


Abstract
Neuromedin U (NmU) is a neuropeptide showing high levels of structural conservation across different species. Since its discovery in 1985, NmU has been implicated in numerous physiological roles, including smooth muscle contraction, energy homeostasis, stress, intestinal ion transport, pronociception, and circadian rhythm. Two G-protein-coupled receptors have been identified for NmU and cloned from humans, rats, and mice. Recombinantly expressed NmU receptors couple to both Gαq/11 and Gαi G-proteins, and NmU binds essentially irreversibly, preventing signaling to repetitive applications of NmU. However, it is unclear whether these properties reflect those of endogenously expressed NmU receptors or how these properties influence the functional consequences of NmU receptor signaling. Here, we have explored the signaling by rat NmU receptors expressed endogenously in cultured rat colonic smooth muscle cells and explore the functional consequence of this signaling by investigating the NmU-mediated contraction of ex vivo rat colonic smooth muscle preparations. We demonstrate that endogenous rat NmU receptors couple to both Gαq/11 and Gαi G-proteins. Furthermore, we show complex patterns of Ca2+ signaling, including oscillations, and provide evidence of essentially irreversible binding of NmU to smooth muscle cells. Challenge of either circular or longitudinal rat isolated colonic smooth muscle preparations with NmU resulted in robust contractions. Stimulation was direct, and paradoxically, repetitive applications of NmU mediated repetitive contractions with no evidence of desensitization, highlighting a major discrepancy in the behavior of NmU in single cells and in intact tissues. The reason for this discrepancy is presently unknown.
The neuropeptide, neuromedin U (NmU), was first isolated from porcine spinal cord based in its ability to contract strips of rat uterus (hence the suffix “U”) (Minamino et al., 1985). Subsequent characterization of NmU from human, rabbit, dog, frog, rat, and chicken has revealed a remarkable degree of structural homology, with identical C-terminal pentapeptides containing elements essential for biological activity (for review, see Brighton et al., 2004a). Physiologically, NmU has been implicated in smooth muscle contraction (Minamino et al., 1985; Maggi et al., 1990; Westfall et al., 2001; Prendergast et al., 2006; Dass et al., 2007); regulation of regional blood flow and blood pressure (Gardiner et al., 1990); the function and development of the pituitary-adrenal-cortical axis (Malendowicz et al., 1993); arterial pressure and heart rate (Westfall et al., 2001; Chu et al., 2002); stress (Hanada et al., 2001; Zeng et al., 2006); pronociception (Cao et al., 2003; Yu et al., 2003); food intake and body weight (Howard et al., 2000; Kojima et al., 2000; Nakazato et al., 2000; Ivanov et al., 2002; Wren et al., 2002; Jethwa et al., 2005); gross locomotor activity, body temperature, heat production, and oxygen consumption (Howard et al., 2000; Nakazato et al., 2000); cancer (Yamashita et al., 2002); and the circadian oscillator (Nakahara et al., 2004) (for review, see Brighton et al., 2004a).
The identification of a human (h) orphan G-protein-coupled receptor as a specific target for NmU [human neuromedin U-receptor 1 (hNmU-R1)] (Fujii et al., 2000; Hedrick et al., 2000; Howard et al., 2000; Kojima et al., 2000; Raddatz et al., 2000; Shan et al., 2000; Szekeres et al., 2000) and the subsequent identification of an additional receptor [human neuromedin U-receptor 2 (hNmU-R2)] (Hosoya et al., 2000; Howard et al., 2000; Raddatz et al., 2000; Shan et al., 2000) has enabled progress in understanding the cellular signaling mediated by NmU receptor activation. Receptors of both human and rodent origin mediate phosphoinositide hydrolysis and cellular Ca2+ signaling dependent upon activation of Gαq/11 (Fujii et al., 2000; Hedrick et al., 2000; Hosoya et al., 2000; Howard et al., 2000; Kojima et al., 2000; Raddatz et al., 2000; Shan et al., 2000; Szekeres et al., 2000; Brighton et al., 2004b). Both human receptors also activate extracellular signal-regulated protein kinase (Brighton et al., 2004b), and there is evidence for inhibition of adenylyl cyclase (Hosoya et al., 2000; Brighton et al., 2004b). Indeed, we have shown direct activation of Gαi (Brighton et al., 2004b). Intriguingly, both recombinantly expressed human receptors bind NmU in an essentially irreversible manner, internalizing the ligand rapidly after agonist addition. The consequence of this is that repetitive application of NmU, even when separated by extensive washing with physiological solutions, does not evoke repetitive Ca2+ signaling (Brighton et al., 2004b), and this property may have important implications for the physiological actions of NmU. Initial studies with smooth muscle cells isolated and cultured from rat stomach fundus suggested that NmU may also bind essentially irreversibly to endogenously expressed receptors (Brighton et al., 2004b). However, with our understanding of NmU-mediated signaling derived predominantly from recombinant systems and an understanding that the cellular background can influence receptor properties, we have now examined the signaling of NmU receptors expressed endogenously in rat colonic smooth muscle cells. In particular, this has allowed an assessment of the relationship between single-cell signaling and functional responses in intact, isolated colonic smooth muscle.
Materials and Methods
Materials
Medium 231 and smooth muscle growth supplement (SMGS) were supplied by Cascade Biologics (Nottingham, UK). All other cell culture media, supplements, and reagents were supplied by Invitrogen (Paisley, UK). Dithiothreitol, hyaluronidase (330 FALGPA units/mg), papain (22 FALGPA units/mg), and collagenase (type F) (2.2 FALGPA units/mg) were all obtained from Sigma-Aldrich Co. Ltd. (Poole, UK). The primary α-actin- and secondary fluorescein isothiocyanate (FITC)-tagged goat anti-mouse antibodies were supplied by Sigma-Aldrich Co. Ltd., and the fluorescence protecting mounting medium was supplied by Dako North America Inc. (Carpinteria, CA). Fluo-3-acetoxymethyl ester was supplied by TEF Labs (Austin, TX), and fluo-4-acetoxymethyl ester and Pluronic F-127 were from Invitrogen. Biocoat 384-well black-walled clear-bottomed microtiter plates were from BD Biosciences (Bedford, MA). Lipofectamine Plus was purchased from Invitrogen. Brilliant black was supplied by Molecular Devices (Wokingham, UK). Male Wistar rats were obtained from Charles River Laboratories (Margate, Kent, UK). Tetrodotoxin (TTX) was supplied by Tocris (Bristol, UK). Porcine (p) NmU-8 and rat (r) NmU-23 were obtained from Bachem Ltd. (St. Helens, UK), and hNmU-25 and Cy3B-pNmU-8 (see Materials and Methods) were made at GlaxoSmithKline (Harlow, UK). 125I-Labeled human neuromedin U-25 (2000 Ci/mmol) was from Amersham Biosciences (Amersham, UK). All other chemicals and reagents were supplied by either Sigma-Aldrich Co. Ltd. or Fisher Scientific (Loughborough, UK). Molecular biology reagents were from BD Biosciences Clontech (Palo Alto, CA).
