


Abstract
κ Opioid agonists were at one time proposed as candidate pharmacotherapies for cocaine addiction, mainly because of their ability to decrease dopamine neurotransmission and attenuate the behavioral effects of cocaine in laboratory animals. Recent studies, however, suggest that κ agonists also may mimic and/or enhance some of the effects of cocaine through mechanisms related to stress. The current study used a reinstatement procedure to examine the ability of the κ agonists spiradoline and enadoline to reinstate extinguished cocaine seeking in squirrel monkeys previously trained to self-administer cocaine under a second-order schedule of i.v. drug injection. Opioid- and stress-related mechanisms were evaluated in antagonism studies with the opioid antagonists naltrexone and nor-binaltorphimine (nor-BNI), the corticotropin-releasing factor receptor antagonist butyl-ethyl-[2,5-dimethyl-7-(2,4,6-trimethylphenyl)-7H-pyrrolo [2,3-d]pyrimidin-4-yl]amine (CP 154,526), and the α2-adrenoceptor agonist clonidine combined with either spiradoline or enadoline. When tested alone, priming with spiradoline and enadoline induced significant reinstatement of cocaine-seeking behavior to approximately 45% of the maximum reinstatement induced by cocaine. Reinstatement of cocaine seeking induced by intermediate doses of spiradoline was greater in the presence than in the absence of response-contingent presentations of a cocaine-paired stimulus. Spiradoline- and enadoline-induced reinstatement of drug seeking was attenuated by naltrexone but not by nor-BNI. Spiradoline-induced reinstatement of cocaine seeking was also antagonized by CP 154,526 and clonidine. The results point to interactions between a subpopulation of κ opioid receptors and central corticotropin-releasing factor and noradrenergic stress systems in the reinstatement of cocaine seeking induced by κ agonists.
κ Opioid agonists have been proposed as potential pharmacotherapies for cocaine addiction because of their ability to decrease dopamine neurotransmission (Devine et al., 1993) and suppress certain behavioral effects of cocaine (Mello and Negus, 1998; Schenk et al., 1999; Beardsley et al., 2005). Recent evidence suggests, however, that reductions in cocaine-related behaviors induced by κ agonists may be secondary to a generalized response-suppressing effect of these drugs, and that the interaction between cocaine and κ agonists depends on the context and timing of their administration. When tested using a choice procedure of cocaine versus food reinforcement, rhesus monkeys showed an overall decrease in response rate but an increase in cocaine preference after intermittent infusions of the κ agonist U50,488 (Negus, 2004). McLaughlin et al. (2006a) demonstrated enhancement of cocaine-conditioned place preference when mice were given U50,488 1 h before cocaine and suppression of cocaine-conditioned place preference when U50,488 was administered at a later time point.
The effects of κ agonists on cocaine preference may partially reflect activation of physiological stress mechanisms, as κ opioid receptors and the endogenous κ opioid peptide dynorphin have been shown to regulate aspects of the stress response. For example, mice subjected to force swim stress before administration of cocaine showed enhanced cocaine-conditioned place preference (McLaughlin et al., 2006a). Administration of the κ antagonist nor-binaltorphimine (nor-BNI) or disruption of the prodynorphin gene blocked enhancement of the cocaine-conditioned place preference induced by U50,488, forced swimming, or social defeat (McLaughlin et al., 2006a,b). Stressful stimuli also can induce reinstatement of drug seeking in animals trained to self-administer cocaine (Shaham et al., 1998; Erb et al., 2000), an effect that can be attenuated by pretreatment with a κ opioid antagonist (Beardsley et al., 2005).
κ Receptor mechanisms appear to play a role in the regulation of stress in other situations. People who have taken κ agonists report adverse emotional and perceptual experiences (Pfeiffer et al., 1986). Furthermore, κ opioid antagonists significantly attenuate stress-induced analgesia and stress-induced immobility in mice (McLaughlin et al., 2006a). Immobilization and forced swim stress enhance dynorphin immunoreactivity in the hippocampus and nucleus accumbens, and administration of a κ antagonist into the hippocampus attenuates “learned helplessness” in rats (Shirayama et al., 2004).
κ Opioid modulation of stress responses may be partially due to interactions with other neurotransmitter systems that serve to regulate stress. Corticotropin-releasing factor (CRF) and norepinephrine (NE) play key roles in the regulation of stress-induced behavior (Koob, 1999), including stress-induced relapse to cocaine seeking. CRF receptor antagonists and α2-adrenoceptor agonists attenuate reinstatement of cocaine seeking induced by footshock in rats (Erb and Stewart, 1999; Erb et al., 2000) and pharmacological (yohimbine-induced) stress in squirrel monkeys (Lee et al., 2004; G. R. Valdez, unpublished observations). The ability of CRF antagonists to attenuate the reinstatement of drug seeking appears to be specific to stress-induced relapse because these drugs do not consistently block cocaine-induced reinstatement of drug seeking (Erb and Stewart, 1999; Lee et al., 2003). The ability of α2-adrenoceptor agonists to attenuate the reinstatement of cocaine seeking also appears to be specific to stress in rats (Erb et al., 2000), although it is less so in monkeys (Lee et al., 2003; Platt et al., 2007).
With regard to interactions between κ opioids and CRF, dynorphin is colocalized with CRF in the hypothalamus (Roth et al., 1983), and administration of κ agonists can increase CRF release (Buckingham and Cooper, 1986). κ Agonists stimulate the hypothalamic-pituitary-adrenal axis in rodents (Laorden et al., 2000) and humans (Ur et al., 1997), which is primarily regulated by CRF release in the hypothalamus (Vale et al., 1983). Within the noradrenergic system, κ agonists increase NE turnover in the paraventricular nucleus of the hypothalamus (Laorden et al., 2000) and NE release in the hippocampus (Jackisch et al., 1986). The κ agonist spiradoline has also been shown to increase NE metabolism (Kunihara et al., 1992).
