


Abstract
Three agents, verapamil, cepharanthine, and 2-[4-(diphenylmethyl)-1-piperazinyl]ethyl-5-(trans-4,6-dimethyl-1,3,2-dioxaphosphorinan-2-yl)-2,6-dimethyl-4-(3-nitrophenyl)-3-pyridinecarboxylate P-oxide (PAK-104P), that reverse drug resistance in P-glycoprotein (P-Gp)-mediated multidrug-resistant cells were examined for their activity to reverse drug resistance in multidrug resistance-associated protein (MRP)-mediated multidrug-resistant C-A120 cells. Agents other than PAK-104P could not reverse the resistance to doxorubicin in C-A120 cells. PAK-104P moderately reversed the doxorubicin resistance. In contrast, PAK-104P almost completely reversed the resistance to vincristine (VCR) in C-A120 cells as well as in KB-8–5 cells, and other agents moderately reversed the VCR resistance in C-A120 cells. PAK-104P at 10 μm enhanced the accumulation of VCR in C-A120 cells to the level of that in KB-3–1 cells without the agent. PAK-104P competitively inhibited the ATP-dependent [3H]leukotriene C4 uptake in membrane vesicles isolated from C-A120 cells. These findings demonstrate that PAK-104P can completely reverse the resistance to VCR in both P-Gp- and MRP-mediated multidrug-resistant cells and that PAK-104P directly interacts with MRP and inhibits the transporting activity of MRP.
The mechanism of MDR has been vigorously studied in tissue culture cells and human cancers. Overexpression of an Mr 170,000 transmembrane glycoprotein called P-Gp has been observed in various MDR cell lines (1). P-Gp is believed to function as an energy-dependent efflux pump (2). P-Gp-mediated MDR is reversed by a variety of compounds, including calcium channel blockers and their analogs that have considerably low calcium channel-blocking activity (3). Most agents that reversed MDR interacted with P-Gp and blocked the drug efflux from the cells (3-5).
Two other types of MDR, an “atypical” MDR caused by alterations in the level or drug sensitivity of topoisomerase IIα (6) and non-P-Gp-mediated MDR (7), have been reported. Recently, increased expression of a novel MDR gene, MRP, was found in many non-P-Gp-mediated MDR cells (8-13). MRP is a 1531-amino acidMr 190,000 membrane glycoprotein of the ATP-binding cassette superfamily of membrane proteins (8). MRP seems to confer MDR by transporting anticancer agents outside the cells (14). LTC4 was transported in the presence of ATP in membrane vesicles from cells expressing MRP (15-17), suggesting that MRP is the GS-X pump.
Little is known about the clinical significance of MRP-mediated MDR. However, non-P-Gp-mediated tumors that express MRP and are nonresponsive to chemotherapy have been reported (18). Expression ofMRP gene was increased in relapsed acute leukemia and the leading edge of the human lung tumors (19, 20).
Few agents reverse the MRP-mediated MDR. Cole et al. reported that ADM resistance of H69AR cells could not be reversed by most of the MDR-reversing agents (21). Buthionine sulfoximine, an inhibitor of glutathione synthesis, enhanced the toxicity of anticancer agents in the MRP-expressing MDR cells by inhibition of the enhanced efflux (22). The LTD4 receptor antagonist MK571 completely reversed VCR resistance in MRP-expressing MDR cells (23). MK571 efficiently inhibited the photolabeling of MRP using [3H]LTC4 and the transport of [3H]LTC4 into membrane vesicles from MRP-overexpressing MDR cells. These findings also point to a relationship between MRP and the GS-X pump.
In this study, we examined whether three agents, PAK-104P {2-[4-(diphenylmethyl)-1-piperazinyl]ethyl-5-(trans-4,6-dimethyl-1,3,2-dioxaphosphorinan-2-yl)-2,6-dimethyl-4-(3-nitrophenyl)-3-pyridinecarboxylate P-oxide}, verapamil, and cepharanthine, that reverse P-Gp-mediated MDR (24-26) also reverse the resistance to VCR as well as to ADM in MRP-mediated MDR cells.
Materials and Methods
Cell culture and cells.
Human epidermoid KB carcinoma cells were obtained from Dr. Michael M. Gottesman (National Cancer Institute, Bethesda, MD). KB cells were subcloned twice; a single recloned line, KB-3–1, was used as the parental cell line for the current study (27). KB-3–1 cells were cultured in minimal essential medium (Nissui Seiyaku, Tokyo, Japan) containing 10% bovine calf serum/iron supplement (Cell Culture Laboratories, Cleveland, OH). The P-Gp-mediated MDR mutant KB-8–5 was originally selected from KB-3–1 cells with increasing concentrations of colchicine and maintained in medium containing 10 ng/ml colchicine (27). MDR C-A120 cells, which overexpressed MRP mRNA, were isolated from KB-3–1 cells and maintained in the medium with 1 μg/ml cepharanthine, 100 nmmezerein, and 120 ng/ml ADM, as described previously (9).
Cell survival by MTT assay.