Dissociation and Culture of Rat Colonic Smooth Muscle Cells
Adult male Wistar rats (<300 g) were handled in accordance with the UK Animals (Scientific Procedures) Act, 1986. After culling, a midline, frontal incision was made in the stomach, and a 1.5- to 2-cm portion of the distal colon was removed. Cells were then isolated using an adaptation of a previously described method (Dart and Standen, 1993). In brief, tissue was washed, diced in dissociation buffer (137 mM NaCl, 5.4 mM KCl, 10 mM HEPES, 1 mM MgCl2, 0.44 mM Na2HPO4, and 4.2 mM NaHCO3, pH 7.4), and subject to enzymatic digestion in dissociation buffer with papain (16.5 units/ml) and dithiothreitol (3.2 mM) (35°C, 30 min) followed by collagenase type F (1.65 units/ml) and hyaluronidase (165 units/ml) (35°C, 45–60 min). Tissue pieces were washed and dissociated by mechanical sheer through fire-polished glass Pasteur pipettes. Cells were collected by centrifugation (500g, 3 min, room temperature) and either resuspended in buffer for immediate experimentation [Krebs-Henseleit bicarbonate (KHB); 10 mM HEPES, 4.2 mM NaHCO3, 11.7 mM d-glucose, 1.18 mM MgSO4.7H2O, 1.18 mM KH2PO4, 4.69 mM KCl, 118 mM NaCl, 1.29 mM CaCl2.2H2O, 0.1% (w/v) bovine serum albumin, pH 7.4] or alternatively resuspended and cultured in Medium 231 supplemented with 5% SMGS, streptomycin (50 μg/ml), penicillin (50 IU/ml), and gentamycin (50 μg/ml) at 37°C/5% CO2. Cells for either immediate experimentation or culture were added to six-well plates containing uncoated, sterile 25-mm glass coverslips. Where cells were cultured, these were not passaged.
Culture of HEK 293 Cells and Human Umbilical Vein Endothelial Cells
HEK 293 Cells. HEK 293 cells were cultured exactly as described previously (Brighton et al., 2004b). The generation and characterization of stable cell lines expressing hNmU-R1 or hNmU-R2 has also been reported previously (Brighton et al., 2004b).
Human Umbilical Vein Endothelial Cells. These cells were generously provided by the Department of Biochemistry, University of Leicester and were routinely cultured in Medium 199 with Glutamax supplemented with 20% fetal calf serum, streptomycin (50 μg/ml), penicillin (50 IU/ml), heparin (2.4 IU/ml), and Endothelial Cell Growth Supplement (50 μg/ml) in 75-cm2 flasks at 37°C in a 95% air/5% CO2 environment.
Staining of α-Actin
Immunocytochemistry. Rat colonic smooth muscle cells were cultured on glass coverslips for up to 15 days as described above. Cells were washed three times with phosphate-buffered saline (PBS) (without Mg2+ or Ca2+), permeabilized, and fixed by incubation with 100% methanol (10 min, –20°C). Cells were then washed a further three times as above. The α-actin primary antibody was diluted 1:500 (42 μg protein/ml) in PBS containing 10% goat serum (v/v) and incubated with cells overnight at 4°C. Cells were then washed and incubated with FITC-tagged goat anti-mouse antibody [diluted 1:200 (23 μg protein/ml) in PBS/10% goat serum] for 2 h at room temperature in the dark before being washed and fixed onto glass slides with a fluorescence-protecting mounting medium. Slides were left to dry before being sealed with clear nail varnish and stored at 4°C until imaging.
Imaging. Slides were mounted onto the stage of an UltraVIEW confocal microscope with a 40× oil-immersion objective lens and excited at 488 nm using a Kr/Ar laser. Emitted light was collected above 510 nm, and images were captured using a charge-coupled device camera.
Human Umbilical Vein Endothelial Cells. At the time of assay (passage 10), human umbilical vein endothelial cells (HUVECs) were cultured onto 1% gelatin-coated 25-mm glass coverslips and incubated overnight. HUVECs were stained with α-actin and imaged by confocal microscopy exactly as described above.
HEK 293 Cells. HEK 293 cells were cultured onto poly-d-lysinecoated 25-mm coverslips and stained exactly as described above.
Generation of cDNA Encoding Rat NmU Receptors and Transient Expression in HEK 293 Cells
cDNA encoding either rNmU-R1 or rNmU-R2 was subcloned into EcoRI and EcoRI/HindIII restriction sites, respectively, of the mammalian expression vector pCDN (Aiyar et al., 1994). Plasmids were purified using BD Biosciences Clontech MaxiPrep kits according to the manufacturer's instructions, adjusted to 1 μg/μl, and transfected into HEK 293 cells for transient expression of receptors using Lipofectamine Plus according to the manufacturer's instructions using 1 μg of DNA for each centimeter squared of cell culture surface area.
Measurement of Intracellular Ca2+ Signaling
Single-Cell Ca2+Imaging in Rat Colonic Smooth Muscle Cells. Single-cell Ca2+ confocal imaging of cultured rat colonic smooth muscle cells was performed using a PerkinElmer UltraVIEW confocal microscope (PerkinElmer Life and Analytical Sciences, Beaconsfield, Bucks, UK) and was based on a protocol previously used for HEK 293 cells (Brighton et al., 2004b). In brief, rat colonic smooth muscle cells, either acutely isolated or cultured for 7 to 9 days, were loaded with 5 μM fluo-3-acetoxymethyl ester and 0.044% (w/v) Pluronic F-127 for 30 min at room temperature diluted in KHB. These parameters gave an even cytosolic loading of smooth muscle cells suitable for single-cell Ca2+ imaging (data not shown). For imaging, coverslips were mounted onto the stage of an Olympus IX50 inverted microscope (Olympus, Tokyo, Japan) and maintained at 37°C using a Peltier heated coverslip holder. The field of cells was excited using a 488-nm laser line, and the emitted fluorescence was captured at wavelengths >505 nm, with images collected at 1-s intervals. Analysis was carried out using software supplied by the manufacturer (PerkinElmer Imaging Suite), with raw fluorescence data exported to Microsoft Excel (Microsoft, Redmond, WA) and expressed as F/Fo (fluorescence/basal fluorescence) for each cell. In experimental protocols requiring a wash step, the chamber was perfused with KHB at a rate of 5 ml/min at 37°C (unless otherwise stated), which rapidly replaced the chamber volume of 500 μl. Where required, experiments were performed in the nominal absence of Ca2+ by the exclusion of 1.29 mM CaCl2.2H2O from KHB.
Population Ca2+Signaling Using a Fluorometric Imaging Plate Reader (Molecular Devices, Sunnyvale, CA). HEK 293 cells were cultured in 175-cm2 flasks until 60 to 70% confluent. Vectors containing cDNA for either the rNmU-R1 or rNmU-R2 were transiently transfected into HEK 293 cells using Lipofectamine Plus transfection reagent as described above and cultured for 24 h. These were then seeded into black-walled, clear-bottomed, 384-well plates at a density of 50 000 cells/well and cultured for a further 24 h. Ca2+ measurements used a “no-wash” brilliant-black procedure. Cells were loaded with 2 μM fluo-4-acetoxymethyl ester diluted in Tyrode's buffer (145 mM NaCl, 2.5 mM KCl, 10 mM HEPES, 10 mM d-glucose, 1.2 mM MgCl2, 1.5 mM CaCl2.2H2O, 2.5 mM probenecid, and 250 μM brilliant black) for 1 h at 37 °C and 95% air/5% CO2 before assay. Analysis of changes in [Ca2+]i using a fluorescence imaging plate reader (FLIPR) was performed exactly as described previously (Brighton et al., 2004b).
Determination of G-Protein Activation
Rat colonic smooth muscle cells were dissociated and cultured in 175-cm2 flasks for 7 to 9 days. Cells were then harvested, membranes were prepared, and G-protein activation was determined as described previously (Akam et al., 2001; Brighton et al., 2004b). In brief, membranes were challenged for 10 min with 10 nM hNmU-25 in the presence of [35S]GTPγS followed by specific immunoprecipitation of Gαi(1–3) and Gαq/11 subunits and the subsequent determination of associated radioactivity.
Generation and Imaging of Fluorescently Labeled NmU
The methods used for the generation and imaging of Cy3B-pNmU-8 (an N-terminally conjugated fluorescent analog of pNmU-8) for use with cultured rat smooth muscle cells was identical to that described previously for HEK 293 cells with stable expression of either hNmU-R1 or hNmU-R2 (Brighton et al., 2004b).
