


Abstract
Prostaglandin (PG) E2 induces dendritic cell maturation in cooperation with proinflammatory cytokines [such as tumor necrosis factor (TNF)-α and interleukin (IL)-1β]. To clarify the involvement of E-prostanoid (EP) receptors in the effect of prostaglandin E2 on human monocyte-derived dendritic cell (MoDC) maturation, we examined the effect of four types of EP receptor-selective agonists on MoDC maturation. PGE2 as well as 11,15-O-dimethyl prostaglandin (E2ONO-AE1-259-01) (EP2 receptor agonist) and ONO-AE1-329 (EP4 receptor agonist) concentration dependently enhanced the expression of CD80, CD86, CD83, and HLA-DR on MoDCs during maturation, especially in the presence of TNF-α, whereas 17S-2,5-ethano-6-oxo-17,20-dimethyl prostaglandin E1 (EP1 receptor agonist) and 16S-9-deoxy-9β-chloro-15-deoxy-16-hyfroxy-17,17-trimethylene-19,20-didehydro prostaglandin F2 (EP3 receptor agonist) showed no effect. The maximal effect of ONO-AE1-259-01 was higher than that of ONO-AE1-329; however, the stimulation with ONO-AE1-259-01 was less effective than that with PGE2. Simultaneous stimulation with both EP receptor agonists produced additive effects and 11-deoxy-PGE1 (EP2/EP4 receptor mixed agonist) mimicked the effects of PGE2. Dibutyryl cAMP mimicked the effects of PGE2, indicating the mediation of PGE2 action by cAMP. Matured MoDCs induced by PGE2 or EP2 and/or EP4 receptor agonists showed a decrease in lipopolysaccharide (LPS)-stimulated IL-12p70, IL-6, and IL-10 production. The coculture of naive T cells with matured MoDCs induced under different conditions showed that EP2/EP4-stimulated MoDCs preferentially induced alloresponsive helper T (Th)2 cells. Together, it was concluded that the cooperative stimulation of EP2 and EP4 receptor subtypes by PGE2 promoted MoDC maturation and inhibited LPS-induced cytokine production in MoDCs. The matured MoDCs under such conditions preferably induced Th2 polarization, indicating the importance of EP2 and EP4 receptors in the determination of Th1/Th2 development of naive T cells.
Dendritic cells are the most potent antigen-presenting cells and are highly responsive to inflammatory stimuli such as bacterial LPS and TNF-α (Steinman, 1991). The phenotypic and functional changes in DCs induced by these stimuli have often been referred to as DC maturation (Cella et al., 1997). DCs establish surveillance networks throughout body surfaces against pathogen entry (Banchereau and Steinman, 1998). DCs, which take up pathogens, develop their maturation processes including a decrease in antigen uptake by endocytosis (Sallusto et al., 1995), an increase in antigen-presenting ability, and up-regulation of the surface expression of major histocompatibility complex and costimulatory molecules during the migration from local tissues to regional lymph nodes where they prime naive antigen-specific T cells (Cella et al., 1997).
By the action of cyclooxygenase (COX), the precursor prostaglandin of PGE2 is synthesized from arachidonic acid liberated from membrane phospholipids by the action of phospholipase A2 (Smith et al., 1996). Two isoforms of COX exist. COX-1 is constitutively expressed on various cells and responsible for homeostasis, whereas COX-2 can be induced by several stimuli such as cytokines and mediators (Smith et al., 1996), playing important roles in PGE2 production in the case of inflammation. PGE2 stimulates four PGE2 receptor subtypes: EP1, EP2, EP3, and EP4 receptors. EP2 and EP4 receptors are coupled to a Gs protein and adenylate cyclase activation (Coleman et al., 1994). EP1 receptor is coupled to inositol phospholipid-related signal transduction (Okuda-Ashitaka et al., 1996) and increases the intracellular Ca2+ concentration (Narumiya et al., 1999). The major signaling pathway of the EP3 receptor is inhibition of adenylate cyclase via Gi, but plural EP3 splicing variants exhibit various signaling pathways (Namba et al.,1993).
PGE2 is produced by stromal cells and infiltrating mononuclear cells (Noguchi et al., 2000; Harizi et al., 2002). In an inflammatory environment, PGE2 induces vasodilation and promotes tissue edema in cooperation with other mediators. PGE2 directly inhibits Th1 cytokine production and induces Th2 subset development (Betz and Fox, 1991; Demeure et al., 1997). In monocytes/macrophages, PGE2 also acts as an immunomodulator, inhibiting IL-12 and TNF-α production and increasing IL-10 production (Scales et al., 1989; Betz and Fox, 1991; van der Pouw et al., 1995; Ikegami et al., 2001). Additionally, PGE2 together with TNF-α has been suggested to be a maturation factor for DCs.
It has been reported that PGE2 works as a maturation factor for DCs and is one of the determinants of Th cell polarization (Kalinski et al., 1998). Although EP2/EP4 receptors were suggested to mediate these effects of PGE2 in mice (Harizi et al., 2003), it is unclear which receptor subtypes are involved in these processes in humans. In the present study, we investigated the involvement of PGE2 receptor subtypes using receptor subtype-selective agonists in the human MoDC maturation and MoDC-induced Th cell polarization.