Chemosensitivity in vitro was measured by means of the MTT colorimetric assay performed in 96-well plates (28). The assay is dependent on the reduction of MTT by the mitochondrial dehydrogenase of viable cells to a blue formazan product that can be measured spectrophotometrically. Equal numbers of cells (2000 for KB-3–1; 5000 for KB-8–5 and C-A120) were inoculated into each well with 180 μl of culture medium. After an overnight incubation (37° with 5% CO2), 20 μl of VCR or ADM solution with or without reversing agent was added to the cultures and incubated for 4 days. Thereafter, 50 μl of MTT (1 mg/ml PBS) was added to each well and incubated for an additional 4 hr. The resulting formazan was dissolved with 100 μl of dimethylsulfoxide after aspiration of the culture medium. Plates were placed on a plate shaker for 5 min and read immediately at 570 nm using a Micro Plate Reader MPR-A4i (TOSOH, Tokyo, Japan).
Northern and slot blot analyses.
The genomic probe pMDR1, which was contained within an 800-base-pair PvuII fragment of pBR322, was obtained from Dr. Michael M. Gottesman. The humanMRP probe, 1-kb EcoRI fragment of MRP cDNA, was kindly provided by Dr. Susan Cole and Dr. Roger Deeley (Queen’s University, Kingston, Ontario, Canada). We used nylon membranes for all analyses, and they were moistened in 10 × SSC (1× SSC = 0.15 m NaCl plus 15 mm sodium citrate, pH 7.0) before use. In slot blot analysis, total RNA was applied to the slot blotter (Hybri-Slot TM manifold; Bethesda Research Laboratories, Gaithersburg, MD) under vacuum. For Northern analysis, poly(A)+ RNA was electrophoresed on a formaldehyde-agarose denaturing gel and blotted onto nylon membranes. The membranes were dried at room temperature before being cross-linked by UV irradiation in a Stratalinker (Stratagene, La Jolla, CA). The membranes were prehybridized for 4 hr at 42° in a solution (/ml) of 50% formamide, 5× Denhardt’s solution (1× Denhardt’s solution = 0.02% Ficoll, 0.02% polyvinylpyrrolidone, and 0.02% acetylated bovine serum albumin), 5× SSC, 0.05 m sodium phosphate buffer, pH 6.5, and 200 μg of denatured salmon sperm DNA. The membranes were then hybridized for 18 hr at 42° in a solution (/ml) containing 50% formamide, 1× Denhardt’s solution, 5× SSC, 0.02 m sodium phosphate buffer, pH 6.5, 10% dextran sulfate, and 200 μg of denatured salmon sperm DNA, and MRP or MDR1 probe labeled to a specific activity of >5 × 109 dpm/μg of DNA with [α-32P]dCTP (3000 Ci/mmol; ICN Biomedicals, Irvine, CA) using the Multiprime DNA Labeling System (Amersham International, Buckinghamshire, UK). After hybridization, the membranes were washed four times with 250 ml of 1× SSC/0.1% SDS for 15 min at room temperature and then twice with 500 ml of 0.2 × SSC/0.1% SDS for 10 min at 50°. The exposure period for MRP mRNA autoradiographs was ∼1–3 days. RNA loading was compared by hybridization with a32P-labeled β-actin probe.
Immunoblotting.
A monoclonal antibody against Chinese hamster P-Gp (C219), which was originally isolated by Kartner et al. (29), was obtained from Centocor (Malvern, PA). Polyclonal antibody against topoisomerase IIα were obtained from Dr. M. Kuwano (Kyushu University, Fukuoka, Japan) (30). Membrane vesicles (50 or 100 μg), nuclear extractions (7.5 μg), or cytosols (100 μg) were mixed with an equal volume of SDS sample buffer consisting of 125 mm Tris·HCl, pH 6.8, 4% SDS, 20% glycerol, and 0.005% bromophenol blue. Electrophoresis on SDS-7.5% (w/v) polyacrylamide minigels was performed according to the method of Laemmli (31) but without heating the samples. Transfer to PVDF membranes (Immobilon-P, Millipore, Bedford, MA), was performed electrophoretically for 30 min at 15 V (constant voltage) (32) using a Transblot SD apparatus (BioRad, Richmond, CA) as described by Kyhse-Anderson (33), except that the buffer consisted of 48 mm Tris and 39 mmglycine containing 20% (v/v) methanol, pH 9.2 (34). After transfer, the membranes were blocked with 3% skim milk in buffer A (0.35m NaCl, 10 mm Tris·HCl, pH 8.0, 0.05% Tween 20) for 1 hr at room temperature, followed by a 4-hr incubation with C219 at a concentration of 0.25 μg/ml or with 1000-fold diluted polyclonal antibody against topoisomerase II-α in buffer A containing 3% skim milk. The membrane was washed·three times with buffer A and then incubated for 1 hr with 1000-fold diluted anti-mouse Ig antibody labeled with HRP (Amersham) or HRP-conjugated anti-rabbit immunoglobulin. PVDF membranes were rinsed once for 15 min and four times for 5 min with buffer A and then evenly coated using the ECL Western blotting detection system (Amersham, Arlington Heights, IL) for 1 min. The membrane was immediately exposed to Kodak X-OMAT AR film at room temperature for various periods in a film cassette.