Although there is evidence that κ agonists can suppress cocaine-induced reinstatement in rats (cf. Schenk et al., 1999), the above considerations suggest that κ agonists in the absence of cocaine might precipitate reinstatement of cocaine seeking via a stress-related mechanism. In the present study we sought to investigate this hypothesis by examining the ability of κ agonists to induce reinstatement of cocaine seeking in monkeys and by determining the extent to which the response-reinstating effects of these drugs could be blocked by the opioid antagonists naltrexone and nor-BNI, the CRF1 antagonist CP 154,526, and the α2-adrenoceptor agonist clonidine.
Materials and Methods
Subjects. Twelve adult male squirrel monkeys (Saimiri sciureus), weighing 0.7 to 1.0 kg, were housed individually in a climate-controlled vivarium where they had unlimited access to water and received a nutritionally balanced diet of monkey chow (Harlan Teklad, Madison WI) supplemented with fresh fruit. Four monkeys were experimentally naive at the beginning of the study, and eight monkeys had previously served in experiments involving cocaine self-administration and reinstatement of cocaine-seeking behavior. All monkeys were maintained in accordance with the guidelines of the Committee on Animals of the Harvard Medical School and of the Guide for the Care and Use of Laboratory Animals (Institute of Laboratory Animal Resources on Life Sciences, National Research Council, National Academy of Sciences, 1996).
Surgery. Indwelling venous catheters (polyvinyl chloride, inside diameter: 0.38 mm; outside diameter: 0.76 mm) were implanted in each monkey using aseptic surgical procedures as described by Platt et al. (2005). Briefly, monkeys were anesthetized with isoflurane, and one end of the catheter was passed by way of either a jugular or femoral vein to the level of the right atrium. The distal end of the catheter was passed s.c. to a midscapular exit point. Catheters were flushed daily with 0.9% saline solution and were sealed with stainless steel obturators when not in use. Monkeys wore nylon-mesh jackets (Lomir Biomedical, Toronto, ON, Canada) at all times to protect the catheter.
Apparatus. Experimental sessions were conducted in ventilated sound-attenuated chambers, which were provided with white noise to mask external sounds. Within the chamber, monkeys were seated in a Plexiglas chair facing a panel that was equipped with a response lever and colored stimulus lights above the lever (MED Associates, St. Albans, VT). Catheters were connected to motor-driven syringe pumps located outside the chamber. Each operation of the pump lasted 1 s and delivered a volume of 0.18 ml into the catheter.
Second-Order Schedule of Cocaine Self-Administration. Monkeys were trained to self-administer cocaine under a second-order schedule of i.v. drug injection similar to the schedule described by Khroyan et al. (2000). Briefly, in the presence of a white light, completion of every 10th or 30th response (FR10 or FR30, depending on the subject) during a 10-min fixed interval (FI) resulted in a 2-s change in illumination from white to red. Completion of the first complete FR after expiration of the FI resulted in an i.v. injection of cocaine simultaneous with the onset of the 2-s red light (cocaine-paired stimulus). A 60-s time-out (TO) period, during which all lights were off and responses had no scheduled consequences, followed each cocaine-stimulus pairing. If the FR requirement was not completed within 8 min after the expiration of the FI, the component ended automatically without an injection followed by a 60-s TO. Daily sessions ended after completion of five cycles of the second-order schedule or a maximum time of 95 min. Initially, the dose of self-administered cocaine was varied over a 3 to 10-fold range to determine the dose that maintained the maximum rates of responding for each monkey. The selected doses (0.18 mg/kg/injection in eight monkeys and 0.3 mg/kg/injection in four monkeys) were then kept constant during periods of cocaine self-administration for the remainder of the experiment.
Extinction. After at least 1 month of stable cocaine self-administration under the second-order schedule, responding was extinguished by substituting saline for cocaine and omitting presentations of the cocaine-paired stimulus. Extinction sessions were otherwise identical to the self-administration sessions described above. Extinction sessions were conducted daily until responding declined to ≤10% of the response rate maintained by cocaine self-administration for at least three consecutive sessions.
Priming-Induced Reinstatement of Drug Seeking. After extinction, a range of priming doses of cocaine and the κ agonists spiradoline and enadoline were tested for their ability to reinstate drug-seeking behavior during test sessions in which only saline was available for self-administration. Response-contingent presentations of the cocaine-paired stimulus also were restored during test sessions because earlier studies showed that reinstatement of cocaine-seeking behavior was greatest when priming injections were accompanied by restoration of the cocaine-paired stimulus (Spealman et al., 2004). Priming injections were administered i.v. immediately before the session followed by a saline flush to clear the catheter of residual drug solution. The drugs were studied in different orders in different subjects, but cocaine was studied first in most cases. Different doses of each drug and vehicle were tested on different days, with each test session separated by one or more extinction sessions as described above. The order of testing was varied across monkeys with the restriction that all doses of a particular drug be tested before tests with another drug were initiated. Between experiments with different drugs, cocaine self-administration was reestablished under the second-order schedule of i.v. drug injection and then extinguished again according to the criteria described above. In previous studies, periodically reestablishing and then extinguishing cocaine self-administration between test periods resulted in consistent cocaine-induced reinstatement of responding over the course of a year or more (Khroyan et al., 2000).
