


Abstract
There is a need to better understand the mechanism of airway hyper-reactivity, a key feature of asthma. Evidence suggests that sphingosine-1-phosphate (S1P) could be a major player in this phenomenon. The purpose of this work was to define the S1P receptor responsible for this phenomenon. We have studied, in the rat, the effect of two S1P synthetic receptor ligands, 2-amino-2-[2-(4-octylphenyl)ethyl]propane-1,3-diol (FTY720) (which in its phosphorylated form is a potent agonist at each S1P receptor except S1P2) and 3-[[2-[4-phenyl-3-(trifluoromethyl)phenyl]-1-benzothiophen-5-yl]methylamino]propanoic acid (AUY954) (a selective S1P1 agonist) on lung function in vivo. This was complemented by in vitro studies using isolated trachea from the rat, the S1P3 receptor-deficient mouse, and its wild-type counterpart. After oral administration, FTY720 induced a generalized airway hyper-reactivity to a range of contractile stimuli. This was observed as early as 1 h postdosing, lasted for at least 24 h, and was not subject to desensitization. In both rat and wild-type mouse isolated trachea, preincubation with the active phosphorylated metabolite of FTY720 induced hyper-responsiveness to 5-hydroxytryptamine. This effect was not seen in the isolated tracheas from S1P3 receptor-deficient mice. AUY954, did not mimic the effect of FTY720 either in vivo or in vitro. Our data are consistent with activation of the S1P pathway inducing a generalized airway hyper-reactivity in rats and mice that is mediated by the S1P3 receptor. S1P3 receptor antagonists might prove to be useful as new therapeutic strategies aimed at blocking the airway hyper-reactivity observed in asthma.
Introduction
Airway hyper-reactivity to bronchoconstrictor stimuli is a key feature of asthma. It is defined as an exaggerated sensitivity and reactivity of the airways to a variety of bronchoconstrictor stimuli. Several cell types and/or mediators have been implicated in the genesis of this phenomenon. However, the cellular and molecular mechanisms that are involved in airway hyper-reactivity remain unclear (Maddox and Schwartz, 2002). Although the inflammatory component of asthma in most patients is well controlled with anti-inflammatory drugs, such as inhaled steroids, airway hyper-reactivity is refractory to such treatments (Lundgren et al., 1988; Brusasco et al., 1998). There is therefore a need to better understand the mechanisms leading to the airway hyper-reactivity observed in asthma.
Sphingosine-1-phosphate (S1P) is a bioactive sphingolipid metabolite that is known to regulate diverse biological functions of many cell types, from proliferation and survival to migration and secretion. S1P exerts most of its actions by the activation of five G protein-coupled receptors designated S1P1–5 (Alexander et al., 2011). S1P has been demonstrated to be an important regulator of vascular and gastric smooth muscle contraction (Zhou and Murthy, 2004; Schuchardt et al., 2011). At the level of airway smooth muscle, S1P has been demonstrated to stimulate contraction of human airway isolated smooth muscle cells (Rosenfeldt et al., 2003) and guinea pig isolated trachea (Kume et al., 2007). In addition, it has been demonstrated that S1P causes increased sensitivity to methacholine-induced contraction of guinea pig isolated trachea (Kume et al., 2007) and mouse isolated bronchus and whole lung (Roviezzo et al., 2007, 2010). Although those studies strongly point toward a role for S1P as an important mediator that induces bronchoconstriction and airway hyper-reactivity in vitro or ex vivo, the same phenomenon has not yet been described in vivo, and none of the above studies have defined unequivocally the S1P receptors responsible for the effect. In the present study, we have used two synthetic S1P receptor agonists, 2-amino-2-[2-(4-octylphenyl)ethyl]propane-1,3-diol (FTY720), which in its phosphorylated form is a potent agonist at each S1P receptor except S1P2, and 3-[[2-[4-phenyl-3-(trifluoromethyl)phenyl]-1-benzothiophen-5-yl]methylamino]propanoic acid (AUY954) (a selective S1P1 receptor agonist), and S1P3 receptor-deficient mice in an attempt to characterize the S1P receptor subtype responsible for the in vivo and in vitro airway hyper-reactivity induced by activation of the S1P pathway. Taken together, our results suggest that, in rats and mice, activation of the S1P pathway induces airway hyper-reactivity that is mediated by activation of the S1P3 receptor.
