for the other concentrations of allosteric modulator. b, logarithms of the equiactive agonist concentrations are analyzed by nonlinear regression analysis using eq. 9. The curves are analyzed simultaneously sharing the estimates of KX among the curves and estimating a unique value of p and q for each curve. c, summary of nonlinear regression analysis for the data simulated at A = 10-3 M. The value of KX was constrained as a constant over the range of values shown on the abscissa and the values of the other parameters that yielded the least-squares fit to the simulated data were estimated by regression analysis as shown by the indicated curves. It can be seen that the estimates of p and q are proportional to one another such that their ratio (q/p or RA) remains constant over the range of KX values shown.
For heuristic purposes, this analysis has emphasized the utility of the RA estimate in understanding allosteric interactions. Consequently, the log RA value has been treated as the dependent variable in nonlinear regression analysis. However, this strategy might not be the best statistical approach when the RA value is estimated from the EC50 and Emax values in situations where the Hill slope is equivalent to 1. For example, if there is error in the estimates of the Emax and EC50 values of the control concentration-response curve, then this error will introduce a systematic error in the RA estimates for all the concentration-response curves measured in the presence of the allosteric modulator. A better strategy might be to calculate log ratio of the Emax/EC50 values measured in the absence and presence of the various concentrations of the allosteric modulator and use the following equation to analyze the data by nonlinear regression analysis:  Regression analysis should yield estimate of γ, KA, and P, the log ratio of Emax′/EC50′ for the control agonist concentration-response curve. The latter equation can be derived from eqs. 12 and 25, and it only applies to situations where the Hill slope of the concentrations response curves are equivalent to 1. In addition, when the Hill slope is equivalent to 1 and the allosteric modulator has no influence on Emax, the above-mentioned equation reduces to that described previously (Ehlert, 1988b):
Regression analysis should yield estimate of γ, KA, and P, the log ratio of Emax′/EC50′ for the control agonist concentration-response curve. The latter equation can be derived from eqs. 12 and 25, and it only applies to situations where the Hill slope of the concentrations response curves are equivalent to 1. In addition, when the Hill slope is equivalent to 1 and the allosteric modulator has no influence on Emax, the above-mentioned equation reduces to that described previously (Ehlert, 1988b):  Regression analysis would yield estimates of γ, KA, and the log control EC50 value. A similar statistical argument might seem appropriate when the RA values are estimated from Kobs and τobs (i.e., error in the estimates of KX and τ will introduce a systematic error in the estimate of RA); however, this concern is unnecessary because the values of KX and τ (Kobs′ and τobs′) are estimated by global nonlinear regression analysis (eq. 10) of all of the agonist concentration-response curves simultaneously. Thus, all of the data are considered in the estimation of KX and τ and not just the control data. It should be apparent from the foregoing that it is possible to estimate the dissociation constant of the agonist (KX) through the analysis of an allosteric modulator that influences the intrinsic efficacy of the agonist to such an extent that a change in Emax is observed.
Regression analysis would yield estimates of γ, KA, and the log control EC50 value. A similar statistical argument might seem appropriate when the RA values are estimated from Kobs and τobs (i.e., error in the estimates of KX and τ will introduce a systematic error in the estimate of RA); however, this concern is unnecessary because the values of KX and τ (Kobs′ and τobs′) are estimated by global nonlinear regression analysis (eq. 10) of all of the agonist concentration-response curves simultaneously. Thus, all of the data are considered in the estimation of KX and τ and not just the control data. It should be apparent from the foregoing that it is possible to estimate the dissociation constant of the agonist (KX) through the analysis of an allosteric modulator that influences the intrinsic efficacy of the agonist to such an extent that a change in Emax is observed.
The present report also describes a null method, whereby the values of KA and γ were determined by nonlinear regression analysis of pairs of equiactive agonist concentrations in the absence and presence of various concentrations of the allosteric modulator. This type of approach should be applicable in situations where the response of the agonist deviates from logistic behavior.
