


Abstract
α2-Adrenergic receptors (α2-ARs) regulate many physiological functions and are targets for clinically important antihypertensive and anesthetic agents. Three human and mouse genes encoding α2-AR subtypes (α2A, α2B, and α2C) have been cloned. We investigated the involvement of the α2C-AR in α2-adrenergic pharmacology by applying molecular genetic techniques to alter the expression of α2C-AR in mice. The effects of dexmedetomidine, a subtype-nonselective α2-AR agonist, on monoamine turnover in brain and on locomotor activity were similar in mice with targeted inactivation of the α2C-AR gene and in their controls, but the hypothermic effect of the α2-AR agonist was significantly attenuated by the receptor gene inactivation. Correspondingly, another strain of transgenic mice with 3-fold overexpression of α2C-AR in striatum and other brain regions expressing α2C-AR showed normal reductions in brain monoamine metabolism and locomotor activity after dexmedetomidine, but their hypothermic response to the α2-AR agonist was significantly accentuated. The hypothermic effect of α2-AR agonists thus seems to be mediated in part by α2C-AR. Some small but statistically significant differences between the strains were also noted in brain dopamine metabolism. Lack of α2C-AR expression was linked with reduced levels of homovanillic acid in brain, and mice with increased α2C-AR expression had elevated concentrations of the dopamine metabolite compared with their controls.
α2-ARs mediate many physiological functions and pharmacological effects in the central nervous system, mainly by inhibiting neuronal firing and release of NE and other neurotransmitters. α2-ARs are also involved in a wide range of functions in peripheral tissues (e.g., in the regulation of NE release from sympathetic nerves, smooth muscle contraction, platelet aggregation, insulin secretion, glomerular filtration, and energy metabolism) (1). Activation of α2-ARs with the highly specific α2-AR agonist dexmedetomidine results in bradycardia, hypotension, hypothermia, locomotor inhibition, anxiolysis, analgesia, sedation, and, with higher doses, anesthesia. Dexmedetomidine also reduces the turnover of the monoamine neurotransmitters NE, DA, and 5-HT (serotonin) in brain (2).
Recent pharmacological and biochemical research has led to a subdivision of α2-ARs into three distinct subtypes: α2A-, α2B-, and α2C-ARs. This classification was first based on the pharmacological properties of the receptors and was confirmed through the cloning of three distinct α2-AR genes in humans, rats, mice, and other species (3). Each receptor has a distinct tissue distribution. In the central nervous system of the rat, α2A-ARs are widely expressed, whereas the other α2-AR subtypes have more limited distributions. α2C-ARs are present in the basal ganglia, olfactory tubercle, hippocampus, and cerebral cortex, but α2B-ARs are expressed only in thalamic nuclei (4, 5). The α2A-AR seems to have an important role as a presynaptic and somatodendritic autoreceptor in noradrenergic nerve cells because this receptor subtype has been identified through both in situ hybridization (4, 5) and immunohistochemistry (6, 7) in the cells of the locus ceruleus and other noradrenergic centers.
Pharmacological experiments have not revealed the functions of the individual receptor subtypes because suitable subtype-selective agonists or antagonists have not been available. An alternative approach to this problem is to use transgenic techniques to genetically alter the expression of a single receptor gene and thereby gain knowledge about the functions of a particular receptor protein (8-13). Recent results obtained with transgenic mice with dysfunctional α2A-ARs and mice with targeted disruption of α2B- and α2C-AR genes demonstrate the involvement of α2A-AR in central cardiovascular regulation (14) and of α2B-AR in the peripheral vasoconstriction induced by α2-AR agonists (15). We studied the physiological and pharmacological functions of α2C-AR in mice with tissue-specific overexpression of this receptor and in mice with targeted disruption of the α2C-AR gene and lack of functional α2C-AR protein. We concentrated on locomotor functions and temperature control because the distribution of α2C-AR in rodent brain points to an involvement of these receptors in striatal functions. In addition, we studied whether the neurochemical effects of a potent, subtypenonselective α2-AR agonist on monoamine turnover in brain would be influenced by altered α2C-AR expression.
Materials and Methods
Animals
Two strains of genetically engineered mice were generated at Stanford University (Stanford, CA). One had a targeted inactivation of the α2C-AR gene (16), and the other had tissue-specific overexpression of α2C-ARs. A total of 60 mice were shipped to the Central Laboratory Animal Facility of the University of Turku, Finland, where they were maintained and bred according to standard procedures.
The generation of mice with targeted disruption of the gene encoding α2C-AR (KO mice) has been described previously (16). Briefly, the mutation was made by inactivating one copy of theAdra2c gene (the murine gene of the α2C-adrenergic receptor) in R1 129/Sv embryonic stem cells (17). Cells containing a targeted mutation of theAdra2c locus were injected into C57BL/6J blastocysts, and the resulting chimeric mice were bred to F1 (C57BL/6J × DBA/2J) animals. These animals were back-crossed for several generations to C57BL/6J mice. The resulting offspring from heterozygous intercrosses formed the colony maintained in Turku. The genetic background of the strain is thus mainly C57BL/6J with a small contribution from 129/Sv and DBA/2J strains.Adra2c −/− mice are viable and fertile and appear grossly normal. The germline transmission of the mutation was continuously monitored from mouse tail DNA by Southern blot analysis.
Mice with overexpression of the α2C-AR gene were generated by pronuclear injection using standard techniques (18, 19). In brief, one-cell fertilized eggs were harvested from 5-week-old superovulated FVB/N female mice. The microinjected constructs contained 4.5 kb of 5′ flanking sequence and 5 kb of 3′ flanking sequence surrounding the Adra2c open reading frame, which encodes the murine α2C-AR (20). In addition, we attached the influenza hemagglutinin epitope recognized by the antibody 12CA5 to the extreme amino terminus of the Adra2c protein. Previous research has demonstrated that the amino-terminal epitope does not impair expression of the Adra2c receptor or alter its ligand-binding properties to a significant degree (21). A tyrosinase minigene construct was coinjected for visual identification of the transgenic progeny on the basis of different coat color. This construct contained 2.2 kb of 5′, 0.7 kb of 3′, and 1.3 kb of coding sequence from pBS-Tyrosinase (22). Eggs that survived microinjection, as judged by morphology, were transferred to the oviduct of pseudopregnant foster mothers. DNA was collected from tail biopsies of weanling animals from these transfers and analyzed by Southern blot analysis or PCR for the presence of the adrenergic transgene (Fig. 1). For transgene-specific PCR, the sense primer (5′-AGC TTC CAT GGG CTA CCC ATA CGA CGT CCC AGA CTA CGC CAG-3′) was located in the 12CA5 epitope, whereas the antisense primer (5′-TTT CTC GCT GAG CGT ACG-3′) was located immediately after the fifth transmembrane domain of theAdra2c protein. Southern blot analysis was performed as described previously (16) using a 0.72-kbNcoI/MluI Adra2c probe. The adrenergic transgene and dark coat color were found to cosegregate during breeding. In addition, the intensity of dark coat color was correlated with the number of copies of both the tyrosinase minigene and the adrenergic transgene as detected by Southern blot analysis. The transgenic mouse line (281#17) that had the greatest amounts of gene copies was chosen for further breeding and experimentation (23).