Materials and Methods
Animals.
Male Brown Norway rats, weighing 200 to 300 g, were supplied by Charles River Laboratories (L'Abresle, France). Wild-type (C57BL/6) or S1P3 receptor-deficient mice (Ishii et al., 2001) were bred in house and used at the age of 11 to 15 weeks. Animals were kept at an ambient temperature of 22 ± 2°C under a 12-h normal-phase light/dark cycle. Food and drinking water were freely available. Animals were acclimatized for at least 7 days upon arrival in the laboratory before any experimental work began. All experiments were carried out with the approval of the Veterinary Authority of the City of Basel (Kantonales Veterinaeramt, Basel-Stadt).
Lung Function Measurements.
Measurement of airway resistance (RL) and cardiovascular parameters was carried out in anesthetized rats as described previously (Ellis et al., 2004). In brief, naive or ovalbumin-sensitized rats were anesthetized with pentothal (80 mg/kg), then given vecuronium bromide (12 mg/kg) to suppress natural breathing. Animals were ventilated via a tracheal cannula with a mixture of air and oxygen (1:1). Heparinized polyethylene catheters were inserted into the right carotid artery for recording blood pressure and heart rate and into the left jugular vein for bronchospasmogen administration. Body temperature was maintained at 37°C by means of a heated table. Airway resistance was calculated with a digital electronic respiratory analyzer (LFR; Mumed Systems Ltd., London, U.K.) for each respiratory cycle.
For the measurement of ovalbumin-induced bronchoconstriction, cumulative intravenous injections of ovalbumin (0.3, 1, and 3 mg/kg) were given to ovalbumin-sensitized animals (Ellis et al., 2004) with a 10-min interval between two doses. The measurement of responses to other bronchoconstrictor stimuli was performed in nonsensitized rats. The spasmogens (5-HT, methacholine, and adenosine) were injected intravenously with 5 min between doses, which was sufficient to allow the effects of the preceding dose to have returned to baseline. The time interval between each sequence of bronchoconstrictor agents was 15 min.
Drug Treatment for In Vivo Experiments.
FTY720 was dissolved in 0.9% saline and given either intravenously (bolus of 0.2 ml) or orally by gavage (2 ml/kg). AUY954, a selective S1P1 receptor agonist (Pan et al., 2006), was dissolved in 10% Tween 80 and 40% polyethylene glycol 200 in distilled water and given orally by gavage (2 ml/kg).
Isolated Tracheal Preparation.
Animals were killed by exposure to carbon dioxide. The trachea was removed, and two (mouse) or four (rat) segments, containing four cartilaginous rings, were cut. Each segment was opened longitudinally, opposite to the smooth muscle band, and set up for recording isotonic tension in 10-ml organ baths containing modified Krebs' solution (118 mM NaCl, 4.8 mM KCl, 1.2 mM MgSO4, 2.5 mM CaCl2, 1.2 mM KH2PO4, 25 mM NaHCO3, and 11 mM glucose) at 37°C bubbled with 95% O2/5% CO2. Resting tension was maintained at 1 and 0.5g for rat and mouse segments, respectively. After a stabilization period of 1 h, during which time the tissues were repeatedly washed, the experiment started as follows. For the rat segments, after addition of a supramaximal concentration of KCl (50 mM) and several washes, compounds were added for 30 min before the start of a concentration response curve to 5-HT (10−7 to 10−5 M). For the mouse segments, the same sequence was used without the initial addition of KCl. 2-Amino-2[2-(4-octylphenyl)ethyl]-1,3-propanediol mono dihydrogen phosphate ester (FTY720-P) and AUY954 were dissolved in 100% dimethyl sulfoxide (final bath concentration 0.2%).
Data Analysis.