NmU-Mediated Contraction of the Rat Isolated Distal Colon
Isolation of the Rat Distal Colon. Male Wistar rats (<300 g) were culled in accordance with Schedule 1 of the UK Animals (Scientific Procedures) Act, 1986. A midline incision was made, and 2 to 3 cm of the distal colon was removed and placed immediately into Krebs' buffer (121.5 mM NaCl, 2.5 mM CaCl2, 1.2 mM KH2PO4, 4.7 mM KCl, 1.2 mM MgSO4, 25 mM NaHCO3, 5.6 mM glucose, pH 7.4) pre-equilibrated with 5% CO2/95% O2 for 15 min at room temperature. The tissue was washed in Krebs' buffer and opened longitudinally by sharp dissection. Submucosa and mucosa were removed by gentle peeling and sharp dissection, and full-wall thickness preparations of rat distal colon were achieved by cutting tissue strips (approximately 15 × 6 mm) parallel to either circular muscle or longitudinal muscle as required.
Contractile Studies. Tissues were suspended between two parallel platinum ring recording electrodes in 5-ml tissue baths containing Krebs' buffer continually bubbled with 5% CO2/95% O2 at 37°C. Tissues were equilibrated for 60 min, during which time the bath solution was changed once every 15 min. Test agents (agonists, antagonists, and inhibitors; see below) were applied directly to the tissue bath, and tension was measured using a Pioden dynamometer UF1 force-displacement transducer (LCM Systems, Newport, Isle of Wight, UK). Initial analysis of contractile responses to a maximally effective concentration of carbachol (100 μM) demonstrated optimal contractions of both circular and longitudinal muscle when the tissue was suspended under 1g tension (data not shown), and all subsequent experiments were carried out under these conditions. Where indicated, experiments were performed in the presence of 1 μM atropine and/or following 5 μM TTX 15-min pretreatment.
Isolated tissue contraction data were acquired and analyzed using MP100 hardware and AcqKnowledge software (BIOPAC Systems, Inc., Goleta, CA). The effects of NmU on resting muscle tension were expressed as a percentage of the contractile response mediated by 100 μM carbachol.
Data Analysis
Concentration-response curves were fitted by a four-parameter logistic equation using GraphPad Prism (GraphPad Software Inc., San Diego, CA). All data are expressed as the mean ± S.E.M. Both mean and representative data were performed to an n of three or more as indicated. Where appropriate, individual n numbers relate to the number of animals used.
Results
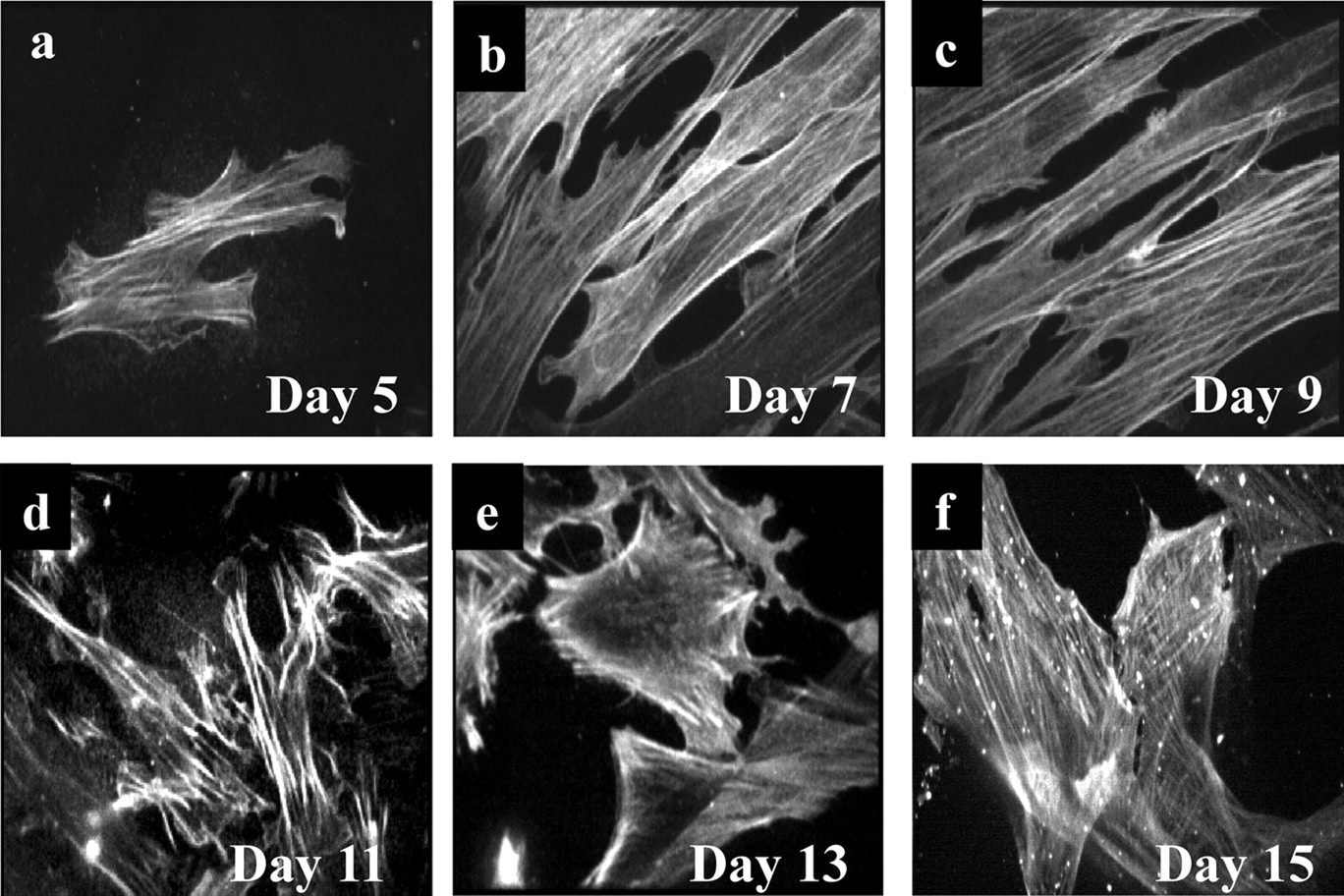
Characterization of Cultured Colonic Smooth Muscle Cells. The enzymatic digestion of rat distal colon resulted in suspensions of smooth muscle cells, which were long and thin, resembling smooth muscle cells (data not shown). Initial efforts to adhere these cells onto a substratum for the analysis of NmU-mediated single-cell Ca2+ imaging were relatively unsuccessful with exceptionally few cells adhering over a reasonable time frame to uncoated glass coverslips or coverslips coated with poly-d-lysine (50 μg/ml), fibronectin (10 μg/ml), gelatin (10% gelatin/90% PBS), conditioned media (from cultured smooth muscle cells, discussed below), and rat collagen (50 μg/ml) (data not shown). Of the very few cells that did adhere, addition of NmU evoked a rapid increase in [Ca2+]i, but as this occurred, the cells contracted vigorously and detached from the coverslip, thereby preventing further imaging and analysis. To overcome this difficulty, cells were cultured in Medium 231 supplemented with SMGS and antibiotics for up to 15 days. Adherent cells were apparent by days 3 to 5 and began to both proliferate and develop processes (Fig. 1a). By days 7 (Fig. 1b) and 9 (Fig. 1c), cells were approximately 50 to 70% confluent, had elongate morphology, and grew parallel to each other. After 11 days in culture (Fig. 1d), cells began to lose their elongated appearance and became more triangular in shape. By days 13 to 15 (Fig. 1, e and f), cells were 90 to 100% confluent and predominantly triangular in shape.