PCR and Southern blot characterization of transgenic mouse lines carrying the Adra2coverexpression transgene. a, Ethidium-stained agarose gels showing detection of transgenic founder animals using the PCR assay described in Materials and Methods. Arrowhead, position of the 777-bp transgene-specific fragment. Left, sizes of relevant size standards. Standards were HindIII cut λ and HaeIII cut PhiX174 DNA. Open arrows, PCR-positive founder lines (numbered). The coat color positive line no. 6 (filled arrow), which carries only integrated copies of the tyrosinase minigene, was confirmed to be PCR negative in this assay. b, Quantitative Southern blot analysis comparing transgene copy number from different transgenic lines with the endogenousAdra2c gene from a wild-type (wt) mouse. Equal amounts of genomic DNA from tail biopsies were digested withEcoRI and detected as described in Materials and Methods. With this strategy, the Adra2c coding sequences derived from the wild-type locus and the transgene concatamers comigrate as a 6.3-kb fragment.
Mice were housed in groups of 7–10 at 22 ± 1°, with a 12:12-hr light/dark cycle (lights on at 6:00 a.m. and off at 6:00 p.m.). Standard certified pelleted food (Rat/Mouse 1 Maintenance Expanded SQC; Special Diet Services, Essex, UK) and water were available ad libitum. The mice were transferred to the neuropharmacological test laboratory for behavioral experiments at least 1 week before testing. All tested KO mice were 2.5–4.5 months old and homozygous for the mutation, and all OE mice were 1.5–2.5 months old and heterozygous. Wild-type control animals for each strain were age- and sex-matched littermates or closely related animals. Each animal was used only once.
Analysis of α2C mRNA Expression in Brain
In situ hybridization.
Four KO and OE mice and four respective control animals (age, 3–4 months) were used forin situ hybridization. The mice were killed with carbon dioxide, and the brains were rapidly excised and cooled in ice-cold 0.9% saline. The brains from KO and OE animals and their controls were frozen together on the same specimen holder. The tissues were sectioned in a Microm HM 500 cryostat at 14 μm and thaw-mounted onto Probe-On glass slides (Fisher Scientific, Pittsburgh, PA). The sections were stored at −20° until use.
Hybridization probes.
Two oligonucleotides [nucleotides 186–233 and 815–862 of the Adra2c coding sequence (20)] were labeled to specific activity of 1 × 109 cpm/μg at the 3′ end with [33P]dATP (New England Nuclear Research Products, Boston, MA) using terminal deoxynucleotidyltransferase (Amersham, Buckinghamshire, UK). Both probes produced similar results. Several control probes with the same length and similar GC content and specific activity were used to determine the specificity of the hybridizations. The addition of 100-fold excess of respective unlabeled probe abolished all hybridization signals.
In situ hybridization was carried out as described previously (24) with minor modifications. The slides were incubated in humidified boxes at 42° for 18 hr with a 5 ng/ml concentration of the labeled probe in a hybridization mixture containing 50% formamide (G.T. Baker, Deventer, Holland), 4× SSC (1× SSC consists of 0.15m NaCl and 0.015 m sodium citrate), 1× Denhardt’s (0.02% bovine serum albumin, 0.02% Ficoll, 0.02% polyvinylpyrrolidone), 1% sarkosyl (N-laurylsarcosine; Sigma Chemical, St. Louis, MO), 0.02 m sodium phosphate, pH 7.0, and 10% dextran sulfate (Pharmacia, Uppsala, Sweden). Sections were subsequently rinsed in 1× SSC at 55° for 60 min with four changes of SSC and finally in 1× SSC starting at 55° and slowly cooled to room temperature (∼1 hr), transferred through distilled water, briefly dehydrated in 60% and 95% ethanol for 30 sec each, air-dried, and covered with Hyperfilm-βmaxautoradiographic film (Amersham) for 30 days. Films were then developed with LX 24 developer (Kodak, Rochester, NY) for 2 min and fixed with Unifix (Agfa-Gevaert, Leverkusen, Germany) for 15 min.
Analysis of α2C-AR Binding Sites in Brain
Receptor autoradiography.
Three 7-week-old mice from each group were killed by decapitation, and their brains were rapidly dissected and frozen by immersion into cold isopentane in a dry ice bath. Coronal 14-μm sections were cut on a Microm cryostat and thaw-mounted onto gelatin-coated slides. The slides were first dried at room temperature for 2 hr and subsequently stored at −70° with desiccant in sealed containers.
Receptor autoradiographic methods were modified from earlier reports on the use of the subtype-nonselective α2-AR radioligand [3H]RX821002 (59 Ci/mmol; Amersham; eight radioligand concentrations: range, 0.1–16 nm) and the α2C-AR-preferring radioligand [3H]rauwolscine (83 Ci/mmol; DuPont-NEN, Bad Homburg, Germany; 0.1–13 nm) (25, 26). The assay conditions were optimized and validated in preliminary experiments. Incubations were carried out at room temperature in 14-ml plastic slide mailers in 50 mm potassium phosphate buffer, pH 7.4. For [3H]RX821002, the incubation time was 20 min, and washes were performed twice for 2 min in the same buffer on ice. For [3H]rauwolscine, the incubation time was 60 min, and washes were performed for 20 and 40 min in ice-cold buffer. Next, the slides were briefly dipped in cold water to remove salts and dried under a stream of cool air. Specificity of α2-adrenergic binding was determined in parallel incubations supplemented with 100 μm (−)-epinephrine (Sigma).
