

Abstract
This study examined the role of metabotropic glutamate receptors (mGluRs) in hippocampal long-term depression (LTD) in vivo. The group 1 mGluR antagonist (S)4-carboxyphenylglycine (4CPG), group 1/2 antagonist (RS)-α-methyl-4-carboxyphenylglycine (MCPG), and group 2 antagonists (RS)-α-methylserine-O-phos-phate monophenyl ester (MSOPPE) and (2S)-α-ethylglutamic acid (EGLU) were used. The NMDA receptor antagonistd(−)-2-amino-5-phosphonopentanoic acid (AP5) was used to examine the NMDA receptor contribution to the observed LTD. Adult male Wistar rats underwent implantation of stimulating and recording electrodes into the Schaffer collaterals and CA1 stratum radiatum, respectively. After recovery of 5–7 d, the field EPSP was measured from freely moving animals. Drugs were applied either before or after 1 Hz low-frequency train (LFT) or 100 Hz stimulation via a cannula implanted in the lateral cerebral ventricle. Nine hundred pulses at 1 Hz produced an LTD that was marked and long-lasting. This LTD was completely inhibited by pre-LFT application of AP5. MCPG inhibited LTD from 2 hr post-LFT. 4CPG partially impaired LTD. MSOPPE and EGLU completely blocked induction of LTD, although short-term depression remained intact. MSOPPE did not block long-term potentiation (LTP) induced by 100 Hz stimulation, whereas 4CPG produced a significant inhibition. When MSOPPE was present, LTD could not be induced either before or after LTP induction, whereas LTD could be induced in an identical protocol in vehicle-injected animals.
These results suggest a differential role for mGluRs in NMDA receptor-dependent hippocampal LTD in vivo. Group 1 mGluRs may play a role in both LTD and LTP, whereas group 2 mGluRs may be critically involved only in LTD induction.
- metabotropic glutamate receptors
- long-term depression
- long-term potentiation
- in vivo
- hippocampus
- NMDA receptors
Long-term depression (LTD) is a long-lasting nonpathological decrease in synaptic transmission that has been studied mostly in cerebellum (Ito, 1989; Linden, 1994) but has also been reported in the hippocampus (Bear and Malenka, 1994). Hippocampal LTD shares characteristics with another form of synaptic plasticity, namely long-term potentiation (LTP) (Bliss and Lomo, 1973), in that it is dependent on calcium entry through NMDA receptor channels (Dudek and Bear, 1992; Mulkey and Malenka, 1992). It has also been postulated that LTD represents an inverse form of LTP (Bear and Malenka, 1994; Stevens and Wang, 1994).
It has been reported that in the hippocampus, LTD requires activation of metabotropic glutamate receptors (mGluRs) for robust induction to take place (Bashir et al., 1993; O’Mara et al., 1995), although others have refuted this claim (Selig et al., 1995). As yet, little information exists as to which mGluR subtypes, if any, may participate in this phenomenon. To date, eight mGluR subtypes (mGluR 1–8) have been identified (Masu et al., 1991; Abe et al., 1992; Tanabe et al., 1992, 1993; Nakajima et al., 1993; Okamoto et al., 1994; Duvoisin et al., 1995). Group 1 mGluRs comprise mGluRs 1 and 5, which are mainly postsynaptically localized in hippocampus, are coupled to phospholipase C, and are activated by 1S3R-1-aminocyclopentane-1,3- dicarboxylic acid (ACPD). Group 2 mGluRs include mGluRs 2 and 3, which are also activated by ACPD but are coupled to adenylate cyclase and localized mainly presynaptically. mGluRs 4, 6, 7, and 8 also couple to adenylate cyclase but share an agonist preference for l-2-aminophosphono butyrate (l-AP4) and have therefore been classified as group 3 mGluRs.
Group 1 mGluRs seem to play a critical role in LTP inductionin vivo (Manahan-Vaughan and Reymann, 1996; Manahan-Vaughan et al., 1996). Furthermore, activation of group 3 mGluRs can both depress basal synaptic transmission and inhibit LTP in the CA1 regionin vivo (Manahan-Vaughan and Reymann, 1995a). To further characterize the involvement of mGluR subtypes in hippocampal synaptic plasticity, the present study investigated the contribution of group 1 and 2 mGluRs to induction of LTD and LTP in the hippocampal CA1 region of freely moving rats.
MATERIALS AND METHODS
Surgical preparation. Seven-week-old male Wistar rats were prepared as described previously (Manahan-Vaughan and Reymann, 1995a). Briefly, under sodium pentobarbitone anesthesia (Nembutal, 40 mg/kg, i.p), animals underwent implantation of a monopolar recording and a bipolar stimulating electrode (made from 0.1 mm diameter Teflon-coated stainless steel wire). A hole was drilled for the recording electrode (1 mm diameter, 2.8 mm posterior to bregma, 1.8 mm lateral to the midline), and a second hole (1 mm diameter, 3.1 mm posterior to bregma, 3.1 mm lateral to the midline) was drilled for the stimulating electrodes [coordinates based on Paxinos and Watson (1986)]. The dura was pierced through both holes, and the recording and stimulating electrodes were lowered into the CA1 stratum radiatum and the Schaffer collaterals, respectively. Recordings of evoked field potentials via the implanted electrodes were taken throughout surgery. A cannula was implanted in the lateral cerebral ventricle (0.08 mm posterior to bregma, 1.6 mm lateral to the midline). Once verification of the location of the electrodes was complete, the entire assembly was sealed and fixed to the skull with dental acrylic (Paladur, Heraeus Kulzer GmbH). The animals were allowed between 5 and 7 d to recover from surgery before experiments were conducted. During this period they were monitored closely for infection or distress and handled regularly. Twenty-four hours before the commencement of the experiments, animals were placed in the recording chamber with full access to food and water, to allow familiarization to occur. Throughout the experiments, the animals could move freely within the recording chamber (40 cm × 40 cm × 40 cm), because the implanted electrodes were connected by a flexible cable and swivel connector to the stimulation unit and amplifier. Aside from the insertion of the connector cable and injection cannula at the start of the experiment, disturbance of the animals was kept to an absolute minimum. Throughout the experiments the electroencephalogram of each animal was monitored continuously.
At the end of the study, brains were removed and histological verification of electrode and cannula localization was carried out. Brain sections (16 μm) were embedded in paraffin, stained according to the Nissl method using 1% toluidine blue (Bock, 1989), and then examined using a light microscope. Brains in which an incorrect electrode or cannula localization was found were discarded from the study.
Measurement of evoked potentials. The field EPSP (fEPSP) slope function was used as a measure of excitatory synaptic transmission in the CA1 region. To obtain these measurements, an evoked response was generated in the stratum radiatum by low-frequency (0.025 Hz) stimulation with single biphasic square wave pulses of 0.1 msec duration per half wave, generated by a constant current isolation unit. For each time point measured during the experiments, five records of evoked responses were averaged. fEPSP was measured as the maximal slope through the five steepest points obtained on the first positive deflection of the potential. By means of input/output curve determination the maximum fEPSP was found, and during experiments all potentials used as baseline criteria were evoked at a stimulus intensity that produced 40% of this maximum.