In vivo data are expressed as variations from the baseline measurements. As an index of airway sensitivity to the contractile agents, the provocative dose of the contractile agents required to increase lung resistance by 100% from baseline (PC100) was calculated, and data were expressed as average and 95% confidence interval.
For isolated rat and mouse trachea, data are expressed as a percentage of the contraction obtained with 50 mM KCl or 10 μM 5-HT during the first concentration-response curve, respectively. All other data are presented as means ± S.E.M. Statistical significance, for the in vivo experiments, was established with Student's t test and the Hommel-Hochberg multiple comparison test by using the Excel version 8.0 software package (Microsoft, Redmond, WA). For the in vitro experiments, Student's t test for paired data was used, and the Hochberg correction was applied to adjust for multiple comparisons. Significance was assumed at the 5% probability level.
Materials.
Unless stated otherwise, all reagents were obtained from Sigma-Aldrich (Buchs, Switzerland). FTY720, FTY720-P, and AUY954 were synthesized by the Novartis Global Department of Chemistry (Basel, Switzerland). The S1P receptors' selectivity profile for the compounds, as determined by guanosine 5′-O-(3-thio) triphosphate binding assay, were as follows: FTY720-P, S1P1, 0.3 ± 0.1 nM; S1P2, >10,000 nM; S1P3, 3.1 ± 0.4 nM; S1P4, 0.6 ± 0.2 nM; and S1P5, 0.3 ± 0.2 nM; AUY954, S1P1, 1.2 ± 0.6 nM; S1P2, > 10,000 nM; S1P3, 1210 ± 127 nM; S1P4, > 1000 nM: and S1P5, 340 ± 80 nM (Pan et al., 2006).
Results
Intravenous Injection of FTY720 but Not AUY954 Induces a Delayed and Long-Lasting Bronchoconstriction That Is Reversed by Salbutamol.
When given as a bolus intravenous injection to the rat, 10 mg/kg FTY720 induced an immediate and short-lasting (less than 2 min) decrease in mean arterial blood pressure and heart rate. Although the hypotensive effect was transient, the bradychardia was long-lasting and still present at 1 h; at the latest time point studied (Fig. 1), approximately 10 min after the intravenous bolus injection of FTY720, airway resistance began to increase. The bronchoconstrictor response developed slowly in a dose-dependent manner and reached a maximal effect at approximately 50 min (Figs. 1 and 2). The effect on the airway resistance, induced by 10 mg/kg FTY720, was almost completely reversed by an intravenous bolus injection of 10 μg/kg salbutamol (inhibition of 95 ± 2%; n = 4) (Fig. 1), consistent with the increase in airway resistance induced by FTY720 being caused by a constrictor effect on the airway smooth muscle. In contrast, when 10 mg/kg AUY954 was given as a bolus intravenous injection no bronchoconstriction was observed (Figs. 1 and 2). However, AUY954 induced a bradycardia and a hypotensive effect with a similar magnitude and kinetic aspect to FTY720 (Fig. 1).

Representative traces for the effect of intravenous FTY720 (left) or AUY954 (right) on lung function and cardiovascular parameters in the Brown Norway rat. Anesthetized rats were given intravenously 10 mg/kg FTY720 or 10 mg/kg AUY954 at time 0, and airway resistance and cardiovascular parameter were measured after 60 min. For the animals given FTY720, the rats were injected with 10 μg/kg i.v. salbutamol 60 min after dosing.

Intravenously administered FTY720 but not AUY954 induces a delayed and sustained increase in RL in the Brown Norway rat in vivo. Anesthetized rats were treated intravenously with vehicle, increasing doses of FTY720, or 10 mg/kg AUY954, and airway resistance was monitored over time. Results are shown as changes from baseline values and expressed as means ± S.E.M. of three to six animals per group. *, p < 0.05, significant difference by comparison with vehicle-treated animals.