Radiolabeled, dried tissue sections were apposed to tritium-sensitive film (Hyperfilm [3H], Amersham) along with autoradiographic [3H] microscales (Amersham and American Radiolabeled Chemicals, St. Louis, MO) for 5 or 12 weeks. Films were developed with Kodak D-19, and the autoradiographic images were analyzed with a computerized image analysis system (MCID M4; Imaging Research, St. Catharine’s, Ontario, Canada). With shading correction, the images were captured with a CCD video camera (Hamamatsu C3077, Hamamatsu Photonics, Hamamatsu City, Japan) and digitized into an array of 640 × 480 pixels with a density range of 0–255. A standard curve was generated by measuring and plotting the absorbances of the images of the plastic microscales versus their radioactivity. Areas of interest were identified and traced, and their absorbance was measured and converted to radioactivity through interpolation. Values for nonspecific binding were subtracted from total binding. Nonspecific binding, determined with 100 μm (−)-epinephrine, always represented <10% of total binding. The results for specific binding are expressed as fmol of radioligand bound/mg of tissue protein. Nonlinear analysis of regression was used to derive equilibrium dissociation constants and receptor densities (KD and B max). The computer program package GraphPAD InPlot (GraphPAD Software, San Diego, CA) was used for this purpose. Two forebrain regions were analyzed that correspond to plates 24–29 and 41–43 of the anatomical mouse brain atlas of Sidman et al. (27). The following structures were identified and measured in both hemispheres: caudate-putamen, fundus striatum, frontal cortex, CA1 field of the hippocampus, CA3 field of the hippocampus, and dentate gyrus. The values from the two hemispheres were averaged for subsequent calculations. Histological control sections were stained with hematoxylin and eosin or cresyl violet according to standard procedures, or they were stained for acetylcholinesterase by theS-acetylthiocholine method (28).
Striatal homogenates.
The 7-week-old mice were killed by decapitation, and their striata were rapidly dissected and homogenized in 10 volumes of potassium phosphate buffer. Crude membrane fractions were prepared by centrifugation. The membranes were washed once and used for binding assays using the α2 subtype-nonselective radioligand [3H]RX821002 (0.1–4 nm) or the α2C-AR-preferring radioligand [3H]rauwolscine [0.8 nm with or without 100 nm oxymetazoline (Sigma) to block α2A-ARs] (29). (−)-Epinephrine (100 μm) was used to determine specificity of binding.
Spontaneous Motor Activity Tests
Spontaneous locomotor activity was measured by placing individual animals into a polypropylene animal cage (38 × 22 × 15 cm) with a transparent polypropylene lid and a ∼1.5-cm-thick layer of granulated aspen bedding on the floor. The cages were surrounded with a photobeam frame system designed for activity measurements (Photobeam Activity System PAS, Cage Rack, San Diego Instruments, San Diego, CA). The system consisted of 16 separate frames connected to a computer control unit. The system enabled the simultaneous measurement of the activity of eight mice at two levels (3 and 6 cm from the bottom of the cage). Three different parameters were recorded: 1) ambulations [large movements (breaks of two adjacent beams of light at the lower level)], 2) fine movements (two consecutive breaks of a single beam of light at the lower level), and 3) rearings (one break of a beam of light at the upper level).
The base-line locomotor activity of all mouse strains (eight 12–16-week-old male mice per group) was measured over 1-hr intervals for 22 hr after a 3-hr habituation period to detect possible differences in spontaneous locomotor activity or its diurnal rhythm.
The effects of the potent and specific α2-AR agonist dexmedetomidine (Orion Corporation, Orion-Pharma, Turku, Finland) (2) on locomotor activity were tested in two separate experiments with adult males. In the first experiment, three different doses (5, 10, or 20 μg/kg) or control solution (distilled water) was administered subcutaneously to OE and OE-wt mice (10 mice per group; total, 80). In the second experiment, groups of KO and KO-wt mice (11–14 mice per group; total, 116) were administered dexmedetomidine (5, 10, 20, or 30 μg/kg) or vehicle. The mice were weighed on the day of the experiment, and allocation of the mice to different treatment groups was randomized according to the Latin square principle. Dexmedetomidine or vehicle was administered 20 min before the animal was placed into the measurement cage, and activity was measured over 10-min intervals for 30 min. The mice were decapitated 1 hr after drug administration, and their brains were rapidly dissected and frozen (−70°) for neurochemical assays.
Body Temperature Experiments
Core body temperatures were measured with a rectal probe and a digital thermometer (Ellab, Roedovre, Denmark). The probe was inserted 2.5 cm inside the anal sphincter. The animals were familiarized to the procedure 1 day before the pharmacological experiments. Three doses (10, 20, and 30 μg/kg) of dexmedetomidine or vehicle (distilled water) was administered subcutaneously, and body temperature was measured at base-line and 30, 60, and 90 min after the injection. KO mice and controls (10 mice per group; total, 80) were female, and OE mice and controls (8–10 mice per group; total, 76) were male. Similar groups of male KO and KO-wt mice were also challenged with three different hypothermic doses of the nonselective DA agonist (−)-apomorphine (0.1, 0.3, and 1.0 mg/kg; Research Biochemicals, Natick, MA) and the 5-HT1A receptor agonist (±)-8-OH-DPAT (0.1, 0.3, and 1.0 mg/kg; Research Biochemicals).
Neurochemical Assays
Biogenic amines and their metabolites were determined from striatal and frontal cortical samples and from whole-brain homogenates in 0.1 m perchloric acid using electrochemical detection (ESA Coulochem, 5011, Bedford, MA) after separation by HPLC on a reversed-phase C18 column (Ultrasphere ODS, 4.6 × 250 mm; Beckman Instruments, Fullerton, CA). The buffer systems described by Mefford (30) were used, with separate assays for indoles and the DA metabolite HVA and for catecholamines after their purification on activated alumina. The minor modifications to the original published procedures have been described elsewhere (31). The NE metabolite MHPG was determined in a separate HPLC assay using a modified procedure previously validated for plasma samples (32). The brain homogenates were centrifuged, and the supernatants were neutralized before extraction with Bond Elut PH columns (Analytichem International, Harbor City, CA). The intra-assay coefficient of variation was 6.6% at 0.40 nmol/g, and the interassay coefficient of variation was 10.0% at 0.28 nmol/g.
