


Abstract
Previous studies showed that human airway smooth muscle (HASM) cells treated with lysophosphatidic acid (LPA), a pertussis toxin (PTX)-sensitive G protein-coupled (GPC) mitogen, simultaneously with epidermal growth factor (EGF), a receptor tyrosine kinase (RTK) mitogen, exhibit markedly synergistic stimulation of mitogenesis. We now show that the RTK mitogens basic fibroblast growth factor, insulin-like growth factor-1, insulin, platelet-derived growth factor-AA, and platelet-derived growth factor-BB, as well as transforming growth factor-β, all induced synergistic stimulation of mitogenesis in the presence of LPA. The PTX-sensitive GPC mitogens carbachol and endothelin-1 and the PTX-insensitive GPC mitogens sphingosine-1-phosphate and thrombin exhibited synergistic stimulation together with EGF. Several RTK-RTK growth factor pairs and GPC-GPC mitogen pairs were also synergistic. HASM cells showed synergistic responses to serum plus EGF but not to serum plus LPA. Testing various other cell types showed that synergism between LPA and EGF occurred in other smooth muscle cells because both vascular smooth muscle cells and mesangial cells exhibited synergism. Additionally, human fetal lung fibroblasts also showed striking synergism. These results indicate that HASM cells can respond synergistically to a wide variety of mitogen combinations and that this synergism is a feature shared with other contractile cell types.
Lysophosphatidic acid (LPA) is a simple endogenous phospholipid growth mediator (Jalink et al., 1994). Release from activated platelets accounts for its presence in serum at the relatively high concentrations of 2 to 20 μM (Moolenaar, 1995). LPA mediates its effects by activating G protein-coupled (GPC) receptors; at least three GPC receptors mediating effects of LPA have recently been cloned (Goetzl and An, 1998; Bandoh et al., 1999; Im et al., 2000). Studies of signaling pathways activated by these receptors have suggested that LPA receptors couple to Gi, Gq, and G12 (An et al., 1998; Fukushima et al., 1998;Bandoh et al., 1999; Im et al., 2000). In contrast, epidermal growth factor (EGF) is a peptide growth factor that mediates its effects by activating a receptor tyrosine kinase (RTK). Thus, the two mitogens LPA and EGF represent different classes of mitogens, the GPC mitogens and the RTK mitogens, respectively. Previously our laboratory showed that LPA plus EGF synergistically stimulated mitogenesis of human airway smooth muscle (HASM) cells, measured by both [3H]thymidine incorporation assays and cell counting (Cerutis et al., 1997). One purpose of the current studies was to determine the specificity of synergistic stimulation of HASM cell mitogenesis for different GPC and RTK mitogens.
A second goal was to determine the extent to which synergism occurs in other cell types. LPA has been reported to exhibit synergistic stimulation of mitogenesis in other contractile cells from other species. In rat aortic vascular smooth muscle cells, LPA plus either EGF or fibroblast growth factor (FGF) induced synergistic stimulation of mitogenesis; in this system, stimulation by LPA plus platelet-derived growth factor (PDGF) was additive (Tokumura et al., 1994). In contrast, LPA plus PDGF-BB synergistically stimulated mitogenesis of rat mesangial cells (Inoue et al., 1997). Because of these studies, human aortic smooth muscle cells and human mesangial cells were included in our studies, as well as other noncontractile cell types, for comparison to HASM cells.
HASM cells provide a synthetic cell culture model of physiological airway smooth muscle responses (Hall and Kotlikoff, 1995; Halayko et al., 1996). As such, studies of HASM cell mitogenesis are relevant to pulmonary disease states showing hypertrophic and/or hyperplastic airway smooth muscle, such as asthma, chronic bronchitis, and bronchiolitis obliterans (Hirst, 1996). In addition, the studies presented herein address the responsiveness of HASM cells and other cell types to simultaneous exposure to multiple growth factors, a situation undoubtedly more common in vivo than the isolated responses to individual mitogens more typically studied, and therefore likely to be of greater physiological relevance.
