


Abstract
The genetic mechanisms underlying cisplatin (DDP) resistance in yeast were investigated by examining the cytotoxicity of DDP toSchizosaccharomyces pombe mutants that were either hypersensitive or resistant to Cd. Despite reports that have linked glutathione (GSH) to DDP resistance in human cancer cells, we found that a mutant of S. pombe that was hypersensitive to Cd by virtue of a 15-fold reduction in GSH level and lack of phytochelatin production was as tolerant as the wild-type strain to DDP. A mutant that harbored a mutation in hmt1, the gene encoding an ATP-binding cassette-type transporter for vacuolar sequestration of a phytochelatin/Cd complex, exhibited only mild hypersensitivity to DDP even though it was 100-fold more sensitive to Cd. Overexpression ofhmt1 in wild-type or mutant cells conferred tolerance to Cd but failed to do the same for DDP. However, a strain that produced 6-fold more sulfide than wild-type cells was found to be 6-fold more resistant to DDP and twice as resistant to Cd; an association between DDP resistance and sulfide production was observed in three other strains that were examined, and overproduction of sulfide was accompanied by reduced platination of DNA. These results indicate that GSH and the GSH-derived phytochelatin peptides do not play critical roles in determining sensitivity to DDP in S. pombe but rather identify increased production of sulfide as a possible new mechanism of DDP resistance that may also be relevant to human cells.
The chemotherapeutic agent DDP is widely used for the treatment of ovarian, head and neck, and small cell lung cancer as well as a variety of squamous cell cancers; however, the development of resistance during treatment remains a major obstacle to its effective use. The mechanisms that underlie the development of acquired resistance to DDP in human tumors are not well understood. There have been reports that DDP and Cd share detoxification mechanisms in mammalian cells. Despite the chemical differences between Cd and DDP, both are capable of reacting with nucleophilic sites on small molecules such as thiol compounds and large molecules such as metallothioneins and DNA. Overproduction of metallothioneins protects mammalian cells from Cd toxicity and, under some circumstances, has been reported to protect against DDP toxicity as well (1). GSH also seems to play a role in the defense against Cd and DDP. Depletion of GSH with buthionine sulfoximine increases the sensitivity of human cells to both metals (2, 3), and in some cell lines selected for resistance to DDP, there is an associated increased in GSH content (4). To gain insight into whether the two cytotoxins might share some common defense mechanisms, we examined the link between Cd tolerance and DDP resistance in a fission yeast model system.
In yeasts, several resistance mechanism are known that are activated on exposure to toxic metals, and molecular genetic analysis has been used to identify some specific genes involved in heavy metal detoxification pathways (5). In S. cerevisiae, the CUP1 gene, which codes for a copper-metallothionein, mediates resistance to Cd (6). In contrast, the fission yeast S. pombe lacks metallothioneins (7) but responds to heavy metal stress with increased synthesis of metal-binding peptides enzymatically synthesized from GSH (7). These peptides, known as PCs, are frequently found in the plant kingdom and have the general structure of (γ-Glu-Cys)nGly, where n is usually 2–4. PCs with as many as 11 γ-Glu-Cys units have been reported (8-10). Candida glabrata uses both the metallothionein- and the PC-mediated detoxification pathways and activates copper-metallothio-nein production in response to Cu and PC production in response to Cd (5).
In the case of Cd, PCs form a loose complex with the metal, which is then sequestered into the vacuoles, where a more stable complex is formed consisting of PCs, Cd, and acid-labile sulfide (11, 12). Mutations that decrease the production of PCs, including those that affect GSH levels (13), the vacuolar sequestration of the PC/Cd complex (12), or the production of Cd-induced sulfide (14), result in a Cd-hypersensitive phenotype. In addition, certain mutants of C. glabrata and S. pombe can achieve Cd resistance through the anomalous production of high sulfide levels (15).
The aim of the current study was to determine whether alterations that affect the detoxification of Cd in S. pombe would also affect sensitivity to DDP. Our results indicate that the level of GSH, which is critical for Cd tolerance, does not significantly affect DDP sensitivity. The vacuolar sequestration mechanism mediated by PCs likewise does not play a significant role in DDP tolerance. However, the high level of production of sulfide found in one mutant is capable of reducing the sensitivity of the cells to both DDP and Cd. Thus, there is at least one common resistance mechanism shared by both DDP and Cd. Because a defect in the assimilatory sulfate reduction pathway does not affect DDP cytotoxicity, it is possible that the mutation conferring resistance to both Cd and DDP resides in a different sulfide-generating pathway.
Materials and Methods
Reagents.
DDP was obtained from Bristol-Myers Squibb (Princeton, NJ). CdCl was purchased from Sigma Chemical (St. Louis, MO).
Genetic materials and manipulation.