Statistical Analysis
The statistical significance of differences between groups was determined by ANOVA using STATISTICA 4.5 computer software (StatSoft, Tulsa, OK). Two-way ANOVA was used for the locomotor activity results and neurochemical measurements to test the differences between genotypes and drug doses, and a comparison of different doses was performed with Tukey’s post hoc t test. The body temperature measurements were evaluated using three-way ANOVA for repeated measurements using genotype, dose, and time as independent factors. Values of p < 0.05 were considered statistically significant. The results are presented as mean ± standard error.
Results
Analysis of α2C mRNA expression in brain by in situ hybridization.
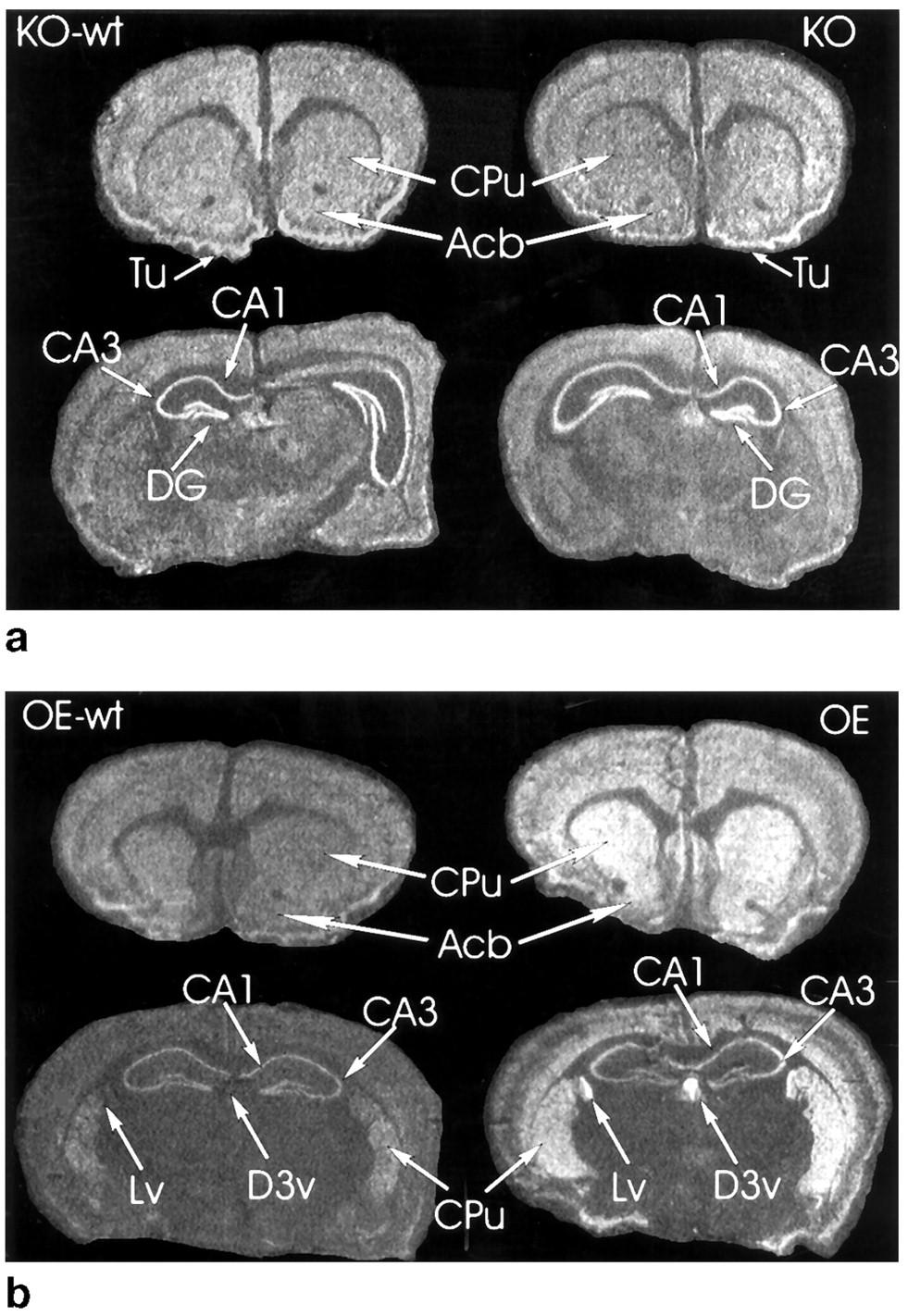
No clear differences were observed in the intensity or distribution of the hybridization signals between KO and KO-wt animals (Fig. 2, top). Brain sections from OE mice had clearly increased α2CmRNA in regions that also in OE-wt mice were labeled for α2C mRNA (Fig. 2, bottom). In addition, very strong hybridization was seen in the ependyma of the ventricles and choroid plexus of OE mice (Fig. 2, bottom).

Top, in situhybridization of α2C mRNA in the brains of KO-wt and KO mice. No clear difference in the expression of α2C mRNA can be seen. Bottom, in situhybridization of α2C mRNA in the brains of OE-wt and OE mice. The hybridization signal is increased in OE mice. The signal in OE animals is in the same brain areas that in OE-wt mice exhibit α2C mRNA except that a clear signal is seen in the ependyma of the ventricles and choroid plexus. Acb, accumbens nucleus; CA1 and CA3, fields of Ammon’s horn; DG, dentate gyrus; CPu, caudate-putamen; Tu, olfactory tubercle;Lv, lateral ventricle; D3v, dorsal third ventricle.
Analysis of α2C-binding sites in brain by receptor autoradiography.