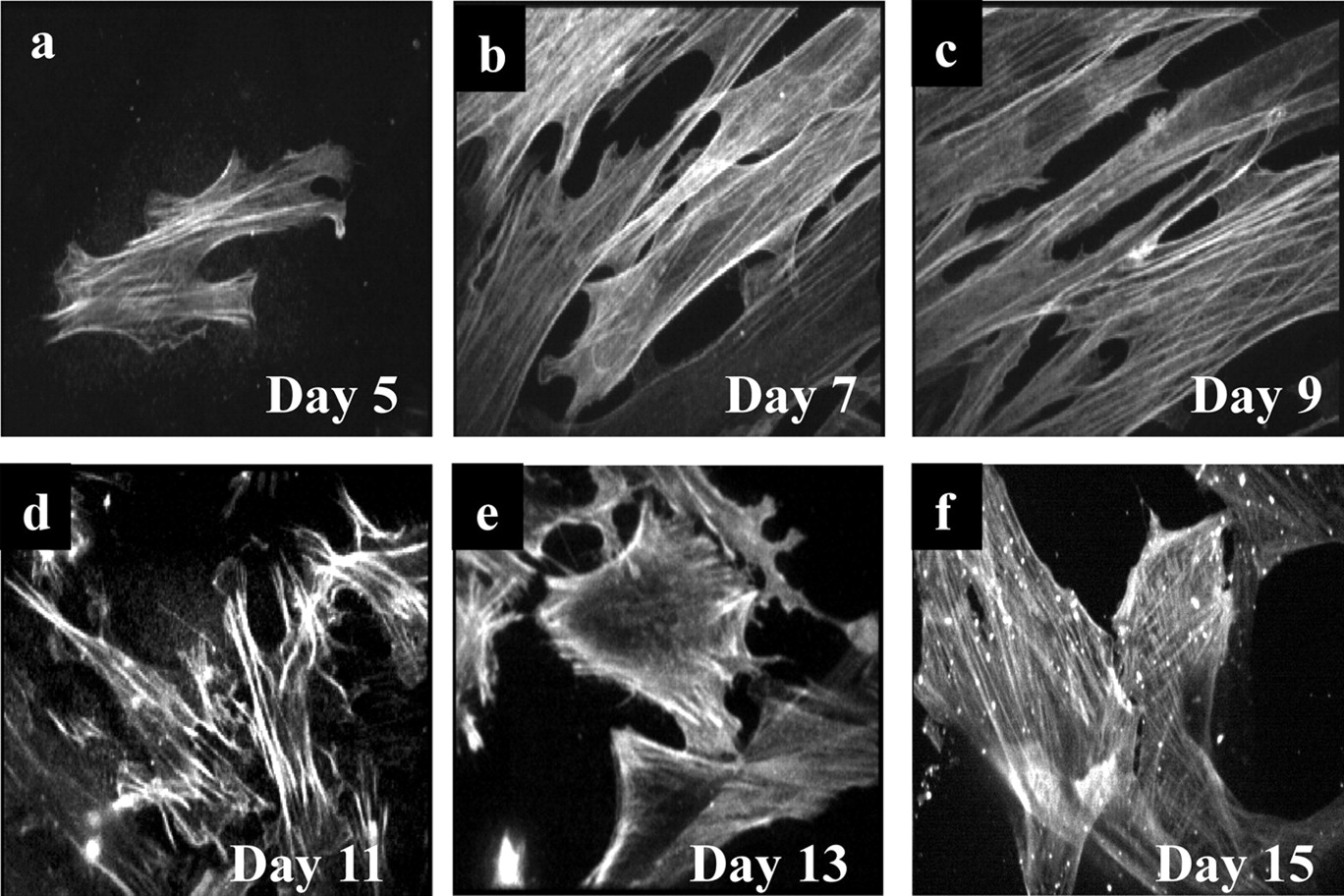
Immunocytochemical staining of α-actin, a smooth muscle cell phenotype marker (Skalli et al., 1988), throughout the 15-day culture period revealed strong staining from day 5 in culture (Fig. 2a). Staining was evident as fluorescent strands or filaments within the cytoplasmic regions of the cells, which is characteristic of smooth muscle cells (Skalli et al., 1988). This pattern of staining was most pronounced on days 7 to 11 (Fig. 2, b–d). Although still evident at days 13 to 15 of culture, α-actin staining had lost most of its filamentous characteristic (Fig. 2, e and f). Omission of the α-actin primary antibody when processing cells at 9 days resulted in no visible staining, and neither HEK 293 cells nor cultured HUVECs showed α-actin staining (data not shown). Smooth muscle cells cultured for between 7 and 9 days were used in subsequent experiments.
G-Protein Coupling of Endogenously Expressed Rat NmU Receptors. In membrane preparations from cultured (7–9 days) rat colonic smooth muscle cells, binding of [35S]GTPγS to immunoprecipitated Gαq/11 (Fig. 3a) or Gαi(1–3) (Fig. 3b) increased by ∼4- and ∼3-fold over basal, respectively, upon activation with 10 nM hNmU-25. Nonspecific binding using 10 μM GTPγS was ∼40 to 50% of basal (unstimulated) [35S]GTPγS binding (Fig. 3).
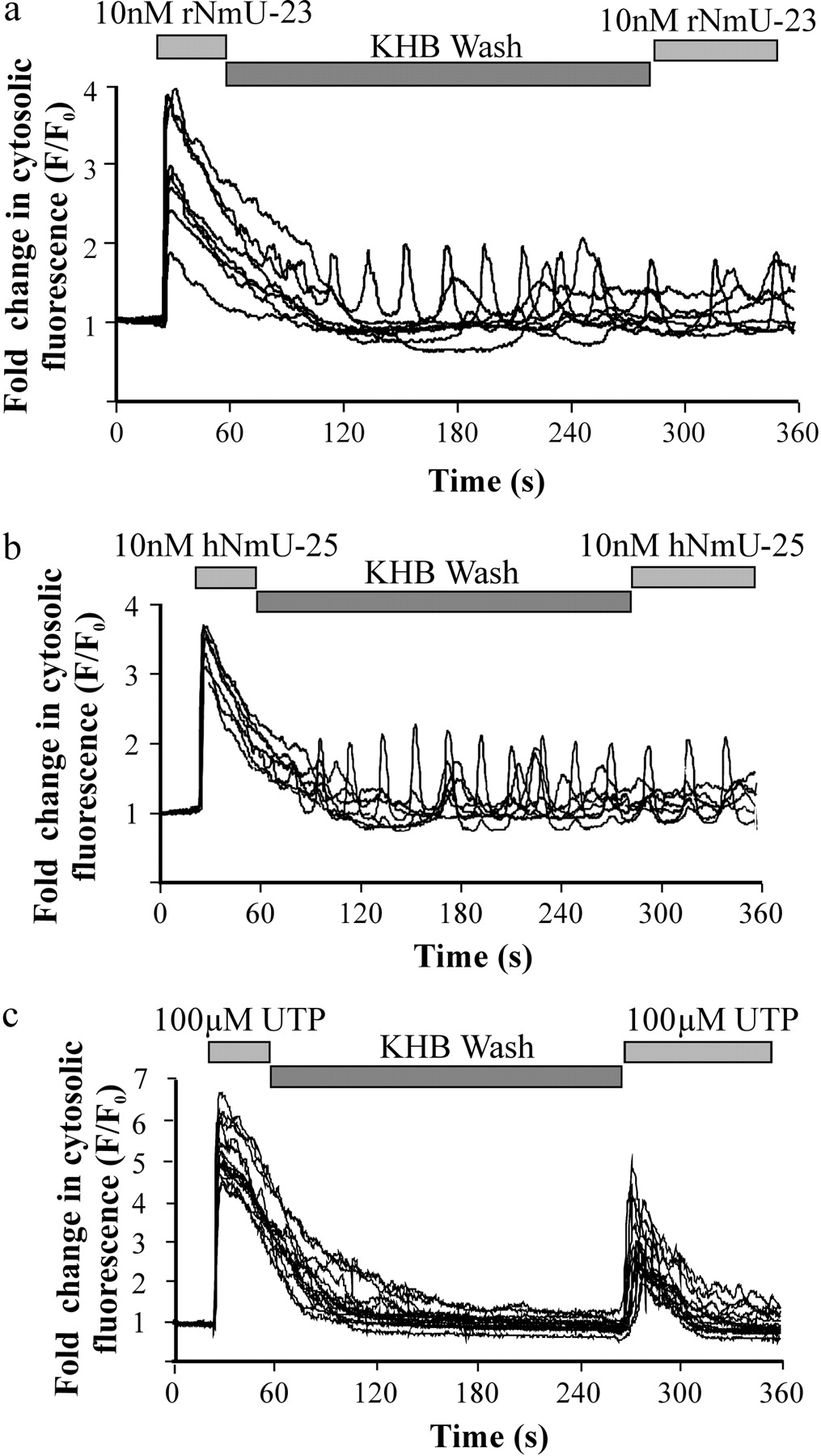
NmU-Mediated Signaling in Cultured Rat Colonic Smooth Muscle Cells: Ca2+Responses. Challenge of cells with either 10 nM rNmU-23 (Fig. 4a) or hNmU-25 (Fig. 4b) resulted in robust (∼3–6-fold over basal) and rapid (<5s) elevations of [Ca2+]i. The rapid initial elevations in [Ca2+]i were followed by a sustained but lower (1–2-fold over basal) signal where oscillations were seen in approximately 20 to 30% of cells. Oscillatory patterns were seen with hNmU-25 at both 100 nM and 1 μM (data not shown). The magnitude of the initial response to either rNmU-23 or hNmU-25 was unaffected by the removal of extracellular Ca2+, whereas the sustained signal was abolished (data not shown). In addition, pretreatment of cells for 10 min with 1 μM thapsigargin abolished all NmU-mediated responses (data not shown). The pEC50 values for the peak Ca2+ responses were 8.94 ± 0.42 and 8.64 ± 0.58 for hNmU-25 and rNmU-23, respectively (Fig. 4c).