Antagonism of κ Agonist-Induced Reinstatement of Drug Seeking. After completion of the drug priming experiments described above, cocaine self-administration was reestablished and then extinguished again before initiation of drug pretreatment experiments with the opioid antagonists naltrexone and nor-BNI, the CRF1 antagonist CP 154,526, and the α2-adrenoceptor agonist clonidine. Experiments with naltrexone, CP 154,526, clonidine, and their respective vehicles were conducted by injecting the drugs 10 min before an i.v. priming dose of spiradoline or enadoline. Experiments with nor-BNI were conducted by injecting the drug at least 2 days before priming with spiradoline or enadoline. This latter pre-treatment protocol was selected on the basis of previous studies showing that nor-BNI exhibits selective κ antagonist effects beginning 1 or 2 days after administration and lasting at least 25 days after administration in squirrel monkeys (Carey and Bergman, 2001). Only one dose of nor-BNI was tested because the results obtained after administration of the 10 mg/kg dose precluded the testing of lower doses that were previously shown to be effective in squirrel monkeys. Experiments with different priming doses of spiradoline or enadoline in the presence of nor-BNI were conducted over two separate 18-day periods separated by 1 or more weeks of reestablished cocaine self-administration and subsequent extinction. Extinction levels remained consistent for all subjects throughout this period. Priming doses of spiradoline or enadoline were tested in irregular order across subjects to control for any possible diminishing effects of nor-BNI over time.
Priming with Spiradoline in the Presence and Absence of the Cocaine-Paired Stimulus. Because combining pharmacological stress with cocaine-paired stimuli has been reported to potentiate reinstatement of drug seeking in rats (Shelton et al., 2004; Shelton and Beardsley, 2005), an additional study was conducted to directly compare the level of reinstatement induced by spiradoline in the presence and absence of response-contingent presentations of the cocaine-paired stimulus. These studies were conducted by administering priming injections of spiradoline before test sessions in which the cocaine-paired stimulus was either presented (as in experiments described previously) or was omitted entirely (as during extinction, see above).
Drugs. Cocaine hydrochloride, enadoline hydrochloride [[5R-(5α,7α,8β)]-N-methyl-N-[7-(1-pyrrolidinyl)-1-oxaspiro[4.5]dec-8-yl]-4-benzofuranacetamide hydrochloride], spiradoline mesylate [(5α,7α,8β)-(±)-3,4-dichloro-N-methyl-N-(7-[1-pyrrolidinyl]-1-oxaspiro[4,5]dec-8-yl)benzeneacetamide methane mesylate], naltrexone hydrochloride [(5a)-17-(cyclopropylmethyl)-4,5-epoxy-3, 14-dihydroxymorphinan-6-one hydrochloride], nor-BNI dihydrochloride [4,8:11,15-dimethano-20H-bisbenzofuro[2,3-α:3′,2′-i] dipyrido[4,3-β:3′,4′-h]carbazole-1,8α,10α,18-tetrol, 7,12-bis(cyclopropyl methyl)-5,6,7,8,9,10,11,12,13,14,19α,20β-dodecahydro-(4βS,8R,8αS,10αS,11R,14αS,19αR,20βR)], and clonidine hydrochloride [N-(2,6-dichlorophenyl)-4,5-dihydro-1H-imidazol-2-amine hydrochloride] were dissolved in 0.9% saline solution. CP 154,526 was dissolved in 40% 0.1 N HCl and 60% propylene glycol and then diluted to the desired concentration with 0.9% saline solution. Cocaine, spiradoline, clonidine, and nor-BNI were purchased from commercial sources (Sigma/RBI, St. Louis, MO; Tocris Bioscience, Ellisville, MO). Enadoline and CP 154,526 were generously provided by Parke-Davis Pharmaceuticals (Ann Arbor, MI) and Pfizer Inc. (Groton, CT), respectively.
Data Analysis. The rate of responding in individual monkeys was computed each session by dividing the total number of responses by the total session time (excluding responses and time during TO periods). Mean ± S.E.M. response rates were then calculated for groups of 4 to 12 monkeys, depending on the particular experiment. Reinstatement of drug seeking induced by cocaine, spiradoline, and enadoline and the effects of pretreatment with naltrexone were analyzed by one-way repeated measures ANOVAs followed by Bonferroni t tests for planned comparisons (α level, p < 0.05). Two-way repeated measures ANOVAs followed by Bonferroni t tests were used to analyze the effects of pretreatment with nor-BNI, CP 154,526, and clonidine on spiradoline- and enadoline-induced reinstatement of drug seeking and the effects of spiradoline with and without presentations of the cocaine-paired stimulus. Other specific comparisons were made using paired t tests.
Results
Consistently high rates of responding were maintained for self-administered cocaine under the second-order schedule of i.v. drug injection throughout the study. Averaged over all cocaine self-administration sessions that preceded extinction sessions, the mean ± S.E.M. response rate for the group of 12 monkeys was 0.86 ± 0.14 response/s. Monkeys received all five injections of cocaine (0.18 or 0.3 mg/kg/injection) during self-administration sessions. During extinction sessions that preceded drug priming sessions, responding declined in all monkeys to a mean response rate of 0.05 ± 0.01 response/s.
Priming with Cocaine, Spiradoline, and Enadoline. Priming injections of cocaine immediately before test sessions in which the cocaine-paired stimulus also was restored resulted in a dose-related reinstatement of drug seeking (F3,33 = 15.47, p < 0.001, n = 12) (Fig. 1A). Planned comparisons showed significantly higher response rates after priming with 0.3 and 1.0 mg/kg compared with vehicle (Bonferroni t test, p < 0.05). The maximum level of reinstated cocaine seeking (0.83 ± 0.15 response/s) was comparable with the level of responding maintained by active cocaine self-administration (see above).