The adenine prototropicS. pombe strain Sp223 (h − leu1.32 ura4.294 ade6.216) was the wild-type progenitor from which all strains other than DS5 were derived: LK100 is isogenic to Sp223 but is hmt1 − and has been described previously (12). Also, the adenine prototropic JS21 (h − leu1.32 ura4.294) is the progenitor of LK37, a GSH-deficient mutant; of Y10, a strain that hyperproduces sulfide; and of DS31, a sulfite reductase mutant constructed through gene disruption of the sulfite reductase gene.1 DS1 and DS5, which contain engineered disruptions of the ade2 locus in the Sp223 and B1048 (h + ura4.294 ade7.50) backgrounds, respectively, produce abnormally low levels of sulfide on Cd induction and fail to accumulate the high-molecular-weight PC/CdS complex (14). Cultures were grown at 30° in YES medium (consisting of 5 g of Difco yeast extract, 30 g of glucose/liter, 0.075 g of adenine/liter, and 0.075 g of uracil/liter). For construction of strains carrying either the yeast expression vector pART1 or thehmt1 expression construct pDH35, cells were transformed through electroporation (13), and transformants were selected in minimal medium supplemented with amino acids other than leucine.
Measurement of PC/Cd complexes.
PC/Cd complexes were measured as described elsewhere (15). Briefly, LK37 and wild-type cells were cultured in standard medium and induced with 0.2 mm Cd for 30 hr. A cell-free extract containing 2 mg of protein was fractionated on a Sephadex G50 column, and the Cd concentration of the fractions was measured with the use of atomic absorbance spectroscopy. The first and last peaks shown in Fig. 1A represent the excluded and included volumes, respectively. The middle two peaks contain high- and low-molecular-weight PC/Cd complexes, respectively.
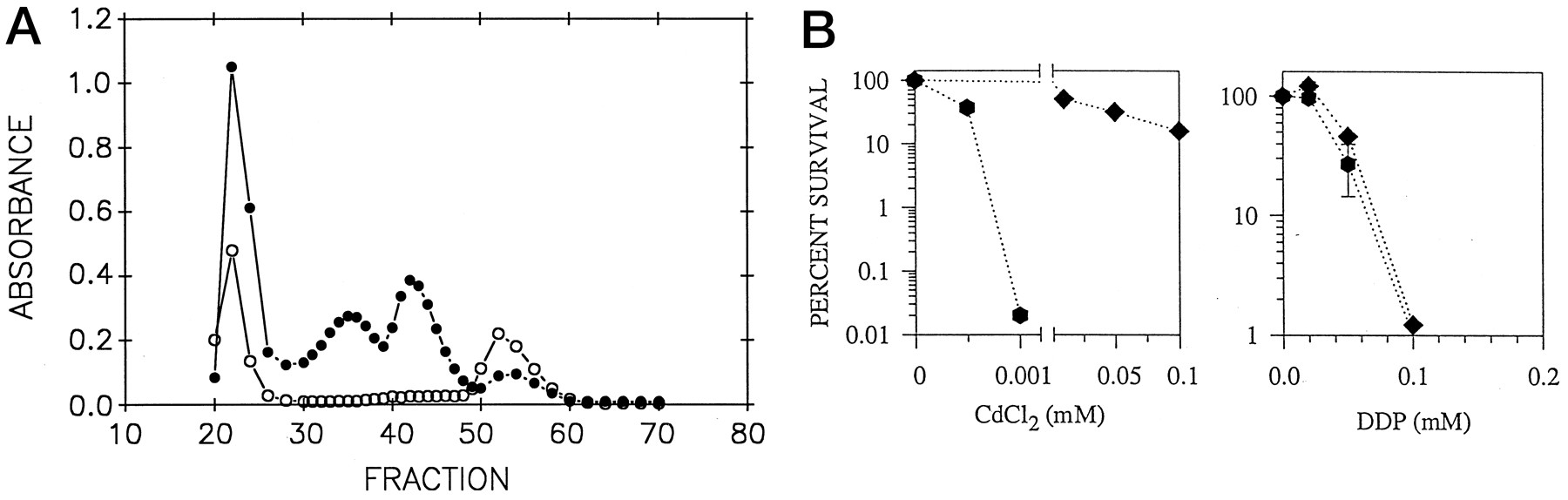
A, PC profile of LK37 and parental wild-type cells. The middle two peaks represent the high and low molecular weight PC/Cd complexes, respectively. •, Parental wild-type cells. ○, LK37. B, Plating efficiency (number of colony-forming units) of S. pombe strains JS21 (GSH proficient) and LK37 (GSH deficient) after a 48-hr exposure to graded concentrations of CdCl2and DDP. ♦, JS21 (GSH proficient). ⬡, LK37 (GSH-deficient).Data points, mean values from two or three independent experiments performed with triplicate cultures. Vertical lines, standard deviation.
Metal sensitivity assay.
To evaluate the toxicity of DDP or Cd, the viability of cells after drug exposure was determined by clonogenic assay. Specifically, 3-ml cultures containing a total of 2.5 × 105 cells were exposed for 48 hr to graded concentrations of either DDP or Cd and subsequently plated at various dilutions onto 60-mm agar plates lacking DDP or Cd and incubated for 4 days at 30°. The IC50 was defined as the drug concentration that reduced the number of colony-forming units to 50% of the value obtained from a control culture not exposed to drug. Each experiment was repeated two or three times with triplicate cultures for each drug concentration.
GSH level.
The intracellular GSH levels in exponentially growing cells were determined through conjugation of GSH with monobromobimane followed by high performance liquid chromatography quantification as previously described (16). Each experiment was performed three times with triplicate samples.
DDP accumulation and DNA platination.