LTD was induced by a low-frequency train (LFT) of 1 Hz (900 pulses) and a stimulus intensity that was 60–70% of the maximum (determined by means of the input/output curve). The input specificity of the LTD obtained was verified via implantation of a control stimulating electrode in the contralateral commissural path. Alternate stimulation via the contralateral and ipsilateral stimulating electrodes demonstrated that basal synaptic transmission in the CA1 was evoked by low-frequency stimulation (0.025 Hz) through the contralateral electrode, whereas LTD was evoked via the ipsilateral electrode through which LFT had been given (data not shown). LTP was induced by a high-frequency tetanus (HFT) of 100 Hz (10 bursts of 10 stimuli, 0.1 msec stimulus duration, 10 sec interburst interval) and a stimulus amplitude that was 20% of the maximum (determined by means of the input/output curve).
Compounds and drug treatment.(RS)-α-methyl-4-carboxyphenylglycine (MCPG), (S)-4-carboxyphenylglycine (4-CPG), (2S)-α-ethylglutamic acid (EGLU),d(−)-2-amino-5-phosphonopentanoic acid (AP5), and (RS)-α-methylserine-O-phosphate monophenyl ester (MSOPPE) were obtained from Tocris Cookson (Bristol, UK). For injection, all drugs were first dissolved in 5 μl sodium hydroxide solution (1 mm) and then made up to a 100 μl volume with 0.9% sodium chloride. Throughout the experiments, drugs or vehicle were applied into the lateral cerebral ventricle via an injection cannula that was inserted through the surgically implanted outer cannula. Drug or vehicle injection occurred either after measurement of the baseline for 30 min or 5 min after HFT or LFT. The injection cannula was inserted before the baseline measurements were taken and left in place for the duration of the experiment so that an artifact in recordings attributable to its insertion or removal would not be created.
The drugs were injected in a 5 μl volume over a 6 min period via a Hamilton syringe. In LTD and LTP experiments, an HFT or LFT was applied 30 min after drug injection (or 60–90 min after the start of the experiment when injections occurred post-HFT/LFT), with measurements then taken at t = 5, 10, and 15 min and then at subsequent 15 min intervals up to 4 hr. In some cases, an additional measurement was taken 24 hr post-HFT/LFT. In baseline experiments, the same protocol was followed, except that no HFT or LFT was given. To approximate drug concentrations in vivo and to match the reported effective concentrations of the drugs in vitro, drug dose calculations were initially based on the assumption that the rat brain volume is ∼2 ml. Thus for an estimated brain concentration of 50 μm AP5, 20 mm (100 nmol) in a 5 μl injection volume was used. Dose–response relationships were subsequently carried out to determine the effective concentration ranges of the drugs used.
Data analysis. The baseline fEPSP data were obtained by averaging the response to stimulation of the Schaffer collaterals, to obtain five sweeps at 40 sec intervals, every 5 min over a period of 30 min. Drug or vehicle injections were then applied and followed by an additional six recordings in a 30 min period. At this point LFT or HFT was given, and three additional measurements at 5 min intervals were taken, followed by recordings at 15 min intervals for 4 or 24 hr. The data were then expressed as mean% preinjection baseline fEPSP reading ± SEM. Statistical significance was estimated using ANOVA with repeated measures, followed by post hoc Student’st tests. The probability levels interpreted as statistically significant were p < 0.001 (***), p < 0.01 (**), p < 0.05 (*).
RESULTS
Hippocampal LTD in freely moving rats is NMDA receptor dependent
The involvement of NMDA receptors in the induction of LTD was examined using the NMDA receptor antagonist AP5. When an LFT (900 pulses at 1 Hz) was applied via the Schaffer collateral-commissural pathway to the stratum radiatum of the CA1 region, an LTD of the fEPSP (n = 6) was generated that was still present 24 hr after LFT. No change in the profile of LTD was seen when a vehicle injection was applied 30 min before LFT via a cannula implanted in the lateral cerebral ventricle (n = 19) (Fig. 1). The fEPSP 5 min after LFT was 64 ± 3% of the preinjection fEPSP baseline values. In six of the experiments, the profile of the depression was followed for 24 hr; it was found that the fEPSP was 73 ± 5% of preinjection values. This was not different from the degree of depression seen in noninjected LFT animals but was significantly different from non-LFT animals (n = 12; ANOVA: F(1,25)= 6.7; p < 0.001).

LTD in the CA1 region in vivo is dependent on activation of both NMDA receptors and mGluRs.A, A low-frequency train (LFT) in the presence of a vehicle injection (n = 19) results in a robust LTD that persists for 24 hr. Previous injection of the NMDA receptor antagonist AP5 (20 mm/5 μl;n = 9) completely inhibits the induction of LTD, from t = 5 min post-LFT. B, Original analog traces showing evoked responses in the CA1 region at three time points: preinjection, t = 5 min, andt = 4 hr post-LFT, in (1) a vehicle-injected animal and (2) an animal injected with MCPG (200 mm/5 μl).C, MCPG (200 mm/5 μl;n = 8) significantly inhibits LTD, fromt = 120 min post-LFT, compared with vehicle-injected controls (n = 10). *p < 0.05, **p < 0.01, ***p < 0.001. Line breaks(//) indicate change in time scale.
When AP5 was applied before LFT, in a concentration that had previously been found to block LTP in vivo [20 mm (100 nmol)/5 μl] (Manahan-Vaughan et al., 1996), a complete inhibition of LTD occurred (Fig. 1A) (ANOVA:F(1,25) = 13.3; p < 0.001). Thus, 5 min after LFT, the fEPSP in the AP5 group was 94 ± 4% of preinjection fEPSP baseline values (n = 9; p < 0.01 compared with controls). This significant inhibition of LTD was maintained throughout the duration of the experiment. Therefore, activation of NMDA receptors during LFT is essential for initiation of LTD to occur in vivo.
It was shown previously that the same concentration of AP5 (20 mm/5 μl) has no effect on hippocampal basal synaptic transmission when injected into the lateral cerebral ventricle (Manahan-Vaughan et al., 1996). This finding rules out the possibility that a direct effect of AP5 on basal synaptic transmission explains the inhibition of LTD obtained by AP5.