Materials and Methods
Reagents.
Dulbecco's modified Eagle's medium (DMEM), fetal bovine serum (FBS), basic FGF (bFGF), and platelet-derived growth factor (PDGF)-AA and -BB were obtained from Life Technologies (Grand Island, NY). LPA was purchased from Avanti Polar Lipids (Alabaster, AL), EGF was from Biosource International (Camarillo, CA), and transforming growth factor-β1 (TGF-β) was from R&D Systems (Minneapolis, MN). Pertussis toxin (PTX) was obtained from List Biologicals (Campbell, CA) and [3H]thymidine was from NEN (Boston, MA); insulin-like growth factor-1 (IGF-1) was a generous gift from Dr. Richard McDonald (University of Nebraska Medical Center, Omaha, NE). Other chemicals were obtained from Sigma (St. Louis, MO).
Cell Culture.
HASM cells previously isolated from human trachea by enzymatic dissociation were kindly provided by Dr. Michael Kotlikoff (University of Pennsylvania, Philadelphia, PA) (Hall and Kotlikoff, 1995). Human fetal lung fibroblasts (HFL-1 cells) were originally obtained from the American Type Culture Collection (Manassas, VA). Human foreskin fibroblasts (HFF cells) isolated from human foreskin explants (Freshney, 1987) were provided by Dr. Roseann Vorce (University of Nebraska Medical Center). Human vascular smooth muscle (VSM) cells obtained from a proximal aortic surgical specimen were provided by Dr. B. Timothy Baxter (University of Nebraska Medical Center) (Halloran et al., 1995). Human mesangial cells (HMCs) originally isolated by Dr. Hanna Abboud (University of Texas Health Sciences Center, San Antonio, TX) (Shultz et al., 1988) were provided by Dr. Steve Sansom (University of Nebraska Medical Center). All cells were cultured in high-glucose (4.5 g/l) DMEM with 10% FBS at 37°C in a humidified 5% CO2 incubator, except for 1321N1 human astrocytoma cells, which were cultured in low-glucose (1.0 g/l) DMEM with 5% FBS at 37°C in a humidified 8% CO2 incubator (Kreps et al., 1993).
[3H]Thymidine Incorporation Assays.
HASM cells were plated at 15,000 cells/well in 24-well plates. HFL-1 and HFF cells also were plated at 15,000 cells/well; HMC and VSM cells were plated at 25,000 cells/well, and 1321N1 cells were plated at 50,000 cells/well. 1321N1 cells were starved 2 days after plating; all other cells were grown to confluence before starvation. All cells were starved in serum-free medium for 24 h followed by treatment with mitogens for 24 h. Because LPA was reconstituted at 10 mM in 0.25% essentially fatty acid-free BSA, the appropriate dilution of BSA was used as a vehicle control. For experiments with PTX, cells were starved in serum-free medium containing 100 ng/ml PTX; 100 ng/ml PTX also was included in the stimulation medium. [3H]Thymidine (2 μCi/ml) was added for the final 2 h of mitogen treatment. Cells were then washed once with PBS and twice with 10% trichloroacetic acid (one 10-min incubation followed by one wash). Precipitated DNA was dissolved with 0.2 N NaOH and [3H]thymidine incorporation was quantitated by scintillation counting.
Flow Cytometric Analysis of Cell Cycle.
