


Abstract
Muscarinic acetylcholine receptors bind allosteric modulators at a site apart from the orthosteric site used by conventional ligands. We tested in cardiac tissue whether modulator binding to ligand-occupied muscarinic M2 receptors is a preferential event that can be detected using a radioactive allosteric agent. The newly synthesized dimethyl-W84 (N,N′-bis[3-(1,3-dihydro-1,3-dioxo-4-methyl-2H-isoindol-2-yl)propyl]-N,N,N′,N′-tetramethyl-1,6-hexanediaminium diiodide) has a particular high potency at M2 receptors occupied by the conventional antagonistN-methylscopolamine (NMS); dissociation of [3H]NMS is half-maximally retarded at an EC50,diss value of 3 nm. Using obidoxime as an “allosteric antagonist,” evidence was found that dimethyl-W84 interacts with the postulated common allosteric site. Binding of [3H]dimethyl-W84 (0.3 nm; specific activity, 168 Ci/mmol) was measured in porcine heart homogenates (4 mm Na2HPO4, 1 mmKH2PO4, pH 7.4, 23°) in the presence of 1 μm NMS. Homologous competition experiments revealed two components of saturable radioligand binding: one with a high affinity (K D = 2 nm) and small capacity (≈30% of total saturable binding) and the other with a 20,000-fold lower affinity. The B max value of the high affinity sites (68 fmol/mg protein) matched muscarinic receptor density as determined by [3H]NMS (79 fmol/mg). Prototype allosteric agents, alcuronium, W84 (the parent compound of the radioligand), and gallamine, displaced high affinity [3H]dimethyl-W84 binding concentration-dependently (pK i values = 8.62, 7.83, and 6.72, respectively). The binding affinities of the modulators were in excellent correlation with their potencies to allosterically stabilize NMS/receptor complexes (EC50,diss = 8.40, 7.72, and 6.74, respectively). We conclude that high affinity binding of [3H]dimethyl-W84 reflects occupation of the common allosteric site of M2 receptors.
The allosteric modulation of ligand-gated ion channels is a well known phenomenon that is therapeutically used with the benzodiazepines at γ-aminobutyric acidA receptors. However, also in the comparably small G protein-coupled receptors, simultaneous occupation by two different ligands of the transmitter binding site and of a distinct allosteric site seems possible [e.g., in adenosine A1 receptors (Bruns and Fergus, 1990), dopamine D2 receptors (Hoare and Strange, 1996), and α2-adrenergic receptors (Wilson et al., 1992)]. Ternary complexes especially have been studied in muscarinic acetylcholine receptors (Lee and El-Fakahany, 1991; Lazareno and Birdsall, 1995; Tuc̆ek and Proška, 1995). As a consequence of alloster binding to the muscarinic receptor protein, the interaction between a ligand, such as the endogenous agonist acetylcholine or the antagonist NMS, and the orthosteric site of the receptor is altered. Ligand binding may be elevated, which opens the perspective to develop enhancers of acetylcholine binding that amplify cholinergic transmission at muscarinic synapses in a subtype-specific fashion (Birdsall et al., 1997;Jakubı́k et al., 1997). The event underlying an elevation of ligand binding to muscarinic receptors on the molecular level is a retardation of ligand dissociation that results from binding of the allosteric agent to the ligand/receptor complex. Allosteric agents also inhibit ligand association by an interaction with the free receptor, yet phenomenologically, this action resembles the action of competitive antagonists (Kostenis and Mohr, 1996).
Until now, the binding characteristics of allosteric agents had to be derived from the effects they induced on ligand binding to the orthosteric site. Due to the lack of a radiolabeled allosteric agent, it has not been possible so far to investigate directly the binding of agents at the allosteric site. Furthermore, it has not been possible to reject by direct measurements the criticism that allosteric actions on ligand-occupied muscarinic receptors might be only an aspect of a rather nonspecific attachment to surface structures of cellular membranes, although there is ample evidence for a specific and preferential interaction.