LTD in vivo requires activation of mGluRs
To investigate the involvement of mGluRs in LTD, the general mGluR antagonist MCPG was used. This compound, in a concentration of 200 mm (1 μmol)/5 μl, significantly inhibits LTP in vivo, allowing only short-term potentiation (STP) to occur (Manahan-Vaughan and Reymann, 1995a; Manahan-Vaughan et al., 1996). When the same concentration of MCPG (200 mm/5 μl;n = 8) was administered 30 min before LFT, a significant inhibition of LTD occurred (ANOVA:F(1,24) = 20.6; p < 0.001). At first, however, a depression appeared that was not statistically different from controls (58 ± 7% of preinjection fEPSP baseline values at 5 min post-LFT in the MCPG-injected animals; 61 ± 6% in controls; n = 12) (Fig.1B,C), but by 120 min post-LFT the fEPSP values of the MCPG-injected group had recovered significantly, being 92 ± 8% of preinjection values compared with 76 ± 3% in controls (p < 0.05). By 135 min post-LFT, the fEPSP values in the MCPG-injected animals had returned to pre-LFT levels (Fig. 1C). It was determined previously that this concentration of MCPG (200 mm/5 μl) has no effect on basal synaptic transmission in the CA1 region when injected into the lateral cerebral ventricle (Manahan-Vaughan and Reymann, 1995b).
Group 1 mGluRs play a role in LTD in vivo
To attempt to dissect the mGluR-mediated component of LTD into the receptor subtypes involved, the group 1 mGluR antagonist 4CPG was used. 4CPG has an IC50 value of 4 × 10−5m at mGluR 1 and an EC50 value of 5 × 10−4m at mGluR 2 and is inactive at mGluR 4 (Hayashi et al., 1994; tested in chinese hamster ovary cells). In addition, it has been reported that 4CPG inhibits ACPD-stimulated phosphoinositide hydrolysis in cerebrocortical slices (Eaton et al., 1993).
When 4CPG in a concentration of 2 mm (n = 4), 4 mm (n = 4), or 10 mm/5 μl (n = 4) was applied 30 min before LFT, no significant effect on LTD was seen (Fig. 2C). When 4CPG (n = 7) was administered 30 min before LFT, however, in a concentration that was found previously to be effective in blocking LTP in vivo (20 mm (100 nmol)/5 μl), a significant impairment of LTD expression was seen compared with controls (n = 12; ANOVA:F(1,24) = 21.9; p < 0.001). As was the case with MCPG, application of 4CPG did not affect short-term depression (Fig. 2A,B). By 120 min post-LFT, however, a recovery of fEPSP values toward preinjection levels had occurred, and a significant difference was evident from this time point onward compared with LFT controls (88 ± 6% of preinjection fEPSP baseline values compared with 74 ± 3% in controls;p < 0.05) (Fig. 2A,B). A complete recovery back to preinjection fEPSP baseline values did not take place in the 4CPG group; rather, the fEPSP values showed a significant difference when compared with control non-LFT animals (n = 8; ANOVA: F(1,24) = 3.2;p < 0.001) (Fig. 2A,B). Thus, 4CPG seemed to produce a partial but significant block of LTD.

LTD in the CA1 region in vivo is modulated by group 1 mGluRs. A, The group 1 mGluR antagonist 4CPG (20 mm/5 μl; n = 7) partially impairs LTD when compared with vehicle-injected controls (n = 12), from t = 120 min post-LFT. This effect was significantly different from baseline values.B, Original analog traces showing evoked responses in the CA1 region at three time points: preinjection,t = 5 min, and t = 4 hr post-LFT, in an animal injected with 4CPG (20 mm/5 μl).C, Dose–response curve for the antagonist effect of 4CPG (2–40 mm/5 μl) on LTD in the CA1 region. The values represent the magnitude of LTD observed at 4 hr post-LFT.D, Application of 4CPG (20 mm/5 μl;n = 5) 5 min after LFT results in a significant inhibition of LTD, from t = 135 min post-HFT compared with vehicle-injected controls (n = 5). *p < 0.05. Line breaks(//) indicate change in time scale.
When basal synaptic transmission was monitored for 4 hr in the presence of 4CPG (20 mm/5 μl), no effect on synaptic transmission was observed (n = 4). In addition, when the higher concentration of 40 mm 4CPG/5 μl was applied 30 min before LFT, no further inhibition of LTD was seen compared with the 20 mm/5 μl concentration (Fig. 2C).
To examine whether 4CPG modulates LTD by influencing its initiation, experiments were carried out in which the compound was applied 5 min after the termination of LFT (n = 5). In this case an impairment of LTD was obtained similar to that seen when 4CPG was applied before LFT (ANOVA: F(1,25) = 15.2;p < 0.001). Thus, no difference in the degree of depression was seen initially when compared with controls (n = 6) (Fig. 2D). From 135 min post-LFT, however, the magnitude of depression of fEPSP in the 4CPG group had reduced (85 ± 2% of preinjection values compared with 67 ± 6% in controls; p < 0.05) (Fig.2D), and a significant difference occurred from this time point onward until the end of the experiment. A complete recovery back to preinjection fEPSP baseline values did not occur in the 4CPG group; rather, the values showed a significant difference when compared with control non-LFT animals (n = 8; ANOVA: p < 0.001).
These data indicate that group 1 mGluRs do not modulate LTD through an interaction with NMDA receptors during LTD initiation, for example, but rather seem to modulate the maintenance phase of LTD.
Group 1 mGluR activation is necessary for persistent LTPin vivo
When a vehicle injection (n = 6) was applied 30 min before an HFT, LTP was induced that persisted for >24 hr (Fig.3A). This potentiation was statistically significant from control (nontetanized), from t = 5 min post-HFT onward. When 4CPG (n = 5) in a concentration of 20 mm (100 nmol)/5 μl was administered 30 min before HFT, a significant difference between 4CPG- and vehicle-treated animals was seen (ANOVA: F(1,25) = 24.2;p < 0.001). Thus, although STP remained intact, by 60 min post-HFT, fEPSP values had declined to 126 ± 10% in the 4CPG group compared with 155 ± 6% in controls. At 24 hr post-LFT, the control group was still potentiated, whereas in the 4CPG group fEPSP values returned to pre-HFT baseline levels by 90 min. Therefore, a concentration of 4CPG that significantly inhibits LTD also successfully inhibits LTP. This finding supports a role for group 1 mGluRs in bidirectional modulation of synaptic plasticity.