The uptake of DDP was measured after a 4-hr exposure of 3-ml cultures containing a total of 1 × 106 cells to 1.6 mm DDP at 30°. After incubation with DDP, the cells were washed four times with cold phosphate-buffered saline at 4°, and an aliquot was removed for cell counting using a hemocytometer. The Pt content of the remaining cells was quantified through atomic absorption spectrometry. Pt in DNA was measured using the same technique after extraction of DNA as previously described (17).
Sulfide production.
Sulfide production was determined as previously described (14). Briefly, a 50-ml culture was grown in a side-arm flask capped with a rubber stopper through which a glass pipette was inserted. The external pipette was attached to a sterile air filter through which humidified air was supplied to the culture by being bubbled through H2O. The side arm of the flask was connected to a tube containing ZnOAc to trap the volatile sulfide. After 24 hr, 10 ml of HCl was added to the culture to release intracellular sulfide. Total (extracellular and intracellular) sulfide was measured in a colorimetric assay usingN,N-dimethyl-p-phenylene diamine HCl as a substrate, and normalized values were expressed relative to the level for the wild-type strain.
Northern analysis.
Total RNA was prepared as described previously (18), and 20 μg was fractionated through electrophoresis on a formaldehyde-containing 1% agarose gel and transferred to a nylon membrane. DNA probes were labeled with a random primer kit (Amersham, Little Chalfont, UK). Hybridization was carried out as previously described using the hmt1 probe retrieved from pDH35 through treatment with EcoRI (12). A leu1 probe (kindly provided by Dr. Paul Russel, Scripps Institute, La Jolla, CA) was used as a control for DNA loading.
In vitro interaction between DDP and sulfide.
We incubated 1 mm DDP with different concentrations of Na2S at room temperature, and the two compounds were allowed to react overnight. After centrifugation, the supernatants were assayed for content of Pt by atomic absorption spectrometry.
Results
Role of GSH.
GSH plays a major role in the detoxification of xenobiotics in eukaryotic cells. However, other than the associations that GSH depletion sensitizes human cells to DDP and that high GSH content is found in some resistant cell lines (1, 3, 19), the role of GSH in DDP detoxification remains unclear. GSH is known to play an indirect role in Cd tolerance in S. pombe. Because PCs are produced enzymatically from GSH, a reduction in cellular GSH content results in subnormal production of PCs (20, 21). One such mutant is LK37, which was isolated as a Cd-hypersensitive strain from the parental JS21 strain. Fig. 1A shows that LK37 contains no detectable high- or low-molecular-weight PC-bound Cd complexes. Based on measurements made using the monobromobimane conjugation assay, LK37 cells were found to have a 15-fold reduction in GSH level (GSH content was 0.86 ± 0.17 and 0.059 ± 0.012 μmol/106cells for the JS21 and LK37 cells, respectively). Fig. 1B shows a comparison of the sensitivity of the GSH-deficient LK37 and GSH-replete JS21 cells to DDP; both strains exhibited the same concentration-survival curve. This contrasts with the substantial difference in tolerance to Cd, with LK37 being clearly more sensitive to this metal. These results indicate that the wild-type level of neither GSH nor PCs significantly influences the sensitivity ofS. pombe to DDP.
Role of PC-mediated sequestration.
After exposure of cells to Cd, the induced production of PCs is followed by the vacuolar sequestration of PC-bound Cd into the vacuole. This process is mediated by an ATP-binding cassette-type transport pump that resides in the vacuolar membrane (12, 22). The gene encoding this transporter is known as hmt1, and the mutant harboring a nonsense codon mutation in this gene is LK100. As reported previously, LK100 is hypersensitive to Cd (12). Fig. 2 shows the survival curve for LK100 and its isogenic parent, Sp223, exposed to Cd and DDP. Based on IC50 values of 0.00092 and 0.1 mm (standard deviation < 5%) for hmt1 − LK100 andhmt1 + Sp223, respectively, LK100 is 108-fold more sensitive to Cd. In contrast, LK100 exhibited only 3-fold greater sensitivity to DDP (LK100: IC50 = 0.015 mm; Sp223: IC50 = 0.054 mm; standard deviation < 5%). The slight sensitivity exhibited by thehmt1 − strain to DDP could indicate a role for this gene in protection against DDP; however, it is also entirely possible that the defect in hmt1 has disrupted some other aspect of metal homeostasis and reduced the overall fitness of this strain, particularly when subjected to stress.

Plating efficiency of S. pombestrains Sp223 (hmt1 +) and LK100 (hmt1 −) after a 48-hr of exposure to graded concentrations of CdCl2 and DDP. •, LK100. ▪, Sp223.Left, values are from a representative experiment.Right, data points, mean values from three experiments performed with triplicate cultures. Vertical lines, standard deviation (where absent, standard deviation was less than the size of the symbol).
Overproduction of the hmt1-encoded transport pump is known to enhance Cd tolerance, presumably as a result of increased vacuolar sequestration of Cd (12). We sought to determine whether overproduction of hmt1 would also produce resistance to DDP. Sp223 and LK100 cells were transformed with the hmt1 cDNA expression construct pDH35 or, as a control, the empty vector pART1. Fig.3 shows that the pDH35 transformants indeed produced a marked increase in the accumulation ofhmt1 mRNA. However, although Cd tolerance was enhanced,hmt1 overexpression failed to significantly alter the sensitivity to DDP of either strain (Fig. 4). Thus, despite the important role for hmt1 in Cd tolerance, neither loss of nor increase in hmt1 expression had a significant impact on DDP tolerance.