Materials and Methods
Reagents and Drugs. PGE2 and lipopolysaccharide (LPS) were purchased from Sigma-Aldrich (St. Louis, MO). 17S-2,5-Ethano-6-oxo-17,20-dimethyl prostaglandin E1 (ONO-DI-004), 11,15-O-dimethyl prostaglandin E2 (ONO-AE1-259-01), 16S-9-deoxy-9β-chloro-15-deoxy-16-hyfroxy-17,17-trimethylene-19,20-didehydro prostaglandin F2 (ONO-AE-248), 16-{3-methoxymethyl}phenyl-omega-tetranor-3,7-dithia prostaglandin E1 (ONO-AE1-329), 4-{4-cyano-2-[2-(4-fluoronaphthalen-1-yl) propionylamino] phenyl} butylic acid (ONO-AE3-208), and 11-deoxy-PGE1 were provided by Ono Pharmaceutical Co. (Tokyo, Japan). Dibutyryl cAMP was purchased from Wako Pure Chemicals (Tokyo, Japan). FITC-conjugated mouse IgG1 mAb against CD80 (B7.1) and HLA-DR and PE-conjugated mouse IgG2b mAb against CD83 were purchased from Immunotech (Marseilles, France). FITC-conjugated mouse IgG1 mAb against CD86 (B7.2) was purchased from BD Biosciences PharMingen (San Diego, CA). Recombinant human TNF-α, interferon (IFN)-γ, granulocyte-macrophage colony-stimulating factor (GM-CSF), and IL-4 were purchased from Genzyme-Techne (Minneapolis, MN).
Generation of MoDCs. Peripheral blood mononuclear cells (PBMCs) were isolated by Ficoll-Paque (Pharmacia AB, Uppsala, Sweden) density gradient centrifugation of heparinized blood obtained from buffy coat of healthy volunteers. These PBMCs were plated (1 × 106 cells/ml) in RPMI 1640 medium (Nissui, Tokyo, Japan) supplemented with 10% (v/v) heat-inactivated fetal calf serum, 20 μg/ml kanamycin, and 100 μg/ml streptomycin and penicillin (Sigma-Aldrich). Two hours later, nonadherent cells were removed, and the adherent cells were cultured. In accordance with a previous report (Brossart et al., 1998), the population of the adherent cells was composed of >90% CD14+ cells. The remaining adherent cells were cultured in RPMI 1640 medium, including GM-CSF (50 ng/ml) and IL-4 (50 ng/ml) for 6 days and generated into immature MoDCs as described previously (Lee et al., 2002). In other cases, monocytes were isolated from PBMCs by counterflow centrifugation using the elutriator system (JE-6B-elutriator system; Beckman Coulter Inc., Fullerton, CA). The resultant MoDCs showed CD1a+CD14-HLA-DR+CD83- phenotype, consistent with the previous report (Brossart et al., 1998).
RT-PCR for Human PGE2 Receptors in MoDCs. Immature MoDCs and mature MoDCs induced in the presence of TNF-α (25 ng/ml) were homogenized with RNAzol B (Tel-Test, Inc., Friends-wood, TX), and total RNA was isolated according to manufacturer's instructions. The cDNA synthesized from total RNA using GeneAmp RNA PCR kit (Roche Diagnostics, Indianapolis, IN) was subjected to PCR using 35 cycles at 94°C for 1 min, 55°C for 2 min, and 72°C for 2 min. The oligonucleotide primers for EP1, EP2, and EP4 were used according to previous report (Asano et al., 2002). The primers are as follows: EP1, sense 5′-CGCGC TGCCC ATCTT CTCCAT-3′, anti-sense 5′-CCCAG GCCGA TGAAG CACCAC-3′; EP2, sense 5′-GCTGC TGCTT CTCATT GTCTCG-3′, antisense 5′-TCCGA CAACA GAGGAC TGAACG-3′; EP3, sense 5′-GGACT AGCTC TTCGC ATAACT-3′, antisense 5′-GCAGT GCTCA ACTGA TGTCT-3′; and EP4 sense 5′-ATCTT ACTCA TTGC CACC-3′, antisense 5′-TCTAT TGCTT TACTG AGCAC-3′. The PCR products were identified by electrophoresis on 2% agarose gels with standard size markers and visualized by ethidium bromide staining. We also performed semi-quantitative PCR with GAPDH internal standard.
Flow Cytometric Analysis. On day 6, the immature MoDCs were harvested. After washing, they were cultured (1 × 105 cells/200 μl/well) in RPMI 1640 medium including GM-CSF (50 ng/ml), IL-4 (50 ng/ml), increasing concentrations of PGE2 or EP receptor agonist (10-9-10-5 M), and/or the optimal concentration of TNF-α (25 ng/ml). On day 8, MoDCs were washed once with washing buffer (phosphate-buffered saline supplemented with 2.5% normal horse serum, 0.1% NaN3, and 0.01 M HEPES, pH 7.3). Then, the MoDCs were incubated with 1 μg of FITC-conjugated anti-CD80 Abs, anti-CD86 Abs, anti-HLA-DR Abs, or PE-conjugated anti-CD83 Abs for 20 min at 4°C. After washing, the MoDCs were fixed with 2% paraformaldehyde and analyzed with a FACSCalibur (BD Biosciences, San Jose, CA), and the data were processed using the CellQuest program (BD Biosciences), as described previously (Takahashi et al., 2002). We also evaluated the effects of each EP receptor agonist (ONO-AE1-259-01, ONO-AE1-329 and PGE2;10-7 M, 10-6 M) in the presence of increasing concentrations of EP4 receptor antagonist (ONO-AE3-208) on the expression of CD83 and CD86 on MoDCs. The data are expressed as fold increase in the mean fluorescence intensities against the corresponding values in immature MoDCs. The results are the means ± S.E.M. of five donors.