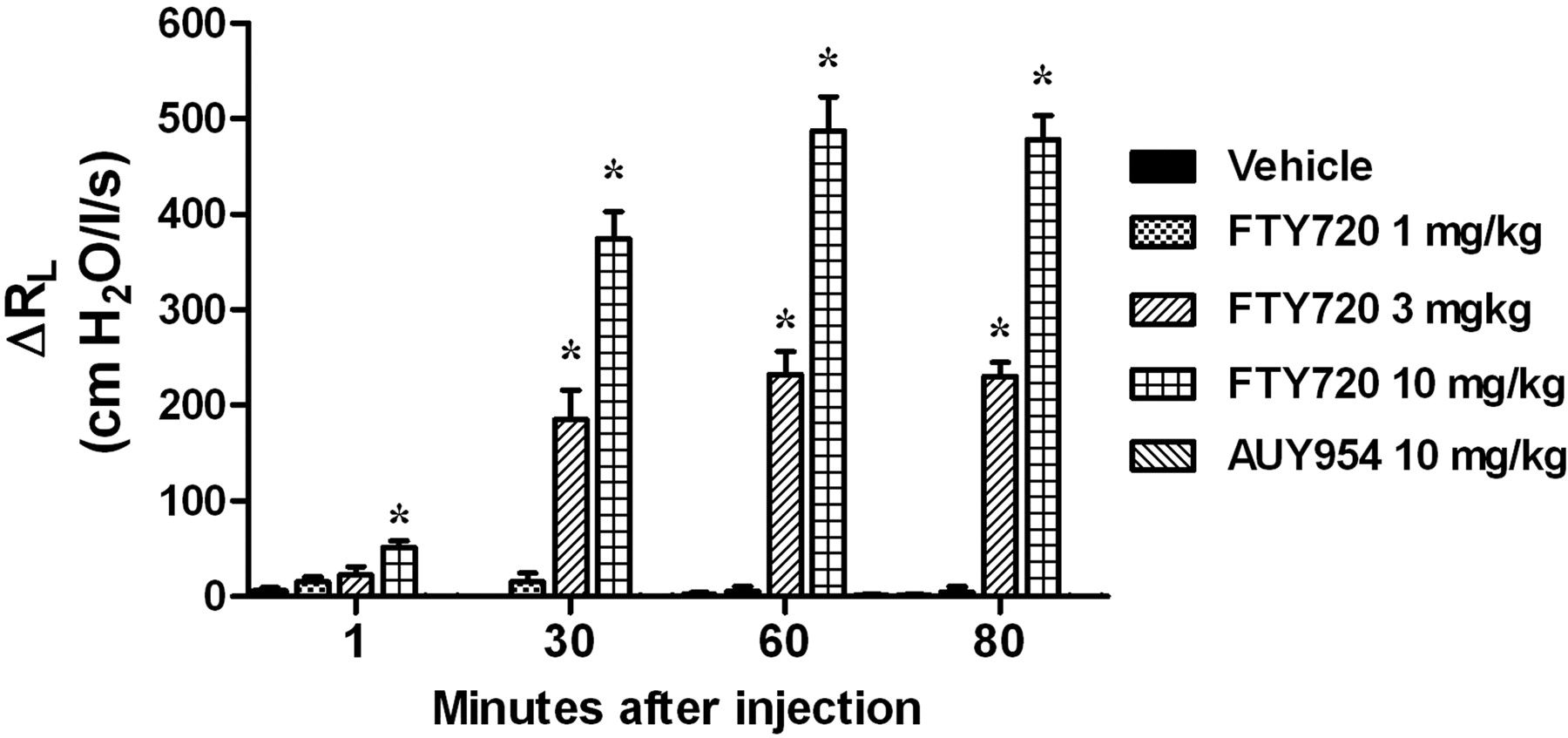
Oral Administration of FTY720 but Not AUY954 Induces Long-Lasting Generalized Airway Hyper-Reactivity That Does Not Show Tachyphylaxis.
