


Abstract
In our study, ampicillin (AMP)-mediated decrease of enterobacteria caused increases in hepatic bile acid concentration through (at least in part) elevation of bile acid synthesis in C57BL/6N mice. We investigated the involvement of enterobacteria on intestinal bile acid absorption in AMP-treated mice in the present study. Fecal enterobacterial levels and fecal bile acid excretion rates were markedly decreased in mice treated with AMP (100 mg/kg) for 3 days, whereas bile acid concentrations in portal blood were significantly increased compared with those in mice treated with a vehicle. Ileal apical sodium-dependent bile acid transporter (SLC10A2) mRNA levels and ileal SLC10A2 protein levels in brush-border membranes were significantly increased compared with those in mice treated with the vehicle. In AMP-treated mice, total bile acid levels were increased, whereas levels of enterobacteria-biotransformed bile acid, taurodeoxycholic acid, and cholic acid were decreased in intestinal lumen. These phenomena were also observed in farnesoid X receptor-null mice treated with AMP for 3 days. Discontinuation of AMP administration after 3 days (vehicle administration for 4 days) increased levels of fecal enterobacteria, fecal bile acid excretion, and taurodeoxycholic acid and cholic acid in the intestinal lumen, whereas the discontinuation decreased ileal SLC10A2 expression and bile acid concentrations in the portal blood. Coadministration of taurodeoxycholic acid or cholic acid decreased ileal SLC10A2 expression in mice treated with AMP. These results suggest that enterobacteria-mediated bile acid biotransformation modulates intestinal bile acid transport and homeostasis through down-regulation of ileal SLC10A2 expression.
Introduction
Enterobacteria have been proposed to have a potential association with human health. They have been implicated in the pathogenesis of host disease such as nonalcoholic steatohepatitis, allergy, and inflammatory bowel disease (Noverr et al., 2004; Dumas et al., 2006; Frank et al., 2007; Alverdy and Chang, 2008). The composition of enterobacterial flora is significantly influenced by alterations in diet (Turnbaugh et al., 2006) or antibiotic use (Jernberg et al., 2007). In symbiosis with the host, enterobacteria play a crucial part in the metabolism of endogenous compounds such as bile acids as well as exogenous compounds. Some enterobacteria catalyze deamidation and 7α-dehydroxylation of amidated primary bile acids, resulting in the production of unconjugated primary bile acids and more hydrophobic secondary bile acids such as deoxycholic acid (DCA) and lithocholic acid (LCA). Thus, reduction of the number of enterobacteria by the administration of antibacterial drugs causes alteration of the composition of bile acids (Carulli et al., 1981; Berr et al., 1996). High concentrations of hepatobiliary secondary bile acids are linked to the pathogenesis of gallstone disease, hepatic injury, and colon cancer. The physiological significance of enterobacteria-mediated biotransformation of bile acids (deamidation and 7α-dehydroxylation) is incompletely understood.
Bile acids have roles not only in the intestinal absorption of lipid and lipid-soluble vitamins, but also in the activation of nuclear receptor and membrane receptor signaling to regulate the metabolism of lipids and carbohydrates (Fiorucci et al., 2009; Lefebvre et al., 2009). Bile acids are amphipathic and cytotoxic, and intense elevation of the concentration of bile acids in the liver causes hepatobiliary injury. Thus, the size of the bile acid pool is strictly regulated by bile acid-mediated negative feedback mechanisms via the synthesis of bile acids in the liver (Goodwin et al., 2000; Lu et al., 2000). More than 95% of the secreted bile acids from the liver are reabsorbed from the intestine and returned to the liver through portal blood. Thus, the hepatic synthesis of bile acids and intestinal absorption of bile acids have been suggested to be crucial determinants of the size of the bile acid pool.
Most absorption of bile acids in the intestine consists of an ileal active transport system through the apical sodium-dependent bile acid transporter (SLC10A2), ileal bile acid binding protein (IBABP), and organic solute transporter α/β (OSTα/β) (Kosters and Karpen, 2008; Dawson et al., 2009). In SLC10A2-null mice, marked increases in the excretion of fecal bile acid and decreases in the size of the bile acid pool have been observed (Dawson et al., 2003). Mutations in the human SLC10A2 gene are related to malabsorption of intestinal bile acids (Oelkers et al., 1997). Those reports highlight the crucial role of SLC10A2 in intestinal bile acid absorption.
Bile acid signaling suppresses liver receptor homolog-1 or retinoic acid receptor-mediated transcriptional activation of the SLC10A2 gene through the FXR/SHP pathway (Chen et al., 2003; Neimark et al., 2004; Li et al., 2005). Furthermore, bile acid activated FXR/FGF15/19 signaling acts through FGF receptor 4 in an autocrine manner to suppress SLC10A2 expression in mice (Sinha et al., 2008). Conversely, bile acids are also found to increase human SLC10A2 transcription through FXR-independent signaling such as mitogen-activated protein kinase and epidermal growth factor receptor (Duane et al., 2007). The regulation of SLC10A2 expression in vivo by bile acids remains controversial. Conflicting observations that SLC10A2 expression and ileal bile acid absorption are induced, repressed, or unaffected by bile acids have been reported (Lillienau et al., 1993; Stravitz et al., 1997; Arrese et al., 1998).
We have found that decreased levels of enterobacteria caused by administration of antibacterial drugs elevated levels of hepatic bile acids and bile acid synthesis (at least in part) through reduction of ileal FGF15 expression caused by enterobacteria-mediated biotransformation of bile acids (Miyata et al., 2009b). The relationship between enterobacterial overgrowth and malabsorption of bile acids has been suggested in many patients with irritable bowel syndrome (Fan and Sellin, 2009). These observations underpin the hypothesis that enterobacteria negatively regulate intestinal bile acid absorption and hepatic bile acid synthesis through the biotransformation of amidated primary bile acids. To test this hypothesis, we examined the influence of decreases in the number of enterobacteria on intestinal absorption of bile acids and ileal SLC10A2 expression in mice treated with the antibacterial drug ampicillin (AMP).
Materials and Methods
Materials.