HASM cells were plated at 100,000 cells/60-mm dish. Once confluent, cells were starved in serum-free medium for 24 h before treatment with mitogens for 24 h. After treatment, cells were washed once with PBS and removed from the dish by trypsinization. Further trypsinization was blocked by adding serum-containing medium. Cells were pelleted by centrifugation for 10 min at 200g in a Beckman GPKR centrifuge. Cells were then washed once with serum-containing medium and once with PBS before being resuspended in Vindelov's reagent [75 μg/ml propidium iodide, 3.5 U/ml ribonuclease A, 0.1% Nonidet P-40 in Tris-buffered saline, pH 7.6 (3.5 mM Tris, 10 mM NaCl)]. Flow cytometric analysis was performed with a Becton Dickinson (San Jose, CA) FACSCalibur flow cytometer, modeling 10,000 events, and data were analyzed with ModFit LT software from Verity (Topsham, ME).
Data Analysis.
Data are presented as means ± S.E. Synergism was defined with the summation method described by Berenbaum (1989). If two mitogens are using two completely separate pathways, the stimulation seen with the two mitogens together should be the sum of the individual effects (the “zero interaction” or additive state). Synergism is thus defined as stimulation clearly greater than the sum of the stimulations seen with each individual mitogen, representing interaction between the mitogenic signaling pathways activated by each individual mitogen. For the analyses presented herein, we have chosen to define synergism as stimulation greater than or equal to 150% of the sum of the stimulations seen with each individual mitogen (denoted “SYN”). Other combinations were either slightly greater than additive, but not 150% of the sum (denoted “>ADD”); purely additive (“ADD”); or less than additive (“<ADD”).
Results
Synergistic Mitogenesis by LPA plus EGF Assessed by DNA Synthesis and S Phase Progression
Treating HASM cells with LPA plus EGF yields synergistic stimulation of mitogenesis as measured by [3H]thymidine incorporation assays (Fig.1A), as previously described (Cerutis et al., 1997). Stimulation with LPA alone produced 13.2 ± 2.9-fold increase in [3H]thymidine incorporation, EGF alone showed 6.1 ± 0.8-fold increase, and LPA plus EGF yielded 54.0 ± 11.9-fold increase. Zero interaction between pathways would predict that the stimulation by LPA plus EGF should be additive of individual stimulation levels; in this case LPA (13-fold) plus EGF (6-fold) should yield an increase of approximately 19-fold, not the 54-fold stimulation observed, clearly indicating synergism. Similar results were seen with flow cytometric analysis (Fig. 1B), where the percentage of cells in S phase was 1.6 ± 0.4% for control cells, 8.4 ± 1.2% for LPA alone, 3.8 ± 0.8% for EGF alone, and 26.8 ± 1.0% for LPA plus EGF. These values showed that the percentage of cells in S phase after treatment with LPA plus EGF was greater than the sum of the individual values, in agreement with the synergism observed with [3H]thymidine incorporation. Flow cytometric analysis of cells in S phase thus confirmed that the [3H]thymidine incorporation assay results reflect new DNA synthesis, not DNA repair or another artifact, and corroborated previous data with cell counting (Cerutis et al., 1997). All subsequent experiments presented below used the [3H]thymidine incorporation assay to compare stimulation due to a variety of mitogens, and the results of these [3H]thymidine incorporation assays were analyzed as described in Fig. 1A.

Mitogenesis and S phase progression of HASM cells treated with LPA plus EGF. After serum starvation for 24 h, HASM cells were treated for 24 h with 10 μM LPA, 60 ng/ml EGF, the combination of LPA plus EGF, or the vehicle BSA. A, incorporation of [3H]thymidine during the last 2 h of mitogen treatment was measured to monitor DNA synthesis. Values are presented as fold stimulation of treated cells over control cells and are the means ± S.E. of three experiments performed in parallel with the cells used for flow cytometry (B). Each experiment was performed in duplicate or triplicate (total n = 8). B, HASM cells treated in parallel with those from A were treated with propidium iodide to stain DNA and analyzed by flow cytometry. Values shown are the percentage of cells in S phase and are the means ± S.E. of three experiments (total n = 5).
