


Abstract
There are species differences between human histamine H1 receptor (hH1R) and guinea pig (gp) histamine H1 receptor (gpH1R) for phenylhistamines and histaprodifens. Several studies showed participation of the second extracellular loop (E2-loop) in ligand binding for some G protein-coupled receptors (GPCRs). Because there are large species differences in the amino acid sequence between hH1R and gpH1R for the N terminus and E2-loop, we generated chimeric hH1Rs with gp E2-loop (hgpE2H1R) and gp N terminus and gp E2-loop (hgpNgpE2H1R). hH1R, gpH1R, and chimeras were expressed in Sf9 insect cells. [3H]Mepyramine binding assays and steady-state GTPase assays were performed. In the series hH1R > hgpE2H1R > hgpNgpE2H1R, we observed a significant decrease in potency of histamine 1 in the GTPase assay. For phenoprodifen 5 and the chiral phenoprodifens 6R and 6S, a significant decrease in affinity and potency was found in the series hH1R > hgpE2H1R > hgpNgpE2H1R. In addition, we constructed new active-state H1R models based on the crystal structure of the human β2-adrenergic receptor (hβ2AR). Compared with the H1R active-state models based on the crystal structure of bovine rhodopsin, the E2-loop differs in its contact to the ligand bound in the binding pocket. In the bovine rhodopsin-based model, the backbone carbonyl of Lys187 (gpH1R) interacts with large histaprodifens in the binding pocket, but in the hβ2AR-based model, Lys187 (gpH1R) is located distantly from the binding pocket. In conclusion, the differences in N terminus and E2-loop between hH1R and gpH1R exert an influence on affinity and/or potency for histamine and phenoprodifens 5, 6R, and 6S.
G protein-coupled receptors (GPCRs) represent the largest class of cell-surface receptors, which consist of seven transmembrane helices that are connected by three extracellular and three intracellular loops (Kristiansen, 2004). The histamine H1 receptor (H1R) is a biogenic amine receptor that belongs to class I of the GPCRs (Foord et al., 2005), and it interacts with Gq proteins to activate phospholipase C (Hill et al., 1997).
In histaprodifens (Fig. 1), identified as potent H1R agonists at the guinea pig (gp) ileum (Elz et al., 2000; Menghin et al., 2003), a 3,3-diphenylpropyl moiety is combined with a 2-substituted histamine. A pharmacological characterization of histaprodifens at the human H1R (hH1R) and guinea pig H1R (gpH1R) showed significant species differences (Seifert et al., 2003; Straßer et al., 2008). Several amino acids are involved in histamine binding: Asp3.32 (Ohta et al., 1994; Nonaka et al., 1998), Lys5.39 (Leurs et al., 1995; Bruysters et al., 2004; Jongejan and Leurs 2005), Thr5.42 and Asn5.46 (Leurs at al., 1994; Ohta et al., 1994), and Phe6.55 (Bruysters et al., 2004). The amino acid side chains Trp4.56, Lys5.39, Phe6.52, and Phe6.5 were found to interact with H1R antagonists (Wieland et al., 1999; Gillard et al., 2002).
Asn2.61 acts as a selectivity switch between hH1R and gpH1R for suprahistaprodifen and dimeric histaprodifen (Bruysters et al., 2005). However, pharmacological analysis of suprahistaprodifen and dimeric histaprodifen at the bovine H1R and the rat H1R (Straßer et al., 2008) showed that the amino acid in position 2.61 cannot be exclusively responsible for the observed species differences.
Molecular modeling studies of dimeric histaprodifen in the binding pocket of the gpH1R suggest that the second extracellular loop (E2-loop) is in close contact to dimeric histaprodifen and suprahistaprodifen and participates in binding of large ligands by forming hydrogen bonds (Straßer et al., 2008). Furthermore, the N terminus presumably interacts with the E2-loop. Comparing the amino acid sequences of the N terminus and E2-loop between hH1R and gpH1R, species differences of approximately 60% for the N terminus and of approximately 30% for the E2-loop are found (Fig. 2).

Structures of histamine, histaprodifens, and mepyramine. Histamine 1, histaprodifens 2 to 7, and mepyramine 8.
Several studies have analyzed the contribution of the E2-loop to agonist or antagonist binding or activation of GPCRs, and the same analysis was performed with the adenosine A1, A2a, A3 receptors (Olah et al., 1994; Kim et al., 1996), the dopamine D2 receptor (Shi and Javitch, 2004), the muscarinic acetylcholine M3 receptor (Scarselli et al., 2007), the α1-adrenergic receptor (Zhao et al., 1996), the α2A-adrenergic receptor (Laurila et al., 2007), and the histamine H2 receptor (Preuss et al., 2007). To study the influence of the species differences in N terminus and E2-loop between hH1R and gpH1R, we constructed chimeric hH1R with gp E2-loop (hgpE2H1R) and hH1R with gp N terminus and gp E2-loop (hgpNgpE2H1R) (Fig. 3A). The wild-type and chimeric H1Rs were coexpressed with the regulator of G protein signaling 4 (RGS4) in Sf9 insect cells. We characterized some histaprodifens (Fig. 1) in [3H]mepyramine competition binding assay and GTPase assay. In addition, active-state models based on the crystal structure of bovine rhodopsin and β2 receptor were constructed and compared to each other.
Materials and Methods
Materials. Phusion high-fidelity polymerase, all restriction enzymes, and T4 DNA ligase were obtained from New England Biolabs (Ipswich, MA). The anti-Flag IgG (M1 monoclonal antibody) was obtained from Sigma-Aldrich (St. Louis, MO), and the anti-RGS4 was from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA). [γ-32P]GTP was synthesized as described previously (Preuss et al., 2007), and [3H]mepyramine (30.0 Ci/mmol) was obtained from PerkinElmer Life and Analytical Sciences (Waltham, MA). A Rotiszint ecoplus (Roth, Karlsruhe, Germany) liquid scintillation cocktail was used. Histaprodifens were synthesized as described by Elz et al. (2000), Menghin et al. (2003), and Striegl (2006).