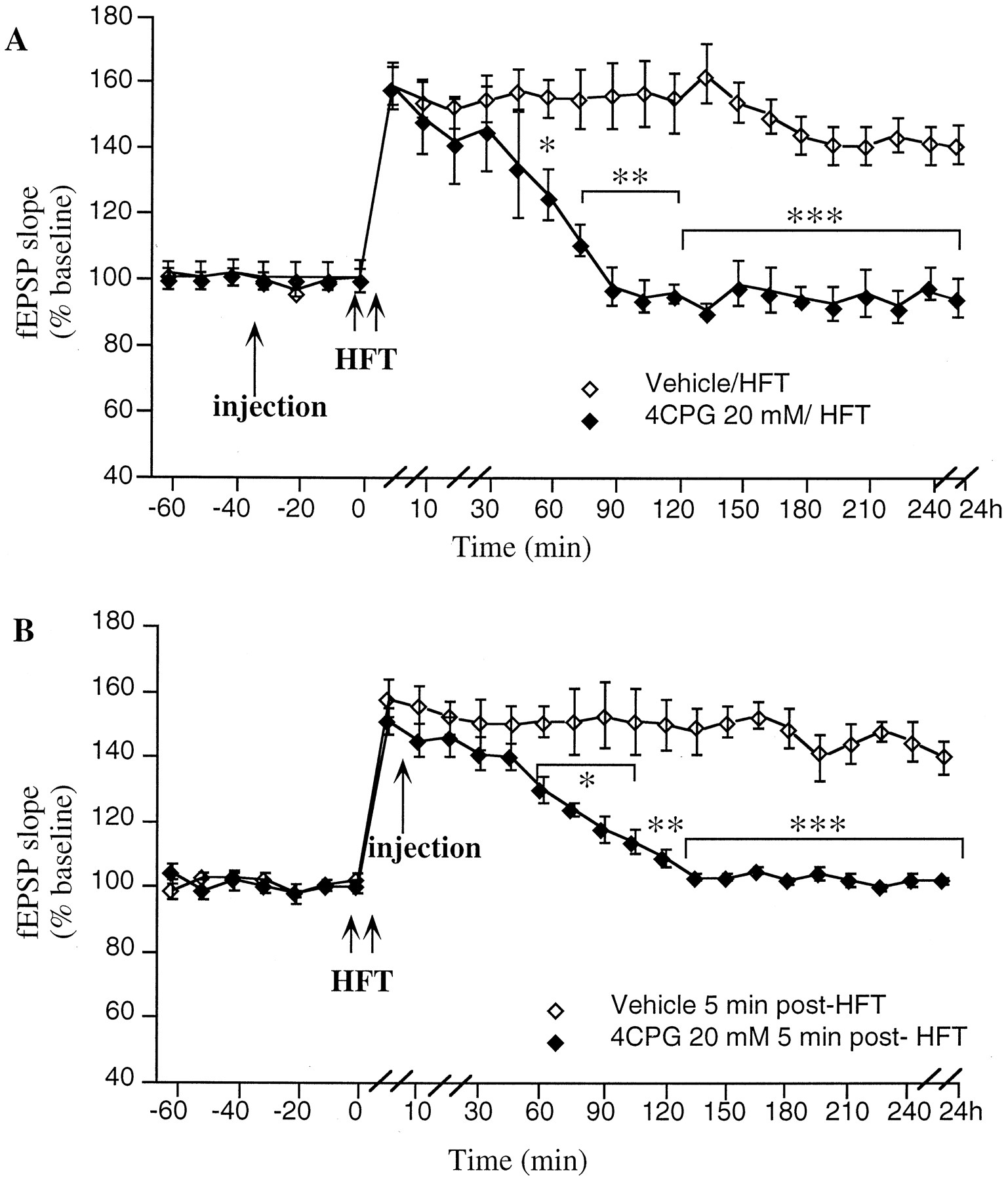
LTP in the CA1 region in vivo is modulated by group 1 mGluRs. A, 4CPG (20 mm/5 μl; n = 5) significantly inhibits LTP, from t = 60 min after a high-frequency tetanus (HFT) compared with vehicle-injected controls (n = 6).B, Application of 4CPG (20 mm/5 μl;n = 5) 5 min after HFT results in a significant inhibition of LTP, from t = 45 min post-HFT compared with vehicle-injected controls (n = 5). *p < 0.05, **p < 0.01, ***p < 0.001. Line breaks(//) indicate change in time scale.
To investigate whether 4CPG inhibits the initiation or maintenance phases of LTP, the compound was applied 5 min after LTP. It was found that an inhibition of LTP occurred (ANOVA:F(1,25) = 34.3; p < 0.001) similar to that seen when the drug was applied before HFT. At 5 min post-HFT, the amplitude of LTP was 151 ± 4% of preinjection fEPSP baseline levels (n = 5) compared with 161 ± 5% in controls (n = 5). By 45 min post-HFT, a significant reduction in the degree of potentiation was seen in the 4CPG group when compared with controls (p < 0.05), and by 120 min post-HFT, fEPSP values had returned to preinjection levels (Fig.3B). Thus, group 1 mGluRs seem to contribute to the maintenance rather than the initiation of LTP.
Group 2 mGluR activation is required for persistent LTDin vivo
MSOPPE is a selective group 2 mGluR antagonist (Jane et al., 1996a; Thomas et al.,1996a) that shows a preference for group 2 mGluRs over group 3 mGluRs. When MSOPPE in a concentration of 4 mm(n = 4), 10 mm (n = 4), or 20 mm/5 μl (n = 4) was applied 30 min before LFT, no effect on LTD was seen (Fig.4C). When the concentration was raised to 40 mm (200 nmol)/5 μl, a significant inhibition of LTD was observed (ANOVA: F(1,24) = 43.8;p < 0.001) (Fig. 4A,B). Although the degree of LTD immediately after LFT was not significantly different in drug (n = 12) and control (n = 12) groups, by 90 min post-LFT a significant inhibition of LTD had occurred in the MSOPPE group compared with controls (94 ± 6% compared with 75 ± 4% in controls; p < 0.05) (Fig.4A). In the MSOPPE group the evoked potentials returned to pre-LFT values by 165 min post-LFT, whereas in the control group values were still depressed at 4 hr post-LFT.