Fig. 3 shows the autoradiographic distribution of [3H]RX821002 and [3H]rauwolscine binding in forebrain sections of OE mice and their controls (OE-wt). Both [3H]RX821002 and [3H]rauwolscine clearly labeled a larger population of sites in many regions of OE mouse brain compared with corresponding regions in control (OE-wt) mice (Table 1). This difference was largest in the caudate-putamen and in the stratum radiatum of the CA1 region of the hippocampus (∼3-fold). In addition, clearly increased α2C-AR binding was noted overlying all brain ventricles. There was no difference inB max values between the strains in frontal cortex and in the dentate gyrus. Receptor affinity for [3H]RX821002 was equal in both strains and in all regions, ∼0.4 nm. In contrast, calculatedKD values for [3H]rauwolscine were clearly smaller in OE mice than in OE-wt mice, especially in brain regions in which α2C-ARs were abundant (fundus striatum, caudate-putamen, stratum radiatum of CA1: 0.3 versus 1.3 nm). Rosenthal plots of [3H]rauwolscine binding could not be reliably resolved into two populations of binding sites because of the small number of radioligand concentrations used in the assays. The reduction in total α2-AR density and high affinity [3H]rauwolscine binding in brains of KO mice has been demonstrated previously (16).

Images of autoradiographic binding of (top) [3H]RX821002 and (bottom) [3H]rauwolscine in coronal sections from two levels of forebrain of (left) OE and (right) OE-wt mice. Acb, accumbens nucleus; CA1 and CA3, fields of Ammon’s horn; LMol, lacunosum moleculare; Rad, stratum radiatum; DG, dentate gyrus; CPu, caudate-putamen; Lv, lateral ventricle;D3v, dorsal third ventricle; Tu, olfactory tubercle; LSD, lateral septal nucleus.
Density (B max) of α2C-AR binding sites in different regions of brains of OE and OE-wt mice determined byin vitro autoradiography with [3H]RX821002 and [3H]rauwolscine
Radioligand binding to striatal membranes.
Striatal membrane preparations from four animals from each group were used to determine receptor affinities (KD ) and receptor densities (B max) in this brain region. Receptor affinities for [3H]RX821002 were similar in membranes from OE, OE-wt, KO, and KO-wt mice: 0.23–0.37 nm. The binding capacity was clearly lower in membranes from KO mice (81 ± 8 fmol/mg of protein) than in membranes from KO-wt mice (122 ± 20 fmol/mg of protein); an intermediate value was obtained with membranes from mice heterozygous for the inactivating mutation (94 ± 7 fmol/mg). Compared with [3H]RX821002, binding of [3H]rauwolscine to striatal membranes was more markedly reduced in KO mice, especially in experiments conducted in the presence of 100 nm oxymetazoline to block binding to α2A-AR (10 ± 4 versus 35 ± 3 fmol/mg; 20 ± 5 fmol/mg in heterozygous mice). The results for OE mice and their controls are shown in Table 2 separately for male and female animals. The capacity of [3H]RX821002 binding to striatal membranes of OE mice was more than double that of membranes from OE-wt mice (162 versus 67 fmol/mg). Binding of [3H]rauwolscine to striatal membranes of OE mice indicated a ∼3-fold higher density of α2C-adrenergic binding sites in striatal membranes from OE mice than in membranes from OE-wt mice (109 versus 36 fmol/mg).
α2-AR density in membrane preparations from striata of 6–7-week-old OE and OE-wt mice of either sex
Spontaneous motor activity and the effects of dexmedetomidine.
OE mice and their controls were more active in the 22-hr base-line activity measurements compared with KO mice and their controls. The average total counts (ambulations plus fine movements) over 22 hr were 4750 ± 136/4910 ± 116 for the OE and OE-wt mice and 3680 ± 92/3440 ± 73 for KO and KO-wt mice. No differences were noted between the mice with altered α2C-AR expression and their respective controls in total counts, ambulations or fine movements, or the diurnal pattern of locomotor activity (Fig.4).

Base-line locomotor activity of (a) OE and OE-wt mice and (b) KO and KO-wt mice. The total activity (ambulations and fine movements) was measured at 1-hr intervals for 22 hr starting at 2:00 p.m. after a 3-hr habituation. Filled bars, OE mice. Open bars, KO mice. Hatched or cross-hatched bars, respective wt controls. Error bars, standard error for eight measurements.
Dexmedetomidine decreased the activity of all strains of mice (Fig.5). Compared with vehicle-treated animals, the reduction in ambulations, fine movements, and rearings achieved statistical significance at doses of 10 and 20 μg/kg in OE and OE-wt mice (p < 0.0002), whereas the 5 μg/kg dose significantly reduced the activity of KO and KO-wt mice (p < 0.0002 for all locomotor variables). Significant strain differences were not noted between OE and OE-wt mice or between KO and KO-wt mice when all time periods were included in the analysis (20–50 min after drug administration). In addition, no significant strain × dose interactions were observed, indicating that the effect of dexmedetomidine on locomotor activity was not affected by the altered α2C-AR expression.