For detection of MRP, blots were incubated with an affinity-purified, rabbit polyclonal antibody raised against a synthetic peptide, the sequence (KEDTSEQVVPVLVKN) of which was predicted from that of the cloned MRP cDNA and is not found in any other known protein (35). Antibody binding was visualized with HRP-conjugated goat anti-rabbit IgG and the ECL Western blotting detection system (35)
Drug accumulation and efflux.
To measure drug accumulation, confluent monolayers of KB-3–1 and C-A120 cells in 24-well plates were incubated with 1 μm [3H]VCR in Hanks’ balanced salt solution with or without PAK-104P for the indicated times at 37° after an incubation with Hanks’ balanced salt solution for 15 min at 37°. After being washed with ice-cold PBS three times, the cells were solubilized in 1% Triton X-100 and 0.2% SDS in 10 mm phosphate buffer, pH 7.4, and harvested, and the radioactivity was determined. In studies of drug efflux, cells were incubated overnight and then further incubated in the presence or absence of 10 μm PAK-104P for 15 min. Medium was changed to fresh medium with or without PAK-104P, and cells were incubated with 1 μm[3H]VCR for 1 hr at 37°. Each dish was washed once with PBS, and fresh medium without [3H]VCR in the absence or presence of 10 μmPAK-104P was added and incubated for the indicated times at 37° and harvested; then, the radioactivity was determined.
Membrane vesicle preparation.
Membrane vesicles were prepared as described previously (36) from cells grown in 24.5 × 24.5-cm tissue culture plates (Nunc, Roskilde, Denmark). Protein concentrations were determined according to the method of Bradford (37).
[3H]LTC4 uptake in membrane vesicles.
LTC4 uptake in the vesicles was measured by filtration essentially as described by Ishikawaet al. (38, 39). The standard incubation medium contained membrane vesicles (50 μg of protein), 1.34 nm[3H]LTC4, 0.25 m sucrose, 10 mm Tris·HCl, pH 7.4, 10 mm MgCl2, 1 mm ATP, 10 mm creatine phosphate, and 100 μg/ml creatine kinase in a final volume of 100 μl. The reaction that proceeded at 37° was stopped with 1 ml of ice-cold stop solution (0.25 m sucrose, 100 mm NaCl, 10 mmTris·HCl, pH 7.4). The diluted samples were passed through Millipore filters (GV, 0.22-μm pore size) under a light vacuum. The filters were washed with 4 ml of ice-cold stop solution and then oven-dried at 50° for 10 min. Each filter was placed in scintillation fluid, and the level of radioactivity was measured by liquid scintillation counting.
Chemicals.
[14,15,19,20-3H]LTC4(150 Ci/mmol) was obtained from DuPont-New England Nuclear (Boston, MA). [3H]VCR and [3H]ADM was obtained from Amersham International. PAK-104P (Fig. 1), a newly synthesized pyridine analog (24), was obtained from Nissan Chemical Industries (Chiba, Japan). Cepharanthine, a biscolclaurine alkaloid, was donated by Kaken Pharmaceutical (Osaka, Japan). Other drugs and chemicals were obtained from Sigma Chemical (St. Louis, MO).

Structure of PAK-104P.
Results
Expression of MRP and MDR1 mRNA in MDR KB cell lines.
Slot-blot analyses of KB cell lines are shown in Fig.2A. MRP mRNA was overexpressed in C-A120 cells but not in KB-8–5 cells, whereas MDR1 mRNA was overexpressed in KB-8–5 cells but not in C-A120 cells. Northern blots detected 6.5-kb MRP mRNA in KB-3–1 as well as in C-A120 cells (Fig. 2B).

Expression of MRP andMDR1 genes in KB-3–1, C-A120, and KB-8–5 cells. A, Slot blot analysis of MRP and MDR1 mRNA from the three KB cell lines. Total RNA (10, 3, or 1 μg) was applied to each well and hybridized with MRP or MDR1 probe. B, Northern blots of KB-3–1 and C-A120 cells. Polyadenylated RNA was Northern-blotted using a 1-kb cDNA fragment of MRP as the probe. To compare the amount of loaded RNA, the blots were also hybridized with β-actin probe.
Expression of MRP, P-Gp, and topoisomerase II in the KB cell lines.
To test for the expression of MRP and P-Gp in membrane vesicles from the KB cell lines, we probed immunoblots of membrane proteins with anti-MRP peptide polyclonal antibody and C219, a monoclonal antibody against P-Gp, respectively. MRP was detected in C-A120 membrane vesicles but not in those from KB-3–1 and KB-8–5 cells (Fig. 3). Because DNA topoisomerase II is the main target of etoposide and ADM, the expression level of topoisomerase II in the KB cell lines was evaluated by using a polyclonal antibody against topoisomerase IIα. The level of topoisomerase IIα was not decreased in KB-8–5 cells but slightly decreased in C-A120 cells (Fig.3).