Stimulation of HASM Cell Mitogenesis by LPA plus Various RTK Growth Factors
To investigate the specificity of the synergistic stimulation of HASM cell mitogenesis reported for LPA plus EGF, stimulation by a variety of RTK growth factors was compared with that by the prototype RTK growth factor EGF (Table 1). The RTK growth factors bFGF, IGF-1, insulin, PDGF-AA, and PDGF-BB were all effective mitogens for HASM cells, with PDGF-BB showing the greatest stimulation. In addition, the receptor serine/threonine kinase growth factor TGF-β also stimulated HASM cell mitogenesis. LPA exhibited synergism with bFGF, IGF-1, insulin, PDGF-AA, PDGF-BB, and TGF-β (Table 1).
Mitogenesis of HASM cells treated with LPA and various RTK growth factors
Stimulation of HASM Cell Mitogenesis by EGF plus Various GPC Mitogens
To investigate the specificity of mitogenic synergism for the GPC mitogen LPA, we compared stimulation by a variety of GPC mitogens to that by LPA (Table 2). Sphingosine-1-phosphate (S1P) and LPA are structurally similar and activate receptors in the same gene family. S1P also can stimulate proliferation (Goetzl and An, 1998), although in guinea pig airway smooth muscle cells, S1P was not mitogenic but did activate mitogen-activated protein kinase (Rakhit et al., 1999). We also included carbachol (CCh), endothelin-1 (ET-1) and thrombin in our panel of GPC mitogens because all were previously reported to be mitogenic in airway smooth muscle cells (Hirst, 1996). EGF exhibited synergism not only with LPA but also with the four other GPC mitogens tested (Table2). Stimulation by LPA, CCh, and ET-1 was markedly PTX sensitive (Fig.2; P < .05), indicating that these GPC mitogens signal primarily through Gi (or Go) proteins. In contrast, stimulation by S1P and thrombin was minimally PTX sensitive, suggesting that S1P and thrombin stimulate mitogenesis primarily through other G proteins. Because synergism was seen when cells were treated with EGF together with either PTX-sensitive or PTX-insensitive mitogens, synergism is not a phenomenon that relies exclusively on signaling by any specific G protein.
Mitogenesis of HASM cells treated with EGF and various GPC mitogens

PTX sensitivity of GPC mitogen responses. [3H]Thymidine incorporation assays were performed to assess the PTX sensitivity of HASM cell mitogenesis induced by the indicated GPC mitogens. Cells were starved and treated with serum-free medium containing 100 ng/ml PTX for 24 h before addition of mitogens. Cells were then treated for 24 h in the absence or presence of 10 μM LPA, 100 μM CCh, 100 nM ET-1, 10 μM S1P, or 2 U/ml thrombin (Thr), in the absence or continued presence of 100 ng/ml PTX. Values are means ± S.E., with the number of experiments, each performed in triplicate, indicated below each mitogen. Asterisks indicate that stimulation in the presence of PTX was significantly less than in the absence of PTX by pairedt test (P < .05).
Stimulation of HASM Cell Mitogenesis by Pairs of RTK Growth Factors
In analogous experiments, RTK growth factors were tested in combination with each other (Table 3). Three of the 13 combinations clearly did not show synergism: stimulation by EGF plus bFGF was the same as with bFGF alone, stimulation with IGF-1 plus insulin was similar to stimulation with either growth factor alone, and the combination of PDGF-AA plus PDGF-BB was only additive. The other combinations tested exhibited mitogenic stimulation greater than the sum of the stimulations with each agent individually; some were only slightly greater than additive, but four combinations (insulin plus EGF, insulin plus bFGF, PDGF-AA plus EGF, and PDGF-AA plus bFGF) exhibited stimulation ≥150% of the sum, thus qualifying as synergism.