The M2 receptor has been postulated to contain a “common allosteric site” used by various structurally heterogenous modulators (Ellis and Seidenberg, 1992; Tränkle and Mohr, 1997). This site is thought to be located at the entrance of the ligand binding pocket of the receptor protein (Tuc̆ek and Proška, 1995; Tränkle et al., 1998). Based on previous structure-activity relationship studies (Holzgrabe et al., 1996; Holzgrabe and Hopfinger, 1996), we intended to develop a highly potent modulator of ligand binding at M2receptors suitable to be applied in radioactive form for direct binding measurements. The compound (Fig. 1) is a dimethylated derivative of the alkane-bis-ammonium compound W84, which previously has been discussed as a lead structure to develop new allosteric agents (Tränkle et al., 1996). In the first part of our study, we characterized the allosteric effect of unlabeled dimethyl-W84 on the binding of the orthosteric ligand [3H]NMS in native M2receptors applying porcine heart homogenates.
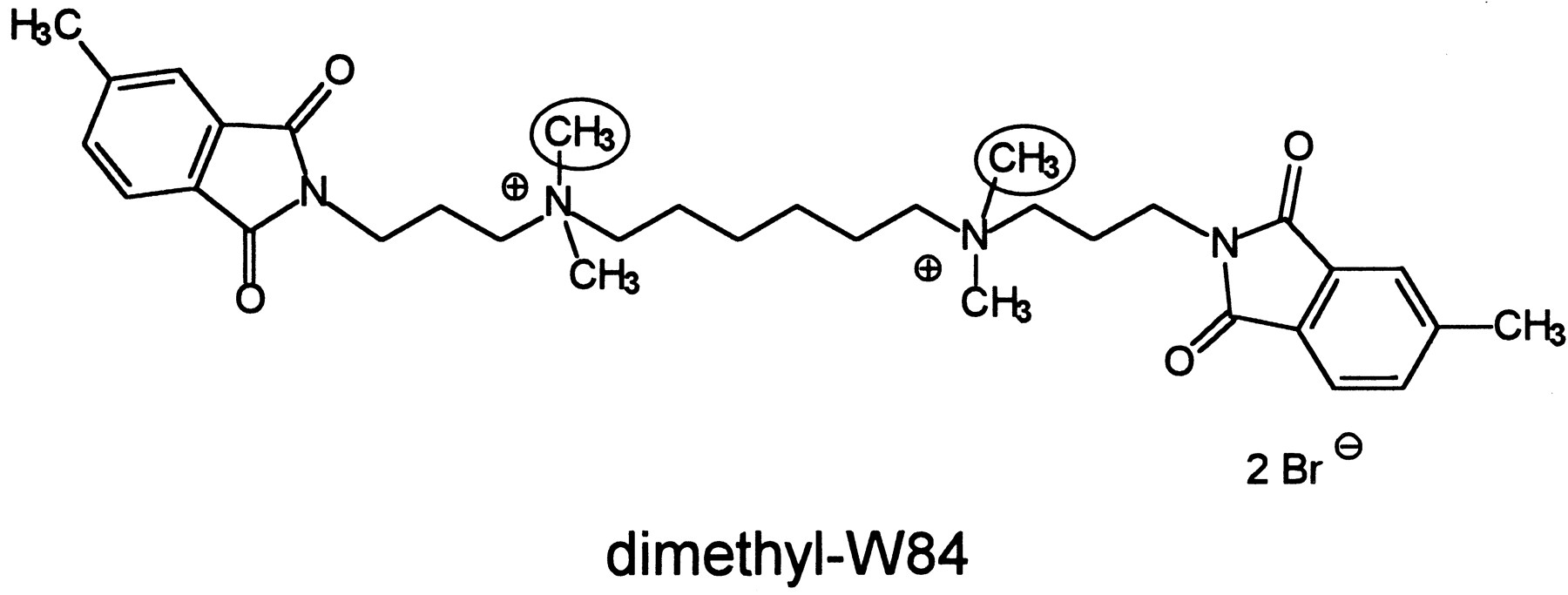
Structure of dimethyl-W84.Encircled, groups replaced by [3H]methyl residues in the radiolabeled compound. The parent compound of dimethyl-W84 (W84) is devoid of the aromatic methyl groups.