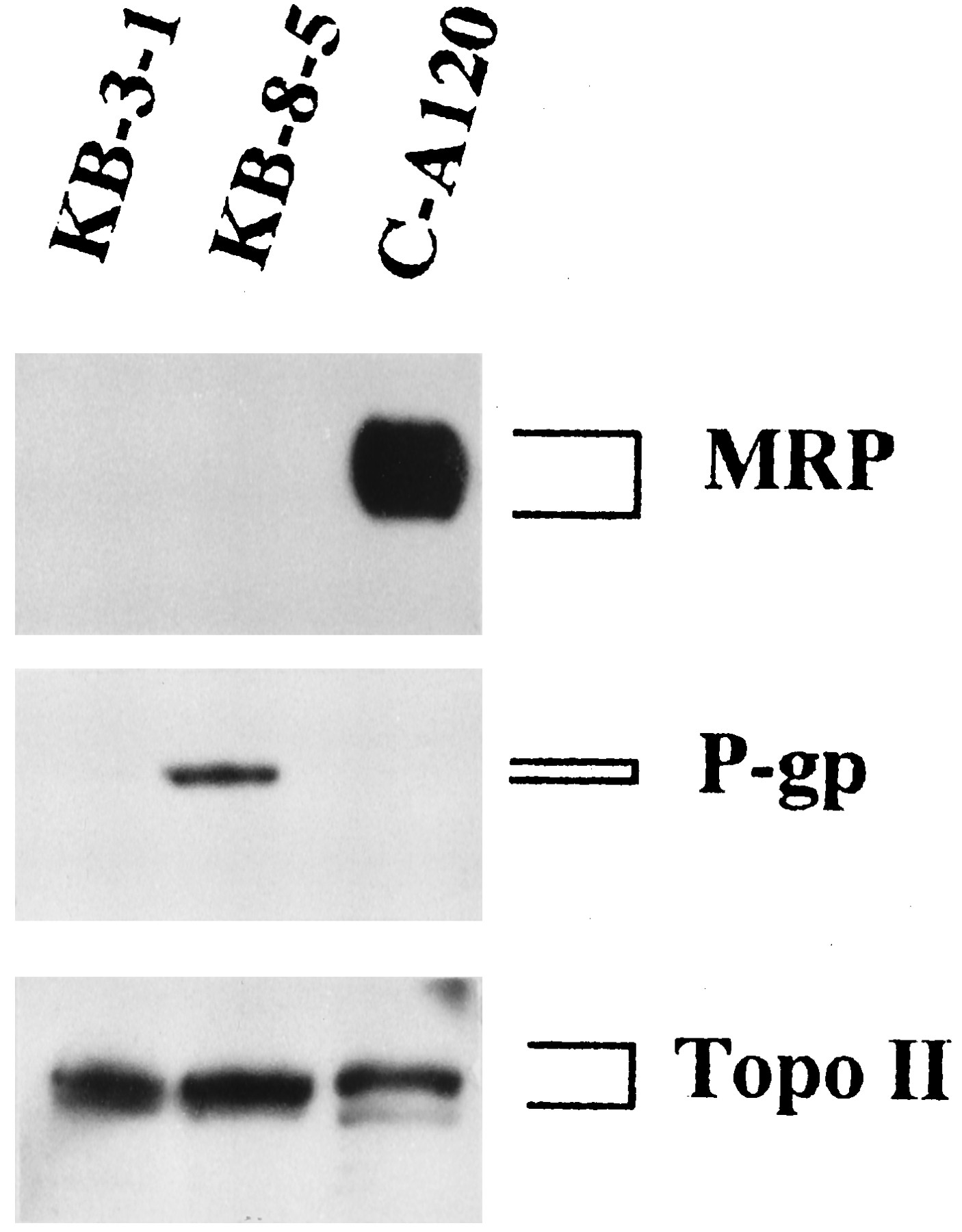
Immunoblot analysis for MRP, P-Gp, and topoisomerase II (Topo II) in MDR KB cells. Membrane vesicles (20 μg of protein) or nuclear protein fractions (7.5 μg) from KB-3–1, KB-8–5, and C-A120 cells were separated by SDS-polyacrylamide gel electrophoresis and transferred to a PVDF membrane. Immunoblot analysis of the transferred proteins with the antibody against MRP, P-Gp, or topoisomerase II was carried out as described in Materials and Methods.
Reversal of resistance to ADM and VCR in C-A120 and KB-8–5 cells by MDR-reversing agents.