Priming with either spiradoline or enadoline also induced a dose-related reinstatement of cocaine seeking (Fig. 1, B and C). Separate repeated measures ANOVAs showed a significant effect of dose for both drugs (spiradoline: F4,40 = 2.80, p < 0.038, n = 11; enadoline: F4,20 = 4.74, p < 0.007, n = 6). Response rates were significantly higher after 0.01 and 0.3 mg/kg spiradoline and 0.001 and 0.1 mg/kg enadoline compared with their respective vehicles (p < 0.05). The maximum rate of reinstated responding induced by either spiradoline (0.33 ± 0.04 response/s) or enadoline (0.37 ± 0.13 response/s) was, however, less than the maximum response rate induced by cocaine. Paired t tests revealed a significant difference in response rates induced by a maximally effective dose of spiradoline compared with cocaine (p < 0.02) and a difference that approached significance in response rates induced by a maximally effective dose of enadoline compared with cocaine (p < 0.06).

Reinstatement of extinguished cocaine seeking by priming with cocaine (A), spiradoline (B), and enadoline (C) in squirrel monkeys. Data are means ± S.E.M. (n = 6–12). Abscissas: dose, logarithmic scale. Ordinates: response rate. Points above V show the effects of vehicle priming. *, significant differences from vehicle control (p < 0.05).
Pretreatment with Naltrexone and Nor-BNI. Pre-treatment with naltrexone administered 10 min before priming with a maximally effective dose of either spiradoline (0.3 mg/kg) or enadoline (0.01 mg/kg) resulted in a dose-related antagonism of spiradoline- and enadoline-induced reinstatement of cocaine seeking (Fig. 2). Separate repeated measures ANOVAs showed a significant main effect of naltrexone dose combined with either spiradoline (F4,15 = 3.75, p < 0.03, n = 7) or enadoline (F4,12 = 4.18, p < 0.03, n = 4). Pretreatment with 1.0 mg/kg naltrexone significantly attenuated the effects of spiradoline (p < 0.05) and enadoline (p < 0.05). Additional post hoc comparisons (paired t test) showed that the rate of responding induced by spiradoline or enadoline after pretreatment with 1.0 mg/kg naltrexone did not differ significantly from the rate of responding during extinction (i.e., in the absence of any drug).
Pretreatment with 10.0 mg/kg nor-BNI a minimum of 48 h before reinstatement testing (see Materials and Methods) did not significantly alter the priming effects of either spiradoline or enadoline compared with the effects of the two drugs in the absence of nor-BNI (Fig. 3). Separate two-way repeated measures ANOVAs with presence versus absence of nor-BNI and dose of either κ agonist as factors revealed no main effect of nor-BNI (F1,7 = 4.05, p = 0.16, n = 8 in the spiradoline study; F1,3 = 0.54, p = 0.51, n = 4 in the enadoline study) and no significant interaction between nor-BNI and either spiradoline (F2,14 = 0.94, p = 0.42) or enadoline (F2,6 = 1.86, p = 0.24).
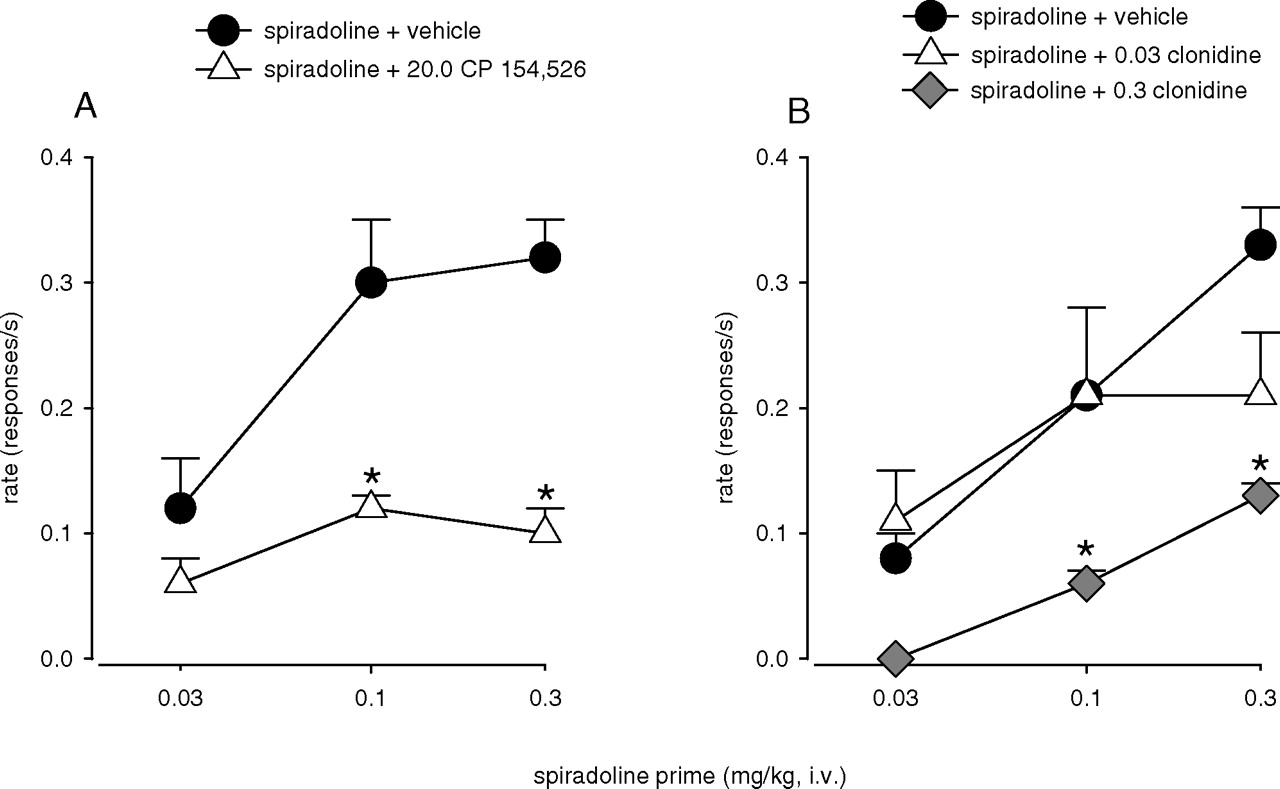
Pretreatment with CP 154,526 and Clonidine. Pre-treatment with 20.0 mg/kg CP 154,526 10 min before priming with spiradoline resulted in a marked reduction in spiradoline-induced reinstatement of cocaine seeking compared with pretreatment with vehicle (Fig. 4A). A two-way repeated measures ANOVA with doses of spiradoline and CP 154,526 as factors revealed a significant main effect of spiradoline (F2,8 = 13.31, p = 0.003) and CP 154,526 (F1,4 = 24.63, p = 0.008) and a significant spiradoline × CP 154,526 interaction (F2,8 = 6.03, p = 0.025, n = 5). Bonferroni t tests revealed that pretreatment with 20.0 mg/kg CP 154,526 significantly attenuated the reinstatement of drug seeking induced by 0.1 and 0.3 mg/kg spiradoline (p < 0.05).