The estimates of the KA (0.56–0.62 μM) and γ (0.019–0.025) values for gallamine antagonism of the action of oxotremorine-M at M2 muscarinic receptors made in this report by regression analysis of the log RA values are in good agreement with those (0.52 μM and 0.020) made in a previous analysis of the same data using regression analysis of the log EC50 values according to eq. 29 above (Ehlert, 1988b). This agreement shows the feasibility of estimating RA values from experimental data. The estimate of the KA of gallamine reported here is also in good agreement with that (0.93 μM) estimated by antagonism of the negative inotropic effect of acetylcholine in the isolated, electrically driven, guinea pig atrium (Christopoulos, 2000) and by competitive inhibition of the binding of [3H]N-methylscopolamine to myocardial muscarinic receptors (Stockton et al., 1983; Ehlert, 1988b) (0.77 and 1.1 μM, respectively).

Analysis of the antagonism of oxotremorine-M-mediated inhibition of adenylyl cyclase activity by gallamine in homogenates of the rat myocardium. a, adenylyl cyclase activity measured in the presence of various concentrations of both oxotremorine-M and gallamine. The data are from Ehlert (1988b). Mean values ± S.E.M. from four experiments are shown. b, summary of global nonlinear regression analysis of the concentration-response curves shown in a at gallamine 10-4 M according to eq. 16. All curves were fitted simultaneously sharing the estimate of m and Msys among the curves and constraining the value of Kobs as a constant at various values between 10-6 and 100. Regression analysis was used to estimate the values of the various parameters in eq. 16 that yielded the best least-squares fit to the data. The plot shows how the estimate of τobs for the data simulated at gallamine 10-4 M is proportional to Kobs such that their ratio (τobs/Kobs) remains constant at approximately 3.7 × 104. c, effect of gallamine on the RA values of oxotremorine-M. The plots represent an analysis of the data shown in a, and the estimates of RA were calculated from the Kobs and τobs values or the Emax and EC50 values listed in Table 5.
In a previous report on the antagonism of the action of the highly efficacious agonist oxotremorine-M and the partial agonist BM-5 at cardiac M2 muscarinic receptors by gallamine, the estimates of γ were calculated to be 0.020 and 0.011, respectively (Ehlert, 1988b). In both cases, gallamine had little or no effect on the Emax values of the agonists. Since BM-5 behaved as a partial agonist, any gallamine-induced change in the intrinsic efficacy of BM-5 should have been manifest as an alteration in Emax. The lack of this change indicates that β = 1 and α = 0.011, approximately, for the allosteric interaction between gallamine and BM-5 (i.e., γ = α/β = 0.011). Additional proof for this hypothesis could be obtained if a similar estimate of the α value for the interaction between gallamine and BM-5 was obtained in ligand binding studies using the technique described by Lazareno and Birdsall (1995).
The presumed lack of effect of gallamine on the intrinsic efficacy of BM-5 has important implications with regard to the action of gallamine and the number of ground and active states of the M2 muscarinic receptor. In a simple two-state model, where there is only one active and one inactive state of the receptor, there should be a correlation between allosteric effects and the intrinsic efficacy of the primary ligand (Ehlert, 2000). However, gallamine has been shown to inhibit the binding of both agonists and antagonists to M2 muscarinic receptors. Moreover, if gallamine reduces the affinity of BM-5 without influencing its intrinsic efficacy, this result implies the existence of another pair of ground and active states of the M2 receptor that are selected by gallamine and whose affinities for BM-5 are both lower by the same amount so that the selectivity of BM-5 for the two states is the same, and hence its efficacy unaltered (i.e., the ratio of microscopic affinity constants of BM-5 for the ground and active states are the same). Alternatively, a more complicated hypothesis involving a group of active and inactive states might explain the data.
Part of the analysis described herein is analogous to a method previously described for analyzing a series of concentration-response curves to different agonists with varying affinity and intrinsic efficacy. In that previous analysis, the concept of intrinsic relative activity was developed (Ehlert et al., 1999; Ehlert and Griffin, 2001). This term denotes the product of the affinity and intrinsic activity of the agonist expressed relative to that of a standard agonist. This estimate is analogous to the RA value described here, which is a measure of the product of affinity and intrinsic efficacy of the agonist in the presence of the allosteric modulator relative to that measured in its absence. Accurate measurements of the components of allosterism are important and likely to be more significant as the number of allosteric drugs used in therapeutics and research increases.
Footnotes
-
This work was supported by National Institutes of Health Grant 69829. Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.105.090886.
-
ABBREVIATIONS: RA, relative activity; RSS, residual sum of squares; BM-5, N-(4-pyrrolidino-2-butynyl)-N-methylacetamide.
- Received June 10, 2005.
- Accepted July 20, 2005.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}