Measurement of cAMP Production in MoDCs. On day 6, immature MoDCs were harvested. After washing, they were stimulated with PGE2 or EP receptor agonist (10-6 M) in the presence of TNF-α (25 ng/ml) and harvested at 0, 10, and 20 min. MoDCs (2 × 105 cells/200 μl/well) were supplemented with trichloroacetic acid to a final concentration of 5% and 3-isobutyl-1-methylxanthine (an inhibitor of phosphodiesterase) (100 μM) and frozen at -80°C. Frozen samples were sonicated and assayed for cAMP using cyclic AMP enzyme immunoassay kit (Cayman Chemical, Ann Arbor, MI) according to manufacturer's instructions. We performed no acetylation procedures. The results are the means ± S.E.M. of three donors.
Cytokine Production in Mature MoDCs. Immature MoDCs were cultured (1 × 105 cells/200 μl/well) in RPMI 1640 medium including GM-CSF (50 ng/ml), IL-4 (50 ng/ml), and TNF-α (25 ng/ml) in the presence or absence of EP receptor agonists (10-6 M) or dbcAMP (10-5 M) for 2 days. Then, the washed MoDCs were cultured under stimulation of the optimal concentration of LPS (50 ng/ml) for 2 days. When IL-12p70 production was induced, 100 ng/ml IFN-γ was added to LPS. The cell-free supernatant fractions were collected and assayed for IL-12p70, IL-6, and IL-10 protein using ELISA using the multiple Abs sandwich principle (for IL-6 and IL-12p70; Amersham Biosciences UK, Ltd., Buckinghamshire, Little Chalfont, UK), and for IL-10 (BioSource International, Camarillo, CA) as described previously (Kohka et al., 2000).
Purification of Naive T Cells and Induction of Naive CD4+CD45RA+CD45RO- T Cell Differentiation by MoDCs. PBMCs were fractionated by counterflow centrifugation using the elutriator system. Naive CD4+CD45RA+ T cells were isolated form the lymphocyte fraction by negative selection using MACS beads against CD8, CD14, CD19, and CD45RO. The purity of the isolated naive CD4+CD45RA+ T cells was >95% as determined by flow cytometry using FACSCalibur cell sorter. Allogeneic naive CD4+CD45RA+ T cells (2.5 × 105 cells/200 μl of complete culture medium) were cocultured with elutriated monocyte-derived DCs matured with TNF-α (25 ng/ml) and/or EP receptor agonists (10-5 M) at a 1:5 stimulator (DCs)/responder (T cells) ratio in 96-well U-bottomed tissue culture plates (Costar, Cambridge, MA) for 5 days. On day 5, the cells were washed out completely and expanded with fresh medium containing 10 U/ml of human rIL-2 (Shionogi, Osaka, Japan) and cultured for another 9 days. On day 14, the cells were washed, counted, and viable cells were restimulated with anti-CD3 Abs (0.2 μg/ml; BD PharMingen) and anti-CD28 Abs (2.0 μg/ml; BD PharMingen) for 24 h. Cell-free supernatant fractions were collected and frozen at -20°C until cytokine measurement or assayed for IFN-γ, IL-12p70, IL-4, and IL-10 using ELISA as described above.
Statistical Examination. The statistical significance in the dose-response experiments was evaluated using analysis of variance, followed by the Dunnett's test. In other cases, the Student's two-tailed t test was used. A probability value less than 0.05 was considered to be significant.
Results
RT-PCR Analysis for mRNA of EP Receptor Sub-types in MoDC. RT-PCR analysis showed the expression of EP2, EP3, and EP4 mRNA in immature and TNF-α (25 ng/ml)-treated mature MoDCs. Semiquantitative PCR revealed relatively higher amount of EP2 mRNA than EP3 and EP4 mRNA. The expression levels of EP2, EP3, and EP4 mRNA in immature and mature MoDCs were not different substantially (data not shown). Immature and mature MoDCs did not express EP1 mRNA.
Flow Cytometric Analysis of Surface Molecules in MoDCs.Figure 1 shows the dose-response relationships for the effects of EP receptor agonists on the expression of CD80, CD86, CD83, and HLA-DR on MoDCs in the absence of TNF-α. The effects of PGE2 on the expression of CD86 and CD83, a human DC-specific maturation marker, were in a concentration-dependent manner and significant at 10-8 M and above (Fig. 1), and the maximal increases were about 5- and 7-fold, respectively. The effects of PGE2 on the expression of CD80 and HLA-DR did not reach significant levels (Fig. 1). ONO-AE1-259-01 and ONO-AE1-329 also produced marginal but significant increases in CD86 and CD83 expression (Fig. 1); however, ONO-DI-004 (EP1 receptor-selective agonist) (Suzawa et al., 2000) and ONO-AE-248 (EP3 receptor-selective agonist) (Suzawa et al., 2000) had no effect.

Dose-response relationships for the effects of PGE2 and EP receptor agonists on CD80, CD86, CD83, and HLA-DR expression in MoDCs. Immature MoDCs were incubated with increasing concentrations of PGE2 or EP receptor agonists without TNF-α for 2 days and then were stained with FITC or PE-conjugated Abs against CD80 (A), CD86 (B), CD83 (C), and HLA-DR (D) as described under Materials and Methods. The results are the means ± S.E.M. of five donors. *, p < 0.05 compared with control value in the absence of EP receptor agonists.