Northern blot analysis of total RNA from LK100 and Sp223 cells carrying pDH35 (hmt1 + cDNA) or pART1 (empty vector). Twenty micrograms of total RNA was loaded in each lane. The hmt1 cDNA was used as a hybridization probe, whereas the leu1 gene probe was used as a control to document RNA lane loading. Basal levels of hmt1 message were detectable when the blot was exposed for longer periods of time.
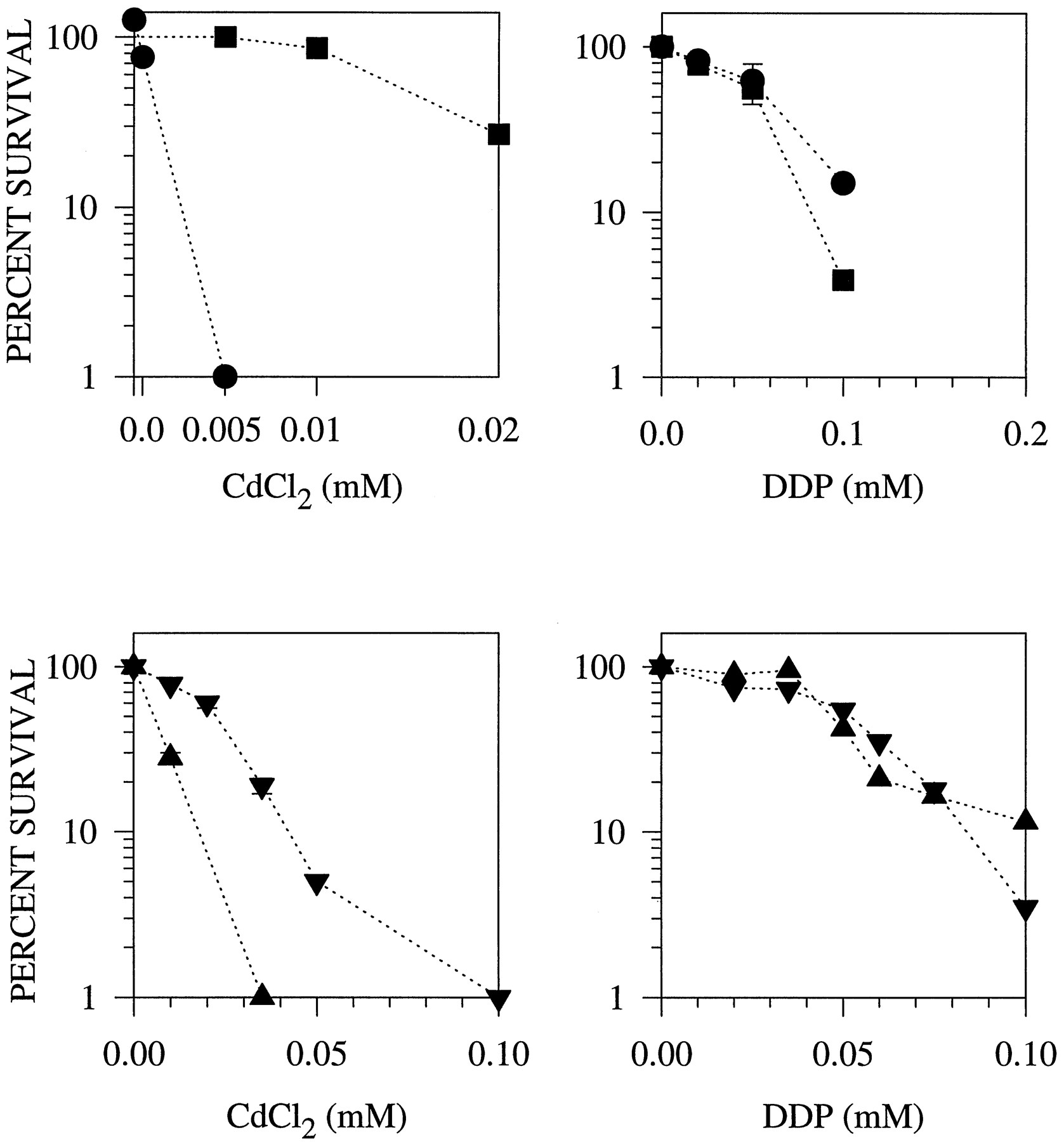
Clonogenic survival as a function of concentration after a 48-hr exposure to CdCl2 and DDP ofhmt1 − LK100 andhmt1 + Sp223 harboring pART1 (empty vector) or pDH35 (hmt1 + cDNA) (Sp223pDH35#1 and LK100pDH35#2). •, LK100 harboring pART1. ▪, LK100 harboring pDH35. ▴, Sp223 harboring pART1. ▾, Sp223 harboring pDH35. Data points, mean values from two or three independent experiments performed with triplicate cultures. Vertical lines, standard deviation.
Sulfide-associated DDP resistance.