AMP was purchased from Nacalai Tesque Inc. (Kyoto, Japan). Cholic acid (CA), taurocholic acid (TCA), chenodeoxycholic acid, taurochenodeoxycholic acid (TCDCA), DCA, taurodeoxycholic acid (TDCA), LCA, and taurolithocholic acid were purchased from Sigma-Aldrich (St. Louis, MO). β-Muricholic acid, tauro-β-muricholic acid (TβMCA), ursodeoxycholic acid, tauroursodeoxycholic acid, etiocholan-3α, 17β-diol (5β-androstan-3α,17β-diol), and 7α-hydroxycholesterol were purchased from Steraloids (Newport, RI). An L-column ODS (2.1 × 150 mm) was obtained from the Chemicals Evaluation and Research Institute (Tokyo, Japan). An Enzymepak 3α-HSD column was purchased from Jasco (Tokyo, Japan). A polyclonal anti-rodent ASBT (SLC10A2) antibody was kindly provided by Dr. Paul A. Dawson (Wake Forest University School of Medicine, Winston-Salem, NC).
Animal Treatment and Sample Collection.
The protocol was approved by the Institutional Animal Care and Use Committee of Tohoku University in Sendai, Japan. Male C57BL/6N mice (Charles River Japan Incorporated, Yokohama, Japan) and farnesoid X receptor (Fxr)-null mice (Sinal et al., 2000) were housed under a standard 12-h light/dark cycle (9:00 AM to 9:00 PM). Before experimentation, mice were fed standard rodent chow (CE-2; CLEA Japan, Tokyo, Japan) and water ad libitum for acclimatization. Age-matched groups of 8- to 9-week-old mice were used for all experiments. Mice were administered (per os) AMP (100 mg/kg body weight, dissolved in 0.9% saline) or the vehicle at 9:00 AM for 3, 7, and 14 days. To examine the influence of the discontinuation of AMP administration, mice were treated with AMP for 3 days followed by vehicle treatment for 4 days (AMP/vehicle). For the bile acid cotreatment experiment, mice were administered (per os) AMP (100 mg/kg body weight, dissolved in 0.9% saline) at 9:00 AM for 7 days and coadministered (per os) TDCA or CA (500 mg/kg body weight) for the final 4 days. They were killed 24 h after the last administration (9:00 AM). Feces were collected 24 h after the last administration. Liver samples were obtained for biochemical assays. The small intestinal lumen was washed with phosphate-buffered saline, and the washed solution was collected. Biliary excretion was monitored in mice anesthetized with ethyl ether. After ligating the common bile duct, bile samples were collected by cannulating the gall bladder using polyethylene tubing (PE-10; internal diameter, 0.28 mm). After a 5-min equilibration period, bile was collected for 30 min. Portal blood was collected from mice anesthetized with ethyl ether.
Analyses of Bile Acid Components.
Bile acid composition in the liver, small intestinal lumen, feces, and portal blood was measured by high-performance liquid chromatography (HPLC) as described previously (Kitada et al., 2003; Miyata et al., 2006). Isolated feces and the washed solution of the small intestinal lumen were homogenized in 20 volumes (v/w) or equal volume (v/v) of 50% tert-butanol, respectively, using Polytron. Homogenates were centrifuged at 20,000g for 10 min. Supernatants were subjected to HPLC analyses. The contents of β-muricholic acid, TβMCA, ursodeoxycholic acid, tauroursodeoxycholic acid, CA, TCA, chenodeoxycholic acid, TCDCA, DCA, TDCA, LCA, and taurolithocholic acid were measured.
Western Blot Analyses.
Ileal brush border membranes (BBMs) were prepared as described previously (Kessler et al., 1978). Ileal segments (5 cm) were opened, and the ileal mucosa was obtained by scraping. The ileal mucosa was suspended in 2 mM Tris-HCl, pH 7.1, buffer containing 50 mM mannitol and 0.1 mM phenylmethanesulfonyl fluoride. The mucosa was homogenized by using a Teflon glass homogenizer. After addition of CaCl2 to a final concentration of 10 mM, the homogenate was incubated on ice for 20 min. The homogenate was centrifuged at 3000g for 15 min, and the supernatant was centrifuged at 27,000g for 30 min. The pellet was suspended in 1 mM HEPES (pH 7.5) buffer containing 100 mM mannitol and 10 mM MgSO4. Ileal BBMs (2.5 μg protein/lane) were subjected to SDS-polyacrylamide gel electrophoresis on 8 or 10% polyacrylamide gels and transferred to nitrocellulose filters. The ileal BBM filters were probed with a polyclonal anti-rodent ASBT (SLC10A2) antibody (Shneider et al., 1995; Wong et al., 1995) or monoclonal β-actin antibody (Sigma-Aldrich). Filters were washed five times with Tween 20 containing phosphate buffer and incubated with phosphatase-conjugated goat anti-rabbit IgG (1:3000 dilution) or rabbit anti-mouse IgG (1:10,000 dilution). Imunoreactive bands were detected by using 5-bromo-4-chloro-3-indolylphosphate disodium salt and nitrotetrazolium blue chloride. Stained membranes were scanned with an Epson GT-8700 scanner (Seiko Epson, Suwa, Japan).
Analyses of mRNA Levels.
Total RNA was prepared from the liver and ileum by using an RNAgents Total Isolation System (Promega, Madison, WI). RNA concentration was determined by measuring the absorbance at 260 nm using a DU800 spectrophotometer (Beckman Coulter, Fullerton, CA). Single-stranded cDNA were synthesized by using an oligo(dT) primer and a Ready-To-Go You-Prime First-Strand Beads kit (GE Healthcare, Chalfont St. Giles, Buckinghamshire, UK). These cDNA templates were subjected to real-time quantitative polymerase chain reaction (qPCR) using SYBR Green 1 with an ABI PRISM 7000 Sequence Detection System (Applied Biosystems, Foster City, CA). Relative mRNA levels were calculated by the comparative threshold cycle method. The following specific forward and reverse primers were used for real-time qPCR: SLC10A2 (ASBT), sense, 5′-TGGGTTTCTTCCTGGCTAGACT-3′ and antisense, 5′-TGTTCTGCATTCCAGTTTCCAA-3′; OSTα, sense, 5′-ATGCATCTGGGTGAACAGAA-3′ and antisense, 5′-GAGTAGGGAGGTGAGCAAGC-3′; OSTβ, sense, 5′-AGATGCGGCTCCTTGGAATTA-3′ and antisense, 5′-TGGCAGAAAGACAAGTGATG-3′; IBABP, sense, 5′-AGATCATCACAGAGGTCCAGC-3′ and antisense 5′-GGTAGCCTTGAACTTCTTGCC-3′; CYP7A1, sense, 5′-AGCAACTAAACAACCTGCCAGTACTA-3′ and antisense, 5′-GTCCGGATATTCAAGGATGCA-3′; SHP, sense, 5′-CGATCCTCTTCAACCCAGATG-3′ and antisense, 5′-AGGGCTCCAAGACTTCACACA-3′; FGF15, sense, 5′-GAGGACCAAAACGAACGAAATT-3′ and antisense, 5′-ACGTCCTTGATGGCAATCG-3′; glyceraldehyde-3-phosphate dehydrogenase, sense, 5′-TGTGTCCGTCGTGGATCTGA-3′ and antisense, 5′-CCTGCTTCACCACCTTCTTGAT-3′; and β-actin, sense, 5′-ACCCTGTGCTGCTCACCGA-3′ and antisense, 5′-CTGGATGGCTACGTACATGGCT-3′.