Construction of pGEMhgpE2H1R, pGEMhgpNgpE2H1R, pVLhgpE2H1R, and pVLhgpNgpE2H1R. First, a pGEM-3Z-SF-hgpNH1R plasmid was constructed. Therefore, pGEM-3Z-SF-gpH1R-plasmid and pGEM-3Z-SF-hH1R plasmid were double-digested with HindIII and PflMI, resulting in the fragments h-I, h-II, gp-I, and gp-II (Fig. 3B). The fragments h-I and gp-I were digested with PvuII, producing four fragments as follows: h-Ia, h-Ib, gp-Ia, and gp-Ib (Fig. 3B). Fragments h-Ia and gp-Ia encode parts of the pGEM-3Z-SF-plasmid, the N terminus, and the very beginning of transmembrane domain (TM) I. After separation and cleaning of the fragments, a triple ligation with gp-Ia, h-Ib, and h-II was performed. For generation of pGEM-3Z-SF-hgpE2H1R and pGEM-3Z-SF-hgpNgpE2H1R, overlap-extension polymerase chain reactions (PCR) (PCR Ia, PCR Ib, and PCR II) with pGEM-3Z-SF-hH1R and pGEM-3Z-SF-hgpNH1R plasmid, respectively, as template were performed. In PCR Ia, the DNA fragment with the signal peptide (S), the FLAG epitope (F), and the first part of the hgpNH1R up to transmembrane domain IV with guinea pig E2-loop was amplified. In PCR Ib, the DNA fragment encoding the guinea pig E2-loop, the second part of the hH1R, and the His6 tag (CACCATCATCACCATCAC) was generated. In PCR II, the products of PCR Ia and PCR Ib were annealed in the gp E2-loop-encoding region, resulting in PCR fragments that encode the SF, hgpNgpE2H1R, the His6 tag, the stop codon, and an XbaI site. For PCRs, the following primers targeting the E2-loop were used: GCTCACTCATTAGGCACC (forward, PCR Ia, PCR II); TTTCTCCCGGGGCTCACTAGTCGGGGCCATGAAGTGGTGCCAGCCTAGAATGGGAATAAC (reverse, PCR Ia); CACCACTTCATGGCCCCGACTAGTGAGCCCCGGGAGAAAAAGTGTGAGACAGACTTCTAT (forward, PCR Ib); and GGATCCTCTAGATTAGTGATGGTGATGATGGTG (reverse, PCR Ib, PCR II). The underlined code indicates a silent mutation for introduction of a unique diagnostic SpeI site. Italic letters designate the E2-loop. The resulting PCR fragment was double-digested with HindIII and XbaI and cloned into the pGEM-3Z plasmid using pGEM-3Z-SF gpH1R as a template. The sequences of hgpE2H1R and hgpNgpE2H1R, cloned into the pGEM-3Z-SF plasmid, were checked for their correctness by sequencing (Entelechon, Regensburg, Germany). pGEM-3Z-SF-hgpE2H1R and pGEM-3Z-SF-hgpNgpE2H1R were used as a template to clone hgpE2H1R and hgpNgpE2H1R into the pVL1392 baculovirus transfer vector using the restriction sites BssHII and XbaI. All wild-type and chimeric H1Rs were N-terminally tagged with the signal peptide and FLAG epitope ATGAAGACGATCATCGCCCTGAGCTACATCTTCTGCCTGGTATTCGCCGACTACAAGGACGATGATGACGCC and C-terminally tagged with the His6 tag CACCATCATCACCATCAC.

Alignment of the amino acid sequences of hH1R and gpH1R. The amino acid sequences are given in the one-letter code. Dots in the sequence of gpH1R indicate amino acids that are identical to hH1R. Hyphens indicate missing amino acids. Bold letters indicate glycosylation sites in the N terminus and E2-loop. Amino acids with gray shading are the most conserved amino acids (X.50, with X being the number of the TM domain), according to the numbering scheme in Ballesteros et al. (2001). Amino acids in white with black shading show the amino acids that are proposed to interact with the dimeric histaprodifen in the binding pocket of H1R, based on the simulation results of the new model. Bold and underlined amino acids interact with dimeric histaprodifen in the binding pocket of H1R only in the bovine rhodopsin-based model, but not in the hβ2AR-based model. The alignment was performed manually.

Scheme of the wild-type hH1R and gpH1R and the chimeras hgpE2H1R, hgpNgpE2H1R, and fragments used for construction of hgpE2H1R. A, hH1R, black; hgpE2H1R, the parts of hH1R are given in black, and the E2-loop according gpH1R is shown in gray; hgpE2gpNH1R, the parts of hH1R are given in black, and the N terminus and E2-loop according gpH1R are shown in gray; gpH1R, gray. B, the sequence of pGEM-3Z-SF-hH1R and pGEM-3Z-SF-gpH1R with PvuII, HindIII, and PflMI sites is given schematically. The fragments resulting from digestion are marked.
Preparation of Compound Stock Solutions. Chemical structures of the analyzed compounds are given in Fig. 1. Compounds 1 and 8 (10 mM each) were dissolved in double-distilled water. Compounds 2 to 6 were dissolved in a solvent mixture containing 30% (v/v) dimethyl sulfoxide (DMSO), 30% (v/v) Tris-HCl, pH 7.4 (10 mM), and 40% (v/v) double-distilled water, 5 mM each. Compound 7 was dissolved in 50% (v/v) DMSO and 50% (v/v) Tris/HCl, pH 7.4 (10 mM), at a concentration of 1 mM. This lead to a 1:10 dilution of DMSO in each assay tube. Reference binding and GTPase assays with three preparations of histamine dissolved in 1) double-distilled water, 2) 30% (v/v) DMSO, 30% (v/v) Tris-HCl, pH 7.4 (10 mM), 40% (v/v) double-distilled water [equivalent to a final DMSO concentration of 3% (v/v)], and 3) 50% (v/v) DMSO, 50% (v/v) Tris-HCl, pH7.4 (10 mM), equivalent to a final DMSO concentration of 5% (v/v), respectively, revealed only a decrease in the radioactivity counted, but to no shift in pKi and pEC50 values.