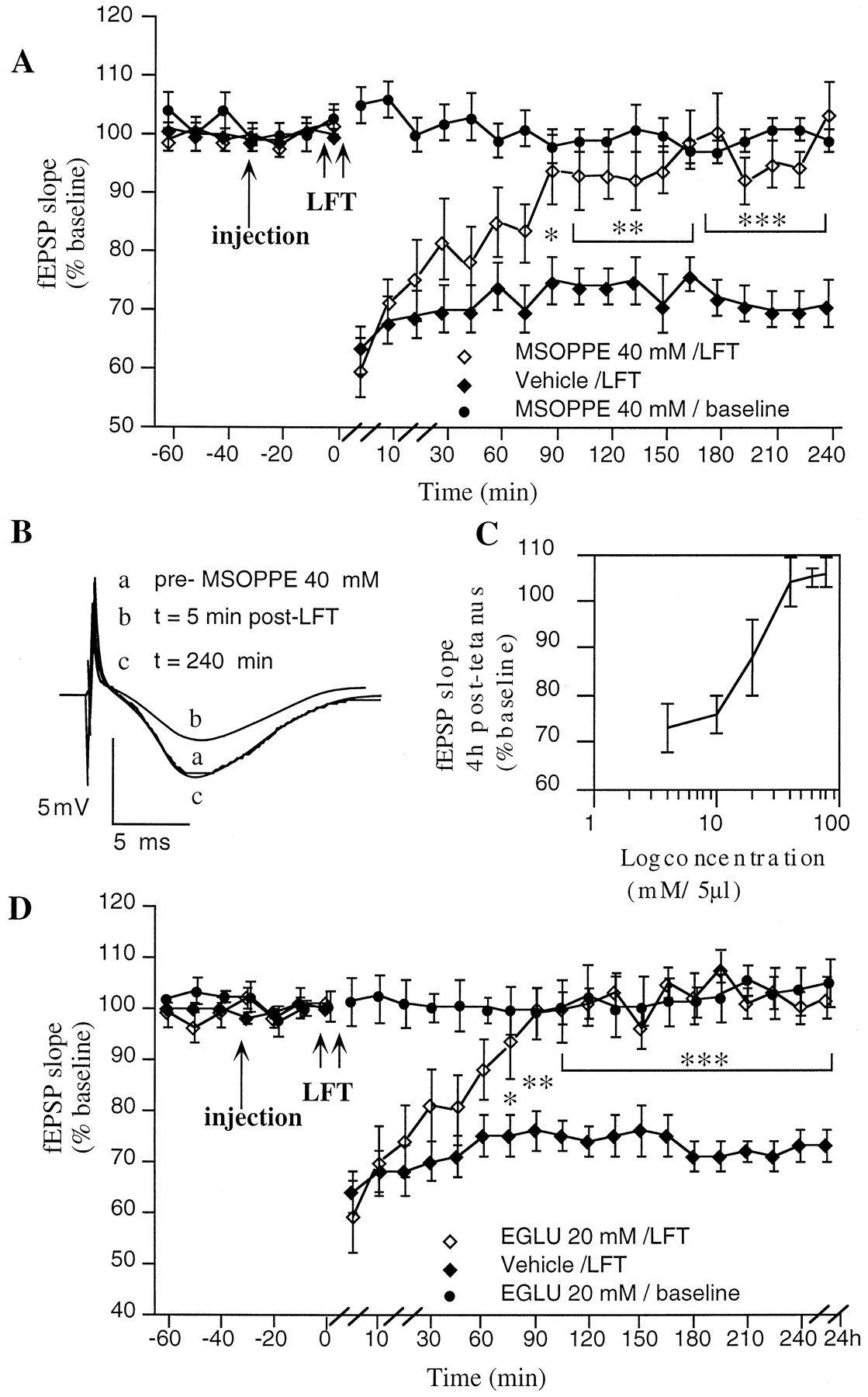
LTD in the CA1 region in vivocritically involves group 2 mGluRs. A, The group 2 mGluR antagonist MSOPPE (40 mm/5 μl;n = 12) completely blocks LTD, fromt = 90 min post-LFT when compared with vehicle-injected controls (n = 12).B, Original analog traces showing evoked responses in the CA1 region at three time points: preinjection,t = 5 min, and t = 4 hr post-LFT, in an animal injected with MSOPPE (40 mm/5 μl).C, Dose–response curve for the antagonist effect of MSOPPE (4–80 mm/5 μl) on LTD in the CA1 region. The values represent the magnitude of LTD observed at 4 hr post-LFT.D, The group 2 mGluR antagonist EGLU (n = 6) completely blocks LTD, fromt = 75 min post-LFT when compared with vehicle-injected controls (n = 11). *p < 0.05, **p < 0.01, ***p < 0.001. Line breaks(//) indicate change in time scale.
When basal synaptic transmission was monitored for 4 hr in the presence of MSOPPE (40 mm/5 μl; n = 8), no effect on synaptic transmission was seen compared with controls (n = 8). When the higher concentrations of 60 mm (n = 4) and 80 mm MSOPPE/5 μl (n = 4) were applied 30 min before LFT, no improvement in the degree of inhibition of LTD obtained was seen compared with the 40 mm/5 μl dose (Fig.4C).
EGLU is a highly selective antagonist for group 2 mGluRs that exhibits a potency at group 2 mGluRs similar to that of MSOPPE but has no antagonist action at group 3 mGluRs (Jane et al., 1996b). Thus, EGLU was used to examine whether the blockade of LTD seen with MSOPPE can be explained via antagonism of group 2 mGluRs. When EGLU was applied in concentrations of either 5 or 10 mm/5 μl before LFT (n = 4 and n = 5, respectively), no inhibition of LTD was observed. A significant inhibition of LTD, however, was obtained when EGLU in a concentration of 20 mm(100 nmol)/5 μl was applied before LFT (Fig. 5D) (ANOVA: F(1,25) = 23.8; p < 0.001).

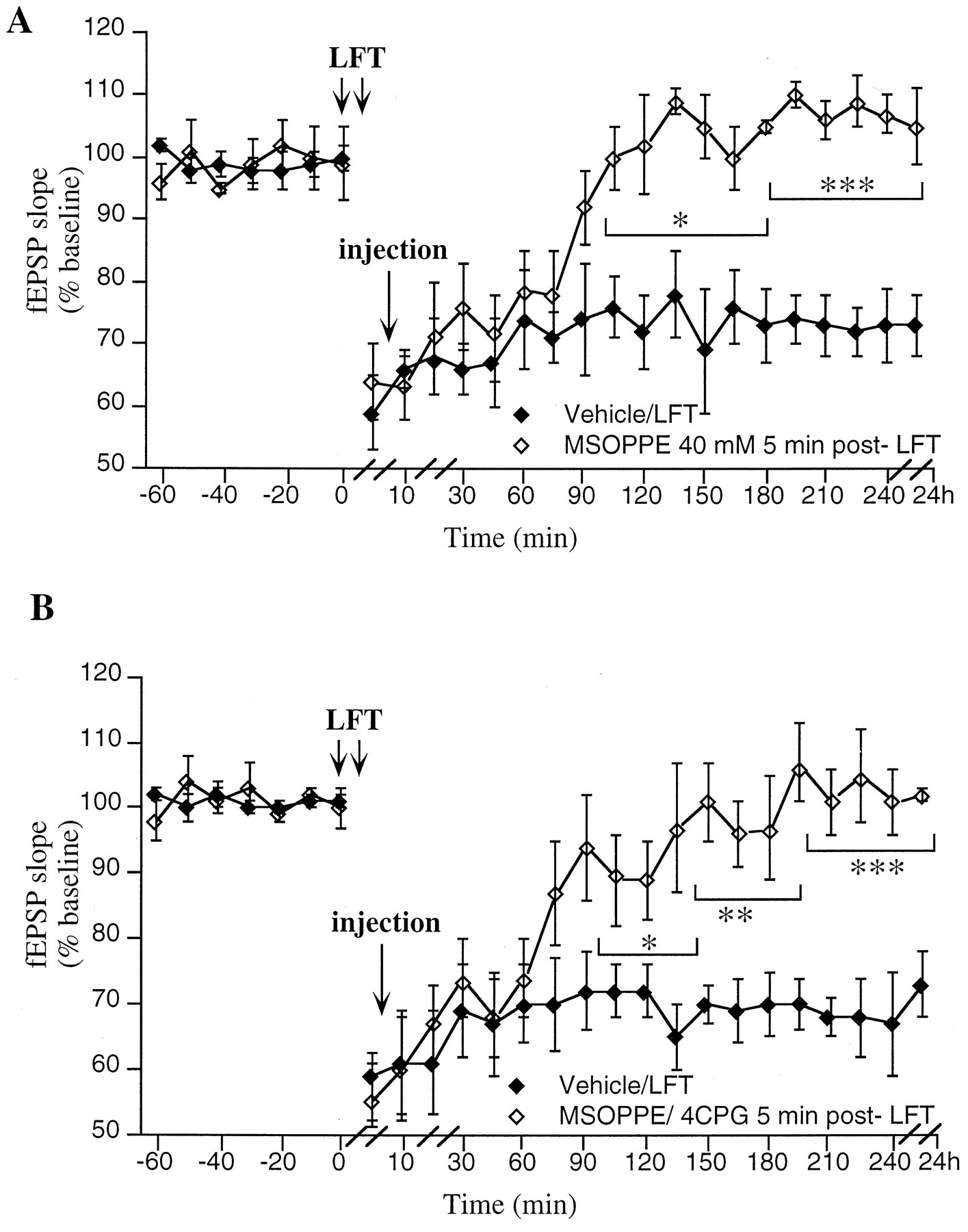
Group 1 and 2 mGluRs contribute to maintenance rather than initiation of LTD. A, Application of MSOPPE (40 mm/5 μl; n = 5) 5 min after LFT results in a significant inhibition of LTD, fromt = 105 min post-LFT compared with vehicle-injected controls (n = 5). B, Application of a subthreshold concentration of MSOPPE (20 mm/5 μl) in conjunction with 4CPG (20 mm/5 μl;n = 5) 5 min post-LFT results in a complete block of the 4CPG-resistant fraction of LTD, from t = 105 min compared with vehicle-injected controls (n = 5).