Quantification of Bacterial DNA in Feces.
The fecal content of bacteria in stools collected over the final 24 h in vehicle and AMP-treated mice was determined. DNA was extracted from 200 mg of feces by using a QIAamp DNA Stool Mini Kit (QIAGEN, Tokyo, Japan). The extracted DNA was diluted in EDTA buffer, and the purity was confirmed by measuring absorbance at 260 and 280 nm using a DU800 spectrophotometer (Beckman Coulter). These DNA samples were subjected to real-time PCR using SYBR Green 1 with an ABI PRISM 7000 Sequence Detection System (Applied Biosystems). Relative DNA levels were calculated by the comparative threshold cycle method. The following specific forward and reverse primers were used for real-time qPCR: Bacteroides fragilis, sense, 5′-CTGAACCAGCCAAGTAGCG-3′ and antisense, 5′-CCGCAAACTTTCACAACTGACTTA-3′; Clostridium clostridiiforme, sense, CCGCATGGCAGTGTGTGAAA-3′ and antisense, 5-CTGCTGATAGAGCTTTACATA-3′; Clostridium scindens, sense, 5′-GCAACCTGCCTGCACT-3′ and antisense, 5′-ACCGAATGGCCTTGCCA-3′; Clostridium sordellii, sense, 5′-TCGAGCGACCTTCGG-3′ and antisense, 5′-CACCACCTGTCACCAT-3′; Fusobacterium prausnitzii, sense, 5′-AGATGGCCTCGCGTCCGA-3′ and antisense, 5′-CCGAAGACCTTCTTCCTCC-3′; and Peptostreptococcus productus, sense, 5′-AACTCCGGTGGTATCAGATG-3′ antisense, 5′-GGGGCTTCTGAGTCAGGTA-3′.
Statistical Analyses.
Values are presented as mean ± S.D. Data were analyzed by unpaired Student's t test or analysis of variance followed by Dunnett's method using Prism 4.0 software (GraphPad Software Inc., San Diego, CA) for evaluating differences between the mean values of each group. p < 0.05 was considered significant.
Results
Influence of AMP Treatment on Levels of Fecal Enterobacteria and Hepatobiliary Bile Acids.
As reported in our previous study, treatment with antibacterial drug (AMP or bacitracin/neomycin/streptomycin) stimulated hepatic bile acid synthesis, which resulted in elevated levels of hepatic bile acid in mice. In the present study, to identify the role of enterobacteria in the regulation of levels of hepatobiliary bile acids, we analyzed fecal levels of enterobacteria and hepatobiliary bile acids in AMP-treated mice. In mice treated with AMP (100 mg/kg) for 3 days, the number of representative enterobacteria in feces was markedly reduced (from 1/10 to 1/10,000) (Fig. 1A). At this time point, consistent with previous experiments, hepatic bile acid concentrations were significantly increased in AMP-treated mice (Fig. 1B). Furthermore, biliary bile acid output rates and portal blood bile acid concentrations were significantly increased in AMP-treated mice (Fig. 1, C and D). Conversely, the excretion rates of fecal bile acid were clearly decreased in mice (Fig. 1E), suggesting that the capacity of absorption of intestinal bile acids in the mice was elevated.

AMP treatment alters fecal enterobacteria level and bile acid homeostasis. Male C57BL/6N mice were treated with AMP (100 mg/kg p.o.) for 3 days. A, fecal enterobacteria DNA levels. B, hepatic bile acid concentration. C, biliary bile acid output rate. D, concentration of bile acids in portal blood. E, fecal bile acid excretion rate. These bile acid levels were analyzed by HPLC. Data are mean ± S.D. (n = 4). Significant differences from the vehicle-treated group are indicated: *, p < 0.05; **, p < 0.01; ***, p < 0.001.
Influence of AMP Treatment on Expression of Ileal Bile Acid-Related Genes.
SLC10A2 transfers bile acids from the intestinal lumen across the apical BBM. Bile acids are then transferred to the basolateral membrane. IBABP seems to be involved in the transfer of bile acids from the apical membrane to the basolateral membrane. OSTα/β extrudes bile acids into the portal circulation. To identify a mechanism for elevated absorption of bile acids in AMP-treated mice, levels of ileal SLC10A2, IBABP, OSTα, and OSTβ mRNA were estimated by real-time qPCR. Specific levels of IBABP, OSTα, and OSTβ mRNA were significantly reduced at 3 days in mice treated with AMP, whereas only the level of SLC10A2 mRNA was significantly increased (Fig. 2A). Consistent with significant increases in the levels of ileal SLC10A2 mRNA, the amounts of SLC10A2 protein in ileal BBMs estimated by Western blot analyses were significantly increased in AMP-treated mice for 3 days compared with those in vehicle-treated mice (Fig. 2B). SLC10A2 protein contents in ileal BBMs of AMP-treated mice were 3-fold higher than those in vehicle-treated mice. Specific mRNA levels of ileal FGF15 and SHP (which are involved in the negative regulation of SLC10A2 expression) were also analyzed. Both mRNA levels were significantly decreased in AMP-treated mice (Fig. 2C). Theses results suggest that reduction of enterobacteria may attenuate FXR signaling. The influence of AMP-treatment was then investigated using Fxr-null mice.