Effects of pretreatment with naltrexone on the reinstatement of drug seeking induced by maximally effective doses of spiradoline (0.3 mg/kg; A) and enadoline (0.01 mg/kg; B). Data are means ± S.E.M. (n = 7). Points above V show the effects of pretreatment with vehicle before priming with spiradoline or enadoline. *, significant differences from spiradoline or enadoline after pretreatment with vehicle (p < 0.05). Other details are as in the legend to Fig. 1.

Effects of pretreatment with 10.0 mg/kg nor-BNI on the reinstatement of drug seeking induced by spiradoline (A) and enadoline (B). Data are means ± S.E.M. (n = 4–8). •, effects of the κ agonists alone; ▵, effects of the κ agonists tested 2 to 18 days after administration of nor-BNI (see Materials and Methods). Other details are as in the legend to Fig. 1.
Pretreatment with clonidine 10 min before priming with spiradoline also resulted in a substantial reduction in spiradoline-induced reinstatement of drug seeking compared with pretreatment with vehicle (Fig. 4B). Two-way repeated measures ANOVA with doses of spiradoline and clonidine as factors revealed significant main effects of both spiradoline (F2,8 = 17.37, p < 0.001) and clonidine (F2,8 = 5.58, p = 0.037). Although no significant spiradoline × clonidine interaction was detected (F4,12 = 1.80, p = 0.195), Bonferroni t tests showed a significant attenuation of reinstated drug seeking induced by 0.1 and 0.3 mg/kg spiradoline after pre-treatment with 0.3 mg/kg clonidine (p < 0.05).
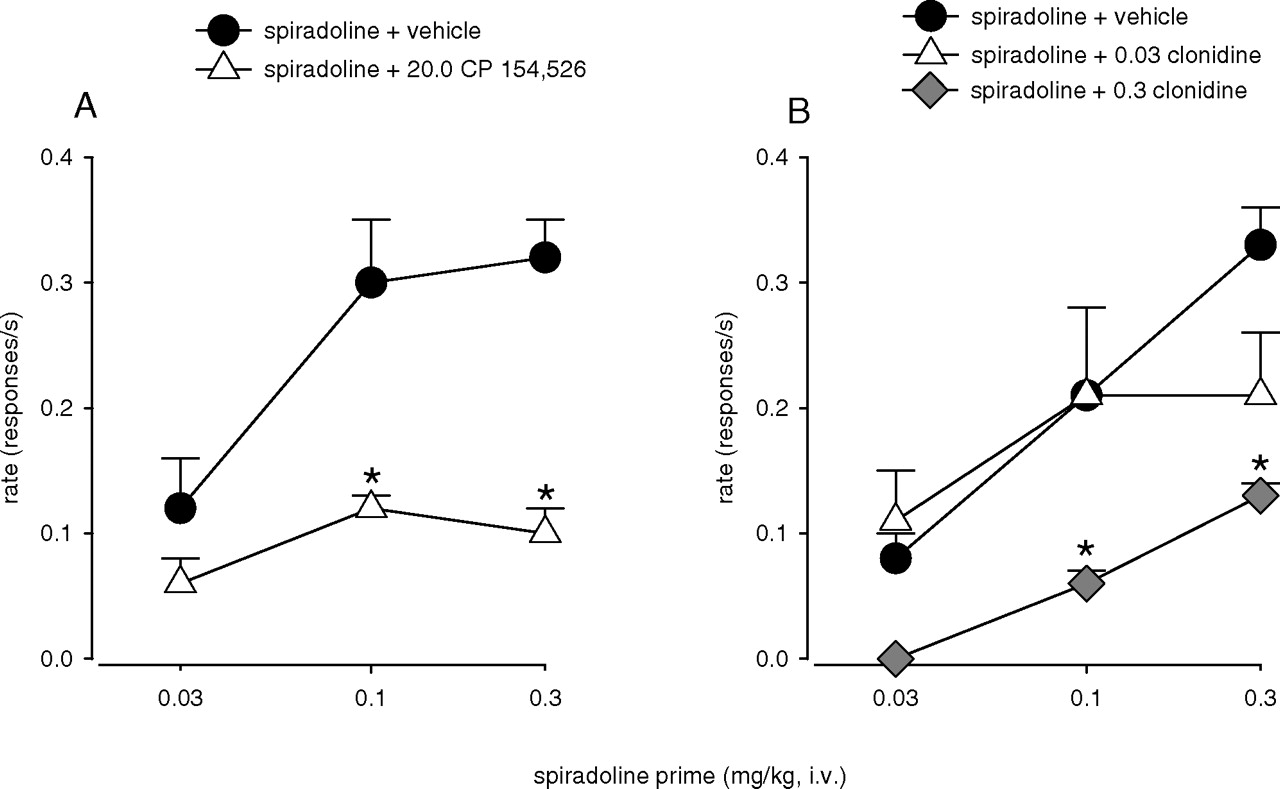
Effects of pretreatment with 20.0 mg/kg CP 154,526 (A) and 0.03 and 0.3 mg/kg clonidine (B) on spiradoline-induced reinstatement of drug seeking. Data are means ± S.E.M. (n = 5). Filled symbols, effects of spiradoline after pretreatment with vehicle; other symbols, effects of spiradoline after pretreatment with CP 154,526 or clonidine. *, significant differences from spiradoline after vehicle pretreatment. Other details are as in the legend to Fig. 1.