Although the magnitude of LTD immediately after LFT was not significantly different in drug (n = 6) and control (n = 11) groups, by 75 min post-LFT a significant inhibition of LTD had occurred in the EGLU group compared with controls (93 ± 7% compared with 75 ± 3% in controls;p < 0.05) (Fig. 4D). In the EGLU group the evoked potentials returned to preinjection values by 90 min post-LFT, whereas in the control group fEPSP values were still depressed at 24 hr post-LFT. When EGLU (20 mm/5 μl;n = 6) was administered and basal synaptic transmission was monitored for 4 hr subsequently, no effect on synaptic transmission was seen compared with controls (n = 8). In addition, no further improvement in the degree of inhibition of LTD was seen when a 40 mm concentration of EGLU was applied before LFT (n = 6).
Both group 1 and 2 mGluRs contribute to LTD maintenancein vivo
To examine whether MSOPPE inhibits LTD through prevention of initiation rather than maintenance of LTD, experiments were carried out in which the compound (40 mm/5μl) was applied 5 min after the termination of LFT. In this case, a profile of response was obtained similar to that seen when MSOPPE was applied before LFT (Fig.5A) (ANOVA: F(1,25) = 13.2;p < 0.001). Thus, although short-term depression was unaffected, MSOPPE (n = 5) produced an inhibition of LTD that became evident from 105 min post-LFT (100 ± 5% of preinjection fEPSP baseline values compared with 76 ± 5% in vehicle-injected controls; n = 5; p< 0.05).
In addition, when EGLU (20 mm/5 μl; n = 6) was applied 5 min after LFT, a significant inhibition of LTD occurred (ANOVA: F(1,25) = 11.7;p < 0.001), and a profile of LTD inhibition was seen similar to that obtained when EGLU was applied before LFT. The magnitude of LTD 5 min after LFT was not significantly different in drug (n = 6) and control (n = 6) groups (54 ± 9% of preinjection values compared with 57 ± 6% in controls). By 105 min post-LFT a significant inhibition of LTD had occurred in the EGLU group compared with controls (93 ± 7% compared with 73 ± 4% in controls; p < 0.05). In the EGLU group the evoked potentials returned to preinjection values by 120 min post-LFT, whereas in the control group values were still depressed at 24 hr post-LFT.
Taken together, these data further support the possibility that group 2 mGluRs are critically involved in the maintenance phase of LTD.
As a partial inhibition of LTD was obtained with 4CPG, we investigated whether application of MSOPPE could inhibit the 4CPG-resistant fraction of LTD. MSOPPE dose-dependently inhibits LTD, with a maximal effect at a concentration of 40 mm/5 μl (Fig. 4C). A concentration of MSOPPE that was subthreshold for significant inhibition of LTD (20 mm (100 nmol)/5 μl) was used therefore in conjunction with 4CPG (20 mm/5 μl). When LFT was applied in the presence of these compounds, a significant inhibition of LTD occurred (ANOVA: F(1,25) = 18.6; p < 0.001). In brief, when MSOPPE and 4CPG were applied together 5 min post-LFT, initially no difference between the drug (n = 5) and control groups (n = 5) was seen (Fig. 5B). By 105 min post-LFT, however, a significant inhibition of LTD developed. At this time point the fEPSP was 89 ± 4% of preinjection fEPSP baseline values in the MSOPPE/4CPG group and 72 ± 4% in controls. By 135 min post-LFT, fEPSP values had returned to preinjection fEPSP baseline levels in the drug group (Fig. 5B). The effect of MSOPPE/4CPG on LTD was significantly different from the effect of MSOPPE (20 mm/5 μl; n = 4) when injected alone (ANOVA: F(1,25) = 7.0;p < 0.001) and was significantly different compared with LTD evoked from animals in which 4CPG (20 mm/5 μl;n = 5) was applied 5 min after LFT (ANOVA:F(1,25) = 15.1; p < 0.001). These data indicate that activation of both group 1 and group 2 mGluRs contributes to LTD maintenance.
Group 2 mGluR activation is not necessary for LTPin vivo
When a vehicle injection (n = 6) was applied 30 min before HFT, LTP was induced that persisted for >24 hr (Fig.6A). This potentiation was statistically significant from control (nontetanized), fromt = 5 min post-HFT onward. When MSOPPE (n = 7) in a concentration of 40 mm (200 nmol)/5 μl was administered 30 min before an HFT was applied, LTP was induced that was not statistically significant from the vehicle-injected LTP group (Fig. 6A,B). Thus, a concentration of MSOPPE that inhibits the induction of LTD has no effect on LTP.

LTP induction does not depend on activation of group 2 mGluRs. A, The group 2 mGluR antagonist MSOPPE (n = 7) does not inhibit LTP in the CA1 region when compared with vehicle-injected controls (n = 6).B, Original analog traces showing evoked responses in the CA1 region at three time points: preinjection,t = 5 min, and t = 4 hr post-HFT, in an animal injected with MSOPPE (40 mm/5 μl). Line breaks (//) indicate change in time scale.