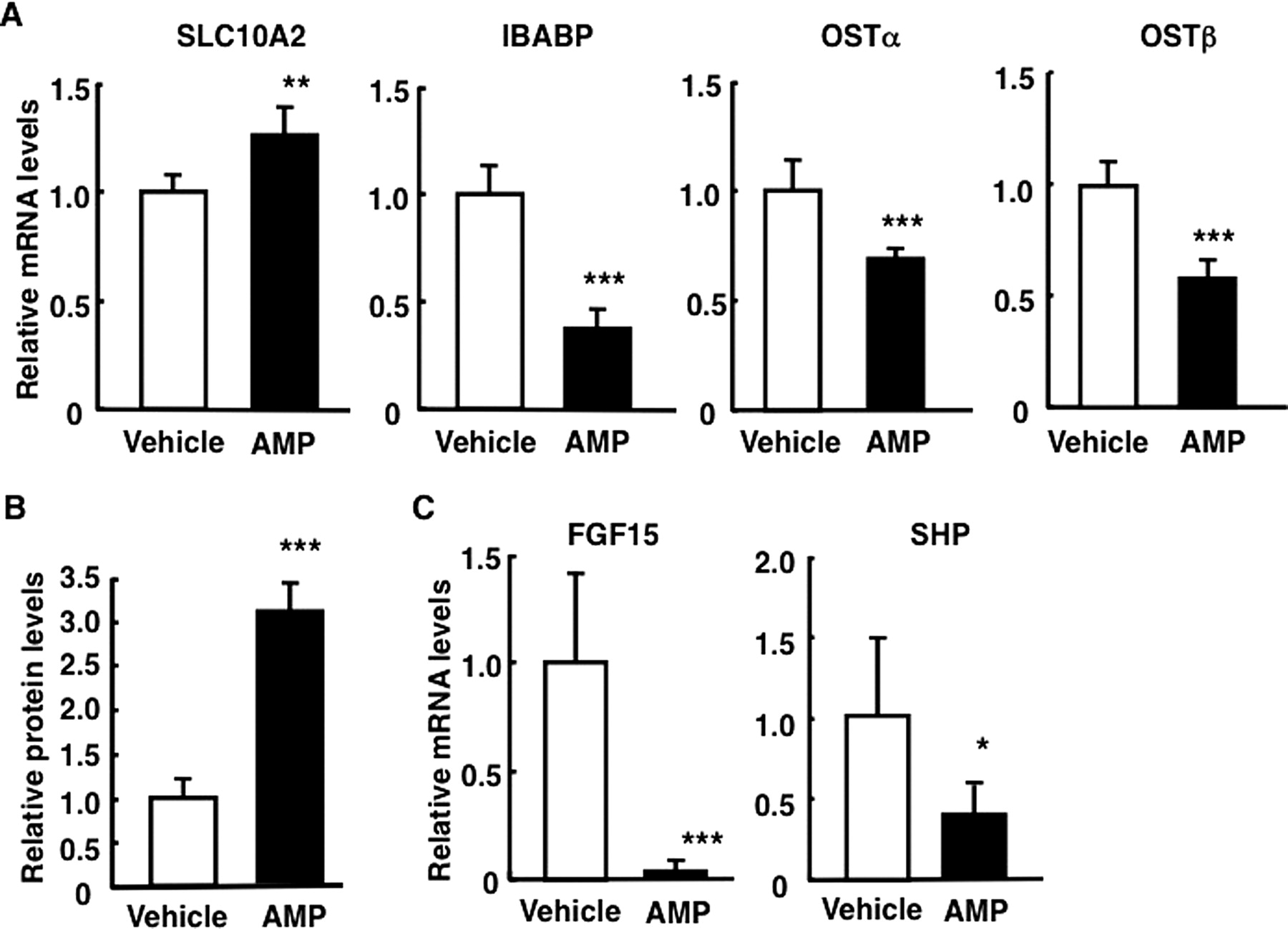
AMP treatment alters expression levels of ileal bile acid-related genes. Male C57BL/6N mice were treated with AMP (100 mg/kg p.o.) for 3 days. A, mRNA levels of ileal bile acid-related genes. B, SLC10A2 protein amounts in ileal BBMs. C, mRNA levels of transcription factors. Data are mean ± S.D. (n = 4). Significant differences from the vehicle-treated group are indicated: *, p < 0.05; **, p < 0.01; ***, p < 0.001.
Influence of AMP Treatment on Levels of Hepatobiliary Bile Acids in Fxr-Null Mice.
Hepatic bile acid concentration is higher in Fxr-null mice than in wild-type mice (Kitada et al., 2003). The level was further enhanced after treatment with AMP in Fxr-null mice (Fig. 3A). Treatment with AMP of Fxr-null mice resulted in the decreased excretion of fecal bile acids and increased concentration of portal blood bile acids (Fig. 3, B and C). Significant increases in ileal SLC10A2 mRNA levels and ileal SLC10A2 protein amounts in BBMs were also observed in AMP-treated Fxr-null mice (Fig. 3, D and E). These alterations in Fxr-null mice were consistent with those in C57BL/6N mice.

AMP treatment alters fecal enterobacteria level and bile acid homeostasis in Fxr-null mice. Male Fxr-null mice were treated with AMP (100 mg/kg p.o.) or the vehicle for 3 days. A, hepatic bile acid concentration. B, fecal bile acid excretion rate. C, concentration of bile acids in portal blood. D, ileal SLC10A2 mRNA levels. E, SLC10A2 protein amounts in ileal BBMs. Data are mean ± S.D. (n = 5). Significant differences from the vehicle-treated group are indicated: *, p < 0.05; **, p < 0.01.
Altered Response after AMP Treatment.
To understand the mechanism of enterobacteria involvement on ileal bile acid absorption, mice were treated with AMP for 3, 7, or 14 days. To examine the influence of the discontinuation of AMP administration, a group of mice were treated with AMP for 3 days followed by vehicle treatment for 4 days (AMP/vehicle). B. fragilis and C. clostridiiforme can mediate bile acid 7α-dehydroxylation and deamino-conjugation. These levels of bacterial DNA in feces were thus estimated by real-time qPCR. These levels of bacterial DNA in feces were markedly low in mice treated with AMP at 3 or 7 days compared with those in mice treated with the vehicle (Fig. 4A). Bacterial levels in feces were gradually increased in a time-dependent manner, and a significant difference was not observed between mice treated with AMP for 14 days and those treated with the vehicle. The discontinuation of AMP administration reversed reduced bacterial DNA levels.

Time-dependent effects of AMP treatment on fecal enterobacteria levels and ileal bile acid absorption. Male C57BL/6N mice were treated with AMP (100 mg/kg p.o.) or the vehicle for 3, 7, or 14 days. To evaluate the effect of AMP discontinuation, mice were treated with the vehicle for 4 days after AMP treatment for 3 days (AMP/vehicle). A, fecal enterobacteria levels. B, fecal bile acid excretion rate. C, concentration of bile acids in portal blood. Data are mean ± S.D. (n = 4). Significant differences from the vehicle-treated group are indicated: *, p < 0.05; **, p < 0.01; ***, p < 0.001.