In contrast to the effects in the absence of TNF-α, the effects of PGE2 on the expression of CD80 and HLA-DR were significant and even more significant were the effects on the expression of CD86 and CD83 in the presence of TNF-α. Figure 2 summarizes the dose-response relationships for the effects of EP receptor agonists and dbcAMP in the presence of TNF-α (25 ng/ml). The effects of PGE2 were almost maximal at the concentration of 10-6 M and above. The effects of ONO-AE1-259-01 and ONO-AE1-329 also mimicked those of PGE2 and the effects were much more significant than those in the absence of TNF-α. The maximal effect of ONO-AE1-259-01 on CD83 were significantly higher than that of ONOAE1-329 (Fig. 2), and the simultaneous stimulation with both agonists produced additive effects equivalent to that with PGE2. Coadministration of PGE2 with the combination of ONO-AE1-259-01 and ONO-AE1-329 had no additive effect on the maximal response of CD83 and CD86 expression in MoDCs induced by PGE2 alone or the combination of ONO-AE1-259-01 and ONO-AE1-329 (data not shown). ONO-DI-004 and ONO-AE-248 also had no effects. The effects of 11-deoxy-PGE1 (EP2/EP4 receptor mixed agonist) mimicked those of PGE2, supporting the requirement of the stimulation of both EP2 and EP4 receptors for MoDC maturation. Dibutyryl cAMP concentration dependently induced the expression of CD80, CD86, CD83, and HLA-DR in the presence of TNF-α. This suggested that PGE2 effects might be through the elevation of cAMP (Fig. 2).

Dose-response relationships for the effects of PGE2, EP receptor agonists, and dbcAMP on CD80, CD86, CD83, and HLA-DR expression in MoDCs in the presence of TNF-α. Immature MoDCs were incubated with increasing concentrations of PGE2, EP receptor agonists, or dbcAMP with TNF-α (25 ng/ml) for 2 days and then were stained with FITC or PE-conjugated Abs against CD80 (A), CD86 (B), CD83 (C), and HLA-DR (D) as described under Materials and Methods. The results are the means ± S.E.M. of five donors. deoxy: 11-deoxy-PGE1. *, p < 0.05, and **, p < 0.01 compared with control value in the absence of both TNF-α and EP receptor agonists. #, p < 0.05 between maximal responses in the presence of EP2 and EP4 receptor agonists.
EP4 receptor antagonist (ONO-AE3-208) concentration dependently inhibited the effects of ONO-AE1-329 and PGE2 (10-7 and 10-6 M) on the expression of CD83 but not the effect of ONO-AE1-259 (Fig. 3).
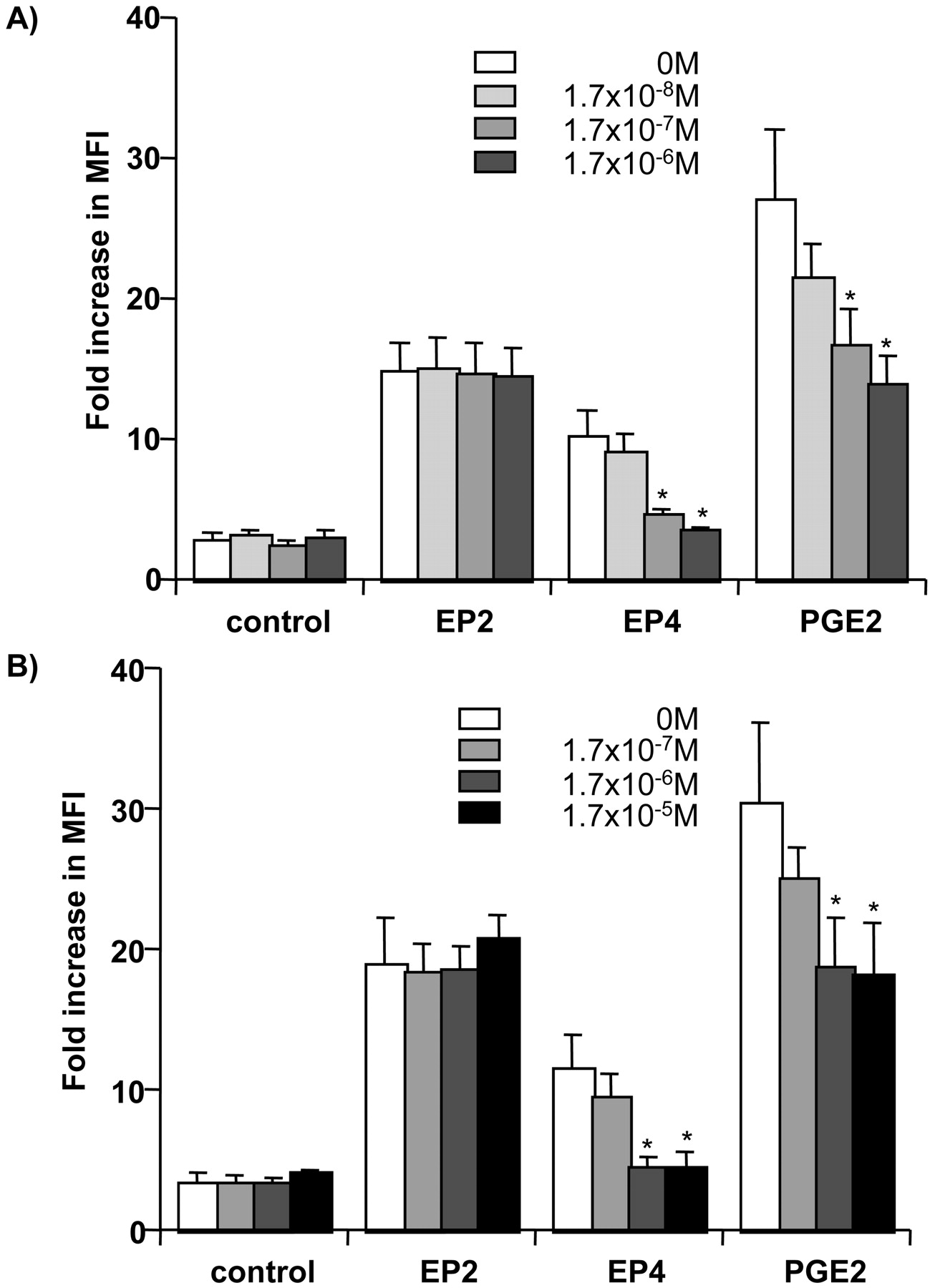
Effects of ONO-AE3-208 (EP4 receptor antagonist) on the enhancement of the expression of CD83 on DC induced by EP receptor agonists. Immature MoDCs were incubated with PGE2 or EP receptor agonists (A, 10-7 M; B, 10-6 M) with TNF-α (25 ng/ml) in the presence of increasing concentration of ONO-AE3-208 (EP4 receptor antagonist) (0-1.7 × 10-6 M). The results are the means ± S.E.M. of three donors. *, p < 0.05 compared with control value in the absence of EP4 receptor antagonist.