MDR-reversing agents verapamil, cepharanthine, and PAK-104P were selected as agents for this study because verapamil is one of the best studied MDR-reversing agents and the other agents have a greater ability than verapamil to reverse drug resistance in KB-C2 cells (24, 40). We examined the cytotoxic effect of the MDR-reversing agents by the MTT method. Cepharanthine (≤5 μm), PAK-104P (≤10 μm), and verapamil (≤3 μm) had no cytotoxic effect on KB-3–1, KB-8–5, or C-A120 cells (data not shown). The dose-response curves of ADM and VCR with and without the reversing agents were assayed by the MTT method. Table 1 summarizes the data from the curves. PAK-104P almost completely reversed the resistance to ADM in KB-8–5 cells, and other reversing agents (cepharanthine and verapamil) moderately reversed the resistance. In contrast, cepharanthine and verapamil did not reverse the resistance to ADM in C-A120 cells, and PAK-104P moderately reversed the ADM resistance. On the other hand, 5 μm cepharanthine and 3 μm PAK 104P completely reversed the resistance to VCR in KB-8–5 cells, and 3 μm verapamil moderately reversed the resistance.
Effects of MDR-reversing agents on ADM and VCR cytotoxicity
Cepharanthine and verapamil moderately reversed the resistance to VCR in C-A120 cells, but 10 μm PAK-104P almost completely reversed the resistance. However, PAK-104P could not enhance the sensitivity of KB-3–1 and C-A120 cells to melphalan or cytosine arabinoside, which are not transporting substrates for MRP.
Effect of agents on cellular accumulation of [3H]VCR.
To investigate how PAK-104P completely reverses the resistance to VCR in C-A120 cells, we examined its effect on the accumulation of VCR in KB-3–1 and C-A-120 cells (Fig. 4).

Accumulation of VCR in KB-3–1 and C-A120 cells. The intracellular levels of VCR in the absence (▩) or presence of the reversing agents PAK-104P [3 μm (░) or 10 μm (▨)], cepharanthine [3 μm (▩)], and verapamil [3 μm (▨)] were determined as described in Materials and Methods. Bars, mean ± standard error of triplicate determinations. ∗∗, p < 0.01; ∗, p < 0.05.
The intracellular concentration of VCR in C-A120 cells was approximately three fifths of that in KB-3–1 cells. The addition of PAK-104P at 10 μm enhanced the accumulation of VCR in C-A120 cells to the level of that in KB-3–1 cells without PAK-104P (Fig. 4). PAK-104P at 10 μm enhanced the accumulation of VCR in KB-3–1 cells by 1.3-fold. Cepharanthine at 3 μm did not enhance the accumulation of VCR in either KB-3–1 or C-A120 cells. Although 3 μmverapamil significantly enhanced the accumulation of VCR in C-A120 cells, the effect was less than that of PAK-104P at the same concentration. Verapamil at 3 μm did not enhance the accumulation of VCR in KB-3–1 cells.