Fecal bile acid excretion in mice treated with AMP for 3 or 7 days was markedly decreased compared with those in mice treated with the vehicle, whereas no clear difference was detected between mice treated with AMP for 14 days and those treated with the vehicle (Fig. 4B). Conversely, the discontinuation of AMP administration reversed the reduced excretion rates of fecal bile acid in mice treated with AMP for 3 days. Bile acid concentrations in portal blood were significantly increased in mice treated with AMP for 3 or 7 days compared with those in mice treated with the vehicle, whereas the concentrations were not increased in mice treated for 14 days (Fig. 4C). The discontinuation of AMP administration decreased concentrations of portal bile acid.
Changes in Expression Levels of Ileal SLC10A2 in AMP-Treated Mice in a Time-Dependent Manner.
Ileal SLC10A2 mRNA levels were significantly increased in mice treated with AMP for 3 or 7 days (Fig. 5A). However, no significant increases in SLC10A2 mRNA levels were observed in mice treated with AMP for 14 days. The discontinuation of AMP administration decreased SLC10A2 mRNA levels. Conversely, changes in ileal SLC10A2 protein amounts in BBMs were similar to those in SLC10A2 mRNA levels (Fig. 5B). Ileal SLC10A2 protein amounts in mice treated with AMP for 14 days were reduced to approximately those in vehicle-treated mice. The discontinuation of AMP administration completely reversed increased SLC10A2 protein amounts in mice treated with AMP for 3 days. As expected, changes in mRNA levels of ileal FGF15 and SHP (which are involved in the negative regulation of SLC10A2 expression) were inversely correlated with those of SLC10A2 (Fig. 5A).

Time-dependent effects of AMP-treatment on ileal SLC10A2 expression. Male C57BL/6N mice were treated with AMP (100 mg/kg p.o.) or the vehicle for 3, 7, or 14 days. To evaluate the effect of AMP discontinuation, mice were treated with the vehicle for 4 days after AMP treatment for 3 days (AMP/vehicle). A, ileal SLC10A2, FGF15, and SHP mRNA levels. Data are mean ± S.D. (n = 4). Significant differences from the vehicle-treated group are indicated: *, p < 0.05; **, p < 0.01. B, SLC10A2 protein amounts in ileal BBMs. Ileal BBMs were individually isolated from four treated mice. Isolated BBMs were pooled and subjected to Western blot analyses using a polyclonal anti-rodent ASBT (SLC10A2) antibody or monoclonal anti-β-actin antibody.
Influence of AMP Treatment on the Composition of Bile Acids in the Intestinal Lumen.
The composition of bile acids in the small intestinal lumen was determined to understand the reason for the increased SLC10A2 mRNA levels. The amounts of enterobacteria-biotransformed bile acid, CA, and TDCA were decreased to below detection limits (<0.01 μmol) in mice treated with AMP for 3 or 7 days, whereas the amounts of the primary bile acids, TCA, and TβMCA were significantly increased in mice treated with AMP for 3 or 7 days, resulting in an increase in the total amounts of bile acids in the intestinal lumen (Table 1). Discontinuation of AMP administration caused the recovery of CA and TDCA in the intestinal lumen of mice treated with AMP for 3 days. Consistent with reversed enterobacteria levels, amounts of CA and TDCA in the intestinal lumen were not decreased to below detection limits in mice treated with AMP for 14 days. Total amounts of bile acids in the lumen of small intestine were increased in mice treated with AMP for 3, 7, or 14 days.
Influence of AMP treatment on bile acid composition in the intestinal lumen
Data are shown as mean ± S.D. (n = 5).
Influence of Coadministration of Bile Acids on SLC10A2 Expression in AMP-Treated Mice.
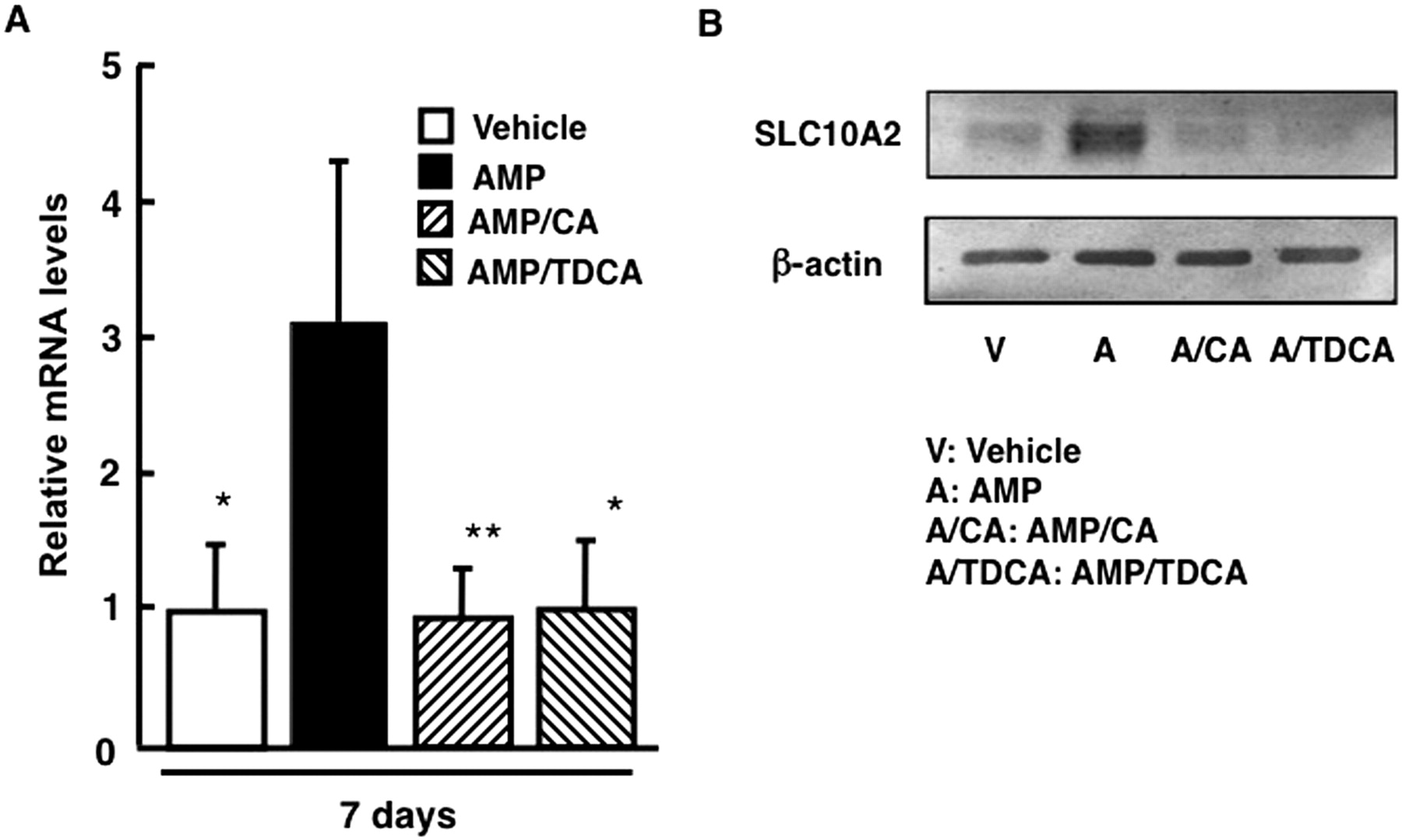
To verify the influence of cotreatment with enterobacteria-biotransformed bile acids in AMP-treated mice on ileal SLC10A2 expression, SLC10A2 expression levels were analyzed in mice cotreated with AMP and CA or TDCA by Western blot analysis and real-time qPCR. Cotreatment with CA or TDCA reversed increased ileal SLC10A2 mRNA levels in AMP-treated mice (Fig. 6). Consistent with SLC10A2 mRNA levels, CA or TDCA-cotreatment reversed increased SLC10A2 protein amounts in ileal BBMs of AMP-treated mice.