Miscellaneous. Cell culture, generation of recombinant baculoviruses, and membrane preparations were performed as described in Straßer et al. (2008). The determination of protein concentration, the SDS-polyacrylamide gel electrophoresis, and the immunoblot analysis were performed as described previously (Seifert et al., 2003; Straßer et al., 2008). The pharmacological assays, i.e., [3H]mepyramine saturation binding assay, [3H]mepyramine competition binding assay, and steady-state GTPase assay, were performed as described previously (Straßer et al., 2008). All assays for comparison of pharmacological data pharmacological data between the wild-type species isoforms of hH1R and gpH1R with the chimerics hgpE2H1R and hgpNgpE2H1R were carried out in parallel and under the same experimental conditions. All data were analyzed with Prism 4.02 (GraphPad Software Inc., San Diego, CA). pKi and pKB values were calculated according to the Cheng and Prusoff (1973) method. All data are the means ± S.E.M. of at least three independent experiments. To compare two pairs of data, the significance of the deviation of zero p was calculated using the t test.
Construction of Active H1R Models with Dimeric Histaprodifen in the Binding Pocket. Besides the well known crystal structure of bovine rhodopsin, in recent studies, the crystal structure of an additional GPCR, namely the human β2-adrenergic receptor (hβ2AR), was solved (Cherezov et al., 2007; Rasmussen et al., 2007; Rosenbaum et al., 2007). Therefore, two homology models of gpH1R, based on the two different crystal structures, were generated. The generation of the gpH1R model based on the crystal structure of bovine rhodopsin was described previously (Straßer and Wittmann, 2007). The gpH1R homology model based on the crystal structure of the hβ2AR (2RH1.pdb) (Cherezov et al., 2007; Rasmussen et al., 2007; Rosenbaum et al., 2007) was constructed in the same manner for all parts of the receptor, with the exception of the N terminus and E2-loop. Because of the lack of the conformation of the N terminus in the crystal structure of the hβ2AR, the N terminus was adopted from the crystal structure of bovine rhodopsin. The alignment of the E2-loop was performed manually and is given in Fig. 4. The modeling of the E2 regions His175 - Phe177, Ala179 - Pro184, and Glu190 - Tyr194 was performed using the loop-search module of the software SYBYL 7.0 (Tripos, St. Louis, MO). The helix structure of the E2-loop in the crystal structure of hβ2AR was not implemented in the homology model of gpH1R. In the crystal structure of the hβ2AR, two disulfide bridges, Cys191(E2) - Cys-184(E2) and Cys192(E2) - Cys106 (TM3, 3.25), are found. In the gpH1R model, only one disulfide bridge, Cys189(E2) - Cys109(TM3, 3.25), is modeled. The resulting homology model was energetically minimized and embedded in an environment consisting of 1-palmitoyl-2-oleoyl-phosphatidylcholine molecules, intracellular and extracellular water molecules, sodium, and chloride ions. Molecular dynamics (MD) simulations to generate an active gpH1R model based on the crystal structure of hβ2R were performed in an analogous way as described previously (Straßer and Wittmann, 2007). In the active model, a positively charged dimeric histaprodifen was docked manually and after MD simulations were performed, as described previously (Straßer et al., 2008). Inactive and active models of hH1R and hgpNgpE2H1R, based on the crystal structure of hβ2AR, were constructed in analogy to the corresponding gpH1R model.

Alignment of the E2-loop of hH1R and gpH1R to the amino acid sequence of bovine rhodopsin and human β2-adrenergic receptor. The cysteine forming a disulfide bridge to a cysteine residue in transmembrane domain III is shaded in gray. Hyphens indicate missing amino acids.
Based on the active-state models of the H1Rs, additional MD simulations were carried out. The equilibration phase and the productive phase were performed as described previously (Straßer et al., 2008). For all calculations, the software package GROMACS 3.3.1 (van der Spoel et al., 2004) was used in combination with the ffG53A6 force field (Oostenbrink et al., 2004). The force-field parameters for dimeric histaprodifen 7 were adopted from the ffG53A6 force field.
Results
Immunological Detection of the H1R Constructs. All H1R constructs were immunologically detected with the M1 antibody (Fig. 5). hH1R showed a strong band at ∼85 kDa. In contrast, gpH1R showed a strong band at 25 kDa, intermediate bands at ∼30 and ∼36 kDa, and faint bands at ∼50 and ∼100 kDa. For hgpE2H1R, a strong band was detected at ∼60 kDa and weak bands were detected in a range from 25 to 37 kDa. For hgpNgpE2H1R, a weak band was visible at ∼60 kDa, corresponding to hgpE2H1R. The ∼60-kDa band of the chimeric constructs hgpE2H1R and hgpNgpE2H1R corresponds very well to the theoretical mass of ∼56 kDa. The increase by 4 kDa relative to the theoretical mass is probably due to N-glycosylation. However, both chimeric constructs show different behavior with respect to the wild-type species isoforms, namely hH1R and gpH1R. These experimental results may be explained with different N-glycosylation states. At the N terminus, hH1R exhibits two and gpH1R exhibits one glycosylation site(s). Homology models of H1Rs suggest that there is one additional N-glycosylation site for hH1Rinthe E2-loop, but not for gpH1R. Thus, hH1R exhibits three (hgpE2H1R two, hgpNgpE2H1R one, and gpH1R one) glycosylation site(s).