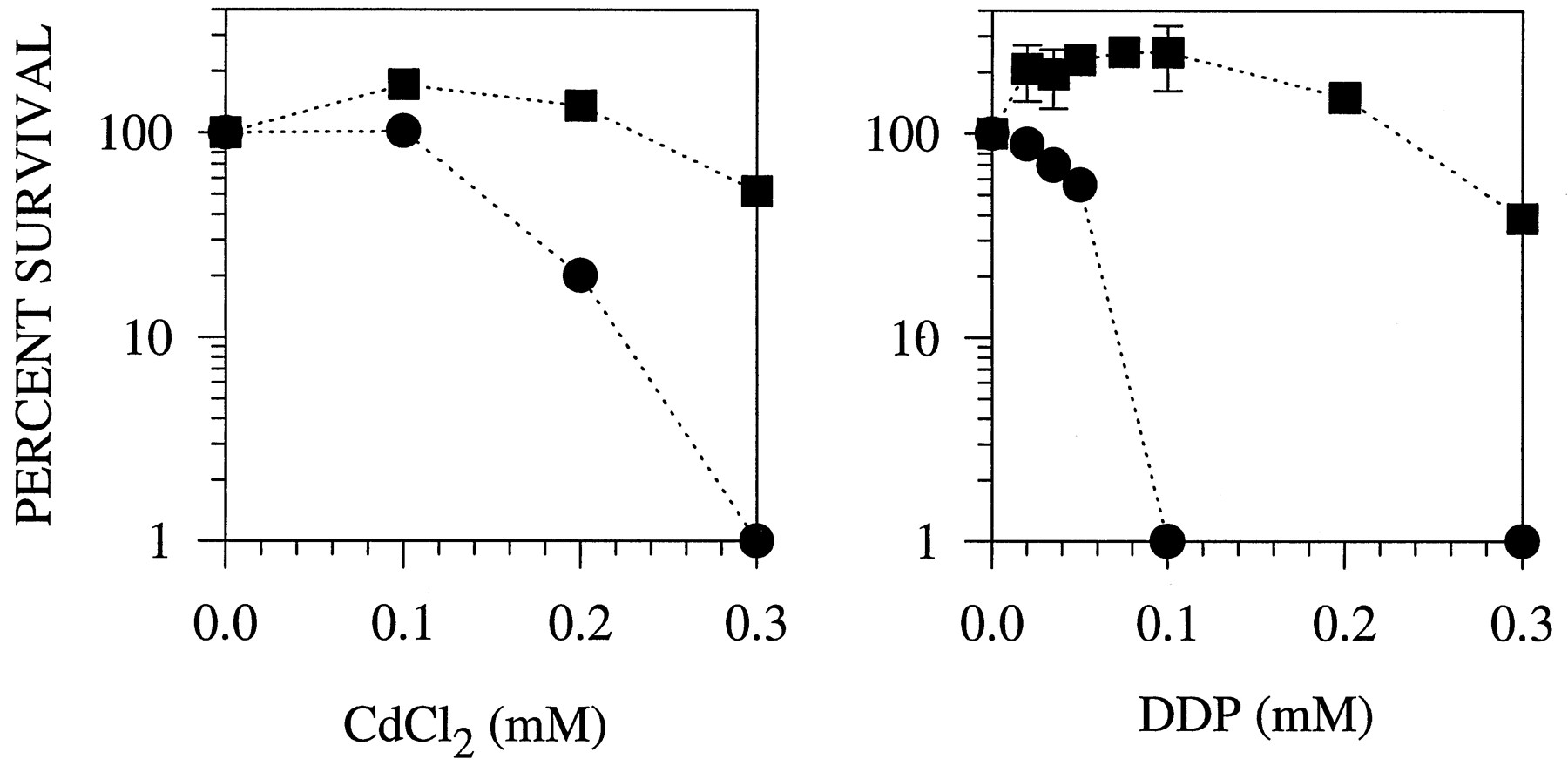
Certain mutants that form yellow colonies on Cd plates also show greater tolerance to Cd. Like the mutants reported for C. glabrata, these mutants produce anomalously high amounts of sulfide (15). One such mutant is Y10, which had a sulfide production rate that was 5.99 ± 0.85-fold greater than that of the parental strain JS21 (values are given as mean ± standard deviation). The abnormally high sulfide production rate was not accompanied by a correspondingly high level of GSH. The GSH levels found in Y10 and its isogenic parent JS21 were 1.01 ± 0.21 and 0.86 ± 0.17 mmol/106 cells, respectively, reflecting no significant difference in GSH content between the strains. Fig.5 shows that when Y10 was exposed to Cd or DDP for 48 hr, it demonstrated a 2-fold increase in the IC50 value for Cd and a 6-fold increase in the IC50 value for DDP compared with its parental strain. Examination of a second strain that had a 3.1 ± 0.3-fold increase in sulfide production demonstrated it to be 2.7-fold resistant to DDP.

Clonogenic survival of the S. pombewild-type strain JS21 and the sulfide-overproducing strain Y10 after a 48-hr exposure to CdCl2 and DDP. •, JS21. ▪, Y10.Data points, mean values from three to six independent experiments performed with triplicate cultures. Vertical lines, standard deviation.