When given orally FTY720 (0.1–10 mg/kg) did not induce bronchoconstriction but from the dose of 1 mg/kg long-lasting bradycardia was observed (Table 1). Three hours after oral dosing, FTY720 induced a dose-dependent increase in the airway responses to a range of spasmogens, including agonists that act directly on the smooth muscle (5-HT and methacholine), an agent that activates mast cells (adenosine), or an allergen challenge with ovalbumin (Fig. 3; Table 2). The increase in sensitivity was selective for the airways because the hypotensive response and effects on heart rate induced by all spasmogens tested were not altered by FTY720 (Fig. 3). In contrast to FTY720, 3 h after oral treatment with 30 mg/kg AUY954 no hyper-reactivity to 5-HT, methacholine, or adenosine was observed (Fig. 4), suggesting that the phenomenon observed with FTY720 was not mediated by the S1P1 receptor. Treatment with AUY954 did not modify the hypotensive response or the effects on heart rate induced by all spasmogens (data not shown).
Baseline values for heart rate, blood pressure, and airway resistance 3 h after oral administration of AUY954 or FTY720
Data are expressed as mean ± S.E.M. of the number of animals indicated in parentheses.

Orally administered FTY720 induces airway hyper-reactivity to a range of spasmogens in the Brown Norway rat in vivo. Increasing doses of FTY720 or its vehicle were administered orally to conscious rats. Three hours later, animals were anesthetized, and the effects of sequentially intravenously injected 5-HT, methacholine, and adenosine on airway resistance (top) blood pressure (middle), and heart rate (bottom) were monitored. In a separate experiment, ovalbumin-sensitized rats were challenged with increasing doses of ovalbumin injected intravenously. Results are shown as changes from baseline values and expressed as means ± S.E.M. of the number of animals indicated in parentheses. *, p < 0.05, significant difference by comparison with vehicle-treated animals.
PC100 values for the various contractile agents after oral administration of FTY720
Data are expressed as mean and 95% confidence interval of the number of animals indicated in Fig. 3.

Orally administered AUY954 does not induce airway hyper-reactivity in the Brown Norway rat. Conscious rats were treated orally with vehicle or 30 mg/kg AUY954. Three hours later, RL to spasmogens was recorded. Results are shown as change from baseline values and expressed as means ± S.E.M. of the number of animals indicated in parentheses.
To investigate the time course for the FTY720-induced increase in airway reactivity to spasmogens, animals were treated orally with 1 mg/kg FTY720, and airway reactivity was measured at different time points thereafter. The increase in airway reactivity was present as early as 1 h after dosing, reached its maximum at 3 h, and was still present 24 h postdosing (Fig. 5). This airway hyper-reactivity phenomenon was not subject to desensitization, because the effect was similar whether the animals were treated acutely with 1 mg/kg FTY720 or after 2 weeks of chronic treatment with 1 mg/kg FTY720, and the measurements were made 3 h after the last dose (Fig. 6).

Time course for oral FTY720-induced airway hyper-reactivity in the Brown Norway rat in vivo. Conscious rats were treated orally with 1 mg/kg FTY720, and RL to spasmogens was measured at different time points thereafter. Results are shown as changes from baseline values and expressed as means ± S.E.M. from six animals per group. *, p < 0.05, significant difference by comparison with vehicle-treated animals.

The effect of oral FTY720 is not subject to desensitization after chronic treatment in the Brown Norway rat. Conscious rats were treated orally with 1 mg/kg FTY720 for 15 days (chronic). As a control, a group of animals was treated with vehicle for 15 days (vehicle) or vehicle for 14 days followed by oral FTY720 (1 mg/kg) on day 15 (acute). RL to intravenously injected spasmogens was measured 3 h after the last treatment. Results are shown as changes from baseline values and expressed as means ± S.E.M. of the number of animals indicated in parentheses. *, p < 0.05, significant difference by comparison with vehicle-treated animals.
FTY720-P-Induced Hypersensitivity to the 5-HT-Induced Contraction in Rat Isolated Trachea.