Immunological detection of hH1R, hgpE2H1R, hgpNgpE2H1R, and gpH1R, expressed in Sf9 insect cell membranes. Sf9 cells, expressing one of the H1Rs and RGS4, were analyzed in an immunoblot as described under Materials and Methods. For immunological detection, the M1 antibody was used. Each line represents one of the four analyzed H1Rs. Numbers at the left indicate the apparent molecular masses of the proteins in kilodaltons.
Analysis of the H1R Wild-Type and Chimeric Species Isoforms in the [3H]Mepyramine Saturation Binding Assay. The KD and Bmax values, which were determined in the [3H]mepyramine saturation binding assay, are given in Table 1. There was no significant difference in the KD values of hH1R, hgpE2H1R, and hgpNgpE2H1R, which were 1.5 to 2-fold higher (p < 0.05) than at gpH1R. The Bmax value for hgpE2H1R was in the same range as the value for hH1R. The Bmax value for hgpNgpE2H1R was approximately 3-fold lower (p < 0.05) than that of hgpE2H1R, corresponding to the weaker immunoreactivity in the immunoblot (Fig. 5). The nonspecific binding of [3H]mepyramine in Sf9 cells expressing wild-type and chimeric species isoforms was maximally 30%.
[3H]Mepyramine saturation binding in Sf9 cell membranes expressing hH1R, hgpE2H1R, hgpNgpE2H1R, and gpH1R with RGS4 Sf9 cell membranes coexpressing one of the H1Rs and RGS4 were incubated with 0.2 to 20 nM [3H]MEP as described under Materials and Methods. Nonspecific binding was determined in the presence of 10 μM diphenhydramine and was subtracted from total [3H]MEP binding. The resulting binding data were analyzed by nonlinear regression and were best fitted to monophasic saturation curves. Data shown are the means ± S.E.M. of three independent membrane preparations, each one analyzed in triplicate.
Analysis of Histaprodifens at H1R Wild-Type and Chimeric Species Isoforms in the [3H]Mepyramine Competition Binding Assay. The affinities of compounds 1 to 7, determined in the competition binding assay, are given in Table 2. For compounds 1 to 4S and 7, no significant difference in pKi values was found between hH1R, hgpE2H1R, and hgpNgpE2H1R. However, for phenoprodifen 5 as well as for the chiral phenoprodifens 6R and 6S, a significant decrease in pKi values was found in the series hH1R > hgpE2H1R ≥ hgpNgpE2H1R (Fig. 6). For compound 5 at hH1R, the pKi value was significantly (p < 0.005) higher than at hgpE2H1R and hgpNgpE2H1R. At hH1R, the pKi value of 6R was significantly lower than that of 6S (p < 0.05).
Affinities of histamine and histaprodifens at hH1R, hgpE2H1R, hgpNgpE2H1R, and gpH1R coexpressed with RGS4 in Sf9 cell membranes in the equilibrium competition binding assay [3H]MEP competition binding in Sf9 membranes expressing hH1R, hgpE2H1R, hgpNgpE2H1R, and gpH1R in combination with RGS4 was determined in presence of 5 nM [3H]MEP as described under Materials and Methods. Data were analyzed by nonlinear regression and were best fit to one-site (monophasic) competition curves. The pKi values were calculated according to the Cheng and Prusoff (1973) method. Data shown are the means ± S.E.M. of at least three experiments, each one performed in duplicate with independent membrane preparations.
Constitutive Activity and Maximal Stimulation with Histamine of hH1R, hgpE2H1R, hgpNgpE2H1R, and gpH1R. The H1R couples to endogenous Gq proteins of Sf9 insect cells (Houston et al., 2002). Agonist activation of the Gq proteins is detected by an increase of high-affinity GTP hydrolysis in membranes expressing H1R and RGS proteins. The basal GTPase activity of the wild-type and chimeric H1R species ranged from 1.1 to 1.5 pmol/(mg*min) without significant differences (Table 3). However, the maximal stimulation with 100 μM histamine 1 relative to the basal activity (ΔHA) at hgpNgpE2H1R was significantly higher (p < 0.005) than at hH1R, hgpE2H1R, and gpH1R (Table 3). In addition, the ratio ΔHA/Bmax was significantly higher (p < 0.01) at hgpNgpE2H1R than at hH1R, hgpE2H1R, and gpH1R. Therefore, it can be concluded that the large stimulatory effect of histamine at hgpNgpE2H1R, despite the low expression level of hgpNgpE2H1R in Sf9 cell membranes, is a consequence of receptor conformation due to binding of the endogenous ligand histamine 1. The inverse agonist mepyramine 8 (Fitzsimons et al., 2004), which stabilizes the inactive state of the H1R and consequently reduces constitutive activity (Seifert and Wenzel-Seifert, 2002), showed only small inhibitory effects on basal GTP hydrolysis at all four H1Rs, indicating a low constitutive activity of the wild-type and chimeric H1Rs (Table 3). The pEC50 values of mepyramine 8 ranged from 7.75 to 8.96 (Table 3). There was no significant difference between the pEC50 values of mepyramine between hH1R and the chimeric H1Rs, but the pEC50 values at the chimeric H1Rs were significant lower (p < 0.005) than at gpH1R.