These findings suggest that sulfide either has a direct role in the protection of S. pombe from DDP and Cd cytotoxicity or is a marker for the primary alteration that controls sensitivity. In the case of Y10, the yellow coloration of the colonies on Cd-containing medium suggested that CdS was formed. We observed similar interactions occurring between DDP and sulfide ions (see below). Interestingly, low concentrations of both Cd and DDP stimulated the survival of sulfide-overproducing Y10 cells. A reasonable interpretation is that at concentrations of the metals that are themselves not toxic, the sequestration of the sulfide as metal/sulfide complexes can ameliorate the growth-suppressing effect of the anomalously high sulfide level.
To determine whether DDP could in fact form a complex with sulfide, 1 mm DDP was incubated with concentrations of Na2S ranging from 0.25 to 4 mm. The formation of a brown precipitate was observed, which was directly proportional to the amount of exogenously provided sulfide. The chemistry of the two reactants suggests that the precipitate consisted of PtS. This interpretation is consistent with atomic absorption measurements presented in Fig. 6 that show a decrease in the concentration of soluble Pt remaining in the supernatant as a function of the concentration of Na2S.
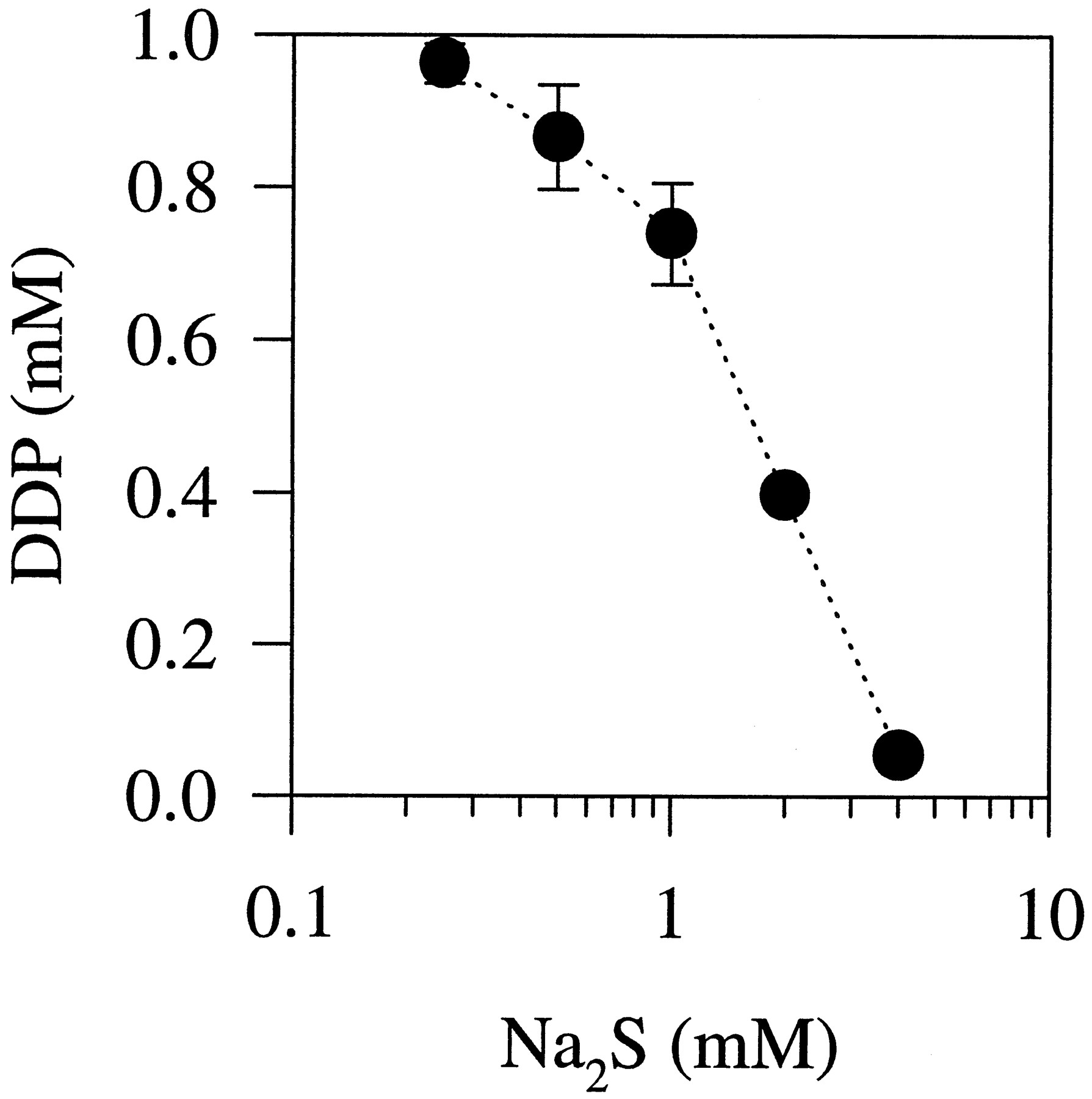
In vitro interaction between DDP and Na2S. DDP (1 mm) was incubated with different concentrations of Na2S. The DDP remaining in solution after removal of precipitation is expressed as a function of Na2S concentration. Data points, average values from two independent experiments. Vertical lines, standard deviation.