cAMP Production. For the evaluation of the signal transduction, we measured cAMP production in MoDCs stimulated with EP receptor agonists. ONO-AE1-259-01, ONO-AE1-329, and PGE2 significantly increased cAMP levels in MoDCs at 10 to 20 min, whereas ONO-DI-004 and ONO-AE-248 had no effect. At 20 min, the production of cAMP in MoDCs stimulated with ONO-AE1-329 was significantly lower than that in ONOAE1-259-01 or PGE2-stimulated MoDCs (Fig. 4).

Measurement of cAMP levels in MoDCs. Immature MoDCs were stimulated with PGE2 or EP receptor agonist (10-6 M) in the presence of TNF-α (25 ng/ml) and harvested at 0, 10, and 20 min. Cyclic AMP was determined using enzyme immunoassay as described under Materials and Methods. The results are the means ± S.E.M. of three donors. **, p < 0.01 compared with the corresponding untreated group (UT). #, p < 0.05 between EP4-treated and PGE2/EP2-treated groups.
Inhibition of IL-12p70, IL-6, and IL-10 Production in MoDCs. After immature MoDCs were cultured with TNF-α for 48 h, LPS stimulation induced a considerable amount of IL-12p70 (Fig. 5A). However, matured MoDCs induced by PGE2 (10-6 M) could not induce the production of IL-12p70 (Fig. 5A). ONO-AE1-259-01 and ONO-AE1-329 (10-6 M) partially inhibited the IL-12p70 production induced by LPS. The combination of these two agonists, 11-deoxy-PGE1 (10-6 M) and dbcAMP (10-5 M), showed a similar extent of inhibition to PGE2. ONODI-004 and ONO-AE-248 had no effect. These results indicated that the stimulation of EP2 and EP4 receptors inhibited the production of IL-12p70 in DCs through the elevation of cAMP. In mature MoDCs induced in the presence of TNF-α (25 ng/ml), LPS (50 ng/ml) stimulation induced IL-6 and IL-10 production (Fig. 5, B and C). However, the LPS-induced production of IL-6 and IL-10 was significantly decreased in matured MoDCs induced by PGE2 (Fig. 5, B and C). Similarly to the case of the inhibition of IL-12 production, ONO-AE1-259-01 and ONOAE1-329 partially inhibited the IL-6 and IL-10 production; however, simultaneous stimulation with both agonists almost completely mimicked the inhibitory effects of PGE2. ONO-DI-004 and ONO-AE-248 had no effect. 11-deoxy-PGE1 and dbcAMP mimicked the inhibitory effects of PGE2 on IL-6 and IL-10 production. Matured MoDCs induced by PGE2 inhibited both Th1 cytokine (IL-12) and Th2 cytokine (IL-6 and IL-10) production, indicating that these MoDCs showed profiles secreting low levels of cytokines through the stimulation of EP2 and EP4 receptors and the elevation of cAMP.

Effects of PGE2, EP receptor agonists, and dbcAMP on the production of IL-12p70, IL-6, and IL-10 in MoDCs. Immature MoDCs were incubated with TNF-α (25 ng/ml), PGE2 (10-6 M), EP receptor agonists (10-6 M), or dbcAMP (10-5 M) for 2 days. On day 8, mature MoDCs were washed and incubated with 50 ng/ml LPS for another 2 days. When the production of IL-12p70 was determined, IFN-γ (100 ng/ml) was added to LPS. On day 10, the concentration of IL-12p70 (A), IL-6 (B), and IL-10 (C) in the supernatant was determined by ELISA as described under Materials and Methods. The results are the means ± S.E.M. of five donors. deoxy: 11-deoxy-PGE1. **, p < 0.01 compared with control value in TNF-α-treated group.