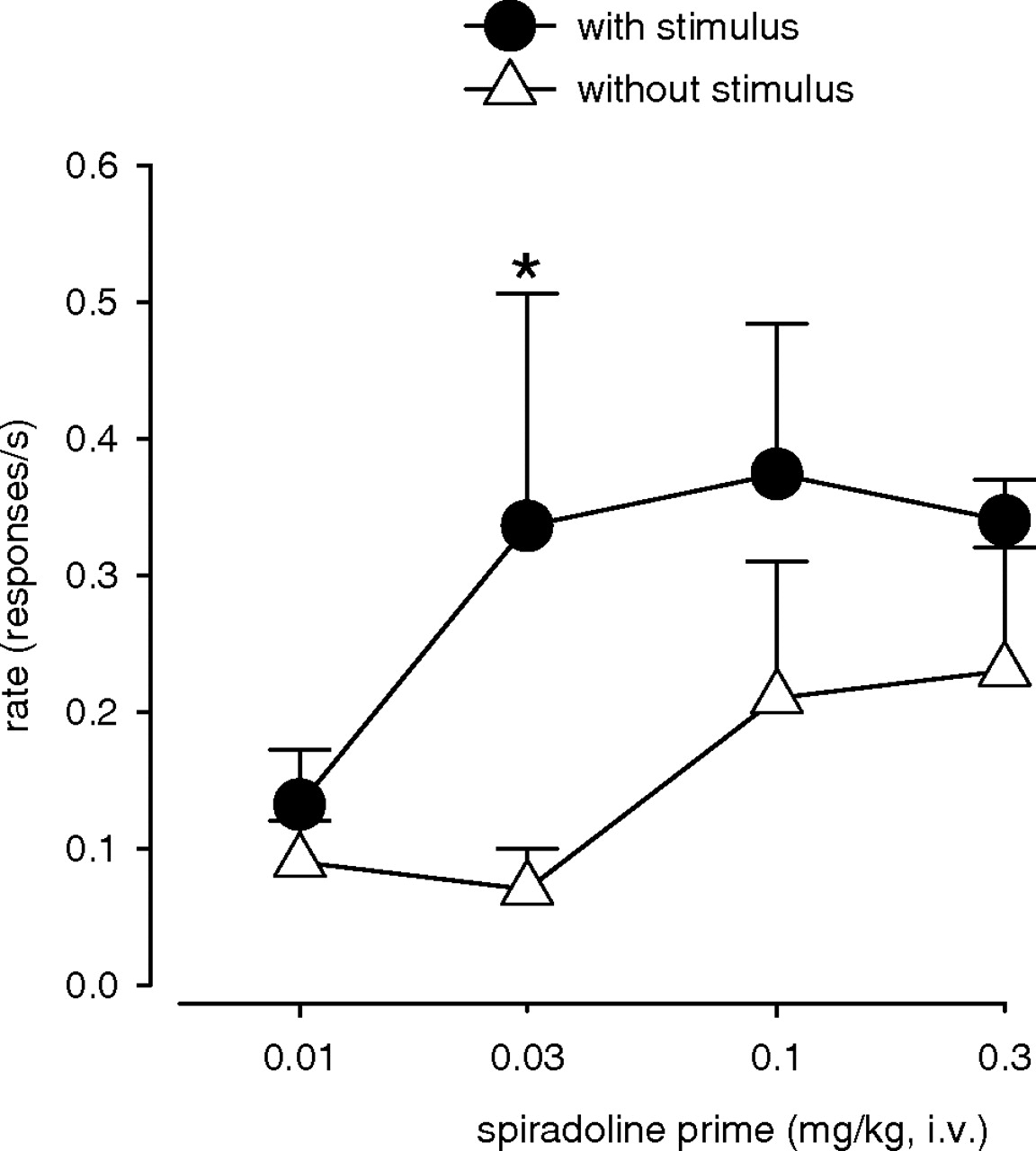
Effects of Presence versus Absence of the Cocaine-Paired Stimulus. Because combining pharmacological stress with cocaine-paired stimuli has been reported to potentiate the reinstatement of cocaine seeking in rats (Shelton and Beardsley, 2005; Feltenstein and See, 2006), we compared the effects of priming with spiradoline in the presence and absence of response-contingent presentations of the cocaine-paired stimulus (Fig. 5). Two-way repeated measures ANOVA with dose of spiradoline and presence versus absence of the stimulus as factors revealed a significant main effect of spiradoline (F4,16 = 4.03, p < 0.02) and the cocaine-paired stimulus (F1,4 = 25.53, p < 0.01), but no significant interaction (F4,16 = 1.82, p = 0.175). Planned comparisons, however, showed that the effects of 0.03 mg/kg spiradoline were significantly greater in the presence than in the absence of response-contingent presentations of the cocaine-paired stimulus (p < 0.05).