Mitogenesis of HASM cells treated with two RTK growth factors
Stimulation of HASM Cell Mitogenesis by Pairs of GPC Mitogens
Because synergism is not exclusively associated with either PTX-sensitive or PTX-insensitive mitogens, we examined whether various GPC mitogens could synergize with each other, and if so, whether synergism would occur only between mitogens showing different sensitivity to PTX treatment (Table 4). Of the three pairs of PTX-sensitive mitogens, only CCh plus ET-1 exhibited synergism; the combinations CCh plus LPA and ET-1 plus LPA were almost exactly additive. In assays pairing PTX-sensitive with PTX-insensitive mitogens, CCh plus S1P and CCh plus thrombin were synergistic. The combinations of LPA plus S1P and LPA plus thrombin were greater than additive but not ≥150% of the sum. In contrast, ET-1 plus S1P and ET-1 plus thrombin were slightly less than additive and thus clearly not synergistic. S1P plus thrombin, the only combination of two PTX-insensitive mitogens, showed stimulation similar to that seen with thrombin alone and thus was not even additive.
Mitogenesis of HASM cells treated with two GPC mitogens
Stimulation of HASM Cell Mitogenesis by LPA plus EGF in the Absence or Presence of Serum
To investigate the upper limits of mitogenic stimulation, HASM cells were treated with LPA, EGF, or LPA plus EGF, in the absence or presence of 1.5 or 5% FBS (Fig. 3). As reported previously, 10 μM LPA and 1.5% FBS yielded similar responses (Cerutis et al., 1997). When HASM cells were treated with LPA plus 1.5 or 5% FBS, stimulations were additive but not synergistic. For example, LPA treatment alone yielded 10-fold stimulation, 1.5% FBS yielded 7-fold stimulation, and together LPA plus 1.5% FBS produced 18-fold stimulation. In contrast, HASM cells treated with EGF plus 1.5 or 5% FBS showed synergistic mitogenic responses. EGF treatment alone yielded 7-fold stimulation, 1.5% FBS yielded 7-fold stimulation, and together EGF plus 1.5% FBS produced 36-fold stimulation of HASM cell mitogenesis.