Group 2 mGluRs may play a differential role in hippocampal synaptic plasticity in vivo
To further investigate whether group 2 mGluRs may play a critical role in LTD as opposed to LTP, an experiment was carried out in which LFT was applied 1 hr after induction of LTP. When a vehicle injection (n = 5) was applied into the lateral cerebral ventricle and followed 30 min later by HFT, LTP occurred (Fig.7A) that was not statistically different from the previous control group (Fig. 6A). When LFT was given 1 hr after HFT, fEPSP values returned to pre-HFT baseline levels. When MSOPPE [40 mm (200 nmol)/5 μl;n = 5] was applied before HFT, a potentiation occurred similar to that seen in the control group. When LFT was given 1 hr later, no significant reduction in the magnitude of LTP was elicited. Thus, although LTD could follow LTP in the control animals, it was not possible to evoke LTD after LTP in the MSOPPE group (ANOVA:F(1,30) = 9.7; p < 0.001).

LTD but not LTP depends on activation of group 2 mGluRs. A, HFT in the presence of a vehicle injection (n = 5) results in a marked potentiation of fEPSP. LFT applied 1 hr after HFT causes evoked potentials to return to baseline values. MSOPPE (n = 5) applied before an identical protocol results in an LTP that cannot be reversed by application of LFT. B, Original analog traces showing evoked responses in the CA1 region at three time points: preinjection,t = 60 min, and t = 4 hr post-tetanus, in (1) an animal injected with MSOPPE (40 mm/5 μl) in which HFT is followed 1 hr later by LFT, and (2) an animal injected with MSOPPE (40 mm/5 μl) in which LFT is followed 1.5 hr later by HFT. C, Vehicle injection (n = 6) before LFT results in a marked depression of fEPSP. HFT applied 1.5 hr after LFT results in a reversal of depression to approximately pre-LFT values. MSOPPE (n = 6) applied before LFT results in a depression of LFT that recovers gradually. Application of HFT 1.5 hr after LFT produces a marked potentiation of fEPSP to LTP levels.Line breaks (//) indicate change in time scale.
To examine whether LTD could be followed by LTP, an experiment was performed in which LFT was applied before HFT. In the control group (n = 6), vehicle injection was followed 30 min later by LFT. A marked LTD was immediately noted, which was still present after 90 min. When HFT was then applied after stabilization of LTD (90 min post-LFT), a complete reversal of LTD to pre-LFT values was obtained (Fig. 7B). When MSOPPE (n = 6) was applied and followed 30 min later by LFT, an initial depression of synaptic transmission occurred, which by 90 min had recovered to pre-LFT values. When HFT was then applied, a marked potentiation of fEPSP occurred (Fig. 7B), which was similar in profile to the “HFT only” group (Fig. 6A,B). This potentiation was still present after 24 hr. Thus, although antagonism of group 2 mGluRs impairs LTD in vivo, LTP is not affected (ANOVA:F(1,32) = 48.3; p < 0.001). These observations support a differential role for group 2 mGluRs in distinct forms of hippocampal synaptic plasticity.
DISCUSSION
Initiation of LTD in the CA1 region in vivo is NMDA receptor dependent
It has been shown previously that administration of 900 pulses at 1 Hz will reliably produce robust input-specific LTD in vitro (Dudek and Bear, 1992; Mulkey and Malenka, 1992) and in anesthetized rats in vivo (Heynen et al., 1996). In this study, a similar stimulation protocol produced a persistent and marked LTD in freely moving rats. When the NMDA receptor antagonist AP5 was applied before LFT, however, a complete block of LTD occurred. This finding is in agreement with the reports of others that blockade of the NMDA receptor prevents homosynaptic LTD in the CA1 region in vitro (Dudek and Bear, 1992; Mulkey and Malenka, 1992) and supports the possibility that initiation of LTD in the hippocampal CA1 region in vivo is dependent on activation of NMDA receptors.
Induction of robust LTD in vivo is mGluR dependent
It has been reported that MCPG blocks LTD in the CA1 region (Bashir et al., 1993; Bolshakov and Siegelbaum, 1994), dentate gyrus (O’Mara et al., 1995), visual cortex (Haruta et al., 1994), and cerebellum (Hartell, 1994) of the rat in vitro, although others claim that MCPG has no effect on hippocampal LTD in vitro (Selig et al., 1995). MCPG remains a controversial compound however, because although some researchers have found it to be a competitive antagonist of both PLC and cAMP-coupled mGluRs (Eaton et al., 1993; Jane et al., 1993; Watkins and Collingridge, 1994), others have found no effect of this compound at these receptors (Chinestra et al., 1993; Saugstad et al., 1994). In previous work it was found that MCPG consistently blocks hippocampal LTP in freely moving rats (Manahan-Vaughan and Reymann, 1995a,b) and antagonizes the action of group 1 mGluRs (Manahan-Vaughan and Reymann, 1996; Manahan-Vaughan et al., 1996). Thus, it may be the case that the action of this compound may be strongly influenced by the conditions under which its efficacy is tested.
Other evidence to support the involvement of mGluRs in LTD has been provided using the group 1 and 2 mGluR agonist ACPD, which induces LTD in the cerebellum (Daniel et al., 1992) and dentate gyrus in vitro (O’Mara et al., 1995). Furthermore, in the present study a marked blockade of hippocampal LTD in vivo in the presence of MCPG was found. The effect became evident >1 hr after LFT was given, suggesting that, as is the case with LTP in vivo(Manahan-Vaughan and Reymann, 1995b; Manahan-Vaughan et al., 1996), mGluRs are critically involved in the induction of stable LTD downstream of the NMDA-dependent “early” component. Thus, the early phase of LTD, which depends on NMDA receptor activation, was present, but in the absence of mGluR activation LTD did not persist longer than ∼1 hr. This finding is in agreement with previous reports fromin vitro studies, which suggest that mGluRs are crucially involved in the maintenance of robust hippocampal LTD.
Group 1 mGluRs contribute to the stable induction of both LTD and LTP in vivo
To identify the subtype of mGluR that may be responsible for LTD induction, an investigation of the effect of the group 1 mGluR antagonist 4CPG (Birse et al., 1993; Hayashi et al., 1994; Watkins and Collingridge, 1994) on LTD was conducted. This drug has an IC50 value of 4 × 10−5m at mGluR 1 and an EC50 value of 5 × 10−4m at mGluR 2 and is inactive at mGluR 4 (Hayashi et al., 1994; tested in chinese hamster ovary cells). In addition it has been reported that 4CPG competitively inhibits ACPD-stimulated phosphoinositide hydrolysis in cerebrocortical slices (Birse et al., 1993; Eaton et al., 1993). Although its selective potency at mGluR 1 has been confirmed by others, it has been claimed that this compound is far less potent at mGluR 5 compared with mGluR 1 (Brabet et al., 1995). In the present study, it was found that although an impairment in the magnitude of LTD occurred in the presence of 4CPG, the remaining depression was still significantly different from control values, implying that only a partial inhibition of LTD took place. Others have reported that blockade of mGluR 1 in cultured Purkinje cells using antibodies (Shigemoto et al., 1994) and “knockout” of mGluR 1 in mouse cerebellum (Aiba et al., 1994a) result in an impairment of LTD. The data presented in this study also suggest a role for group 1 mGluRs in hippocampal LTD in vivo. The observation, however, that the partial blockade of LTD could not be improved on application of higher concentrations of 4CPG indicates that group 1 mGluRs are capable of modulating the degree of LTD but are not critical for hippocampal LTD induction.