Phenotypic characterization of cultured rat colonic smooth muscle cells. Smooth muscle cells were dissociated from the distal colon of male Wistar rats by enzymatic digestion of the extracellular matrix. These were cultured directly onto 25-mm glass coverslips for 5 (a), 7 (b), 9 (c), 11 (d), 13 (e), or 15 (f) days. Coverslips were mounted onto the stage of a PerkinElmer confocal microscope with a 40× oil immersion objective lens, and phase/light images were captured. Each individual image was chosen at random and is representative of three images from each of three separate cultures.
Immunocytochemical staining of α-actin in cultured rat colonic smooth muscle cells. Smooth muscle cells were prepared and cultured exactly as described in Fig. 1 for 5 (a), 7 (b), 9 (c), 11 (d), 13 (e), or 15 (f) days. Cells were permeabilized, fixed, and probed using an α-actin primary antibody and FITC-labeled secondary antibody. Cells were viewed under a 40× oil immersion objective lens using a PerkinElmer confocal microscope. Cells were excited at 488 nm and emitted light collected above 510 nm using a charge-coupled device camera. Images were chosen at random from nine images of cells from three separate rats.

G-protein coupling of endogenously expressed rat NmU receptors. Membrane preparations from cultured rat colonic smooth muscle cells were incubated in the presence of [35S]GTPγS and where applicable stimulated with 10 nM hNmU-25 for 10 min. Immunoprecipitation was then carried out using specific antibodies against either Gαq/11 (a) or Gαi(1–3) (b) subunits and associated [35S] determined. Nonspecific binding (NSB) was determined using 10 μM GTPγS. All data are mean + S.E.M., n = 3.

Single-cell imaging of NmU-mediated Ca2+ signaling in cultured rat colonic smooth muscle cells. Smooth muscle cells were prepared as described in Fig. 1 and cultured on 25-mm glass coverslips for 7 to 9 days. Cells were loaded with fluo-3, and agonist-mediated changes in cytosolic fluorescence were determined by confocal microscopy as an index of [Ca2+]i. Cells were challenged with either 10 nM rNmU-23 (a) or 10 nM hNmU-25 (b) as indicated. Shown are the changes in cytosolic fluorescence of six to 10 individual cells in the field of view. Images A to C were taken at the time points indicated. Data are representative of three experiments. c, concentration-response relationship for maximal NmU-mediated changes in [Ca2+]i. Each data point was obtained using maximal changes in cytosolic fluorescence averaged from 10 individual cells chosen at random in the field of view. Calculated pEC50 values are 8.94 ± 0.42 and 8.64 ± 0.58 for hNmU-25 and hNmU-23, respectively. Data are mean ± S.E.M., n = 3.
Lack of Repetitive NmU-Mediated Ca2+Signaling in Cultured Rat Colonic Smooth Muscle Cells. Following challenge of cells with either 10 nM rNmU-23 or hNmU-25, the sustained [Ca2+]i elevation or oscillatory behavior was unaffected by washing with KHB for 3 min. Furthermore, after a KHB perfusion for 3 min, reapplication of the same analog of NmU (10 nM) did not influence the [Ca2+]i (Fig. 5, a and b). Extending the wash with KHB to 12 min had no impact on the inability of the second addition of NmU to alter [Ca2+]i (data not shown). Challenge of naive cells with either 10 nM rNmU-23 or hNmU-25 also prevented the alternate ligand eliciting a [Ca2+]i response on the same cells after perfusion with agonist-free KHB for 2 min (data not shown).
Single-cell Ca2+ signaling was also assessed by confocal imaging of fluo-3-loaded HEK 293 cells transiently expressing either rNmU-R1 or rNmU-R2. Here, application of 10 nM hNmU-25 or rNmU-23 to cells expressing either rNmU-R1 or rNmU-R2 evoked a peak and plateau of elevated [Ca2+]i in approximately 30% of cells, consistent with typical levels of transfection efficiency achieved in these cells. Following washing of the cells by perfusion, either the same or the alternate ligand was unable to evoke a Ca2+ response (data not shown). Identical data were obtained in HEK 293 cells with stable expression of either hNmU-R1 or hNmU-R2, with the exception that all cells responded to the initial challenge with NmU (data not shown), given these are clonal cell lines.
Ca2+ signaling in cultured smooth muscle cells was also evoked by the application of 100 μM UTP to activate endogenous P2Y nucleotide receptors. UTP rapidly (<5 s) increased [Ca2+]i to between 4- and 10-fold over basal (Fig. 5c). Washing the cells by perfusion of the chamber with agonist-free KHB for >3 min after application of 100 μM UTP allowed a reapplication of 100 μM UTP to evoke a second Ca2+ response, albeit 30 to 50% of the magnitude of the initial response (Fig. 5c).
Binding of a Fluorescently Labeled NmU Analog to Cultured Rat Colonic Smooth Muscle Cells. The inability of a second addition of NmU to evoke a Ca2+ response after an initial challenge with NmU and a wash with buffer suggests either desensitization or an inability to remove receptor-bound NmU. As an approach to determine whether washing was sufficient to remove receptor-bound NmU, we used pNmU-8 with an N-terminally conjugated fluorophore, Cy3B (Cy3B-pNmU-8) (Brighton et al., 2004b). Based on Ca2+ responses in a FLIPR assay, Cy3B-pNmU-8 was equipotent with unlabeled pNmU-8, hNmU-25, and rNmU-23 against transiently expressed rNmU-R1 (Fig. 6a; Table 1) and rNmU-R2 (Fig. 6b; Table 1), indicating the validity of using Cy3B-pNmU-8 against endogenously expressed rat NmU receptors.
pEC50 values for the maximal elevation of [Ca2+]i by NmU analogs in HEK 293 cells transiently expressing either rNmU-R1 or rNmU-R2
Using a FLIPR, concentration-response curves were generated for the maximal (peak) changes in fluorescence (as an index of changes in [Ca2+]i) in response to the various NmU analogs in populations of HEK 293 cells that had been transiently transfected with either rNmU-R1 or rNmU-R2. Data were fit to sigmoidal curves by a four-parameter logistic equation using GraphPad Prism. Data are mean ± S.E.M., n = 3.