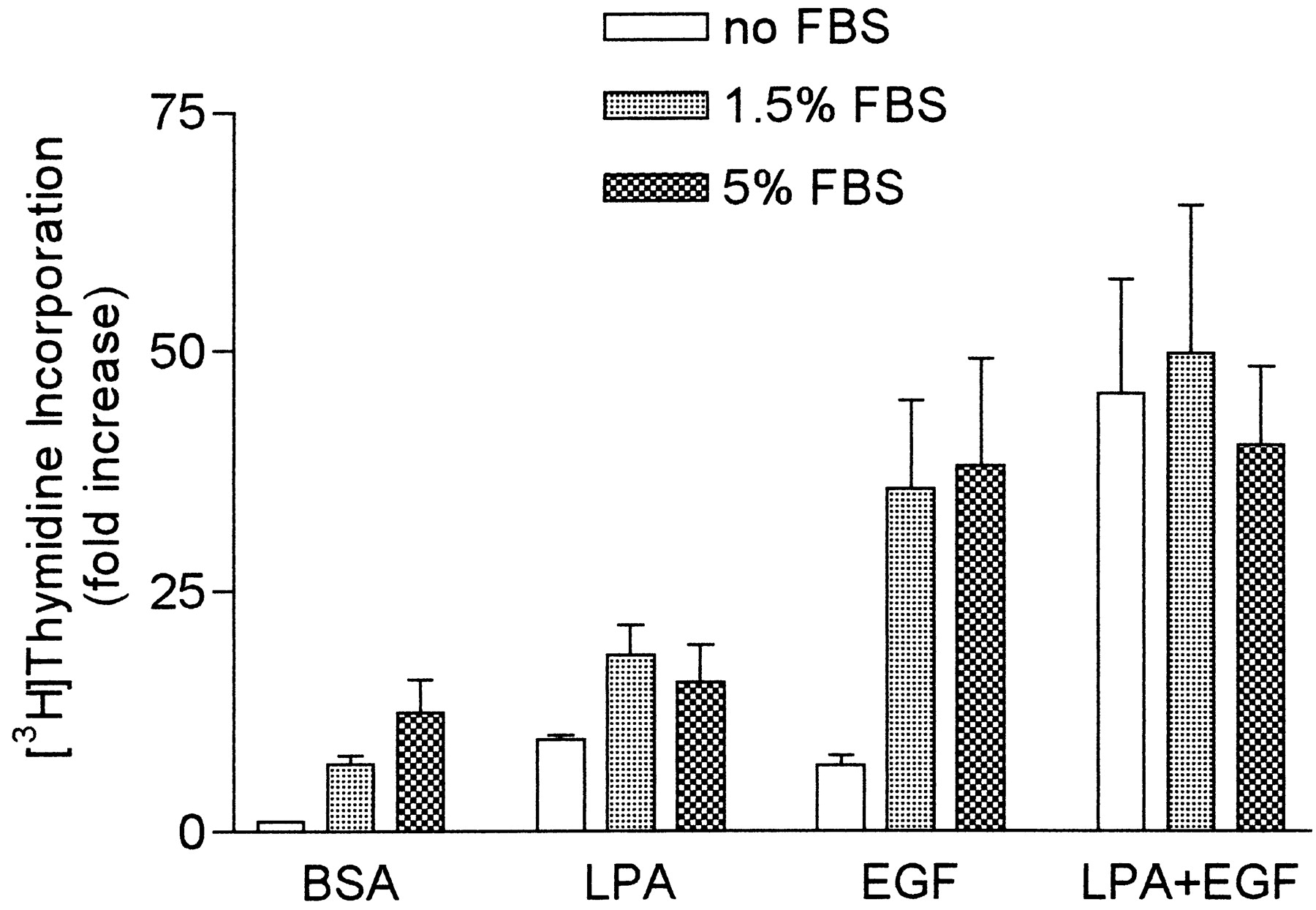
Mitogenic stimulation of HASM cells by FBS together with LPA, EGF, or LPA plus EGF. [3H]Thymidine incorporation assays were performed with HASM cells treated in the absence or presence of either 1.5 or 5% FBS together with 10 μM LPA, 60 ng/ml EGF, LPA plus EGF, or the vehicle BSA. Values shown are the means ± S.E. of three experiments, each performed in triplicate.
HASM cells treated with either 1.5 or 5% FBS together with LPA plus EGF did not exhibit mitogenic stimulation greater than that seen with LPA plus EGF in the absence of serum. In addition, for both LPA and EGF used individually, combination with 5% FBS instead of 1.5% FBS did not generate greater mitogenic stimulation.
Occurrence of Synergism with LPA plus EGF in Other Cell Types
To investigate the specificity of synergism for the HASM cell type, various other cell types were tested for synergistic mitogenic responses to LPA plus EGF and other RTK mitogens (Table5).
Mitogenesis of multiple cell types treated with LPA plus various RTKs
Cell Types Exhibiting Synergism.
Human aortic VSM cells showed mitogenic responses to LPA, EGF, bFGF, and PDGF-BB. LPA plus EGF resulted in mitogenesis moderately greater than the sum of the individual responses, whereas LPA plus PDGF-BB yielded clear synergism. In contrast, the response to LPA plus bFGF was similar to the response to LPA alone; however, bFGF alone was a weak mitogen for VSM cells. HMCs showed mitogenic responses to LPA, EGF, bFGF, and PDGF-BB. When treated with combinations of LPA plus either EGF, bFGF, or PDGF-BB, HMC mitogenic responses were clearly synergistic. HFL-1 cells showed mitogenic responses to LPA, EGF, and bFGF, and both LPA plus EGF and LPA plus bFGF exhibited synergism.
Cell Types Not Showing Synergism.