Because the reversal of the bronchoconstriction observed after intravenous injection of FTY720 by salbutamol suggested a direct effect on smooth muscle, we used the rat isolated tracheal segment model to further study this effect. When applied to the organ bath, the active phosphorylated metabolite of FTY720 (FTY720-P) induced an immediate concentration-dependent contraction (tension in percentage of the response to 50 mM KCl; 0.1 μM, 1.9 ± 0.89; 1 μM, 27.9 ± 5.8; 10 μM, 98.9 ± 16.1). In addition, it increased concentration-dependently the 5-HT-induced contraction compared with control (Fig. 7, top left) but did not affect its potency with calculated pEC50 values ranging from 5.98 to 6.06 (Table 3). Under similar conditions, AUY954 (0.1–10 μM) had neither an intrinsic contractile effect nor any influence on the contractile response to 5-HT (Fig. 7, top right; Table 3).

FTY720-P-induced hypersensitivity to 5-HT in the rat and mouse isolated trachea. Vehicle or increasing concentrations of FTY720-P or AUY954 were incubated for 30 min before the start of the 5-HT cumulative concentration response. For the rat, the 5-HT concentration-response curves in the presence of FTY720 were expressed as increases over the FTY720 effect. Results are expressed as means ± S.E.M. of the number of segments isolated from the different animals indicated in Table 3.
5-HT pEC50 values in the rat and mouse isolated trachea preparations
Data are expressed as mean ± S.E.M. of the number of animals indicated in parentheses.
Effect of FTY720-P on 5-HT-Induced Contraction of Isolated Mouse Trachea.
Because of the lack of selective antagonists for the S1P3 receptor, we used the S1P3 receptor-deficient mouse to study the possible involvement of this receptor in the phenomenon observed. In contrast to the rat isolated trachea, FTY720-P had no intrinsic contractile effect in the mouse trachea isolated from wild-type animals. However, as observed in the rat, FTY720-P, from a concentration of 1 μM, induced an increase in the contractile response to 5-HT in the isolated trachea from wild-type mice (Fig. 7, middle left). When applied under the same conditions to tracheal segments isolated from S1P3 receptor-deficient mice, FTY720-P did not induce hypersensitivity to 5-HT (Fig. 7, bottom left). The 5-HT potency was not different compared with segments from wild-type and S1P3 receptor-deficient mice and was also not affected by FTY720-P as indicated by the pEC50 values ranging from 5.91 to 6.04 and 5.71 to 6.07 for the wild-type and S1P3 receptor-deficient animals, respectively (Table 3). AUY954 (0.1–10 μM) when applied to tracheal segments isolated from rat (Fig. 7, top right), wild-type mice (Fig. 7, middle right), or S1P3 receptor-deficient mice (Fig. 7, bottom right) did not have an intrinsic contractile effect or influence the contractile response to 5-HT (Table 3).
Discussion
Airway hyper-reactivity is a prominent feature of human asthma (Maddox and Schwartz, 2002) that is insensitive to currently available treatments (Lundgren et al., 1988; Brusasco et al., 1998). A significant body of data now indicates that S1P has proinflammatory effects, and studies have demonstrated that this lipid could be involved not only in the inflammatory reaction seen in asthmatic patients (Nixon, 2009) but also could play a role in airway hyper-reactivity.
The role of endogenously produced S1P has been assessed in mouse models of asthma, and in all cases the airway hyper-reactivity was abrogated (Lai et al., 2008; Nishiuma et al., 2008; Haberberger et al., 2009; Chiba et al., 2010b). These data suggest that the increased S1P levels seen in the airways of asthmatic patients after provocation with allergen (Ammit et al., 2001) could drive the airway hyper-reactivity associated with this disease. There can be little doubt that the above studies establish a role for S1P pathways in the airway hyper-reactivity induced by allergen challenge in mice. Our data extend this phenomenon to the rat and demonstrate for the first time that airway hyper-reactivity to a range of spasmogens can be induced in naive animals after oral administration of FTY720, a nonselective S1P receptor agonist. This phenomenon lasted for at least 24 h and did not desensitize after chronic dosing for 2 weeks.