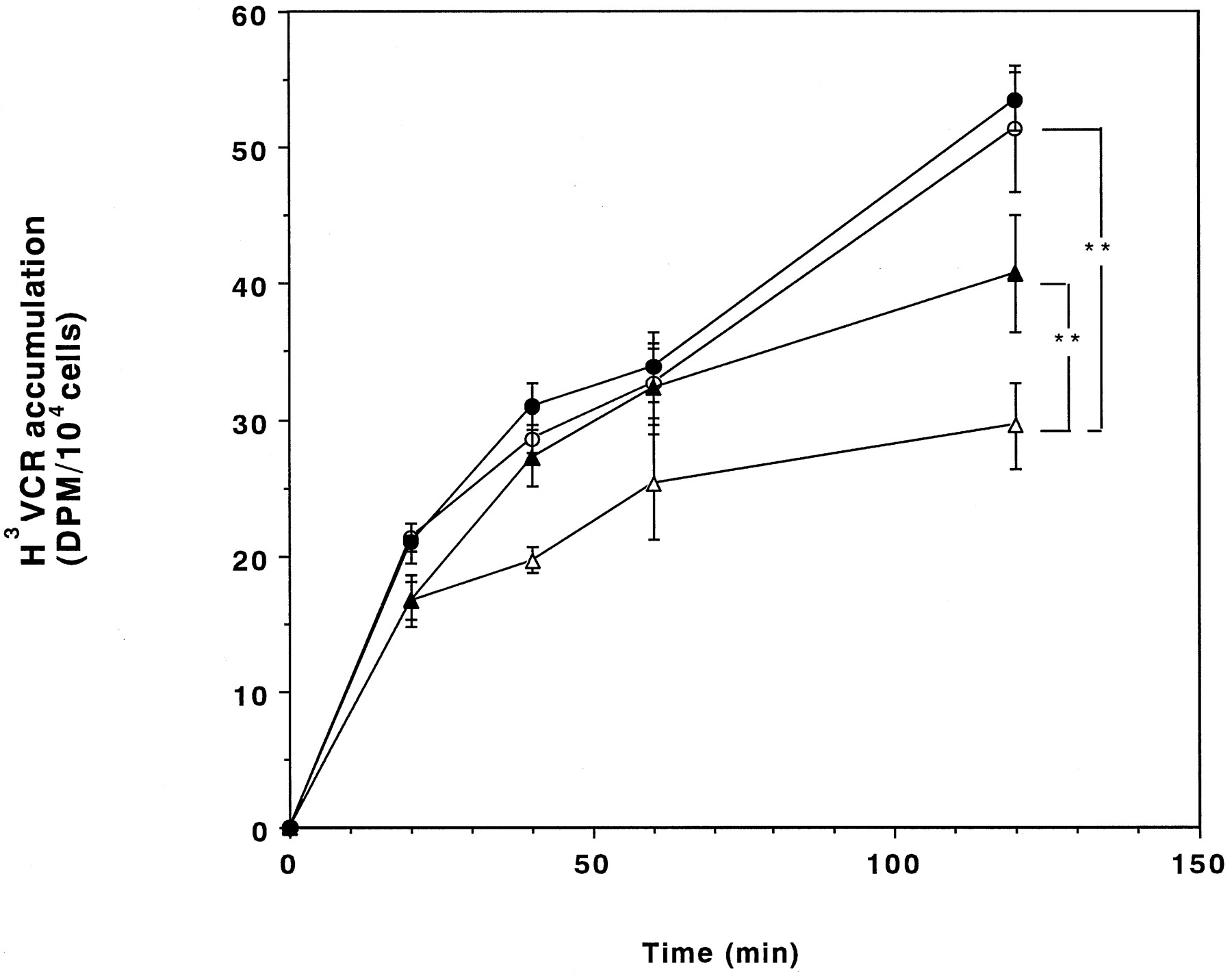
The effects of PAK-104P on the accumulation of VCR as a function of time were compared in KB-3–1 and C-A120 cells. The accumulation of VCR in C-A120 cells was less until 2 hr and enhanced to a greater extent by PAK-104P than that in KB-3–1 cells (Fig. 5).

Accumulation of VCR as a function of time. VCR accumulation in KB-3–1 (○ and •) and C-A120 (▵ and ▴) cells over 2 hr was measured in the absence (○ and ▵) or presence (• and ▴) of 10 μm PAK-104P. Bars, mean ± standard error of triplicate determinations. ∗∗,p < 0.01.
Drug efflux.
We examined whether increased accumulation of VCR in C-A120 cells by PAK-104P was due to inhibition of VCR efflux. Release of VCR as a function of time after a 2-hr period of accumulation is shown in Fig. 6. C-A120 cells released a higher percentage of VCR than KB-3–1 cells. At 20 min, ∼45–50% of cell-associated VCR was lost from C-A120 cells, whereas >85–90% of VCR was retained in KB-3–1 cells. PAK-104P apparently inhibited the efflux of VCR from the both cells, but the extent of the inhibition by PAK-104P in C-A120 cells was greater than that in KB-3–1 cells at 10 and 20 min.

Efflux of VCR. VCR retained in KB-3–1 (○ and •) and C-A120 (▵ and ▴) in the absence (○ and ▵) or presence (• and ▴) of PAK-104P is shown after a 2-hr period of VCR accumulation. Bars, mean ± standard error of triplicate determinations.
Effect of agents on [3H]LTC4 uptake in membrane vesicles.
The effects of cepharanthine, verapamil, and PAK-104P on the ATP-dependent [3H]LTC4uptake in C-A120 vesicles were examined (Fig. 7). The amount of ATP-dependent [3H]LTC4 uptake in C-A120 membrane vesicles was 12-fold higher than that in KB-3–1 vesicles.

Effect of the reversing agents on the ATP-dependent [3H]LTC4 uptake in C-A120 membrane vesicles. Membrane vesicles (50 μg of protein) were incubated with 1.34 nm [3H]LTC4 at 37° in 100 μl of 0.25 m sucrose containing 10 mm Tris·HCl, pH 7.4, 10 mm MgCl2, 10 mm creatine phosphate, 100 μg/ml creatine kinase, and the indicated concentrations of each agent in the presence or absence of 1 mm ATP. ATP-dependent accumulation was calculated from the difference in the radioactivity incorporated into the vesicles in the presence or absence of ATP. Bars, mean ± standard error of triplicate determinations. ∗∗, p < 0.01.