[3H]NMS was chosen because this radioligand has been used previously to define the potency of various allosteric agents to interact with NMS-liganded M2 receptors (Tränkle et al., 1996) and to check the common-site hypothesis for the agents alcuronium, W84, and gallamine (Tränkle and Mohr, 1997), which were used here to test the specificity of [3H]dimethyl-W84 binding. The experiments were carried out in a buffer of low ionic strength devoid of divalent cations to provide optimum conditions of a high affinity at the allosteric site. In the second part of the study, the binding of [3H]dimethyl-W84 was investigated under identical conditions. In these experiments, we applied a high concentration of NMS (1 μm) to occupy the orthosteric site of the M2 receptors because of the following reasons. First, as mentioned above, the ability to bind with rather high affinity to ligand-occupied receptors distinguishes allosteric modulators from conventional ligands. Second, the experiments with [3H]NMS as the radioligand suggest that dimethyl-W84 binds with slightly higher affinity to free receptors than to [3H]NMS-liganded receptors. It cannot be excluded that dimethyl-W84 in the free receptor might have a somewhat different orientation compared with NMS-occupied receptors. Therefore, a saturating concentration of NMS was applied to prevent this possibility. Third, measurement of [3H]dimethyl-W84 binding in NMS-occupied receptors allowed a direct comparison with the functional experiments in which the allosteric actions of nonlabeled dimethyl-W84 and other modulators were characterized at [3H]NMS-liganded receptors. Thus, it could be checked whether the site of dimethyl-W84 binding is identical with the site that mediates the actions of dimethyl-W84 and of the other applied allosteric agents.
Materials and Methods
Synthesis of dimethyl-W84.
Equimolar amounts of methylphthalic acid anhydride and 3-(N,N-dimethylamino)propylamine were refluxed in toluene using a water separator. After evaporation of the solvent, the product was purified by means of column chromatography (silica gel, MeOH/CH2Cl2 = 5:3) to give 85% of pure 6-methyl-N-[3-(N,N-dimethylamino)propyl]phthalimide. 1,6-Dibromohexane was alkylated with the phthalimide derivative by refluxing for 4 hr in acetonitrile. After evaporation of the acetonitrile, the remaining oil was washed several times with mixtures of ethanol/ethylacetate. Finally, the oil was dissolved in dichloromethane/ethanol, and the crystallization was initiated by the addition of diethylether to give 20% of pure dimethyl-W84.
Synthesis of the radioligand.
N-(3-Bromopropyl)-4-methylphthalimide and 1,6-bis-(methylamino)hexane (ratio 2:1) were refluxed in acetonitrile for 5 hr to give 2,2′-[2,6-hexanediylbis[(methylimino)-3,1-propanediyl]]bis[4-methyl-1H-isoindole-1,3-(2H)-dione]. After column chromatographic purification (silica gel, MeOH/CH2Cl2 = 5:3), the bistertiary compound can be obtained in a yield of 20%. Methylation of both tertiary nitrogens can be achieved through treatment of the tertiary dimethyl-W84 with an excess of methyliodide in acetonitrile at room temperature (thin layer chromatography control silica gel, ethanol/concentrated NH3 = 4.5:1.5). The addition of diethyl ether to the reaction solution gives 85% of crystalline dimethyl-W84.
The NMR (1H and 13C) and IR data are in agreement with the structures; the elemental analyses are within ±0.4% of the theoretical values.