Measurement of IFN-γ, IL-4, and IL-10 by ELISA in Supernatant of Naive CD4+CD45RA+CD45RO- T Cells Cocultured with MoDCs. The alloantigen response in Th cells was induced by coculture of naive Th cells with mature MoDCs induced under different conditions. On the last day of the coculture, we performed flow cytometric analysis and confirmed that there were few MoDCs remaining. When stimulated with TNF-α-treated control MoDCs, the alloantigen-responsive Th cells restimulated with anti-CD3 and anti-CD28 Abs produced considerable amounts of IFN-γ, whereas the production of IL-4 and IL-10 was marginal (Fig. 6). However, when stimulated with matured MoDCs induced by ONO-AE1-259-01 (10-5 M), ONO-AE1-329 (10-5 M), and 11-deoxy-PGE1 (10-5 M), the production of IFN-γ was significantly inhibited, whereas that of IL-4 and IL-10 was vice versa. ONO-DI-004 and ONO-AE-248 had no effect on the cytokine production profile in the coculture of naive Th cells with MoDCs compared with the TNF-α-treated control (Fig. 6).
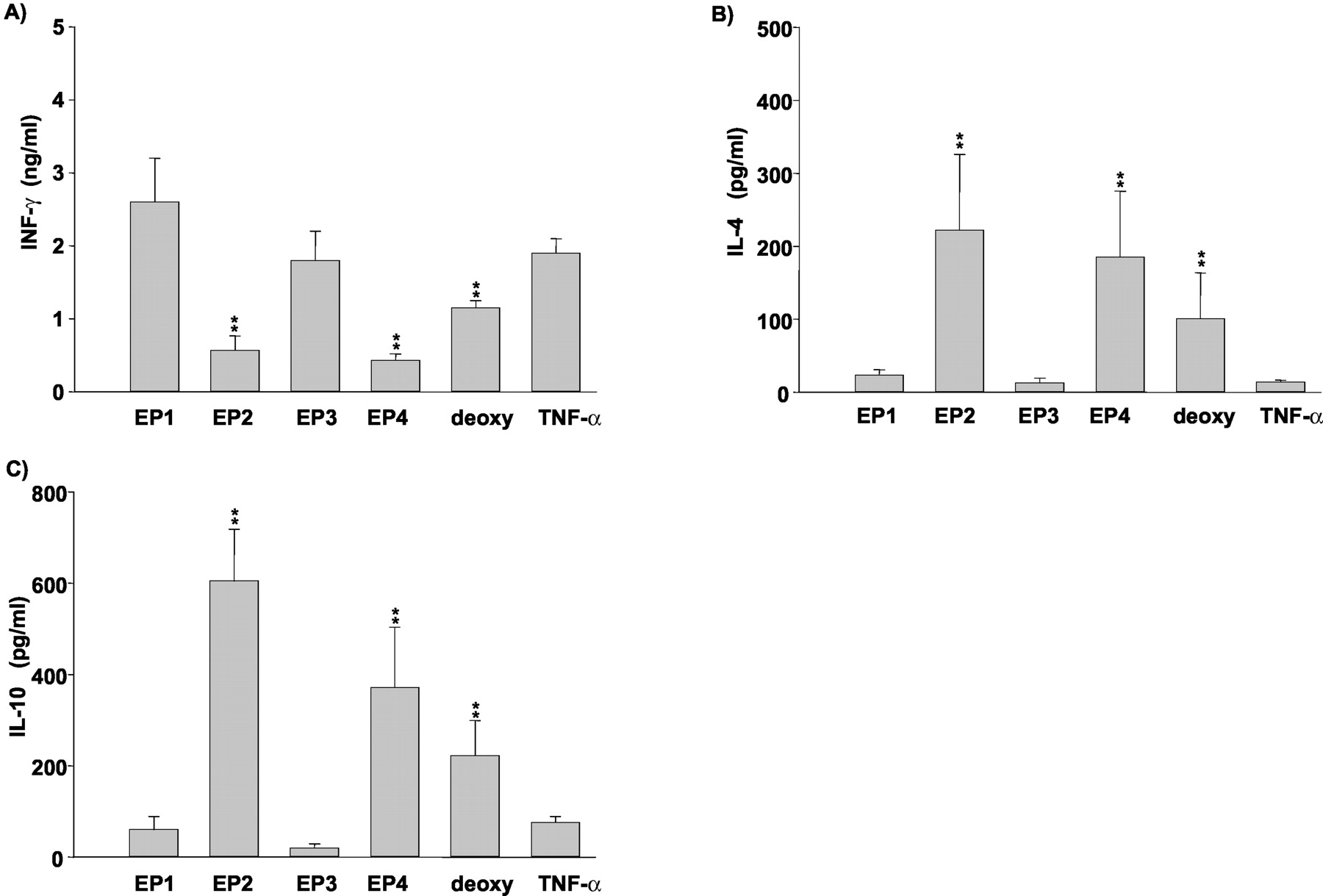
Measurements of IFN-γ, IL-4, and IL-10 by ELISA in supernatant of naive CD4+CD45RA+CD45RO- T cells stimulated by mature MoDCs. Naive T cells were stimulated with MoDCs matured with TNF-α (25 ng/ml) and EP receptor agonists (10-5 M) for 14 days. Five days after the start of coculture, IL-2 (10 U/ml) was added to the culture medium. After washing, the naive T cells were restimulated with anti-CD3 and anti-CD28 antibodies as described under Materials and Methods. The supernatant fractions were harvested and assayed for IFN-γ (A), IL-4 (B), and IL-10 (C) using ELISA. The results are the means ± S.E.M. of five donors. deoxy: 11-deoxy-PGE1. **, p < 0.01 compared with control value in TNF-α-treated group.