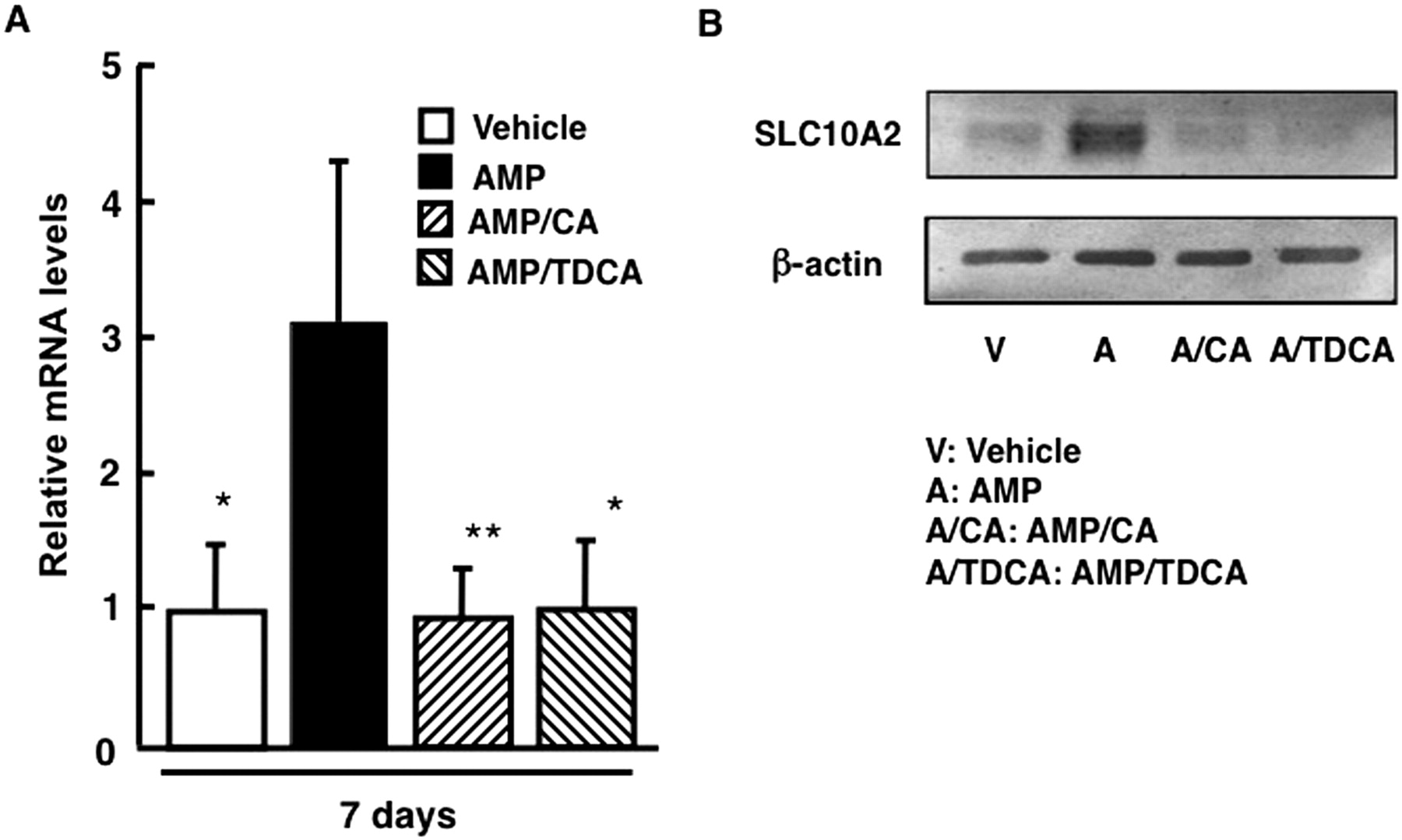
Influence of coadministration of bile acids on ileal SLC10A2 expression. Male C57BL/6N mice were treated with AMP (100 mg/kg p.o.) for 7 days and cotreated with CA or TDCA (500 mg/kg) for the last 4 days. A, ileal SLC10A2 mRNA levels. Data are mean ± S.D. (n = 4). Significant differences from the AMP-treated group are indicated: *, p < 0.05; **, p < 0.01. B, SLC10A2 protein amounts in ileal BBMs. Ileal BBMs were individually isolated from four treated mice. Isolated BBMs were pooled and subjected to Western blot analyses using a polyclonal anti-rodent ASBT (SLC10A2) antibody or monoclonal anti-β-actin antibody.