HFF cells responded to LPA, EGF, and bFGF; responses to LPA plus EGF or bFGF were additive but not synergistic. The human astrocytoma cell line 1321N1 exhibited mitogenic responses to both LPA and EGF but not synergism or even additivity when treated with LPA and EGF together.
Discussion
The purpose of these studies was to investigate the extent of occurrence of the mitogenic synergism previously reported for LPA plus EGF in HASM cells, both in terms of other mitogen combinations and other cell types. The results suggest that synergism can be induced by a diverse group of mitogen combinations. LPA is able to synergize with a variety of RTK mitogens, and EGF synergizes with both PTX-sensitive and PTX-insensitive GPC mitogens. RTK mitogens can synergize with other RTK mitogens, and GPC mitogens can synergize with other GPC mitogens. These data indicate that there are likely to be multiple mechanisms underlying synergism because some combinations of each class of mitogen are able to exhibit mitogenic synergism. However, some specific combinations did not exhibit synergism. Notably, LPA did not synergize with either of the two PTX-sensitive mitogens, CCh and ET-1. There did appear to be some interaction with other GPC mitogens because LPA plus either S1P or thrombin yielded greater than additive results, although stimulation did not achieve ≥150% of the sum, our criterion for synergism. Interestingly, the only combination of two PTX-insensitive GPC mitogens tested was not even additive, let alone synergistic. Thus, the ability of synergism to occur between GPC mitogens activating the same G protein appears limited.
In our initial documentation of LPA plus EGF synergism, we showed that serum stimulation of HASM cell mitogenesis is highly PTX sensitive (Cerutis et al., 1997). This suggested that Gi-coupled mitogens are key mitogenic components of serum. Interestingly, in the current report, HASM cells showed synergistic responses to treatment with serum plus EGF, but treatment with LPA plus serum was not synergistic. Because the other studies reported herein showed that LPA enhanced stimulation by both PTX-insensitive mitogens (Table 4) and a wide variety of RTK mitogens (Table 1) but not by other PTX-sensitive mitogens (Table 4), the finding that LPA did not enhance serum stimulation supports the previous observation that Gi-coupled mitogens found in serum are important mediators of serum-induced mitogenesis in HASM cells.
Several previous reports of synergistic stimulation of mitogenesis have been from studies with smooth muscle cells. LPA was synergistic with EGF and FGF in rat aortic smooth muscle cells (Tokumura et al., 1994), and LPA plus PDGF-BB exhibited synergism in rat mesangial cells (Inoue et al., 1997). In addition, ET-1 was synergistic with PDGF-AA, PDGF-BB, bFGF and EGF in guinea pig airway smooth muscle cells (Fujitani and Bertrand, 1997). In HASM cells, ET-1 was reported to potentiate EGF stimulation of mitogenesis, although in that study ET-1 alone did not stimulate mitogenesis (Panettieri et al., 1996). The fact that all of these studies showing synergism were in cells of smooth muscle origin, either vascular smooth muscle, airway smooth muscle, or mesangial cells (Kreisberg and Karnovsky, 1983; Diamond and Karnovsky, 1988), suggested to us that synergism might be a phenomenon common to smooth muscle cells. Early studies of LPA signaling found that proliferation by LPA was not potentiated by insulin in Rat-1 fibroblasts (van Corven et al., 1989). The data in Table 5, all with human cells, confirm that synergism is indeed a property of multiple smooth muscle cell types. The first cell type that we had tested was the airway smooth muscle (HASM) cell, which then became the prototype; the additional studies presented herein show that mesangial cells and VSM cells exhibit a similar phenotype. Mesangial cells showed strongly synergistic responses to LPA plus EGF, LPA plus bFGF, and LPA plus PDGF-BB. VSM cells had a similar pattern, although the level of stimulation was not high enough to make a strong case for synergism except for the LPA plus PDGF-BB combination. In contrast, both HFF cells and 1321N1 human astrocytoma cells showed mitogenic responses to LPA and EGF individually but no synergism when treated with LPA plus EGF. The only other cell type that did exhibit synergism was HFL-1 cells, which showed strong synergism in response to LPA plus EGF and LPA plus bFGF. HFL-1 cells represent a heterogeneous cell population, including cells with a myofibroblast phenotype that express smooth muscle-specific actin (Schmitt-Graff et al., 1994; Kawamoto et al., 1997). Thus, HFL-1 cells may be similar in phenotype to vascular or airway smooth muscle cells, and synergism could be a feature of the highly responsive myofibroblast or “synthetic” smooth muscle cell phenotype.