Analysis of the effects of histamine and mepyramine and determination of the constitutive activity of hH1R, hgpE2H1R, hgpNgpE2H1R, and gpH1R in the steady-state GTPase assay Sf9 cell membranes expressing hH1R, hgpE2H1R, hgpNgpE2H1R, or gpH1R in combination with RGS4 were used to study the constitutive activity. GTPase assays were performed as described under Materials and Methods. The concentration-response curves of the inverse agonist MEP (8) were determined in a concentration range from 0.1 to 10 μM. Data were analyzed by nonlinear regression and were best fit to sigmoidal concentration-response curves. The efficacy of histamine was set 1.00. The ΔHA value refers to the difference of maximal stimulation with 100 μM histamine relative to basal activity. The coefficient ΔHA/Bmax was calculated based on the data given for ΔHA in this table and Bmax values in Table 1. The ΔMEP value refers to the difference of basal activity relative to GTP hydrolysis in presence of 10 μM MEP. The relative effect of MEP in the last column is the percentage effect of MEP relative to HA. Data shown are means ± S.E.M. of three experiments, each one performed in duplicate.
Analysis of Histaprodifens at H1R Wild-Type and Chimeric Species Isoforms in the Steady-State GTPase Assay. The potencies and efficacies (Table 4), and the pKB values (Table 5), determined in the steady-state GTPase assay are given. A decrease in pEC50 values of histamine 1 in the series hH1R > hgpE2H1R > hgpNgpE2H1R was observed (Fig. 7). The potency of 1 at hgpE2H1R and hgpNgpE2H1R was significantly lower than hH1R(p < 0.05 for hgpE2H1R and p < 0.005 for hgpNgpE2H1R). For compounds 2 to 4S and 7, neither differences in potency nor efficacies were found between hH1R and both chimeras. For the phenoprodifen 5, a decrease in potency was found in the series hH1R > hgpE2H1R > hgpNgpE2H1R. The chiral phenoprodifens 6R and 6S act as antagonists at hgpE2H1R and hgpNgpE2H1R as well as at hH1R (Straßer et al., 2008). In accordance with the results in the competition binding (Fig. 6) assay, a decrease in pKB values was found for 6R as well as for 6S in the series hH1R > hgpE2H1R > hgpNgpE2H1R (Table 4).
Potencies and efficacies of histamine and histaprodifens at hH1R, hgpE2H1R, hgpNgpE2H1R, and gpH1R coexpressed with RGS4 in Sf9 cell membranes in the steady-state GTPase assay. All GTPase experiments were performed as described under Materials and Methods. Data were analyzed by nonlinear regression and were best fit to sigmoidal concentration-response curves. The efficacy (Emax) of histamine was set 1.00. The Emax values of all other compounds were referred to this value. Data shown are means ± S.E.M. of at least three experiments, each one performed in duplicate or triplicate. Membranes were used from independent membrane preparations. The effects of 4S, 6R, and 6S at hH1R, hgpE2H1R, and hgpNgpE2H1R were too small to determine pEC50 values. Therefore, Emax values were determined using ligand concentrations of 10 or 100 μM each.
Potencies of histaprodifens acting as antagonists at hH1R, hgpE2H1R, and hgpNgpE2H1R determined in the steady-state GTPase assay at Sf9 cell membranes expressing hH1R, hgpE2H1R, or hgpNgpE2H1R, and RGS4 The potencies of 4S, 6R, and 6S were determined in the steady-state GTPase assay as described under Materials and Methods. Each assay tube additionally contained 1 μM histamine. Data were analyzed by nonlinear regression and were best fit to sigmoidal concentration-response curves. The pKB values were calculated according to Cheng and Prusoff (1973) method. Data shown are means ± S.E.M. of at least three experiments, each one performed in duplicate or triplicate. Membranes were used from independent membrane preparations.
Binding Mode of Dimeric Histaprodifen in gpH1R Models Based on Two Different Crystal Structures. The binding mode of dimeric histaprodifen 7 docked into the binding pocket of the gpH1R active-state model, based on the crystal structure of bovine rhodopsin (Fig. 8A), was described in detail in Straßer et al. (2008). In comparison, the molecular dynamics simulations with dimeric histaprodifen 7 docked into the active-state model of the gpH1R, based on the crystal structure of hβ2AR (Fig. 8B), revealed some differences in the resulting binding mode. The largest difference between the two active gpH1R models were found in the conformation and flexibility (Fig. 9) of the E2-loop. In the bovine rhodopsin-based gpH1R model, the part of the E2-loop that shows species differences between hH1R and gpH1Risin close contact to the upper part of the binding pocket, near the transmembrane domain II. It is interesting to note that the backbone carbonyl of Lys187 (gpH1R) forms a hydrogen bond to an imidazole moiety of dimeric histaprodifen. Because of a shift of the E2-loop in the hβ2AR-based model, Lys187 does not interact with dimeric histaprodifen. The following amino acids were found to participate in the binding of 7 in the new model: Leu39 (TM1, 1.35), Leu43 (TM1, 1.39), Leu97 (TM2, 2.65), Trp112 (TM3, 3.28), Ile124 (TM3, 3.40), Trp429 (TM6, 6.48), and Trp456 (TM7, 7.40). The quaternary amine moiety in the center of 7 interacts electrostatically with the negatively charged Asp116 (TM3, 3.32) and Tyr459 (TM7, 7.43). One of the imidazole moieties forms stable hydrogen bonds with Ser120 (TM3, 3.36) and Tyr432 (TM6, 6.51). The second imidazole moiety forms stable hydrogen bonds to Glu190 (E2-loop) and Trp456 (TM7, 7.40) (Fig. 8). Both dipenyl propyl moieties of the dimeric histaprodifen are embedded in hydrophobic pockets.

Competition binding isotherms for 6R and 6S at hH1R and hgpNgpE2H1R. The competition binding experiments were performed using Sf9 cell membranes expressing hH1RorhgpNgpE2H1R and RGS4 in the presence of 5 nM [3H]mepyramine ([3H]MEP) as described under Materials and Methods. Data were analyzed by nonlinear regression and were best fit to one-site (monophasic) competition curves. The downward-facing triangle (▾) shows the data for hH1R, and the square (▪) shows the data for the chimerical hgpNgpE2H1R. Data shown are the means ± S.E.M. of at least three experiments, each one performed in duplicate. Membranes were used from independent membrane preparations.