Discussion
In the present study priming injections of the κ agonists spiradoline and enadoline induced significant reinstatement of cocaine seeking in squirrel monkeys previously trained to self-administer cocaine. The effects of the κ agonists appear to be opioid receptor-mediated because spiradoline- and enadoline-induced reinstatement of cocaine seeking was antagonized by naltrexone. The additional finding that the priming effects of spiradoline and enadoline were not significantly attenuated by nor-BNI at doses previously shown to be active in squirrel monkeys (Carey and Bergman, 2001) raises the possibility that κ agonist-induced reinstatement of cocaine seeking was mediated via a non-κ opioid mechanism or a subpopulation of nor-BNI-insensitive κ opioid receptors (see below). Finally, the CRF1 receptor antagonist CP 154,526 and the α2-adrenoceptor agonist clonidine significantly attenuated spiradoline-induced reinstatement, suggesting a possible role for putative stress mechanisms.

Reinstatement of cocaine seeking by priming with spiradoline in the presence (•) and absence (▵) of response-contingent presentations of the cocaine-paired stimulus. Data are means ± S.E.M. (n = 5). *, significant difference from the corresponding value in the absence of stimulus presentations.
The differential antagonism of spiradoline and enadoline by naltrexone and nor-BNI in our study parallels the results of previous studies demonstrating differences in the abilities of naltrexone and nor-BNI to block the effects of enadoline in rhesus and squirrel monkeys (e.g., Butelman et al., 1998; Powell and Holtzman, 1999). Various studies have provided evidence for the existence of multiple κ receptor subtypes (Nock et al., 1988; Clark et al., 1989). Spiradoline and enadoline are members of the arylacetamide class of κ agonists that have been proposed as κ1 receptor agonists (Vonvoigtlander and Lewis, 1988). Another chemical class of κ agonists, the benzomorphans typified by bremazocine, exhibits putative selectivity at the κ2 receptor subtype (Zukin et al., 1988). Previous studies suggested that nor-BNI may be more effective in antagonizing the effects of benzomorphans than of the arylacetamides (Butelman et al., 1998; Powell and Holtzman, 1999; but see Carey and Bergman, 2001). In the study by Powell and Holtzman (1999), for example, the response rate-decreasing effects of enadoline were significantly attenuated by naltrexone but not by nor-BNI, whereas both antagonists blocked the effects of bremazocine. Similarly, nor-BNI did not antagonize the antinociceptive effects of enadoline in rhesus monkeys (Butelman et al., 1998), although it did attenuate the discriminative stimulus effects of enadoline in squirrel monkeys (Carey and Bergman, 2001). Consistent with the findings of Butelman et al. (1998) and Powell and Holtzman (1999), our data suggest that the effects of spiradoline and enadoline may be mediated by a subpopulation of κ receptors that is largely insensitive to antagonism by nor-BNI.
Although it is tempting to conclude that the effects of the κ agonists in our study reflect primarily κ1 receptor mechanisms, this simple conclusion should be viewed cautiously, as nor-BNI shows at least modest selectivity at κ1 compared with κ2 receptors in rhesus monkey brain membranes (Butelman et al., 1998). Furthermore, the finding by Carey and Bergman (2001) that nor-BNI attenuated the discriminative stimulus effects of both enadoline and bremazocine suggests that the behavioral effects of enadoline may not be mediated by a single subtype of κ opioid receptors.
Drug seeking after priming injections of spiradoline was significantly attenuated by the CRF1 receptor antagonist CP 154,526, suggesting a potential interaction between κ opioids and stress-related mechanisms. Dynorphin, which plays a role in regulating the stress response, is colocalized with CRF in the paraventricular nucleus of the hypothalamus (Roth et al., 1983) and has been found to modulate the effects of CRF on autonomic function (Overton and Fisher, 1989). κ Agonists also increase CRF release (Buckingham and Cooper, 1986) and stimulate the hypothalamic-pituitary-adrenal axis (Ur et al., 1997; Laorden et al., 2000). In light of these findings, it seems possible that priming with spiradoline in our study led to downstream activation of brain CRF1 systems involved in the regulation of stress. Consistent with this interpretation, CRF receptor antagonists have been shown to attenuate re-instatement of drug seeking induced by footshock stress in rats (Shaham et al., 1998), and CRF injected directly into the bed nucleus of the stria terminalis has been shown to induce reinstatement of cocaine seeking (Erb and Stewart, 1999). It is unlikely that the ability of CP 154,526 to attenuate spiradoline-induced reinstatement of drug seeking in our study was due to a nonspecific suppression of response rate as CP 154,526, albeit at a lower dose, did not significantly affect cocaine-induced reinstatement of drug seeking in a previous study using similar procedures in squirrel monkeys (Lee et al., 2003).