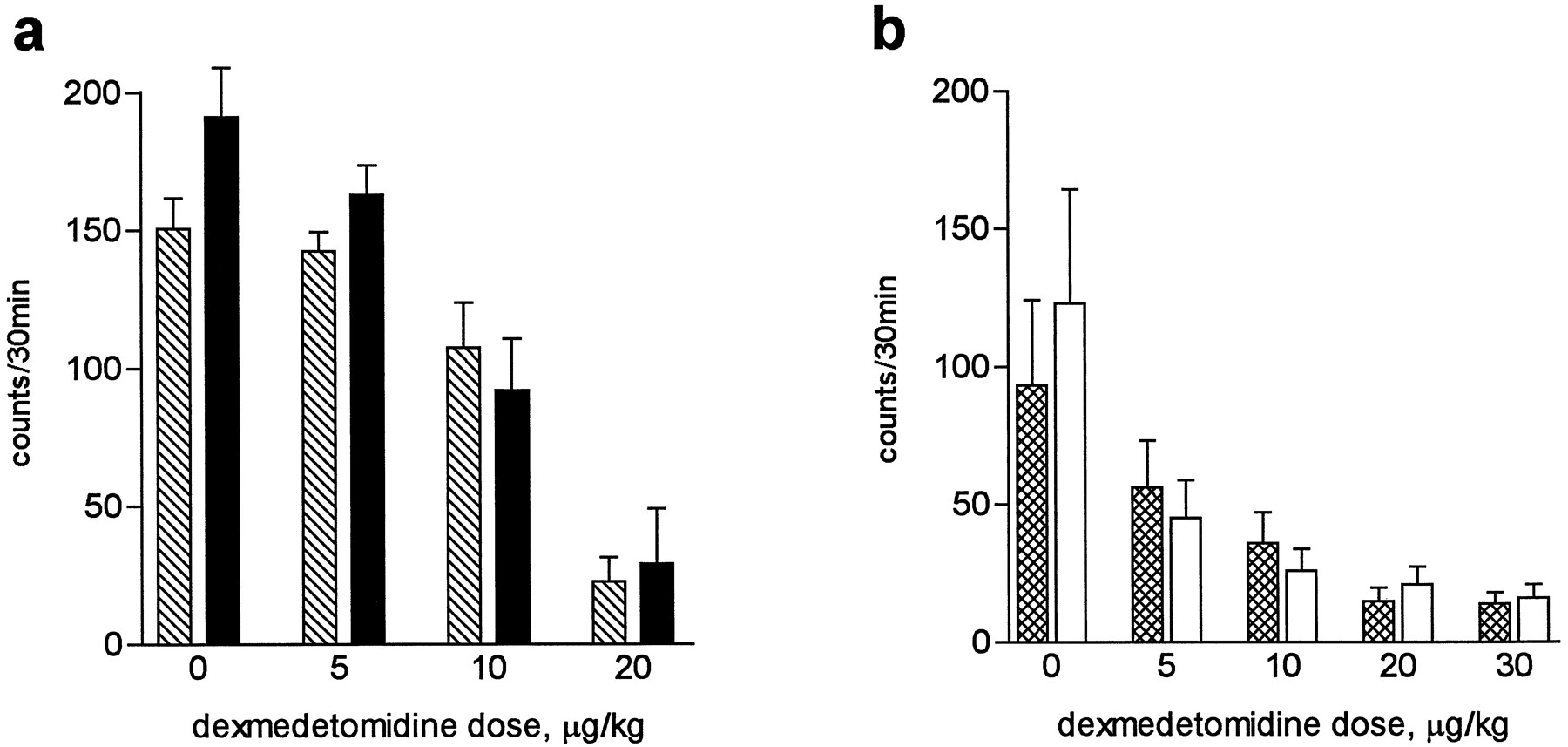
Locomotor activity measured as ambulation counts (large horizontal movements) in two separate drug experiments. a, Effect of dexmedetomidine on OE and OE-wt mice. b, Effect of dexmedetomidine on KO and KO-wt mice. Filled bars, OE mice. Open bars, KO mice. Hatched or cross-hatched bars, respective wt controls. Error bars, standard error for 10–14 measurements.
Effects of dexmedetomidine on body temperature.
The dose-dependent hypothermic effect of dexmedetomidine was more potent in OE animals than in their controls (three-way ANOVA, time × strain interaction: p = 0.0017). Correspondingly, the KO mice were not as sensitive to the hypothermic effect of the drug as their controls (p = 0.0029) (Fig. 6). The average differences from the controls in the reduction of body temperature calculated at 30 min after the injection were 0.80° (17.3%) in KO mice and 0.43° (11.8%) in OE mice when all doses of active drug were included in the analysis. There were no differences in the hypothermic responses of KO and KO-wt mice to the DA agonist apomorphine or the 5-HT1A agonist (±)-8-OH-DPAT. Apomorphine (1 mg/kg) lowered the mean body temperature 30 min after the injection by 4.86 ± 0.32° and 5.57 ± 0.49° in KO and KO-wt mice, respectively (p = 0.10). After 0.3 mg/kg, the reductions were 3.41 ± 0.65° and 3.28 ± 0.53° (p = 0.99). The corresponding temperature reductions induced by 1 mg/kg (±)-8-OH-DPAT were 3.17 ± 0.30° and 2.90 ± 0.36° (p = 0.90). In addition, the two strains of control mice showed differing sensitivity to dexmedetomidine (p < 0.0001), with the OE-wt mice less hypothermic after dexmedetomidine than the KO-wt mice. This difference may have been due to sex or strain differences or both.
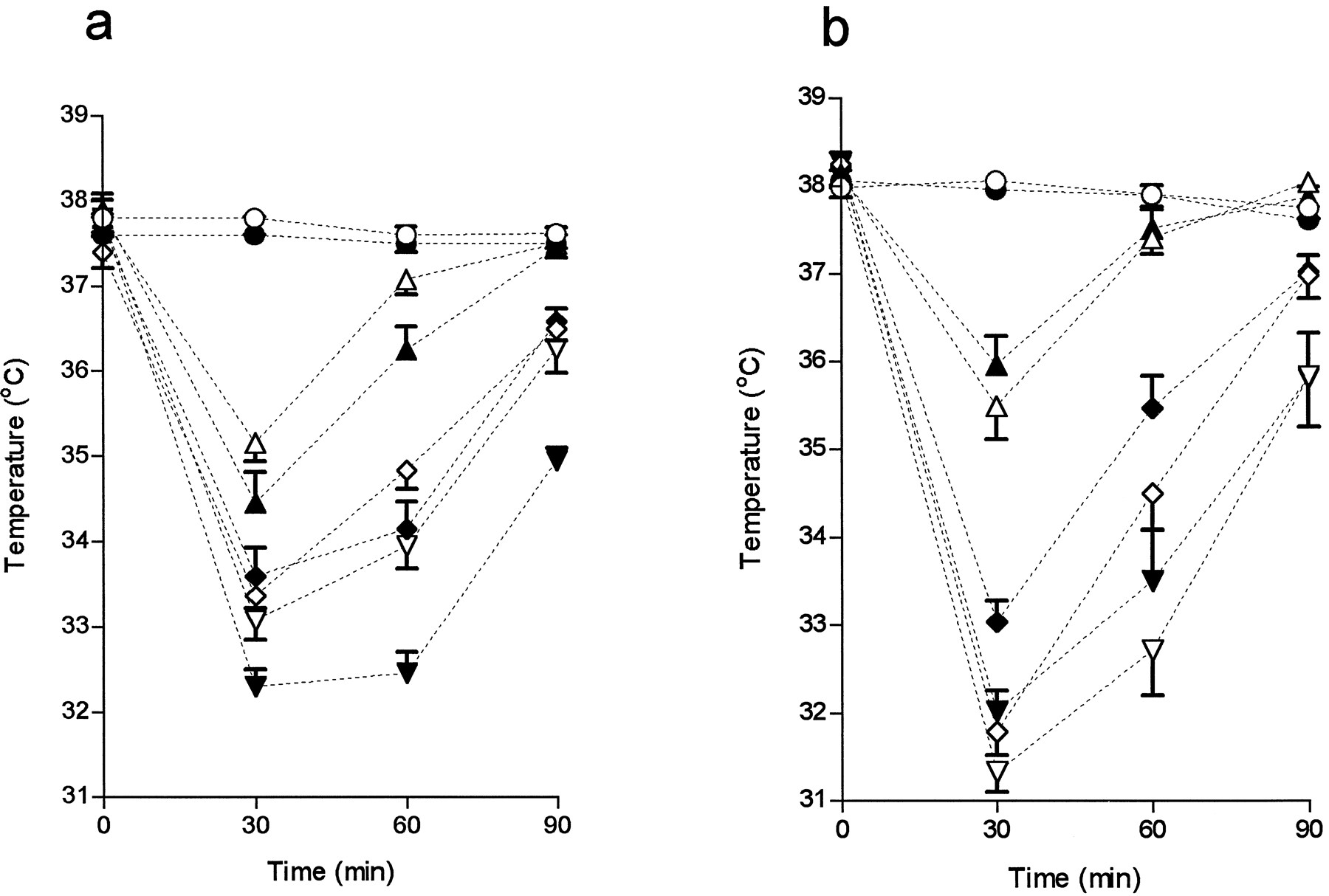
Time course of effects of different doses (0, 10, 20, and 30 μg/kg) of dexmedetomidine on body temperature. a, OE and OE-wt mice (male; 8–10 mice per group). b, KO and KO-wt mice (female; 10 mice per group). Values are mean ± standard error. ○, ▵, ⋄, and ▿, wt mice. •, ▴, ♦, and ▾, (a) OE or (b) KO mice. ○, 0 μg/kg; ▵, 10 μg/kg; ⋄, 20 μg/kg, ▿, 30 μg/kg.