The conserved disulfide bond between two cysteine side chains in TM3 and the E2-loop is responsible for a reduced flexibility in this region of the E2-loop (Fig. 9). However, although molecular dynamic simulations show a high flexibility of the nonrestricted parts of the E2-loop, a movement toward the binding pocket is notably observed. The interaction between the conserved Glu and the ligand is observed only in cases where the E2-loop is close to the binding pocket. This hydrogen bond stabilizes the conformation of the ligand in the binding pocket and reduces the flexibility of the ligand. The exchange of N terminus and E2-loop exerted an influence onto the hydrogen bond network between N terminus, E1-loop, and E2-loop with an effect on E2-loop conformation and flexibility (Fig. 9).
Discussion
GTPase Activation and Potency of Histamine at the Chimeric hgpNgpE2H1R Compared with hH1R. Our steady-state GTPase assay data show that the double-chimeric hgpNgpE2H1R has influence on the maximal Gq-protein stimulation with histamine and potency of histamine, compared with hH1R. Because the single-chimeric hgpE2H1R showed no influence on the maximal Gq-protein stimulation with histamine compared with hH1R, it can be concluded that the N terminus or the N terminus in combination with the E2-loop is responsible for this observation. Molecular dynamics simulations revealed that the exchange of N terminus and E2-loop has an influence on the hydrogen bond network between N terminus, E1-loop, and E2-loop with an effect on the conformation and flexibility of the E2-loop. Based on these data, it can be concluded that the interaction of the N terminus with the extracellular parts of the receptor induces subtle long-range conformational changes, resulting in a larger stimulatory effect of histamine and a decreased potency at hgpNgpE2H1R compared with hH1R.
Influence of the N Terminus and E2-Loop on Pharmacological Differences of Histaprodifens between hH1R and gpH1R. The experimental data show that the chimeric hgpE2H1R and hgpNgpE2H1R are well expressed in Sf9 cell membranes and show full functionality. However, the pharmacological studies specifically binding assays and steady-state GTPase assays indicate that the N terminus and the E2-loop are not responsible for the species differences between hH1R and gpH1R with regard to pKi, pEC50, and Emax values for compounds 2 to 4S and 7. Nevertheless, for the three phenoprodifens (5, 6R, and 6S) with a high structural similarity, a significant decrease in pKi and pEC50 values was found in the series hH1R > hgpE2H1R > hgpNgpE2H1R.
Our molecular dynamics simulations with the new gpH1R model, based on the crystal structure of hβ2AR, show that the backbone carbonyl of Lys187 (gpH1R) is far away from the binding pocket and does not interact with a large histaprodifen in the binding pocket, as suggested by the gpH1R model based on the crystal structure of bovine rhodopsin. In addition, the molecular dynamic simulations based on the hβ2AR model show that the highly conserved Glu181 (hH1R) and Glu190 (gpH1R), respectively, within the H1R family point toward the binding pocket with formation of a stable hydrogen bond to the ligand and point away from the binding pocket forming a stable hydrogen bond to Tyr96 (TM2, 2.42). The MD simulations have shown that the exchange of the N terminus and E2-loop exhibits an influence of the hydrogen bond network in the extracellular part of the receptor with an effect on E2-loop conformation and flexibility, which results in decreased pKi and pKB values for the phenoprodifens 5, 6R, and 6S at the chimeric H1Rs. Furthermore, we assume that 5, 6R, and 6S can bind in two different orientations into the binding pocket (Straßer et al., 2008). It is possible that a change in orientation of 5, 6R, and 6S in the series hH1R - hgpE2H1R - hgpNgpE2H1R can explain our experimental data. Because the largest differences between hH1R, the chimeric H1Rs, and gpH1R occur in the extracellular surface of the receptor, it is assumed that this possible change in orientation is determined kinetically during the early steps of the ligand binding process. To test this hypothesis, sophisticated kinetic binding studies have to be carried out. Furthermore, extensive molecular dynamics simulations with compounds 5, 6R, and 6S at wild-type and chimeric H1Rs are required to quantify the question of orientation.

Concentration-response curves for histamine 1 at hH1R, hgpE2H1R, and hgpNgpE2H1R in the steady-state GTPase assay. The experiments were performed using Sf9 cell membranes coexpressing hH1R, hgpE2H1R, or hgpNgpE2H1R and RGS4 as described under Materials and Methods. The downward-facing triangle (▾) shows the data for hH1R, and the square (▪) shows the data for the chimerical hgpNgpE2H1R. Data shown are the means ± S.E.M. of at least three experiments, each one performed in duplicate with independent membrane preparations.
Our new H1R models indicate that the region of the E2-loop with species differences in amino acid sequence is pointing away from the binding pocket. It is possible that highly conserved amino acids in the E2-loop of H1Rs are involved in ligand binding, e.g., Glu190 (E2-loop, gpH1R), which shows electrostatic interaction with dimeric histaprodifen in the old gpH1R model as well as in the new model. Additional experimental studies will be carried out to analyze the participation of Glu190 (E2-loop, gpH1R) in binding of large histaprodifens.