Discussion
In the present study, we demonstrated that enterobacteria are involved in the suppression of ileal SLC10A2 expression through biotransformation of bile acids, resulting in the reduction of absorption of ileal bile acids and elevation of fecal bile acids. It is noteworthy that excretion of fecal bile acid is decreased in AMP-treated mice, despite increased hepatic bile acid synthesis through the reduction of ileal FGF15 expression (Miyata et al., 2009b). Decreases in the number of enterobacteria after treatment with AMP for 3 days caused disruption of bile acid homeostasis. Furthermore, concomitant with the reversal of decreased numbers of enterobacteria, bile acid homeostasis was restored after long-term administration of AMP (14 days) and discontinuation of AMP administration. These phenomena are consistent with the fact that patients taking antibacterial drugs do not cause severe hepatobiliary injury. Furthermore, theses results indicate that enterobacteria have a high ability to survive antibacterial environment. This ability of enterobacteria is probably convenient for the host to maintain bile acid homeostasis.
The total amounts of bile acids in the intestinal lumen were increased in AMP-treated mice for 3 or 7 days, whereas enterobacteria-biotransformed bile acids such as TDCA and CA were decreased to below the detection limit (0.01 μmol in the intestinal lumen). SLC10A2 expression was increased in AMP-treated mice (3 or 7 days) in which total levels of bile acids in the intestinal lumen and portal blood were elevated. These results raise the possibility that bile acid-mediated suppression of SLC10A2 expression depends on the levels of enterobacteria-biotransformed bile acids (CA and TDCA), but not total levels of bile acids and levels of amidated primary bile acid (TCA). The discontinuation of AMP administration reversed elevated expression of SLC10A2 and reduced the amounts of TDCA and CA in the intestinal lumen. Ileal SLC10A2 expression level was inversely correlated with enterobacteria-biotransformed bile acid amounts in the intestinal lumen. Coadministration of TDCA or CA decreased ileal SLC10A2 expression in AMP-treated mice. These results also support the idea that enterobacteria-biotransformed bile acids such as CA and TDCA are involved in the suppression of ileal SLC10A2 expression.
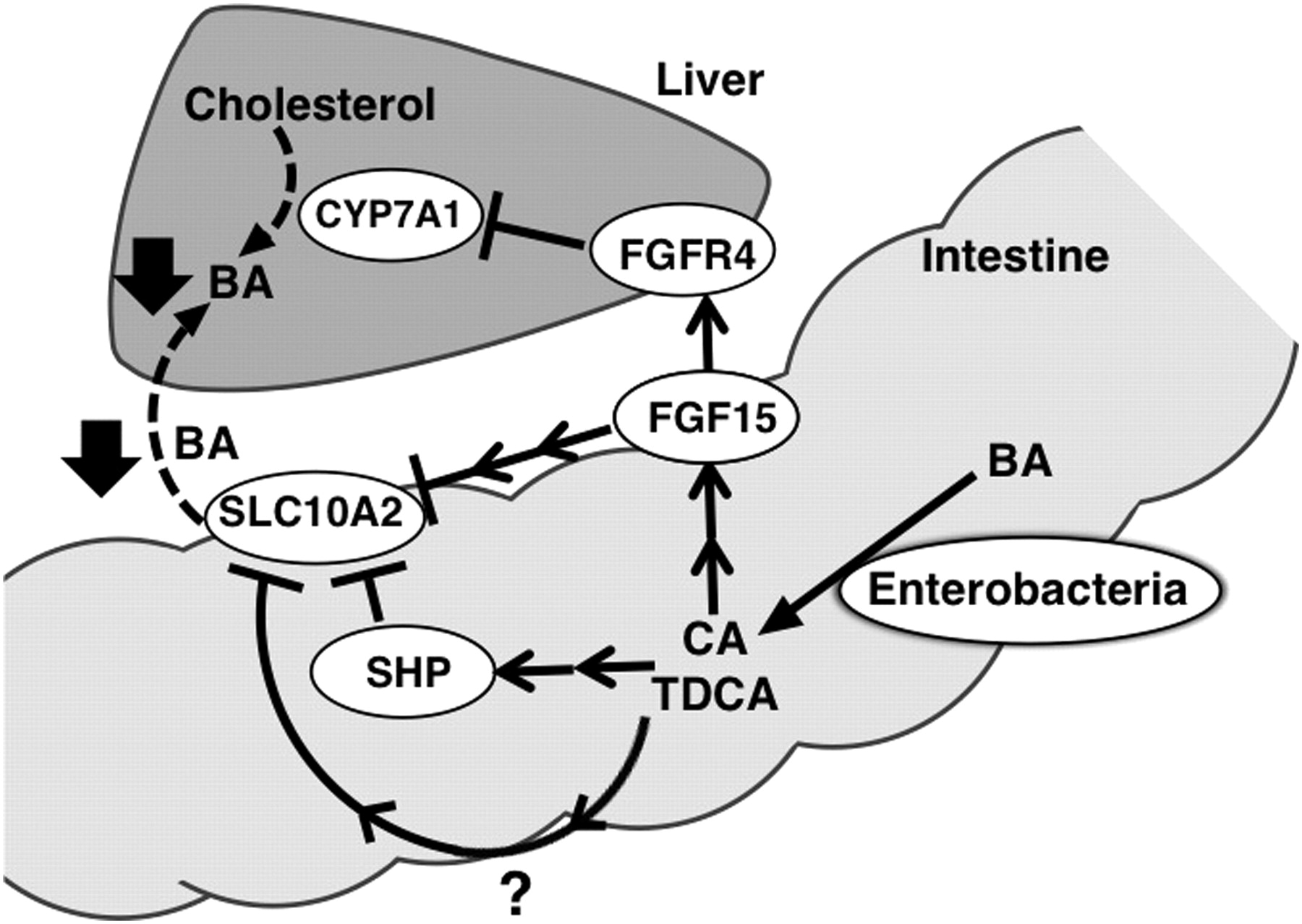
SLC10A2 expression in humans, rabbits, and mice is down-regulated by bile acid-activated FXR signaling through SHP (Chen et al., 2003; Neimark et al., 2004; Li et al., 2005). Thus, decreases in ileal SHP expression may be associated with the elevation of ileal SLC10A2 expression in AMP-treated mice. Reports have demonstrated that FGF15/19 signaling negatively regulates SLC10A2 expression in an autocrine manner (Sinha et al., 2008). Ileal SLC10A2 protein levels are decreased in C57BL/6 mice treated with FGF19 via the intraperitoneal route, whereas ileal FXR and SHP protein levels are increased. These facts suggest that the attenuation of ileal FGF15 signaling in an autocrine manner is involved in the elevation of ileal SLC10A2 expression in a similar manner to hepatic CYP7A1 expression in AMP-treated mice (Miyata et al., 2009b). However, involvement of FGF15 signaling may be limited to ileal SLC10A2 expression compared with hepatic CYP7A1, because of low expression levels of ileal FGF receptor 4 (Inagaki et al., 2005). Furthermore, increases in fecal bile acid excretion are observed in Fgf15-null mice in which hepatic bile acid synthesis is increased (Inagaki et al., 2005). These facts suggest the possible role of a bile acid-mediated mechanism (rather than the previously proposed ileal FGF15 signaling) for the elevation of ileal SLC10A2 expression in AMP-treated mice (Fig. 7).