Effects of dexmedetomidine on brain monoamines and their metabolites.
Table 3 shows the results of brain neurochemistry analyses from whole-brain homogenates. As expected, dexmedetomidine dose-dependently reduced the concentrations of the measured monoamine metabolites in whole brain. There were, however, no significant strain × dose interactions in the two-way ANOVA, indicating that the drug-induced reductions in the metabolism of NE, DA, and 5-HT were not influenced by altered α2C-AR expression. When all treatment groups were included in the ANOVA, statistically significant strain differences were observed as follows: OE mice had higher concentrations of HVA (p < 0.0001) and DA (p = 0.049) in brain than their controls, and KO mice had lower concentrations of HVA (p = 0.013), 5-HIAA (p= 0.041), and MHPG (p = 0.007) in brain than their controls. In brain samples from another group of naive mice, KO mice, compared with KO-wt mice, had lower concentrations of HVA in striatum (−15.1%) (p = 0.021) but not in frontal cortex, and OE mice, compared with OE-wt mice, had higher concentrations of HVA (+19.2%) (p = 0.038) in frontal cortex but not in striatum (Table 4). Dexmedetomidine had no significant effects on the levels of NE, DA, and 5-HT in brains of OE and OE-wt mice, but it increased the amine concentrations in brains of KO and KO-wt mice. It also significantly reduced the brain concentrations of the 5-HT precursor amino acid tryptophan in all strains.
Effects of dexmedetomidine on monoamine metabolism in whole-brain homogenates of OE and OE-wt mice and KO and KO-wt mice
Concentration of HVA in striatal and frontal cortical brain samples of OE and KO mice and their controls
Discussion
The present in situ hybridization, receptor autoradiographic, and in vitro receptor binding results from the brains of OE mice showed that microinjections of the described DNA sequences could be successfully used to generate a strain of mice with tissue-specific overexpression of α2C-ARs. The in situ results and receptor autoradiograms from brains of OE mice showed the expected regional distribution of increased α2C-AR expression, confirming that the expression was tissue specific. The promoter elements present in the gene construct were capable of directing the increased receptor expression to those brain regions, and probably also to those cells, which normally express endogenous α2C-ARs. The only possible exception to this contention detected so far was the presence of α2C mRNA and [3H]rauwolscine binding in ependymal tissue in OE mice. Control (OE-wt) mice did not have detectable α2CmRNA hybridization or [3H]rauwolscine binding in this tissue. Results from receptor binding assays with striatal homogenates support the conclusion that OE mice have ∼3-fold tissue-specific overexpression of α2C-AR in striatum.
As reported previously (16), based on evidence from Northern blot and reverse-transcription PCR analyses, Adra2c −/−(KO) mice do not produce gene transcripts capable of encoding functional α2C-AR protein. This was supported by receptor autoradiographic results from brain sections, in which [3H]rauwolscine binding (in the presence of oxymetazoline to mask α2A-AR) was markedly reduced in the caudate-putamen, the brain region normally expressing the highest proportion of α2C-AR (16). The brain regions that were found to have significantly increased binding of [3H]rauwolscine in OE mice were the same with reduced binding in KO mice (16).1 Also, this distribution pattern agrees with our α2C mRNA in situ hybridization results from normal mice and previous, more-detailed in situ α2C-AR mRNA hybridization results in the rat brain (4, 5). In situhybridization analysis of brain sections from KO mice and their controls yielded similar results, which is not unexpected because the probes were targeted to mRNA sequences upstream of the inactivating mutation.
Despite significantly altered α2C-AR expression, both KO and OE animals are viable and fertile and appear grossly normal. Clearly, the α2C-AR is not fundamental for mouse embryonic development or adult reproductive function, and a moderately increased receptor expression level does not confer a serious disadvantage for survival and reproduction. Also, altered α2C-AR expression did not have major effects on spontaneous locomotor activity or its diurnal rhythm. The altered α2C-AR expression modified the hypothermic response to the potent α2-AR subtype-nonselective agonist dexmedetomidine because OE mice were more hypothermic and KO mice were less hypothermic after the drug than were their controls. However, altered α2C-AR expression had no effect on the sedative response to dexmedetomidine.
According to the HPLC analyses, there were no overall strain-dependent differences in the sensitivities of mice to the α2-AR agonist-induced alterations in brain monoamine levels or metabolism. However, some minor differences were found. Interestingly, the concentrations of the DA metabolite HVA seemed to be influenced by altered α2C-AR expression, with OE mice having increased and KO mice having decreased concentrations of HVA. The similar drug effects on brain monoamine metabolism in the mutant and control mice indicate that the majority of the effects of α2-AR agonists on brain monoamine turnover are mediated by α2A-ARs, as was expected for NE.