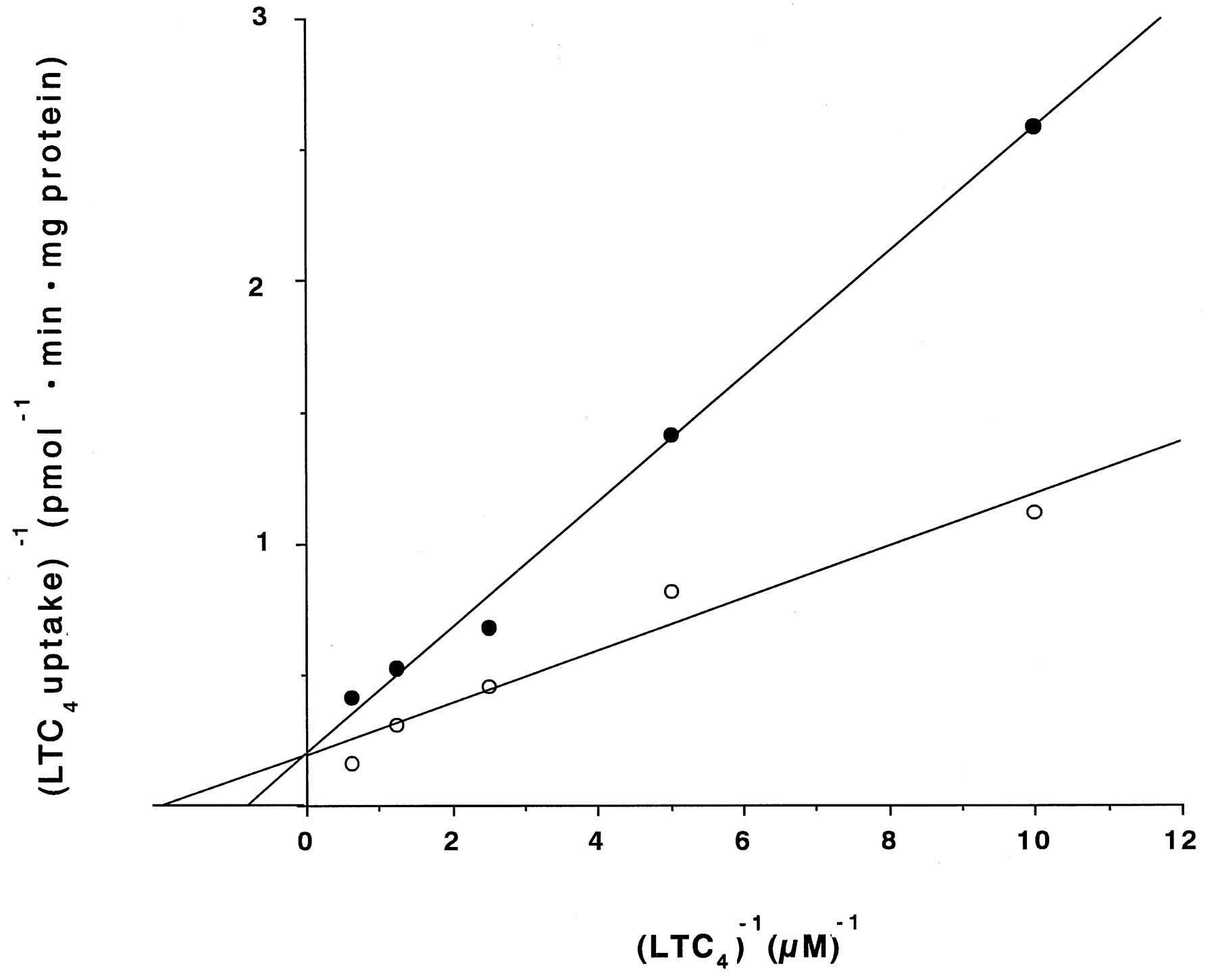
Cepharanthine (100 μm), verapamil (100 μm), and PAK-104P (50 and 100 μm) reduced the [3H]LTC4 uptake in C-A120 vesicles by 5%, 17%, 61%, and 98%, respectively. A Lineweaver-Burk plot in the absence or presence of 50 μm PAK-104P showed that the agent had the property of a competitive inhibitor for the inhibition of the ATP-dependent LTC4 uptake in C-A120 membrane vesicles (Fig. 8).

Double-reciprocal plots of ATP-dependent LTC4 uptake. The amount of LTC4 taken up in C-A120 vesicles within the first 2 min of incubation in the absence (○) or presence (•) of 50 μm PAK-104P was measured as described in Materials and Methods.
Discussion
The use of resistance modifiers, including calcium channel blockers, calmodulin inhibitors, antiestrogens, and the antimalarial drug chloroquine, was reportedly not effective in non-P-Gp-mediated MDR (23). However, the circumvention of ADM resistance was examined using H69AR cells, which overexpress MRP (8) and have a decreased amount of topoisomerase II (23). We recently isolated non-PG-p-mediated MDR cells (C-A120, C-A500, and C-A1000) and indicated that both the increased expression of MRP and decreased levels of topoisomerase IIα underlie the drug resistance in the non-P-Gp-mediated MDR cells (9). Topoisomerase is quantitatively or qualitatively changed in many other non-P-Gp-mediated MDR cells (12, 41, 42). One of the target molecules for ADM is topoisomerase II, and the decreased level of topoisomerase II causes resistance to ADM, which may be difficult to reverse by MDR-reversing agents that interact with P-Gp.