Addition of 10 nM Cy3B-pNmU-8 to cultured rat colonic smooth muscle cells at 12°C resulted in the immediate plasma membrane localization of fluorescence (Fig. 7a, i and ii). No fluorescence was observed following 10 nM Cy3B-pNmU-8 addition to either wild-type HEK 293 cells or HUVECs (data not shown). Furthermore, the fluorescence associated with the membranes of cultured smooth muscle cells was abolished by pretreatment with 1 μM unlabeled hNmU-25 (data not shown). These data are consistent with the binding of Cy3B-pNmU-8 to NmU receptors expressed recombinantly in HEK 293 cells (Brighton et al., 2004b) and indicate that Cy3B-pNmU-8 is able to bind to NmU receptors endogenously expressed in cultured colonic smooth muscle cells. The membrane-associated fluorescence could also not be removed by continual perfusion of KHB (12°C) for up to 12 min (Fig. 7b, i and ii) or by 1 μM hNmU-25 when it was applied 1 min following Cy3B-NmU-8 at 12°C (Fig. 7c, i and ii), highlighting the essentially irreversible nature of the binding.

Ca2+ signaling in response to repetitive stimulation of cultured rat colonic smooth muscle cells with either NmU or UTP. Smooth muscle cells were prepared as described in Fig. 1 and cultured on 25-mm glass coverslips for 7 to 9 days. Cells were loaded with fluo-3 and agonist-mediated changes in cytosolic fluorescence determined by confocal microscopy as an index of [Ca2+]i. Cells were challenged with 10 nM rNmU-23 (a), 10 nM hNmU-25 (b), or 100 μM UTP (c) as indicated. The cell chamber was then perfused with agonist-free buffer, and the cells were rechallenged. Changes in the cytosolic fluorescence of six to 10 cells within the field of view are shown. Data are representative of at least three experiments.

Concentration-response relationships for the elevation of [Ca2+]i by NmU analogs in HEK 293 cells transiently expressing either rNmU-R1 or rNmU-R2. HEK 293 cells in 384-well plates were transiently transfected with either rNmU-R1 (a) or rNmU-R2 (b). Cells were loaded with fluo-4, and changes in cytosolic fluorescence were determined by FLIPR as an index of [Ca2+]i. Shown are data for the maximal changes in fluorescence intensity immediately following agonist addition. Cells were challenged with hNmU-25 (▪), rNmU-23 (▴), pNmU-8 (▿), or Cy3B-pNmU-8 (⋄). The pEC50 values are given in Table 1. Data are mean ± S.E.M., n = 3.
At 37°C (in contrast to 12°C), addition of Cy3B-pNmU-8 again resulted in the immediate appearance of membrane fluorescence. However, after approximately 180 to 240 s, fluorescence associated with the plasma membrane had been largely lost, and fluorescence was almost exclusively punctate within the cell (Fig. 7d, i and ii).
NmU-Mediated Contraction of Rat Isolated Colon. In the absence of electrical stimulation, 1 μM (a maximally effective concentration; see below) of rNmU-23 mediated a contraction of either circular or longitudinal rat distal colon muscle strips maintained under 1g tension (Fig. 8, a and b). NmU induced a biphasic response characterized by an initial rapid contraction followed by a more sustained contraction on which there were repetitive twitches that did not reduce in magnitude (approximately 20–30% of initial contraction for circular muscle preparations and 40–60% for longitudinal preparations) and were maintained for at least 4 h. NmU-mediated contractions of either circular or longitudinal muscle strips were unaffected by the presence of either atropine (1 μM, 15-min pretreatment) or TTX (5 μM, 15-min pretreatment) (data not shown), suggesting a direct effect of NmU on smooth muscle. In contrast, atropine abolished contractions in response to carbachol (1–100 μM), and TTX abolished electrically stimulated, nerve-mediated responses (see Dass et al., 2003) (data not shown). Stimulation of muscle strips with 1 μM hNmU-25 evoked similar responses to those mediated by 1 μM rNmU-23 (see below).

Binding of fluorescently labeled pNmU-8 (Cy3B-pNmU-8) to cultured rat colonic smooth muscle cells. Phase image [a(i)] of cells immediately following addition of 10 nM Cy3B-pNmU-8 at 12°C and the corresponding fluorescence image [a(ii)] showing distinct plasma membrane localization. A fluorescent image of the same cell is shown immediately before [b(i)] and following [b(ii)] perfusion of the cell chamber with buffer (5 ml/min) for 12 min at 12°C, demonstrating an inability of this perfusion to remove membrane fluorescence. c, cells immediately after the addition of 10 nM Cy3B-pNmU-8 [C(i)] and 1 min after the addition of 1 μM hNmU-25 [c(ii)], demonstrating an inability to competitively displace bound Cy3B-pNmU-8. Phase image [d(i)] and fluorescent image 300 s following addition of 10 nM Cy3B-pNmU-8 at 37°C [d(ii)] showing internalization of the fluorescent ligand. All images are representative of at least three separate experiments.

NmU-mediated contraction of rat colon smooth muscle strips. Circular (a) or longitudinal (b) colonic smooth muscle tissue strips were prepared and suspended under 1g tension in organ baths. After equilibration, strips were challenged with 1 μM rNmU-23 as indicated by the bar. After 4 h, agonist-containing buffer was replaced with agonist-free buffer (first wash). After a further 15 min, this buffer was again replaced with agonist-free buffer (second wash); following another 15 min, the strip was rechallenged with 1 μM rNmU-23 as indicated. Data are representative of three experiments from three separate rats.
Repetitive NmU-Mediated Contractions of Rat Isolated Colon. Following the stimulation of either circular or longitudinal colonic smooth muscle preparations with rNmU-23 (1 μM) for up to 4 h, washing the strips with buffer followed by a second addition of rNmU-23 (1 μM) resulted in a contraction that was 90 to 100% of the response mediated by the first addition (Fig. 8, a and b). Similar results were obtained using hNmU-25 (1 μM) for both additions (data not shown). In these experiments, the protocol consisted of a 30-min interval between NmU additions with two complete replacements of buffer at 0 and 15 min. Using this interval and wash protocol, rNmU-23 (1 μM) repetitively (at least five times) contracted both circular and longitudinal smooth muscle preparations with no reductions in the amplitude of the response (Fig. 9, a and b). Similar results were obtained using hNmU-25 (1 μM) (data not shown). In these experiments, muscle strips were initially challenged with 100 μM carbachol to provide a reference for subsequent contractions.
Given this apparent lack of desensitization, noncumulative concentration response curves were generated for rNmU-23 and hNmU25 in both circular and longitudinal smooth muscle preparations (Fig. 10). NmU was added at different concentrations, in a random order, with a 30-min interval and two washes with buffer (as described above) between each addition. Therefore, full concentration-response curves were generated for individual pieces of tissue. Equivalent maximal contractions were achieved with approximately 1 μM rNmU-23 or hNmU-25 in both circular and longitudinal muscle strips. The pEC50 values obtained were 7.59 ± 0.17 and 7.64 ± 0.14 for rNmU-23 and 7.75 ± 0.08 and 7.53 ± 0.16 for hNmU-25, based on contractions of circular and longitudinal smooth muscle, respectively.