In the neurochemical assays performed in the course of this study, the expected reductions in the turnover rates of brain monoamine neurotransmitters after dexmedetomidine were similar within both pairs of strains. However, some minor but consistent differences in the measured levels were noted in all dose groups between mutant and control mice. In KO mice, MHPG (metabolite of NE), 5-HIAA (metabolite of 5-HT), and HVA (metabolite of DA) levels were lower compared with their controls, whereas OE mice had minor elevations in their levels of HVA and DA in brain. Thus, it seems that the lack of α2C-AR expression slightly decreases the rate of monoamine turnover in brain and that overexpression of α2C-AR increases DA stores and metabolism. The opposite findings for the HVA levels in KO and OE mice suggest a possible involvement of α2C-ARs in regulation of brain DA systems. This was supported by the assays of striatal and frontal cortical samples of drug-naive animals, in which KO mice had lower HVA concentrations in striatum and OE mice had higher HVA concentrations in frontal cortex, whereas other monoamine and metabolite levels were unaltered. The difference in frontal cortex was somewhat unexpected because cortical α2C-AR expression is quite low, but it suggests that the involvement of the α2C-ARs in dopaminergic regulation is not confined to striatum.
The α2C-AR distribution in brain and the observed differences in HVA concentrations in both mutant mouse strains compared with their controls point to the possibility that α2C-ARs participate in regulation of dopaminergic function. In addition to locomotor functions, striatal dopaminergic systems have been implicated in other physiological functions, including modulation of body temperature regulation. This is a possible mechanism for the opposite alterations in the hypothermic responses to dexmedetomidine in KO and OE mice. The preoptic area of the anterior hypothalamus is a central site of thermoregulation in rodents, but pharmacological experiments have demonstrated that nigrostriatal dopaminergic neurons also participate in the complex regulation of body temperature (33). It is also possible that the modulatory effects of α2C-ARs on body temperature are due to altered heat production by peripheral muscles, which is indirectly controlled by nigrostriatal pathways. Similar thermal responses of KO and KO-wt mice to apomorphine and (±)-8-OH-DPAT support the assumption that central DA or 5-HT receptors are not compensatorily regulated in KO mice and that the strain-dependent changes in body temperature after dexmedetomidine are consequences of differing α2C-AR activation. KO mice, however, had a tendency to have an attenuated response to the highest dose of apomorphine compared with KO-wt mice, which is in agreement with the suggested α2 agonistic properties of apomorphine.
Our results of unaltered dexmedetomidine-induced locomotor inhibition in both KO and OE mice indicate that α2A-ARs are involved in mediation of the sedative effect of dexmedetomidine. This assumption is now directly testable in transgenic mice with dysfunctional α2A-ARs (14). The inhibition of spontaneous locomotor activity in the current study reflects mainly sedation, and it is not very sensitive to other possible drug-induced changes in motor performance. Thus, α2C-ARs may have more subtle modulatory effects on locomotor functions, as suggested by their presence in the striatum. Marked strain differences were observed between KO-wt and OE-wt mice in all behavioral experiments, complicating the study designs, because both mutant strains needed their own control strains and direct comparisons between KO and OE mice were not appropriate. It has recently been emphasized that phenotypical abnormalities observed in mice with targeted gene disruptions may in fact be due to chance effects attributable to background genes that are unevenly distributed in the breeding process (34). However, the opposite findings in our two gene-manipulated mouse strains, one with targeted disruption of the Adra2c gene and the other with tissue-specific overexpression of the receptor encoded by this gene, point to a specific role of the α2C-AR in mediating modulation of DA metabolism and the hypothermic effect of α2-AR agonists.
In conclusion, genetic alteration of receptor expression is a powerful new method of studying the physiological and pharmacological functions of a particular receptor, especially if selective agonists or antagonists are not available. This study supports the concept that the sedative effects of dexmedetomidine are probably mediated by α2A-ARs, which inhibit the activity of noradrenergic neurons in the locus ceruleus. In addition, because α2Aexpression has also been detected in dopaminergic and serotonergic control centers in rodent brain (4, 5) and α2B expression is very limited in brain, it seems that dopaminergic and serotonergic systems also are mainly regulated by α2A-ARs. Striatal α2C-ARs may, however, have subtle modulatory effects on the dopaminergic systems in brain. This could be studied further in α2C-AR-lacking KO mice and in OE mice (e.g., by exploring interactions in locomotor tests between adrenergic and dopaminergic drugs). Also, methods other than simple motor activity measurements that are specifically designed to monitor more complex aspects of motor performance might reveal a possible role of α2C-AR in locomotor modulation and coordination. The availability of mice with altered α2C-AR expression permits the analysis of the involvement of α2C-AR in many other α2-AR-mediated functions, such as blood pressure control (15), metabolic regulation, sensory modulation, and higher behavioral functions such as learning and memory.
Footnotes
- Received June 24, 1996.
- Accepted October 2, 1996.
-
Send reprint requests to: Dr. Mika Scheinin, MediCity Research Laboratory, University of Turku, Tykistökatu 6A, FIN-20520 Turku, Finland. E-mail: mschein{at}utu.fi
-
↵1 M. Kulatunga and M. Scheinin, unpublished observations.
Abbreviations
- AR
- adrenergic receptor
- DA
- dopamine
- NE
- norepinephrine
- MHPG
- 3-methoxy-4-hydroxyphenylglycol
- HVA
- homovanillic acid
- 5-HT
- 5-hydroxytryptamine
- 5-HIAA
- 5-hydroxyindoleacetic acid
- OE
- mice overexpressing the α2C-adrenergic receptor
- KO
- mice lacking the α2C-adrenergic receptor
- OE-wt
- control mice for mice overexpressing the α2C-adrenergic receptor
- KO-wt
- control mice for mice lacking the α2C-adrenergic receptor
- (±)-8-OH-DPAT
- (±)-8-hydroxy-2-dipropylaminotetralin
- PCR
- polymerase chain reaction
- HPLC
- high performance liquid chromatography
- ANOVA
- analysis of variance
- SSC
- standard saline citrate
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}