The α2-adrenoceptor agonist clonidine also attenuated the reinstatement of cocaine seeking induced by spiradoline, suggesting an interaction between the κ opioid and noradrenergic systems. κ Opioids have been shown to enhance NE turnover in the paraventricular nucleus of the hypothalamus (Laorden et al., 2000), increase NE metabolism in the brain stem and cortex (Kunihara et al., 1992), and stimulate NE release in the hippocampus (Jackisch et al., 1986). In addition, the NE transport inhibitor, imipramine, enhances spiradoline-induced analgesia (Kunihara et al., 1992). In vitro studies also have shown that clonidine can inhibit the effects of the κ agonist ethylketocyclazocine, whereas the α2-adrenoceptor antagonist yohimbine enhances the effects of ethylketocyclazocine (Limberger et al., 1986).
In vivo studies further suggest that noradrenergic mechanisms play an important role in stress-induced relapse (Erb et al., 2000; Lee et al., 2004). In squirrel monkeys, for example, yohimbine induces a clonidine-reversible reinstatement of cocaine-seeking behavior, as well as behavioral and physiological markers of stress (Lee et al., 2004). The interactions observed between the noradrenergic and κ opioid systems discussed above, along with the ability of clonidine to antagonize spiradoline-induced (present study), yohimbine-induced (Lee et al., 2004), and foot shock-induced (Erb et al., 2000) reinstatement of cocaine seeking, suggest that κ agonists may indirectly activate noradrenergic stress systems to induce the reinstatement of drug seeking.
The ability of clonidine to attenuate spiradoline-induced reinstatement of cocaine seeking should be interpreted cautiously, however, because full blockade was observed only with the higher dose of clonidine (0.3 mg/kg) in our study, possibly suggesting a nonspecific rate-suppressing effect. Although the effects of clonidine are similar both quantitatively and qualitatively to those observed in a previous study of yohimbine-induced reinstatement of cocaine seeking in squirrel monkeys (Lee et al., 2004), Platt et al. (2007) recently found that a dose of clonidine as low as 0.03 mg/kg significantly attenuated the reinstatement of cocaine seeking induced by the NE transport inhibitor nisoxetine. This finding notwithstanding, our results taken as a whole suggest that κ agonists may exert their response-reinstating effects through a complex interaction between CRF and noradrenergic stress mechanisms subsequent to binding at a nor-BNI-insensitive population of κ receptors. This conclusion is consistent with the view that reciprocal interactions among elements of the endogenous κ opioid CRF and noradrenergic systems constitute important neurobiological substrates for the behavioral expression of stress (cf. Koob, 1999; Valentino and Van Bockstaele, 2001).
In the majority of our experiments, response-contingent presentations of the cocaine-paired stimulus were available during reinstatement test sessions. The role of drug-associated stimuli in the reinstatement of drug seeking has been well established in rodents and monkeys (Shalev et al., 2002; Spealman et al., 2004). It is, however, unlikely that the reinstatement of cocaine seeking after priming with spiradoline and enadoline was due primarily to presentations of the cocaine-paired stimulus. Although presentations of a cocaine-paired stimulus alone can induce significant reinstatement of cocaine seeking under conditions similar to those used in the present study (Spealman et al., 2004), larger effects were seen after priming with spiradoline both in the presence and absence of the cocaine-paired stimulus.
Shelton and Beardsley (2005) and Feltenstein and See (2006) reported enhanced reinstatement of cocaine seeking in rats when footshock- or yohimbine-induced stress was accompanied by presentations of a cocaine-paired stimulus. Although priming with a maximally effective dose of yohimbine (Lee et al., 2004) or spiradoline (present study) may have overshadowed any synergistic effects of the stimulus in our experiments, priming with an intermediate doses of spiradoline (0.03 mg/kg) in the present study induced significantly greater reinstatement of drug seeking in the presence than in the absence of the cocaine-paired stimulus. These findings and the corresponding results in rats (Shelton and Beardsley, 2005; Feltenstein and See, 2006) may have implications for the development of relapse prevention strategies that address the potentially magnified effects of drug-paired stimuli under conditions of stress.
To summarize our principal findings, the present study demonstrates that spiradoline and enadoline can induce significant reinstatement of cocaine-seeking behavior in squirrel monkeys. Similar to previous findings with yohimbine (Lee et al., 2004), it appears that κ agonists may act as pharmacological stressors, as the CRF1 receptor antagonist CP 154,526 and the α2-adrenoceptor agonist clonidine blocked the response-reinstating effects of spiradoline nearly completely. Although stress is widely implicated as a risk factor in relapse, the mechanisms by which stress triggers relapse are largely unknown. Our data, along with previous reports demonstrating interactions between central κ opioid CRF and noradrenergic systems, suggest a complex interplay among these systems in the expression of stress-induced relapse.
Acknowledgments
We gratefully recognize the expert technical assistance of Kristen Bano, Annemarie Duggan, Lindsey Kettinger, Brett Lewis-Deweese, and Donna Reed.
Footnotes
-
This research was supported by Grants DA11054 and DA17700 from the National Institute on Drug Abuse and RR00168 from the National Center for Research Resources. G.R.V. was supported by National Research Service Award DA019286 from the National Institute on Drug Abuse.
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.107.125484.
-
ABBREVIATIONS: U50,488, trans-(–)-3,4-dichloro-N-methyl-N-[2-(1-pyrrolidinyl)cyclohexyl]benzeneacetamide; nor-BNI, nor-binaltorphimine; CRF, corticotropin-releasing factor; NE, norepinephrine; CP 154,526, butyl-ethyl-[2,5-dimethyl-7-(2,4,6-trimethylphenyl)-7H-pyrrolo [2,3-d]pyrimidin-4-yl]amine; FR, fixed ratio; FI, fixed interval; TO, time out; ANOVA, analysis of variance.
-
↵1 Current affiliation: Department of Psychology, Grand Valley State University, Allendale, Michigan.
- Received May 8, 2007.
- Accepted August 15, 2007.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}