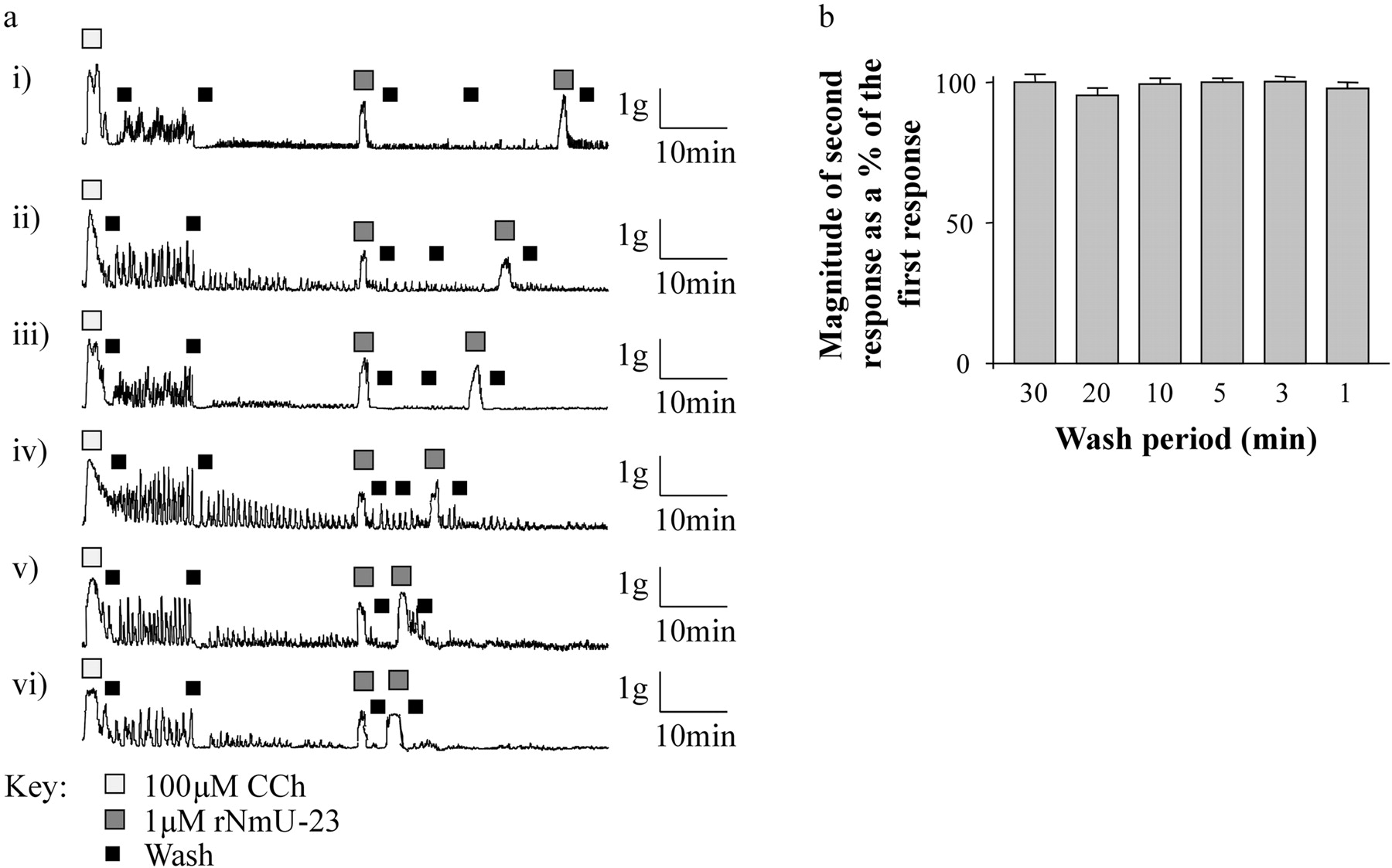
Using preparations of circular smooth muscle, the effect of reducing the wash period between consecutive applications of 1 μM rNmU-23 was explored. In these experiments, the wash period between consecutive applications was progressively reduced from 30 (Fig. 11a, i) to 1 (Fig. 11a, vi) min. The wash protocol was adapted to include at least one complete replacement of buffer at 0 min and, if possible (for wash periods of 5 min and more), a second replacement of buffer midway through the interval between successive applications of NmU. As the wash period was reduced, the magnitude of the response to the second challenge with NmU remained equivalent to the response of the first challenge (Fig. 11, a and b), even when just 3 (Fig. 11a, v) or 1 (Fig. 11a, vi) min, and one wash separated the two challenges.

Repetitive NmU-mediated contractions of rat colonic smooth muscle strips. Circular (a) or longitudinal (b) colonic smooth muscle tissue strips were prepared and suspended under 1g tension in organ baths. After equilibration, strips were challenged initially with 100 μM carbachol (CCh) and then washed twice with buffer over a 30-min period (see Fig. 8). Following this, strips were challenged repetitively with 1 μM rNmU-23 as indicated with two washes with agonist-free buffer during the 30-min intervals between challenges. Data are representative of three experiments from three separate rats.
Discussion
Despite a growing understanding of the physiological roles of NmU and the cloning of its two G-protein-coupled receptors, there is still a relative paucity of information concerning the cellular signaling mediated by endogenously expressed NmU receptors and how this relates to their biological roles. Here, we have examined this, using colonic smooth muscle as a model and demonstrate that although many aspects of the behavior of recombinant receptors are essentially paralleled by endogenously expressed receptors, there are clear differences between the behavior of NmU in isolated cells and intact tissue.
Although very few freshly isolated cells adhered sufficiently well to image, in the very small number that did, NmU evoked aCa2+ response, and the cells contracted, rounded up, and detached from the substratum. Culture of smooth muscle cells results in rapid phenotypic changes that result in a loss of contractility, most likely as a consequence of the loss of key contractile proteins including smooth muscle myosin light-chain kinase (Ma et al., 1998). Despite this, contractile stimuli can often evoke Ca2+ signaling in cultured smooth muscle cells. Here, we have exploited the lack of contractile responses to allow imaging of Ca2+ transients in cultured cells, particularly to allow repetitive challenges with agonist. Cells were used at a time in culture (7–9 days) at which staining for the smooth muscle marker α-actin was present.

Concentration-dependence of NmU-mediated contractions of rat colonic smooth muscle strips. Circular (•) or longitudinal (▪) colonic smooth muscle tissue strips were prepared and suspended under 1g tension in organ baths. After equilibration, strips were challenged initially with 100 μM CCh and then washed twice with buffer over a 30-min period (see Fig. 8). Concentration-response curves were then generated for each strip by the addition of either rNmU-23 (a) or hNmU-25 (b) at varying concentrations, noncumulatively and in a random order. Each successive agonist application was separated by two complete replacements of buffer in the bath over a 30-min period (see Fig. 9). The magnitude of each response was measured as a percentage of the initial carbachol-mediated response. The pEC50 values obtained were 7.59 ± 0.17 and 7.64 ± 0.14 for rNmU-23 and 7.75 ± 0.08 and 7.53 ± 0.16 for hNmU-25, based on contractions of circular and longitudinal smooth muscle, respectively. Data are mean ± S.E.M., n = 4.
In these cells, NmU mediated a robust transient elevation of [Ca2+]i, consistent with its contractile activity in smooth muscle. The initial Ca2+ response was often followed by asynchronous oscillatory Ca2+ signals, and although the role of such oscillations in smooth muscle cells is poorly understood (Savineau and Marthan, 2000), they do point to roles of NmU in aspects of cell function other than contraction. Here, we demonstrate directly that NmU receptors expressed endogenously in colonic smooth muscle cells couple to both Gαq and Gαi. This is consistent with the coupling of recombinantly expressed human NmU receptors, which also elevate [Ca2+]i and inhibit forskolin-stimulated cAMP accumulation with similarly high potency via Gαq and Gαi, respectively (Brighton et al., 2004b). Although we have not explored the signaling consequences of GαI coupling in smooth muscle cells, previous studies have suggested that Gαi can contribute significantly to agonist-mediated contractile events (Spitzbarth-Régrigny et al., 2000), and this would be consistent with offsetting cAMP-mediated relaxation. Although there are clearly exceptions, peripheral tissue generally expresses NmU-R1, whereas NmU-R2 is generally expressed within the central nervous system, suggesting that the NmU receptor involved here is likely to be NmU-R1 (see Brighton et al., 2004a). Although this remains to be established specifically for the rat colon, this is also consistent with studies on the gastrointestinal tract in NmU-R1 (Prendergast et al., 2006) and NmU-R2 (Dass et al., 2007) knockout mice.