In the present study, using synthetic S1P receptor ligands, we have attempted to define the receptor responsible for the S1P-induced airway hyper-reactivity in the rat. Although it has been previously reported that a single subcutaneous injection of S1P was able to induce an increase in cholinergic reactivity of mouse bronchial tissue in vitro (Roviezzo et al., 2010), in the rat we could not see any effect of S1P itself either using a bolus intravenous injection or an infusion (data not shown). This discrepancy is hard to explain, but seems probably to have a pharmacokinetic explanation. Thus, it is known that levels of S1P in most tissues are low, probably because of the metabolic activity of S1P lyase (Schwab et al., 2005), whereas the concentration of S1P in plasma is in the micromolar range (Pappu et al., 2007). Taking these points into account, we do not find it surprising that intravenous administration of S1P has no effect in our model. Because we could not see any in vivo effect using S1P, we used FTY720, a stable synthetic analog of S1P that once phosphorylated in vivo acts as an agonist at each of the S1P receptors except S1P2 (Brinkmann et al., 2002). In contrast to S1P, it has been shown that after oral and intravenous administration FTY720 distributes extensively to various tissues including lungs (Meno-Tetang et al., 2006). Using this agonist, we have shown that after intravenous injection, the compound induced a remarkable, slowly developing bronchoconstriction. When given orally, the intrinsic bronchoconstrictor effect was not observed. However, a generalized airway hyper-reactivity, to a range of contractile stimuli, was observed as early as 1 h after oral dosing. The fact that FTY720 induces bronchoconstriction only when it is given intravenously is a puzzling observation. When dosed to the rat, FTY720 is extensively converted to its active metabolite, FTY720-P, and this has been demonstrated to take place in a wide range of tissues (Billich et al., 2003). We speculate that the lack of bronchoconstriction after oral dosing of FTY720 is related to its peak exposure. Indeed, in the rat intravenous injection of FTY720 gives a high systemic exposure and a rapid diffusion into tissues including the lung. In contrast, when given orally the absorption into tissues is slow and delayed with a very low systemic clearance (Meno-Tetang et al., 2006). Therefore, the peak lung concentration after oral dosing of FTY720 might not be high enough to induce bronchoconstriction, but its very long systemic half-life together with its extensive distribution into the lung might account for the airway hyper-reactivity observed. Indeed, after oral dosing, it takes approximately 3 h for FTY720 to fully distribute to the lung, which is the time needed to observed the maximal airway hyper-reactivity (Fig. 5); whereas the lung diffusion is much faster after intravenous dosing (Meno-Tetang et al., 2006).
To obtain more information on the S1P receptors responsible for airway hyper-reactivity, we used AUY954, a selective S1P1 receptor agonist (Pan et al., 2006; Schuchardt et al., 2011). It was important to establish that the intervention with AUY954 was truly a consequence of a high degree of selectivity for the S1P1 receptor at the dose level used. That this was the case is suggested by the fact that AUY954 did not mimic the effect of FTY720 in vivo in the present study, whereas the compound did block allograft rejection in a rat transplantation model at doses similar to FTY720 (Pan et al., 2006). In addition, in the present experiments the compound did not mimic the effects of FTY720-P in the rat and mouse isolated trachea. These results suggest strongly that the S1P1 receptor does not play a role in the induction of airway hyper-reactivity. The fact that the airway hyper-reactivity induced by FTY720, which has negligible affinity for the S1P2 receptor, in vivo, was also seen on the isolated trachea in vitro and mimics the effect observed with S1P in the mouse isolated bronchi (Roviezzo et al., 2007) and in guinea pig isolated trachea (Kume et al., 2007) essentially rules out that the S1P2 receptor plays a role in this phenomenon.
A previous study, in the mouse isolated bronchus, concluded that the S1P3 receptor was not responsible for the potentiating effect of S1P on acetylcholine-induced contraction (Roviezzo et al., 2007). Nonetheless, this conclusion has to be challenged by the fact that it was based on the ineffectiveness of suramin and 2-undecyl-thiazolidine-4-carboxylic acid (BML-241) to inhibit the S1P-driven increase in sensitivity to acetylcholine. However, suramin is a nonselective anionic polycyclic dye that blocks many receptor/ligand interactions, and BML-241 has been shown not to be a S1P3 antagonist but rather a nonselective inhibitor of increases in intracellular calcium (Jongsma et al., 2006). In contrast, our data showing that FTY720-P-induced hyper-reactivity to 5-HT is abrogated in tracheal segments isolated from S1P3 receptor-deficient mouse provides powerful evidence that this phenomenon is mediated by the S1P3 receptor in this species.