Smooth muscle cells exhibit a spectrum of phenotypes from the purely “contractile” to the purely “synthetic”; grown in culture, synthetic smooth muscle cells have the selective advantage (Chamley-Campbell et al., 1979). Synthetic smooth muscle cells proliferate rapidly, respond to inflammatory mediators, and secrete cytokines (Hirst, 1996). Thus, the synthetic smooth muscle cell, whether it be of airway, vascular, or renal origin, is of particular physiological relevance in inflammatory disease states and hyperproliferative states. Our finding that a variety of smooth muscle cell types respond synergistically to a variety of RTK and GPC mitogens is likely to be especially relevant to the inflammatory environment. EGF, FGF, IGF-1, and PDGF are all secreted by macrophages and thus may be present in the inflammatory milieu (Hirst and Twort, 1992). LPA and PDGF are both secreted by platelets, again placing these mediators at the site of inflammation. When considering the role of inflammatory mediators in producing smooth muscle hyperplasia such as that seen in asthma (Hirst and Twort, 1992), it is imperative to consider the exposure of smooth muscle cells to the wide variety of mediators released in inflammation because the responses are not only robust but often synergistic. The synergistic mitogenic responses of smooth muscle cells shown in this study emphasize the importance of considering the interactions of multiple inflammatory and proliferative mediators, especially in understanding the interplay of inflammatory mediators implicated in disease states involving smooth muscle hyperplasia. Because synergism occurs not only for airway smooth muscle but also for vascular smooth muscle cells and mesangial cells, there are broad implications not only for asthma but also for atherosclerosis and glomerulosclerosis, all diseases marked by smooth muscle hyperplasia of various cell types.
Acknowledgments
We thank Charles Kuszynski at the University of Nebraska Medical Center Cell Analysis Facility for technical support. We also thank our University of Nebraska Medical Center colleagues Roseann Vorce, B. Timothy Baxter, Steve Sansom, Stephen Rennard, and Richard McDonald for cells and reagents.
Footnotes
-
Send reprint requests to: Myron L. Toews, Ph.D. Department of Pharmacology, 986260 Nebraska Medical Center, Omaha, NE 68198-6260. E-mail: mtoews{at}unmc.edu
-
↵1 This work was supported in part by research seed grants from the University of Nebraska Medical Center (to M.L.T.) and a University of Nebraska Medical Center McDonald Fellowship (to T.L.E).
- Abbreviations:
- LPA
- lysophosphatidic acid
- GPC
- G protein-coupled
- EGF
- epidermal growth factor
- RTK
- receptor tyrosine kinase
- HASM
- human airway smooth muscle
- FGF
- fibroblast growth factor
- PDGF
- platelet-derived growth factor
- DMEM
- Dulbecco's modified Eagle's medium
- FBS
- fetal bovine serum
- bFGF
- basic fibroblast growth factor
- TGF-β
- transforming growth factor-β
- PTX
- pertussis toxin
- IGF-1
- insulin-like growth factor-1
- HFL-1
- human fetal lung fibroblasts
- HFF
- human foreskin fibroblasts
- VSM
- vascular smooth muscle
- HMC
- human mesangial cell
- S1P
- sphingosine-1-phosphate
- CCh
- carbachol
- ET-1
- endothelin-1
- Received January 6, 2000.
- Accepted May 9, 2000.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}