Discussion
PGE2 is involved in the inhibition of Th1 cytokine production and Th2 development (Demeure et al., 1997; Minakuchi et al., 1999). It is also reported that PGE2 regulates DC maturation (Rieser et al., 1997; Kalinski et al., 1998; Steinbrink et al., 2000; Luft et al., 2002). However, little was previously known about the distribution and activity of sub-types of EP receptor in human monocyte-derived DCs. Using receptor subtype-selective agonists, we revealed that PGE2 promoted DC maturation, the inhibition of cytokine production, and Th polarization via EP2 and EP4 receptors.
ONO-D1-004 (EP1 receptor-selective agonist), ONO-AE1-259-01 (EP2 receptor-selective agonist), ONO-AE-248 (EP3 receptor-selective agonist), and ONO-AE1-329 (EP4 receptor-selective agonist) were demonstrated to possess high selectivity and were highly specific for each receptor, using a receptor binding assay in Chinese hamster ovary cells transfected with each EP cDNA (Suzawa et al., 2000; Narumiya and FitzGerald, 2001). For example, ONO-AE1-259-01 had at least 2000-fold higher affinity for EP2 receptor compared with that for other EP receptors. In addition, ONO-DI-004 had at least 60-fold, ONO-AE-248 had 400-fold, and ONOAE1-329 had at least 120-fold higher affinity for EP1, EP3, and EP4 receptors, respectively, compared with those for other EP receptors (Narumiya et al., 1999; Suzawa et al., 2000; Narumiya and FitzGerald, 2001). We previously reported that ONO-AE1-259-01 and ONO-AE1-329 regulated the IL-18-induced activation of human monocytes (Takahashi et al., 2002). The potencies of the effects of these agonists on CD83 and CD86 expression on DCs were consistent with those on monocytes (Takahashi et al., 2002; Morichika et al., 2003), indicating that human monocytes and monocyte-derived DCs expressed pharmacologically identical EP2 and EP4 receptors. The EC50 values of ONO-AE1-259-01 and ONO-AE1-329 calculated from our data on the expression of CD86 (8 × 10-8 M and 6 × 10-8 M, respectively) were almost the same order with those reported by Morichika et al. (2003) (3 × 10-8 M and 1 × 10-8 M, respectively), supporting the above-mentioned notion. The selective and complete inhibition by ONO-AE3-208 (EP4 antagonist) of the effect of ONO-AE1-329 (Fig. 3) strongly indicated that the effect of ONO-AE1-329 was solely mediated by EP4. The response induced by PGE2 was partially inhibited by the same antagonist. These results as a whole indicated that the overall effects of PGE2 on the expression of CD83 and CD86 were the additive effects through EP2 and EP4 stimulation.
In our culture condition, MoDCs expressed EP2, EP3, and EP4 receptor mRNA (data not shown). Among the PGE2 receptor subtypes, only EP2 and EP4 receptors were related to MoDC maturation; however, the stimulation of EP3 receptors had no effect. The maximal effect of EP2 receptor agonist was higher than that of EP4 receptor agonist, indicating the functional dominance of EP2 receptor in MoDC maturation. This dominance of EP2 receptor function may be derived from the higher cAMP production (Fig. 4). Semiquantitative PCR showed that EP2 mRNA expression was higher than that of EP4 in both immature and mature MoDCs. The stimulation with EP2 or EP4 receptor agonists alone was less effective than that with PGE2 with respect to the effects on the expression of four molecules, suggesting that the effect of each agonist was partially and not sufficient for the mediation of the effect of PGE2. Simultaneous stimulation with both EP2 and EP4 receptor agonists showed additive effects equivalent to that with PGE2, and 11-deoxy-PGE1 (EP2/EP4 receptor mixed agonist) mimicked the effect of PGE2, indicating that both EP2 and EP4 receptor stimulation by PGE2 is necessary for full activation of the DC maturation processes. Additionally, the mimicry of dbcAMP (a cAMP analog) supported the involvement of Gs protein-coupled EP2 and EP4 receptors in the mediation of PGE2 effects. The fact that the maximal effects induced by PGE2 and by the combination of EP2 and EP4 agonists were almost the same strongly suggested that EP3 receptor stimulation had negligible effect. However, the possibility that EP3 receptor stimulation may has inhibitory modulatory effect (Hatae et al., 2002) could not be excluded.