Despite the lack of detailed knowledge on what seems to be multiple sulfide-generating pathways in fission yeast, much is known about the assimilatory pathway by which sulfate is reduced to sulfite and then sulfide (23). A mutant of this pathway, DS31, fails to convert sulfite to sulfide because of a genetically engineered disruption of the sulfite reductase gene.1 DS31 cells fail to form the sulfide needed for assimilation into cysteine and are therefore cysteine auxotrophs. Despite cysteine supplementation, however, cysteine auxotrophs are known to be sensitive to Cd and fail to form sufficient amounts of PCs. Presumably, the rate of import of cysteine is inadequate to meet the high cysteine demand for PC biosynthesis. As with other cysteine auxotrophs, Fig. 7 shows that DS31 proved to be hypersensitive to Cd, even when this amino acid was available in the growth medium. However, DS31 was as tolerant as the wild-type strain to DDP. This suggests that the sulfide generated by the assimilatory sulfate reduction pathway is not required for the wild-type level of DDP tolerance. In contrast, two other mutants that have defects in sulfide production were hypersensitive to DDP. These two mutants, DS1 and DS5, are Cd hypersensitive and fail to accumulate the high-molecular-weight PC/CdS complex due to mutations that block inducible sulfide production during Cd stress (14, 24) The Cd-inducible production of sulfide is not attributable to the assimilatory sulfate-reduction pathway but rather to another pathway that involves enzymes of the de novo purine biosynthesis pathway. When exposed to DDP, DS1 and DS5 were found to be 4.0- and 3.6-fold more sensitive to DDP, respectively, than the wild-type control. Thus, there seems to be an interesting link between Cd-inducible sulfide production and DDP tolerance.

Plating efficiency of JS21 (wild-type) and DS31 (sulfite reductase mutant) after a 48 hr exposure to CdCl2. ▴, JS21. ▾, DS31. Data points, mean values from two independent experiments. Vertical lines, standard deviation.
Cellular accumulation of DDP.
To determine whether the observed differences in DDP cytotoxicity among the tested strains could be attributed to differential DDP uptake, accumulation studies were performed. The S. pombe strains were exposed to 1.6 mm DDP for 4 hr, a period sufficient for any changes in uptake resulting from exposure to the metal ion to have occurred. Marked differences were not found among the different strains. DDP accumulation in these strains, expressed as micrograms of Pt/106 cells (mean ± standard deviation) was as follows: JS21, 0.08 ± 0.01; LK37, 0.13 ± 0.03; Sp223, 0.12 ± 0.02; LK100, 0.17 ± 0.03; and Y10, 0.05 ± 0.01. This lack of significant difference was not surprising for LK37 and JS21, which exhibited no difference in DDP sensitivity. The fact that there was no substantial reduction in Pt accumulation in the quite resistant sulfide-overproducing Y10 strain indicates that the underlying mechanism of DDP resistance cannot be attributed to changes in overall DDP accumulation in this strain either.
The extent of DNA platination produced by a 4-hr exposure to 1.6 mm DDP was compared for the parental JS21 and sulfide-overproducing Y10 strain to determine whether even in the absence of an effect on total cellular accumulation, the sulfide antagonized the ability of the drug entering the cell to platinate DNA. DNA platination was 0.0219 ± 0.0011 nmol of Pt/μg of DNA for the JS21 cells and 0.0138 ± 0.0022 nmol of Pt/μg of DNA for the Y10 cells (p = 0.007), indicating that the sulfide overproduction was associated with a 34% reduction in the ability of DDP to reach the nucleus and react with DNA.
Discussion
The aim of this study was to determine whether known mechanisms that yeast cells use to protect themselves against toxic metal ions also play a role in the protection against DDP. Cd is of particular interest in this regard because, like the cytotoxic hydrated form of DDP, it is capable of complexing with other molecules that have nucleophilic sites, particularly thiols. Recent observations suggest that there is cross-resistance in human cells between DDP and a variety of toxic metal ions, including arsenite and antimonite (25). Based on the assumptions that the resistance mechanisms might be conserved at the cellular level among diverse organisms and that other reactive metal ions may share detoxification pathways with DDP, we chose to examine the effect of six different mutations known to alter the tolerance of S. pombe to Cd ions.
The role of GSH in the DDP-resistant phenotype remains enigmatic; however, the results obtained in the current study do not support an important role for GSH as a major determinant of DDP sensitivity inS. pombe cells. No appreciable difference in DDP sensitivity was found in a strain that had a 15-fold reduction in GSH level and produced no detectable PCs. The fact that Y10 was 6-fold more resistant to DDP but had a GSH content that was similar to its DDP-sensitive wild-type parent lends further support to the idea that there is no correlation between GSH content and DDP resistance in S. pombe. This finding was somewhat surprising in view of the fact that reduction in GSH through inhibition of its synthesis with buthionine sulfoximine reliably sensitizes human cells to DDP (2, 3,26-28). It is noteworthy that although overproduction of GSH has been documented in some DDP-selected human cell lines, this is not a universal observation. Even in cells in which increased GSH levels have been found, it has not been established that GSH per se is the proximate cause of the DDP resistance. The increase in GSH level could be the result of a deficit in another pathway whose dysfunction results in dysregulation of both GSH level and DDP resistance.