Proposed mechanisms for enterobacteria-mediated regulation of hepatobiliary bile acid levels. Enterobacteria produce unconjugated bile acids and secondary bile acids such as CA and TDCA. These enterobacteria-biotransformed bile acids increase ileal FGF15/19 expression, resulting in the reduction of hepatic CYP7A1 expression and primary bile acid synthesis. Conversely, enterobacteria-biotransformed bile acids (CD and TDCA) suppress ileal SLC10A2 expression through multiple mechanisms, resulting in reduction of ileal bile acid absorption. Taken together, these results indicate enterobacteria suppress bile acid pool size through bile acid biotransformation. The dotted line indicates a decreased pathway. FGFR4, FGF receptor 4.
Furthermore, it is considered that the decreases in expression levels of ileal SHP and FGF15 in AMP-treated mice are (at least in part) caused by the attenuation of FXR signaling through reduction of enterobacteria-biotransformed bile acids such as TDCA and CA. Decreases in expression levels of FXR-targeted genes such as IBABP and OSTα/β in AMP-treated mice support this idea. Conversely, in Fxr-null mice treated with AMP, SLC10A2 mRNA and protein levels were increased, suggesting that SLC10A2 expression is also regulated by enterobacteria-mediated and FXR-independent signaling. Ileal mRNA levels of FGF15 (but not SHP) are significantly decreased in AMP-treated Fxr-null mice. In Fxr-null mice, decreased ileal FGF15 levels may be at least in part involved in the elevation of SLC10A2 expression. FXR-independent regulation of SLC10A2 expression and intestinal bile acid absorption have been demonstrated with Fxr-null mice (Miyata et al., 2009a). SLC10A2 gene expression is activated by peroxisome proliferator-activated receptor α and vitamin D receptor through the response elements in the promoter region (Jung et al., 2002; Chen et al., 2006).
In the present study, changes in SLC10A2 protein amounts in BBMs were not completely correlated with changes in ileal SLC10A2 mRNA levels. Proteasomal degradation of SLC10A2 protein probably is stimulated by cytokines such as interleukin-1β (Xia et al., 2004). It has been suggested that internalization of SLC10A2 from the membrane to intracellular compartment is modulated by protein kinase C signaling (Sarwar et al., 2009). Membrane expression levels of SLC10A2 protein are probably (at least in part) regulated through posttranslational mechanisms by some type of enterobacteria signaling. Enterobacteria-mediated cytokine signaling may be involved in the alteration of ileal SLC10A2 expression in AMP-treated mice. Further study is necessary to identify the mechanisms for suppressive regulation of SLC10A2 expression through enterobacterial signaling.
The present study demonstrated the physiological significance of enterobacteria-mediated bile acid biotransformation in the regulation of bile acid homeostasis. Enterobacteria produce secondary bile acids and unconjugated primary bile acids that act as signaling molecules to suppress hepatic primary bile acid synthesis and ileal bile acid absorption, resulting in reduction of the size of bile acid pool in the host. Enterobacteria may require the ability to survive in the small intestinal lumen through decreases in bile acid levels. Conversely, the host may incorporate the enterobacteria-mediated signaling as one of bile acid-regulating systems and also protect the distal small intestine from enterobacteria by using enterobacteria-biotransformed bile acid/FXR signaling (Inagaki et al., 2006).
In humans, enterobacteria produce mainly DCA and LCA as secondary bile acids. Humans do not have the 7α-hydroxylase conversion ability of secondary bile acids, so the proportion of secondary bile acids (especially amino-conjugated DCA) in humans is higher than in rodents (Ridlon et al., 2006). Thus, the regulation of hepatobiliary bile acid levels in humans may highly depend on enterobacteria-mediated signaling.
In conclusion, we showed that decreases in the number of enterobacteria by the administration of AMP elevated ileal SLC10A2 expression, resulting in the elevation of intestinal bile acid absorption and reduction of fecal bile acid excretion. Furthermore, we suggested that enterobacteria-mediated bile acid biotransformation is a critical determinant of the suppression of intestinal bile acid absorption caused by down-regulation of ileal SLC10A2 expression.
Authorship Contributions
Participated in research design: Miyata, Yamakawa, and Yamazoe.
Conducted experiments: Miyata, Yamakawa, Hamatsu, Kuribayashi, and Takamatsu.
Contributed new reagents or analytic tools: Takamatsu.
Performed data analysis: Miyata, Yamakawa, Hamatsu, and Kuribayashi.
Wrote or contributed to the writing of the manuscript: Miyata, Yamakawa, and Yamazoe.
Other: Miyata and Yamazoe acquired funding for the research.
Footnotes
This study was supported by the Ministry of Education, Culture, Sports Science, and Technology of Japan [Grants 21390039, 20590137] and the Yakult Bio-Science Foundation.
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
doi:10.1124/jpet.110.171736.
-
ABBREVIATIONS:
- DCA
- deoxycholic acid
- AMP
- ampicillin
- ASBT
- apical sodium-dependent bile acid transporter
- SHP
- small heterodimer partner
- FGF
- fibroblast growth factor
- FXR
- farnesoid X receptor
- LCA
- lithocholic acid
- TDCA
- taurodeoxycholic acid
- TβMCA
- tauro-β-muricholic acid
- CA
- cholic acid
- TCA
- taurocholic acid
- TCDCA
- taurochenodeoxycholic acid
- IBABP
- ileal bile acid binding protein
- OSTα/β
- organic solute transporter α/β
- BBM
- brush border membrane
- qPCR
- quantitative polymerase chain reaction
- HPLC
- high-performance liquid chromatography.
- Received June 22, 2010.
- Accepted September 29, 2010.
- Copyright © 2011 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}