The effects of reducing the interval between repetitive challenges with rNmU-23 on the contraction of rat colonic smooth muscle strips. a, circular colonic smooth muscle tissue strips were prepared and suspended under 1g tension in organ baths. After equilibration, strips were challenged initially with 100 μM CCh and then washed twice with agonist-free buffer over a 30-min period. Strips were then challenged with 1 μM rNmU-23 and washed with buffer for 30 (i), 20 (ii), 15 (iii), 10 (iv), 3 (v), or 1 (iv) min before rechallenge with 1 μM rNmU-23. For longer intervals, the buffer in the organ bath was replaced twice, whereas for intervals of 1 and 3 min, the buffer was replaced once as indicated. Data are representative of 3 experiments. b, magnitude of the second rNmU-23-mediated contraction was expressed as a percentage of the first. Data are mean + S.E.M., n = 3.
Challenge of cultured smooth muscle cells with UTP evoked aCa2+ response, most likely through activation of P2Y nucleotide receptors. Furthermore, after removal of UTP by perfusion of buffer, the readdition of UTP was able to evoke another Ca2+ response, albeit somewhat reduced, which is consistent with a partial desensitization of the receptors. Using this protocol, NmU was unable to evoke a Ca2+ response on a second challenge. Although it is possible this results from a full desensitization of the receptors, this would also be entirely consistent with an essentially irreversible binding of NmU, which we have shown with recombinantly expressed human NmU receptors. Indeed, here, using a fluorescently labeled NmU (Cy3B-pNmU-8), we show that once bound at the plasma membrane, NmU cannot be removed by washing the cells with a standard physiological salt solution. The rapid internalization of fluorescence in smooth muscle cells following addition of Cy3B-pNmU-8 is also consistent with high-affinity binding, which presumably requires an intracellular mechanism (low pH and/or proteolysis) to remove the ligand from its receptor. However, to date, little is known about the processing and potential recycling of NmU receptors.
Although the ability of NmU to mediate smooth muscle contraction is not in doubt, there appear to be both species and tissue differences, which include variation along the length of the gastrointestinal tract (Prendergast et al., 2006; Dass et al., 2007) and even differences between longitudinal and circular smooth muscle (Maggi et al., 1990). Of relevance to the present study are the observations that NmU mediates contraction of canine (Westfall et al., 2001) and human colon (Jones et al., 2006) but not colon from rat, guinea pig, or mouse (Benito-Orfila et al., 1991; Prendergast et al., 2006). The lack of a direct effect of NmU on contraction of mouse colon has been supported by a recent study, although it did potently enhance electrically stimulated, nerve-evoked contractions, possibly through prejunctional effects within the enteric ganglia (Dass et al., 2007). Such an effect is consistent with the regulation of peristalsis by NmU, demonstrated in mouse and guinea pig colon (Dass et al., 2007). The ability of NmU to evoke contraction of rat colon in the present study is somewhat in contrast to the findings in rodents. It is worthy to note that similar differences have been reported previously, with NmU either not contracting (Benito-Orfila et al., 1991) or contracting (Prendergast et al., 2006) rat ileum. Interestingly, in the latter study, NmU contracted longitudinal but not circular ileal muscle. The reasons for variation between the present and past studies are unclear.
In the present study, NmU-mediated contractions of circular and longitudinal smooth muscle preparations were equivalent, independent of either neuronal or muscarinic receptor activity, and were of only slightly less magnitude than those mediated by carbachol. Following an initial contraction evoked by a maximally effective concentration of NmU, the exchange of buffer in the organ bath enabled a second application to evoke a contraction of equivalent magnitude. This lack of functional desensitization was apparent irrespective of the interval between applications and at least with a 30-min interval allowed noncumulative concentration-response curves to be constructed with individual tissue strips. The potency of NmU (pEC50 values, 7.5–7.8) is consistent with its potency in other gastrointestinal smooth muscle preparations of the rat (pEC50 values, 6.4–8.4; Prendergast et al., 2006).
A clear paradox in the present study is the ability of NmU to mediate repetitive contractions of colonic smooth muscle strips but an inability to mediate repetitive Ca2+ transients in cultured colonic smooth muscle cells. It is worthy to note that these two preparations are clearly very different, which may complicate interpretation. Moreover, the cells and tissues have undergone different preparative procedures. In particular, the isolated cells have undergone both an enzymatic digestion and subsequent period of culture. Thus, it is possible that NmU receptors or any critical accessory proteins expressed in cultured smooth muscle cells differ in some way from those expressed in intact tissue and thereby account for differences in NmU receptor function. However, a lack of repetitive responses is consistent with the apparently irreversible binding of NmU to its receptors, whether endogenously expressed as in the present study or recombinantly expressed (Brighton et al., 2004b). The ability of NmU to mediate a contraction following an initial challenge with NmU and subsequent wash suggests additional factors regulate NmU signaling in intact tissue compared with isolated cells. It is unlikely that this is accounted for by differences in affinity of NmU for its receptors but could include mechanisms such as the recruitment of a pool of receptors initially inaccessible to ligand or the rapid internalization of receptor followed by ligand dissociation and recycling of receptors. It is also conceivable that a large receptor reserve, coupled with a lack of equilibrium binding and relatively low receptor occupancy after short periods of exposure to NmU, allow previously unoccupied receptors to mediate contraction in the experimental paradigm used. An alternative hypothesis is that the proteolytic degradation of NmU either in the immediate vicinity of the receptors or even when bound to receptors may play a crucial part in the removal of ligand, allowing subsequent additions to provoke responses. The in silico digestion of NmU (http://www.expasy.org/tools/peptidecutter/) indicates many cleavage sites within hNmU-25 and rNmU-23. Furthermore, rat uterine membranes efficiently degrade 125I-rNmU-23 (Nandha et al., 1993) and modification of the N-terminal of porcine NmU-8 to give aminopeptidase resistance increases its contractile activity on chickencrop smooth muscle (Sakura et al., 1995). The degradation of NmU may also account for its paradoxically relatively low potency on tissue contractile responses (pEC50 values of 6.4–8.4 in this study and Prendergast et al., 2006) compared with its affinity for NmU receptors (subnanomolar) (Fujii et al., 2000; Raddatz et al., 2000; Szekeres et al., 2000; Brighton et al., 2004b). Although the proteolysis of neuropeptides is a typical event in the regulation of their concentration and the termination of signaling, it remains to be established whether such a process is involved in regulating the cellular signaling and physiology of NmU.
Acknowledgments
We thank J. Scott and M. Ruediger (GlaxoSmithKline) for the generation and purification of Cy3B-NmU-8. We also thank both E. Appelbaum and E. Dul (Brad; GlaxoSmithKline, Upper Merion, Philadelphia, PA) for generating the stable cell lines and N. Elshourbagy and U. Shabon (Gene Cloning and Expression Proteomics, Glaxo-SmithKline) for cloning the receptors. We express our thanks to S. Ratcliffe (GlaxoSmithKline) for supplying hNmU-25 and F. McKay (GlaxoSmithKline) for help in using the FLIPR.
Footnotes
-
This study was supported by the Biotechnology and Biological Sciences Research Council (Grant 01/A4/C/07909) and by GlaxoSmithKline (Harlow, UK).
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.107.132803.
-
ABBREVIATIONS: NmU, neuromedin U; h, human; hNmU-R1, human neuromedin U receptor 1; hNmU-R2, human neuromedin U receptor 2; SMGS, smooth muscle growth supplement; FITC, fluorescein isothiocyanate; TTX, tetrodotoxin; pNmU-8, porcine NmU-8; hNmU-25, human neuromedin U-25; Cy3B-pNmU-8, porcine Cy3B-neuromedin U-8; KHB, Krebs-Henseleit bicarbonate; HEK, human embryonic kidney; PBS, phosphate-buffered saline; HUVEC, human umbilical vein endothelial cell; rNmU-R1, rat neuromedin U receptor 1; rNmU-R2, rat neuromedin U receptor 2; FLIPR, fluorescence imaging plate reader; rNmU-23, rat neuromedin U-23; CCh, carbachol; FALGPA, N-[3-(2-furyl)acryloyl)]-Leu-Gly-Pro-Ala.
- Received October 10, 2007.
- Accepted January 4, 2008.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}