The observation that 4CPG inhibits LTP in exactly the same concentration that impairs LTD is an interesting finding. Together with the observation that the compound is still effective when applied after induction of LTP or LTD, these data suggest that group 1 mGluRs are involved in the maintenance rather than the initiation of these forms of synaptic plasticity. Furthermore, these data indicate that group 1 mGluRs are capable of bidirectional modulation of synaptic plasticity. One possibility is that differential degrees of activation of group 1 mGluRs can determine which form of synaptic plasticity is expressed. This may occur perhaps through group 1 mGluR-mediated modulation of the level of activation of protein kinase C (Stanton, 1995). This bidirectional capability of group 1 GluRs could have an important significance in terms of the regulation of synaptic plasticity.
Group 2 mGluRs are critically involved in the stable induction of LTD but not LTP
The involvement of group 2 mGluRs in LTD and LTP was investigated by means of the selective group 2 mGluR antagonists MSOPPE (Jane et al., 1996a; Thomas et al., 1996a) and EGLU (Jane et al., 1996b; Thomas et al., 1996b). Both MSOPPE and EGLU exhibit a selective preference for group 2 mGluRs. Their KD values for antagonism of ACPD-induced depression of dorsal root-evoked monosynaptic excitation have been determined as 73 and 63 μm, respectively, at presynaptic (group 2) mGluRs. TheKD values of MSOPPE for antagonism ofl-AP4-induced depression of dorsal root-evoked monosynaptic excitation is 221 μm at presynaptic (group 3) mGluRs (Jane et al., 1996a), whereas EGLU has no detectable actions at group 3 mGluRs (Jane et al., 1996b; Thomas et al., 1996b). One cannot completely rule out, however, the possibility that group 3 mGluRs may be influenced by the concentration of MSOPPE used in the present study. In addition, the characterization of EGLU antagonism of presynaptic mGluRs was conducted using rat spinal cord preparations, in which the complement of group 3 mGluRs may not be identical to that of hippocampal group 3 mGluRs. Thus it may be the case that antagonism of group 3 mGluRs may contribute to the inhibition of LTD seen with MSOPPE and EGLU.
In the present study, a dose-dependent inhibition of LTD was obtained with MSOPPE. A similar block of LTD was obtained with EGLU, which is reported to be a more selective antagonist of group 2 mGluRs than MSOPPE (Jane et al., 1996b; Thomas et al., 1996b). Both MSOPPE and EGLU also effectively blocked the induction of LTD when applied 5 min post-HFT, which suggests that activation of group 2 mGluRs contributes to the maintenance or stabilization of LTD. Interestingly, neither MSOPPE nor EGLU had an effect on the induction or expression of LTP in vivo. The observation that both MSOPPE and EGLU can fully inhibit LTD but have no effect on LTP implies that activation of group 2 mGluRs may be critically required for LTD but unnecessary for LTP.
Group 2 mGluRs may play an exclusive role in LTD
The observation that MSOPPE inhibits LTD but not LTP in vivo provokes the interesting implication that the involvement of mGluRs in these two forms of synaptic plasticity is not simply dependent on the relative levels of activation of these receptors. This possibility is reinforced by the finding that it is possible in vivo to induce LTP and LTD in series, as has been reportedin vitro (Mulkey and Malenka, 1992; Dudek and Bear, 1993), but it is not possible to follow LTP with LTD, or to follow LTD with LTP, when MSOPPE is present. This suggests that distinct mGluR subtypes may be critically involved in one form of synaptic plasticity as opposed to another. With regard to LTP, this question was addressed previously, and it was found that group 1 mGluRs seem to be the critical mGluR class involved, whereas group 3 mGluRs may play merely a modulatory role in this phenomenon (Manahan-Vaughan and Reymann, 1995a,1996; Manahan-Vaughan et al., 1996). The requirement of group 1 mGluRs for LTP has also been demonstrated in mGluR1 knockout mice (Aiba et al., 1994b). Furthermore, the observations of the present study offer strong evidence to support the theory that group 2 mGluRs are critically involved in the maintenance of LTD but not LTP.
This finding, combined with the observation that group 1 mGluRs contribute to LTD maintenance, implies that both a presynaptic (group 2 mGluR) and a postsynaptic (group 1 mGluR) locus for LTD expression exists. It has been shown that induction of hippocampal LTD requires Ca2+ release from distinct pre- and postsynaptic stores (Reyes and Stanton, 1996). Perhaps group 1 mGluRs contribute to LTD by stimulating release of Ca2+ from postsynaptic inositol trisphosphate-gated intracellular Ca2+ stores, whereas group 2 mGluRs influence release of Ca2+ from the presynaptic ryanodine-sensitive Ca2+ stores identified byReyes and Stanton (1996).
Conclusion
The results of this study comprise a number of interesting observations about LTD in vivo. Namely, (1) it is possible to obtain a robust LTD in the CA1 region of freely moving rats; (2) this LTD is dependent on activation of NMDA receptors; (3) in addition, activation of mGluRs is required for a persistent form of LTD to occur; (4) although group 1 mGluRs seem to modulate the degree of LTD, group 2 mGluR activation may be a critical factor for LTD maintenance; and (5) it seems that group 2 mGluRs are involved in LTD but not LTP induction, whereas group 1 mGluRs are involved in the induction or modulation of both phenomena.
Taken together, the observations of this study demonstrate an intriguing involvement of mGluRs in synaptic plasticity and suggest that bidirectional modulation of hippocampal synaptic transmission by distinct mGluR subtypes can occur. Furthermore, it may be the case that certain mGluR subtypes modulate distinct forms of plasticity only. Thus, mGluR involvement in LTD and LTP may correspond to an important system by which the efficacy of synaptic transmission is regulated within the hippocampus.
Footnotes
I am very grateful to Ms. Silvia Vieweg for expert technical assistance and to Professor Klaus Reymann and Dr. Ritchie Brown for valuable discussion of this manuscript.
Correspondence should be addressed to Dr. Denise Manahan-Vaughan, Federal Institute for Neurobiology, Department of Neurophysiology, Brenneckestrasse 6, P.O. Box 1860, D-39008 Magdeburg, Germany.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}