Role of the Extracellular Loop E2 in Ligand Binding and Activation of GPCRs. Compared with other GPCR regions, relatively little attention has been paid thus far to the contribution of the second extracellular loop E2 in ligand recognition and ligand binding in biogenic amine and nucleoside GPCRs. Glutamate residues in the E2-loop of the human A2a adenosine receptor are indirectly or directly involved in ligand binding (Kim et al., 1996). For the α1-adrenergic receptor, amino acids, located in the extracellular loop E2, play a role in subtype-selective antagonist binding (Zhao et al., 1996). In addition, the affinity of yohimbine to the α2A-adrenergic receptor is significantly influenced by interactions with the extracellular loop E2 (Laurila et al., 2007). Moreover, specific residues of the E2-loop are directly involved in forming the binding pocket of the dopamine D2 receptor (Shi and Javitch, 2004). For the muscarinic M3 receptor, several amino acids in the E2-loop are important for efficient agonist-induced activation of the muscarinic M3 receptor (Scarselli et al., 2007). In contrast to these results, the E2-loop does not contribute to the species selectivity of guanidine-type agonists at human and guinea pig histamine H2 receptor (Preuss et al., 2007). Our present study shows that the E2-loop and the E2-loop in combination with the N terminus have ligand-specific influence on the pharmacology of the H1R. Based on these data, it can be concluded that the extracellular loop E2 as well as the N terminus contribute to ligand binding, receptor activation, and selectivity in biogenic amine and nucleoside GPCRs. Therefore, it seems to be worthwhile to study the influence of N terminus and E2-loop on pharmacology and functionality of GPCRs in more detail.
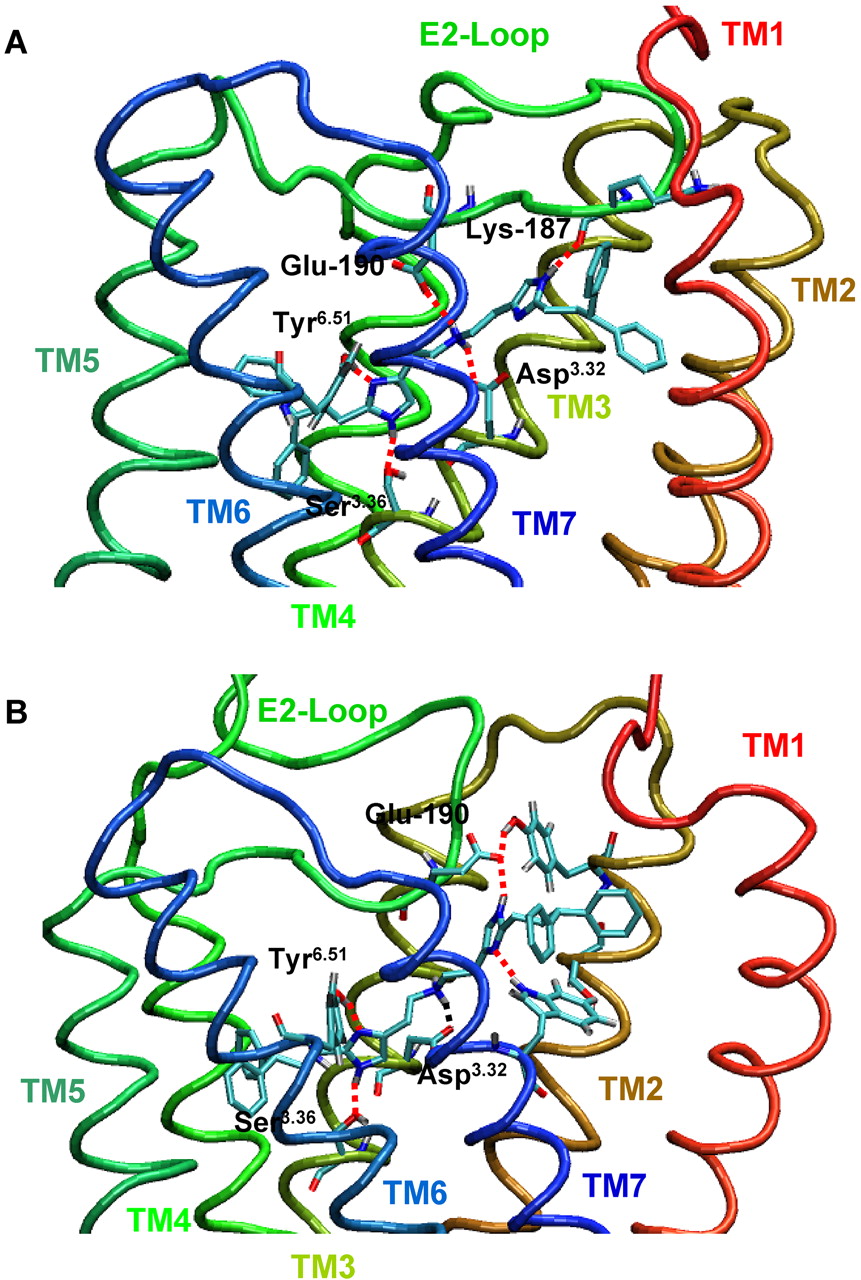
Binding mode of dimeric histaprodifen 7 in the active gpH1R model. Snapshot of dimeric histaprodifen 7 in the binding pocket of the active gpH1R model during MD simulation. A, model based on the crystal structure of bovine rhodopsin. B, model based on the crystal structure of the hβ2AR. MD simulations were performed as described under Materials and Methods. Hydrogen bonds and electrostatic interactions between ligand and amino acids are marked as red dashed lines.