Similar to the case of the induction of MoDC maturation, PGE2 inhibited the LPS-induced cytokine production in MoDCs by the stimulation with EP2 and EP4 receptor agonists, whereas EP1 and EP3 receptor agonists had no effect. IL-12 produced in antigen-presenting cells, including DCs (DeKruyff et al., 1997; Kalinski et al., 1999), plays important roles in inducing Th1 response and cytolytic T cell activity (Trinchieri and Scott, 1994). In some reports, matured DCs induced by PGE2 were demonstrated to induce Th2 responses (Kalinski et al., 1998, 2001; Luft et al., 2002). Added to the mixed culture with naive T cells and DCs matured with PGE2, IL-12 restored Th1 polarization and is thought to be a crucial factor in Th bias (Kalinski et al., 2001). In the present study, PGE2 inhibited LPS-induced IL-12 production in MoDCs through the stimulation of EP2 and EP4 receptors. Together with Th1 cytokine (IL-12) production, Th2 cytokine (IL-6 and IL-10) production was inhibited by PGE2 through the stimulation of EP2 and EP4 receptors. In this study, both Th1 and Th2 cytokine production were inhibited by PGE2, indicating that DCs with a low ability for cytokine production were generated by PGE2, through the stimulation of EP2 and EP4 receptor. Previously, matured DCs induced by IL-1β and TNF-α were reported to produce a reduced amount of IL-12 in response to IFN-γ in the presence of CD40L compared with immature DCs (Kalinski et al., 1999). Matured DCs induced by PGE2 were at the final stage of differentiation (Kalinski et al., 1998) and were reported to exhibit profiles secreting low levels of cytokines, despite the addition of stimulation such as CD40L and LPS (Luft et al., 2002; Scandella et al., 2002). Therefore, the reduced cytokine production in MoDCs may be due to the final maturation of MoDCs by PGE2 through the stimulation of EP2 and EP4 receptors. Because IL-12 is indicated to be a crucial factor for T cell polarization, inhibition of the production of IL-12 by MoDCs via EP2 and EP4 receptors is speculated to induce Th2 bias. However, IL-10 is reported to inhibit DC maturation, when added to the culture medium during DC maturation (Kalinski et al., 1998). Therefore, it is possible that the inhibition of IL-10 production in MoDCs via EP2 and EP4 receptors may indirectly contribute to the facilitation of MoDC maturation.
In DC-T cell cultures, there were few MoDCs remaining on the last day of the culture. Therefore, the cytokines produced during 24-h stimulation seemed to be of T cell origin. T cells showed decreased Th1 cytokine (IFN-γ) production and increased Th2 cytokine (IL-4 and IL-10) production, when stimulated with matured MoDCs induced by EP2 and EP4 receptor agonists, although EP1 and EP3 receptor agonists did not show such modulation, supporting the results that the modulation of DC function was related solely to EP2 and EP4 receptors. Although LPS-induced cytokine production in matured MoDCs stimulated via EP2 and EP4 receptors was generally inhibited regardless of Th1 or Th2 cytokine, these matured MoDCs inhibited only Th1 cytokine production and induced Th2 cytokine production in naive T cells, indicating that matured MoDCs induced by the stimulation of EP2 and EP4 receptors preferentially induced Th2 polarization of naive T cells.
EP2 and EP4 receptors have been suggested to be involved in the regulation of monocytes/macrophages (Hubbard et al., 2001; Ikegami et al., 2001; Takahashi et al., 2002), T cells (Nataraj et al., 2001), and B cells (Fedyk and Phipps, 1996). On the other hand, EP1 and EP3 receptors are involved in IgE/antigen-mediated degranulation and release of IL-6 and GM-CSF in mast cells (Gomi et al., 2000). Thus, PGE2 exerts complex immunological modulation via its four EP receptors. Of course, it is important to note that EP receptor expression and function may be different among species and cell types. Scandella et al. (2002) reported facilitatory effects of PGE2 via EP2 and EP4 receptors on the induction of migration in human monocyte-derived DCs; however, Harizi et al. (2003) reported that EP2 and EP4 receptor stimulation inhibited the maturation of mouse bone marrow-derived DCs. The former also reported the down-regulation of EP2 and EP4 receptor expression by exogenous PGE2 (Scandella et al., 2002); however, the latter reported the up-regulation of EP2 and EP4 receptor expression via COX-2-induced endogenous PGE2 by LPS stimulation (Harizi et al., 2003). Thus, conflicting results about EP receptor expression and function are reported, indicating the difficulty in direct comparison of different studies without considering species and cell types.
Our results clearly demonstrated the cooperative stimulation of EP2 and EP4 receptor produced full maturation of human MoDCs, which in turn preferentially induced Th2 development of naive T cells. To apply PGE2-related agents to the DC-mediated immune response, the differences in EP receptor distribution and activity should be evaluated. These phenotypic features in PGE2-induced MoDCs should be taken into consideration when clinical therapy using in vitro-generated DCs is being planned.
Acknowledgments
We thank Ono Pharmaceutical Co. (Tokyo, Japan) for generously donating ONO-DI-004, ONO-AE1-259-01, ONO-AE-248, ONO-AE1-329, and 11-deoxy-PGE1. We also thank Yumiko Shiotani and Yuki Onoda for excellent technical assistance.
Footnotes
-
This study was in part supported by a Grant-in-Aid C15590467 from the Ministry of Education, Culture, Science and Sports, Japan (to H.K.T.) and a grant (No. 21) from Okayama University (to M.N.).
-
DOI: 10.1124/jpet.103.062646.
-
ABBREVIATIONS: LPS, lipopolysaccharide; TNF, tumor necrosis factor; DC, dendritic cell; COX, cyclooxygenase; PG, prostaglandin; Th, helper T; IL, interleukin; EP, E-prostanoid; MoDC, monocyte-derived dendritic cell; dbcAMP, dibutyryl cAMP; FITC, fluorescein isothiocyanate; mAb, monoclonal antibody; IFN, interferon; GM-CSF, granulocyte-macrophage colony-stimulating factor; PBMC, peripheral blood mononuclear cell; RT-PCR, reverse-transcription-polymerase chain reaction; PCR, polymerase chain reaction; Ab, antibody; PE, phycoerythrin; ELISA, enzyme-linked immunosorbent assay.
- Received November 11, 2003.
- Accepted February 9, 2004.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}