The synthesis of the corresponding radiolabeled compound [3H]dimethyl-W84 was carried out at Amersham Life Science (Braunschweig, Germany), starting with the bistertiary compound and using the procedure described above. The radiochemical purity was 97%, and the specific activity was 168 Ci/mmol (6.22 TBq/mmol). The stock solution contains ethanol/water/methanol (84:15:1). In mixtures of ethanol and water, as well as in distilled water, no chemical decomposition of the unlabeled compound could be detected. In the buffer of the current study (4 mmNa2HPO4, 1 mmKH2PO4, pH 7.4, 23°), dimethyl-W84 undergoes slow hydrolysis with a half-life of 16 hr, which is manyfold beyond the applied incubation time.
Membrane preparation.
Preparation of porcine heart homogenates was carried out as described in detail previously (Tränkle and Mohr, 1997) at an ambient temperature of 3–6°. In short, ventricular myocardium of freshly excised domestic pig hearts obtained from the local slaughterhouse was cut into small pieces, washed in sucrose solution (0.32 m), and homogenized using a Waring blender and a Potter-Elvejhem homogenizer. The homogenate was centrifuged for 11 min at 300 × g (2,000 rpm in a Beckman rotor 35; Beckman Instruments, Columbia, MD). The resulting supernatant was centrifuged for 41 min at 80,000 × g(32,000 rpm in a Beckman rotor model 35). The homogenates intended for use in a centrifugation assay (see below) instead of a filtration assay were pelleted for 20 min at 20,900 × g (15,300 rpm in a Beckman rotor model F241.5). Pellets were resuspended in a buffer composed of 4 mmNa2HPO4 and 1 mm KH2PO4, pH 7.4 (Na,K,Pi buffer). Aliquots of 1 ml were frozen in liquid nitrogen and stored at −80°. Protein content amounted to 4.7–8.5 mg/ml membrane suspension.
Binding assays.
Cardiac membranes at a protein concentration of 300–700 μg/ml were incubated with the appropriate radioligand in the Na,K,Pi buffer at a temperature of 23°. Assays for homologous and heterologous competition measurements were carried out in volumes of 1.5 ml with an incubation time of 2 hr. For kinetic experiments, assays were prepared in larger volumes, and 1-ml aliquots were removed at appropriate intervals over a total period of 120 min.
Effects of dimethyl-W84 on [3H]NMS binding.
The binding characteristics of [3H]NMS (0.2 nm) under control conditions were investigated by homologous competition experiments. Nonspecific [3H]NMS binding was determined in the presence of 1 μm atropine and did not exceed 5% of total binding when membranes were separated by filtration; in centrifugation experiments (see below), nonspecific [3H]NMS binding was <10% of the total. In filtration experiments, the −log equilibrium dissociation constant of [3H]NMS binding amounted to a pKD value of 9.80 ± 0.11, and the density of binding sites toB max was 100 ± 18 fmol/mg protein (mean ± standard error, three experiments); in centrifugation experiments, pKD was 9.56 ± 0.14, and B max was 79 ± 21 fmol/mg protein (mean ± standard error, six experiments). The effect of dimethyl-W84 on the equilibrium binding of [3H]NMS (0.2 nm) was determined in heterologous inhibition experiments with membrane separation by filtration.
To determine the influence of dimethyl-W84 on the dissociation of [3H]NMS, membranes were incubated for 60 min with dimethyl-W84 and the radioligand before radioligand dissociation was revealed by the addition of 1 μm atropine.
To measure the effect of dimethyl-W84 on the association of [3H]NMS, dimethyl-W84 was incubated with the membranes for 30 min before [3H]NMS was added to start the formation of radioligand/receptor complexes; 1-ml aliquots were drawn at appropriate intervals.
Membranes were separated by rapid filtration (glass fiber filters No. 6; Schleicher & Schuell, Dassel, Germany). Filters were washed twice with 5 ml of ice-cold incubation buffer, dried, and placed into vials containing 5 ml of Ready Protein (Beckman) for liquid scintillation counting.
[3H]Dimethyl-W84 binding assay.