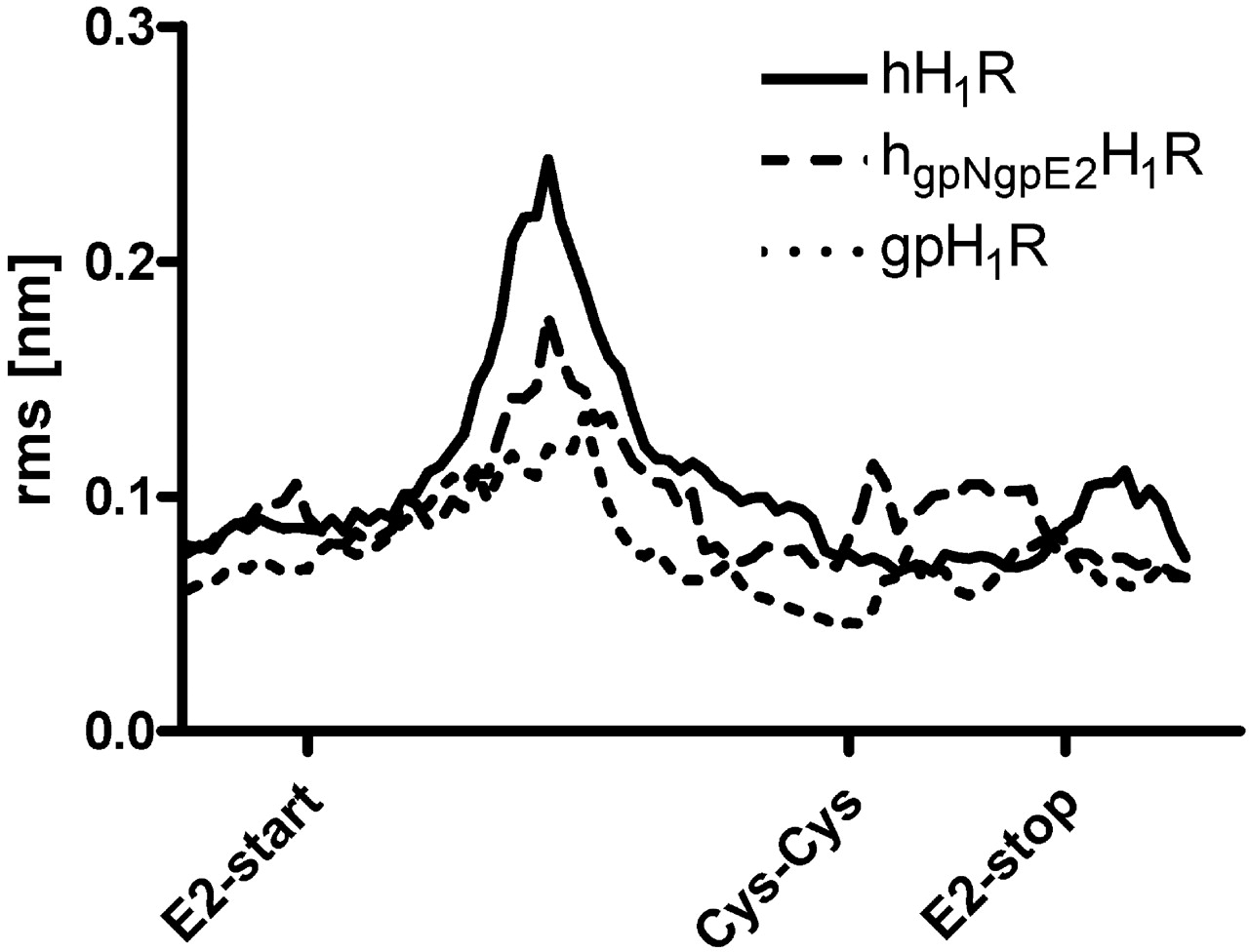
Root mean square (rms) fluctuation of the backbone atoms of the E2-loop. The label Cys-Cys indicates the position of the disulfide bridge between E2-loop and TM3. This part of the E2-loop, which shows large species differences and is in no contact with the binding pocket, exhibits a decrease in backbone fluctuation within the series hH1R > hgpNgpE2H1R > gpH1R (region between the labels E2-start and Cys-Cys). The region of the E2-loop with no species differences and contact to the ligand in the binding pocket shows a higher fluctuation for the hgpNgpE2H1R compared with the wild-type H1Rs. For all species, the fluctuation is reduced in the region of the disulfide bridge (Cys-Cys). All calculations were performed with GROMACS 3.3.1.
Conclusion
Our studies have shown that the N terminus and E2-loop have ligand-specific influence onto the pharmacology of the H1R. For a new class of histaprodifens, the phenoprodifens 5, 6R, and 6S and the pKi and pEC50 values (determined at hH1R and the chimeric hgpE2H1R and hgpNgpE2H1R) are not increased in direction of the gpH1R but decreased to the contrary direction. Thus, despite the decrease in species differences in the amino acid sequence between hH1R and gpH1R, in the series hH1R → hgpE2H1R → hgpNgpE2H1R → gpH1R, the differences in pharmacology increased for the phenoprodifens.
Acknowledgments
We thank Dr. S. Elz (Department of Pharmaceutical and Medicinal Chemistry I, University of Regensburg, Germany) for helpful discussions and support; K. Wohlfahrt for performing the molecular biological experiments and GTPase and binding assays; C. Huber for performing GTPase and binding assays; A. Seefeld for performing GTPase assays; and G. Wilberg for cell culture and immunoblot analysis.
Footnotes
-
This work was supported by the Research Training Program (Graduiertenkolleg) GRK760 “Medicinal Chemistry: Molecular Recognition–Ligand-Receptor Interactions” of the Deutsche Forschungsgemeinschaft.
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.108.140913.
-
ABBREVIATIONS: GPCR, G protein-coupled receptor; H1R, histamine H1 receptor; gp, guinea pig; hH1R, human H1R; gpH1R, guinea pig H1R; E2-loop, second extracellular loop; hgpE2H1R, hH1R with gp E2-loop; hgpNgpE2H1R, hH1R with gp N terminus and gp E2-loop; RGS4, regulator of G protein signaling 4; h, human; TM, transmembrane domain; S, signal peptide; F, FLAG epitope; PCR, polymerase chain reaction; DMSO, dimethyl sulfoxide; hβ2AR, human β2-adrenergic receptor; MD, molecular dynamics; [3H]MEP, mepyramine; Cpd, compound; HA, histamine.
- Received May 7, 2008.
- Accepted June 23, 2008.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}