GSH is needed for Cd tolerance in plants and in S. pombebecause metal-binding PC peptides are produced through the polymerization of GSH (29, 30). Resistance to Cd in cultured tomato cells has been linked to an increase in PC content (31), whereas hypersensitivity to Cd in Arabidopsis thaliana correlates with reduced accumulation of PCs (32).1 The hmt1gene product in S. pombe mediates vacuolar sequestration of Cd, and abundance of this transport protein shows positive correlation with the degree of Cd resistance (12). Therefore, the PC/hmt1 pathway is critical for detoxification of Cd. However, although loss of hmt1 function in S. pombe resulted in a 108-fold increase in Cd sensitivity, it produced a minimal effect on DDP sensitivity. Therefore, this excludes the PC/hmt1 pathway as a major detoxification mechanism for DDP, although it remains possible that it plays a minor role, perhaps serving to sequester DDP/sulfide complexes.
The most interesting observation to emerge from this study is that sulfide overproduction in Y10 was associated with enhanced resistance to both Cd and DDP. Although the magnitude of resistance to Cd was not as great, it was quite substantial for DDP. This shows that the two compounds can be detoxified through a common mechanism involving either sulfide directly or some elements of its biosynthetic pathway. It is noteworthy that low concentrations of either Cd or DDP effectively increased the survival of Y10, which is consistent with the hypothesis that sulfide overproduction may be harmful to these cells and that metal complexation serves to reduce the concentration of free sulfide.
In plants and in microorganisms, including S. pombe, but not in animals, sulfide can be produced by the well-defined assimilatory sulfate reduction pathway, in which inorganic sulfate is added to ATP to produce adenosine phosphosulfate, which is then further converted until sulfide is formed. We determined whether this pathway could affect DDP tolerance by testing a mutant with an engineered defect in the sulfite reductase gene. The results show that a sulfite reductase mutant does not exhibit greater DDP sensitivity. Therefore, this excludes the assimilatory reduction pathway as an important determinant of the wild-type level of DDP tolerance. That is, wild-type cells must possess mechanisms for maintaining normal levels of tolerance that do not depend on sulfide generated through assimilatory sulfate reduction, and thus loss of function of this pathway does not result in hypersensitivity. In the sulfide-overproducing variants, however, the increase in sulfide above normal levels could be the basis for a novel “gain of function” mechanism of DDP resistance. Because there are multiple pathways that generate sulfide, it remains possible that a genetic alteration outside the assimilatory pathway is responsible for sulfide overproduction in these mutants. We cannot exclude the possibility that sulfide overproduction is simply a symptom of an underlying metabolic change that directly mediates DDP resistance. However, an investigation of the genetic defect in Y10 should uncover the pathway responsible for its sulfide overproduction, and this pathway would then be the subject of fruitful study. We are currently isolating the gene that causes sulfide overproduction in Y10.
Concerning the issue of which sulfide-generating pathway is responsible for the enhanced DDP resistance observed in Y10, it is of interest to note that high sulfide production during Cd stress is not attributable to the assimilatory sulfate reduction pathway. During Cd stress, intracellular sulfide levels increase by 5–7-fold concomitant with formation of the vacuolar high-molecular-weight PC/CdS complexes. This Cd-induced sulfide production is also observed in the sulfite reductase mutant DS31 as long as cysteine is available in the growth medium.1 Therefore, Cd-induced sulfide production can proceed from a step downstream of the assimilatory sulfate reduction pathway. This is consistent with the previously proposed hypothesis that Cd-induced sulfide is generated through a novel pathway. This novel pathway is not well defined but has been postulated to begin with the conversion of cysteine to cysteine sulfinate, which is then incorporated into purine precursors (14, 24).
It is not currently known which sulfide pathway is genetically altered in Y10 to produce the anomalous high level of sulfide. Because humans do not assimilate inorganic sulfate, the generation of sulfide would necessarily be due to pathways that degrade sulfur compounds, such as cysteine or GSH. The findings reported here serve to redirect attention from GSH to other sulfur-containing constituents. Through the deciphering of the pathways in S. pombe, homologous pathways in human cells could be revealed as being important in the determination of tumor sensitivity to this important chemotherapeutic agent.
Acknowledgments
We would like to thank Bristol-Myers Squibb for the kind gift of the DDP used in these studies.
Footnotes
- Received July 8, 1996.
- Accepted October 14, 1996.
-
Send reprint requests to: Paola Perego, Ph.D., Experimental Oncology B, Istituto Nazionale Tumori, via Venezian 1, 20133 Milano, Italy. E-mail: perego{at}icil64.cilea.it
-
↵1 D. M. Speiser, K. F. McCue, and D. W. Ow, unpublished observations.
-
This work was supported in part by the Associazione Italiana per la Ricerca sul Cancro (P.P.) and the United States Department of Agriculture Agricultural Research Service Project 5335–21000-009-00D (D.W.O.). This work was conducted in part by the Clayton Foundation for Research, California Division. S.B.H. is a Clayton Foundation Investigator.
Abbreviations
- DDP
- cisplatin
- PC
- phytochelatin
- GSH
- glutathione
- PC/CdS
- phytochelatin/cadmium/sulfide
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}