In the current study, we examined whether the MDR-reversing agents that interact with P-Gp are able to reverse the resistance to VCR and ADM in MRP-mediated MDR cells (C-A120) in which the level of topoisomerase II is slightly decreased. Among three MDR-reversing agents used in this study, PAK-104P alone almost completely reversed the resistance to VCR in C-A120 cells as well as in KB-8–5 cells. Regarding the reversal of the ADM resistance in C-A120 cells, no agents completely reversed the resistance. PAK-104P at 3 μm moderately reversed the ADM resistance, but the other agents at the same concentration did not. The decreased level of topoisomerase II in C-A120 cells may be the reason why PAK-104P, which completely reversed the resistance to VCR, could not completely reverse the resistance to ADM in C-A120 cells, because PAK-104P completely reversed the reduced ADM accumulation in C-A120 cells (data not shown). Cepharanthine almost completely reversed the resistance to VCR in KB-8–5 cells but only moderately reversed that in C-A120 cells. These findings suggest that cepharanthine preferentially interacts with P-Gp but that PAK-104P strongly interacts with both P-Gp and MRP.
Most of the non-P-Gp-mediated MDR cells had a drug accumulation deficit (11, 14, 43-46). The accumulation of VCR in C-A120 cells was also decreased (9), and PAK-104P, but not cepharanthine, restored the accumulation of VCR in C-A120 cells. PAK-104P inhibited the efflux of VCR from KB-3–1 cells, increased the accumulation of VCR in KB-3–1 cells, and enhanced the sensitivity of KB-3–1 cells to VCR by a factor of 2.5. PAK-104P may inhibit the function of the MRP expressed in KB-3–1 cells. However, we assume that unknown molecule or molecules other than MRP are also involved in these phenomena, since we detected MRP mRNA at low level but could not detect any expression of MRP in KB-3–1 cells. Melpharan and cytosine arabinoside seem not to interact with both MRP and the unknown molecule or molecules; C-A120 cells were not resistant to these anticancer agents, and PAK-104P did not enhance the sensitivity of KB-3–1 and C-A120 cells to these agents. The mechanism of the transport of anthracyclines and Vinca roseaalkaloids by MRP is still unknown. MRP was not photolabeled with a photoanalog of vinblastine and a photoactive MDR-reversing agent, azidopine (7). These findings suggested that vinblastine and azidopine do not directly bind to MRP and therefore are not transported by MRP. Recently, Jedlitschky et al. reported that LTC4, a substrate for the GS-X pump (43), is transported by MRP and that MRP is photolabeled with LTC4 (15). ATP-dependent [3H]LTC4 uptake in membrane vesicles from C-A120 cells was enhanced; the uptake was almost completely inhibited by 100 μm PAK-104P but not by cepharanthine. Kinetic analysis of the inhibition type with the Lineweaver-Burk plot suggested that PAK-104P and LTC4 compete for the same binding site on the MRP surface. PAK-104P seems to interact directly with MRP and inhibits the transporting activity of the GS-X pump. Although it is not known whether VCR and ADM are conjugated with glutathione or forms other negatively charged complexes, Ishikawa et al. proposed a putative metabolic pathway that yields the glutathioneS-conjugates of ADM (47). It is probable that MRP transports such a negatively charged complex and PAK-104P inhibits the transport. PAK-104P significantly inhibited the LTC4 uptake in C-A120 vesicles at 10 μm that is needed for the reversal of drug resistance and restoration of the VCR accumulation deficit in C-A120 cells. PAK-104P at 100 μm almost completely inhibited the LTC4 uptake in C-A120 vesicles. The density of the membrane fractions used for the LTC4 transport assay, when converted to cell density, was ∼10-fold higher than that used for the drug accumulation study. This may be why the concentration of PAK-104P needed to inhibit the LTC4 transport was higher than that needed to reverse the drug resistance and the accumulation deficit. Cepharanthine moderately reversed the VCR resistance in C-A120 cells but did not affect the accumulation of VCR in C-A120 cells or the uptake of LTC4 in C-A120 vesicles. Cepharanthine may interact with other molecule or molecules than MRP to reverse the VCR resistance.
Many agents that reverse P-Gp-mediated drug resistance have been reported (3), and some agents, such as MK571, an LTD4receptor antagonist (22), and genistein, a protein kinase inhibitor (48), modulated MRP-associated MDR. However, no agents that completely reverse both P-Gp- and MRP-mediated drug resistance have been reported. PAK-104P should be useful in treating patients with tumors that overexpress both P-Gp and MRP.
Footnotes
-
Send reprint requests to: Dr. Shin-ichi Akiyama, Institute for Cancer Research, Kagoshima University, 8–35-1 Sakuragaoka, Kagoshima 890, Japan. E-mail:akiyamas{at}khosp2.kufm.kagoshima-u.ac.jp
- Abbreviations:
- MDR
- multidrug resistance (or resistant)
- P-Gp
- P-glycoprotein
- LTC4
- leukotriene C4
- LTD4
- leukotriene D4
- MRP
- multidrug resistance-associated protein
- GS-X pump
- ATP-dependent glutathioneS-conjugate export pump
- ADM
- doxorubicin
- HRP
- horseradish peroxidase
- PVDF
- polyvinylidene difluoride
- VCR
- vincristine
- MTT
- 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
- PBS
- phosphate-buffered saline
- SSC
- standard saline citrate
- SDS
- sodium dodecyl sulfate
- Received May 21, 1996.
- Accepted October 23, 1996.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}