In contrast to previous reports describing an intrinsic contractile effect of S1P in human airway smooth cells (Rosenfeldt et al., 2003) and guinea pig isolated trachea (Kume et al., 2007) and an intrinsic contractile effect of FTY720-P in isolated rat trachea (this study), we could not demonstrate such an effect in the mouse isolated trachea with FTY720-P. Our observation is in line with the lack of effect of S1P reported in the isolated bronchus from naive mice (Roviezzo et al., 2007).
We did not directly investigate the mechanism of FTY720-induced airway hyper-reactivity in vivo. However, many different cell types that are responsive to S1P could explain the in vivo airway hyper-reactivity (Ryan and Spiegel, 2008). We believe that the data of the present study strongly suggest that this phenomenon is driven by a direct effect on the bronchial smooth muscle that has been shown to express the S1P3 receptor (Roviezzo et al., 2007; Chiba et al., 2010a). In support of this, the response observed in vivo could be abrogated by salbutamol, a recognized bronchodilator. Furthermore, in vivo airway hyper-reactivity of similar magnitude was observed for a mechanistically diverse range of broncho spasmogens, including agonists that act directly at the levels of smooth muscle (i.e., 5-HT and methacholine), an agent that activate mast cells (i.e., adenosine), or more complex challenges such as ovalbumin. In addition, the in vivo phenomenon could be reproduced in the mouse and rat isolated trachea. All of these observations point toward a crucial role for the smooth muscle in the airway hyper-reactivity induced by activation of the S1P pathway. Although we did not attempt to delineate the intracellular pathways involved in airway hyper-reactivity, we feel that the downstream mechanism of action for S1P-induced contraction and/or airway hyper-reactivity in various airway smooth muscle preparations, including human (Rosenfeldt et al., 2003) and guinea pig (Kume et al., 2007), has been published numerous times and shown to involve the activation of the Rho kinase.
In conclusion, evidence is presented in this article that the activation of the S1P pathway, in rats and mice, induces a generalized increase in airway reactivity in vivo and in vitro that is mediated by the S1P3 receptor. It bears emphasis that the long duration of action of the airway hyper-reactivity induced by FTY720 and the fact that it does not desensitize are features that are characteristic of human asthma. It follows, therefore, that selective antagonists at the S1P3 receptor could help to elucidate the mechanism of this phenomenon and possibly facilitate the development of new therapeutic strategies aimed at suppressing the airway hyper-reactivity observed in conditions such as asthma.
Authorship Contributions
Participated in research design: Trifilieff and Fozard.
Performed data analysis: Trifilieff and Fozard.
Wrote or contributed to the writing of the manuscript: Trifilieff and Fozard.
Acknowledgments
We thank Francois Baur and Emilie Renard for technical expertise and Lazzaro Mazzoni for many helpful discussions.
Footnotes
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
ABBREVIATIONS:
- S1P
- sphingosine-1-phosphate
- RL
- airway resistance
- 5-HT
- 5-hydroxytryptamine
- FTY720
- 2-amino-2-[2-(4-octylphenyl)ethyl]propane-1,3-diol
- FTY720-P
- 2-amino-2[2-(4-octylphenyl)ethyl]-1,3-propanediol mono dihydrogen phosphate ester
- AUY954
- 3-[[2-[4-phenyl-3-(trifluoromethyl)phenyl]-1-benzothiophen-5-yl]methylamino]propanoic acid
- BML-241
- 2-undecyl-thiazolidine-4-carboxylic acid.
- Received January 6, 2012.
- Accepted May 7, 2012.
- Copyright © 2012 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}