The [3H]dimethyl-W84 experiments were carried out in the presence of 1 μm NMS to occupy the orthosteric binding site of the M2 receptor. NMS was allowed to equilibrate for 5–10 min with the receptors before [3H]dimethyl-W84 was added to the assay. Alkane-bis-ammonium compounds closely related to dimethyl-W84 have been reported to interact with acetylcholinesterase (Ohnesorge, 1969); to inhibit binding of [3H]dimethyl-W84 to acetylcholinesterase, 1 μm physostigmine was added to the incubation mixture. Control experiments revealed that at the indicated concentrations, neither NMS nor physostigmine had an allosteric effect on [3H]NMS dissociation and thus did not interact with the allosteric site of the [3H]NMS-occupied M2receptor (data not shown). Membranes were separated by centrifugation with 20,900 × g (15,300 rpm) in a Microfuge (Beckman) for 20 min at 23°. After drawing off the supernatant, the tubes containing the tightly packed pellet were carefully and quickly rinsed with 1.5 ml of cold Na,K,Pi buffer to remove residual radioactivity from the tube wall. After resuspension of the pellet in 1.5 ml of buffer, the suspension was transferred into a scintillation vial filled with 10 ml of Ready Protein (Beckman) for liquid scintillation counting.
Data analysis.
Data from individual experiments were analyzed by computer-aided, nonlinear regression analysis using Prism (ver. 2.01; GraphPAD Software, San Diego, CA). Analysis of homologous competition data obtained with [3H]NMS was based on the general Hill equation. Because the observed Hill coefficients did not differ significantly from unity (partialF test, p > 0.05, data not shown), IC50 values were determined from curve fits withnH fixed to 1. The binding parametersKD andB max were calculated according to DeBlasiet al. (1989). [3H]Dimethyl-W84 inhibition curves were fitted by applying an equation for a two-site model. The binding parameters KD andB max were derived from the high and low affinity components of this fit according DeBlasi et al. (1989). Ki values for the inhibitory action of the test compounds on [3H]dimethyl-W84 binding were obtained from IC50 values according to Cheng and Prusoff (1973).
Analysis of the effect of dimethyl-W84 on the binding of [3H]NMS was based on the ternary complex model of allosteric interactions of Ehlert (1988) as described previously (eq. 3 in Tränkle et al., 1998).
[3H]NMS dissociation data could be described by a monoexponential decay function that yielded the rate constant of dissociation k −1. [3H]NMS association was monophasic. The rate constant of association was obtained by fitting a straight line to the initial data points of specific [3H]NMS binding (at 30 and 60 sec) as described, for instance, by Bennett and Yamamura (1985).
The retarding action of dimethyl-W84 on the dissociation and association of [3H]NMS was expressed in terms of a reduction of the apparent rate constant of associationk +1 (control, 1.131 ± 0.070 nm −1·min−1, mean ± standard error, four experiments) and dissociationk −1 (control, 0.165 ± 0.002 min−1, mean ± standard error, 16 experiments), respectively. Concentration-effect curves for the diminution of the rate constants were obtained by curve fitting based on a four-parameter logistic function.
The antagonistic action of obidoxime on the dimethyl-W84-induced retardation of [3H]NMS dissociation was analyzed according to Lazareno and Birdsall (1993) as described previously (eq. 4 in Tränkle et al., 1998).
Drugs.
[3H]NMS was purchased from DuPont-New England Nuclear (Bad Homburg, Germany). Alcuronium dichloride was generously provided by Hoffmann-La Roche AG (Grenzach-Wyhlen, Germany). Obidoxime chloride was a gift of Merck KG (Darmstadt, Germany). Atropine sulfate, (−)-scopolamineN-methylbromide, and gallamine triethiodide were from Sigma Chemical (München, Germany). W84 was synthesized by Dr. Joachim Pfeffer (University of Kiel, Kiel, Germany).
Results
Allosteric interaction of unlabeled dimethyl-W84 with [3H]NMS binding.
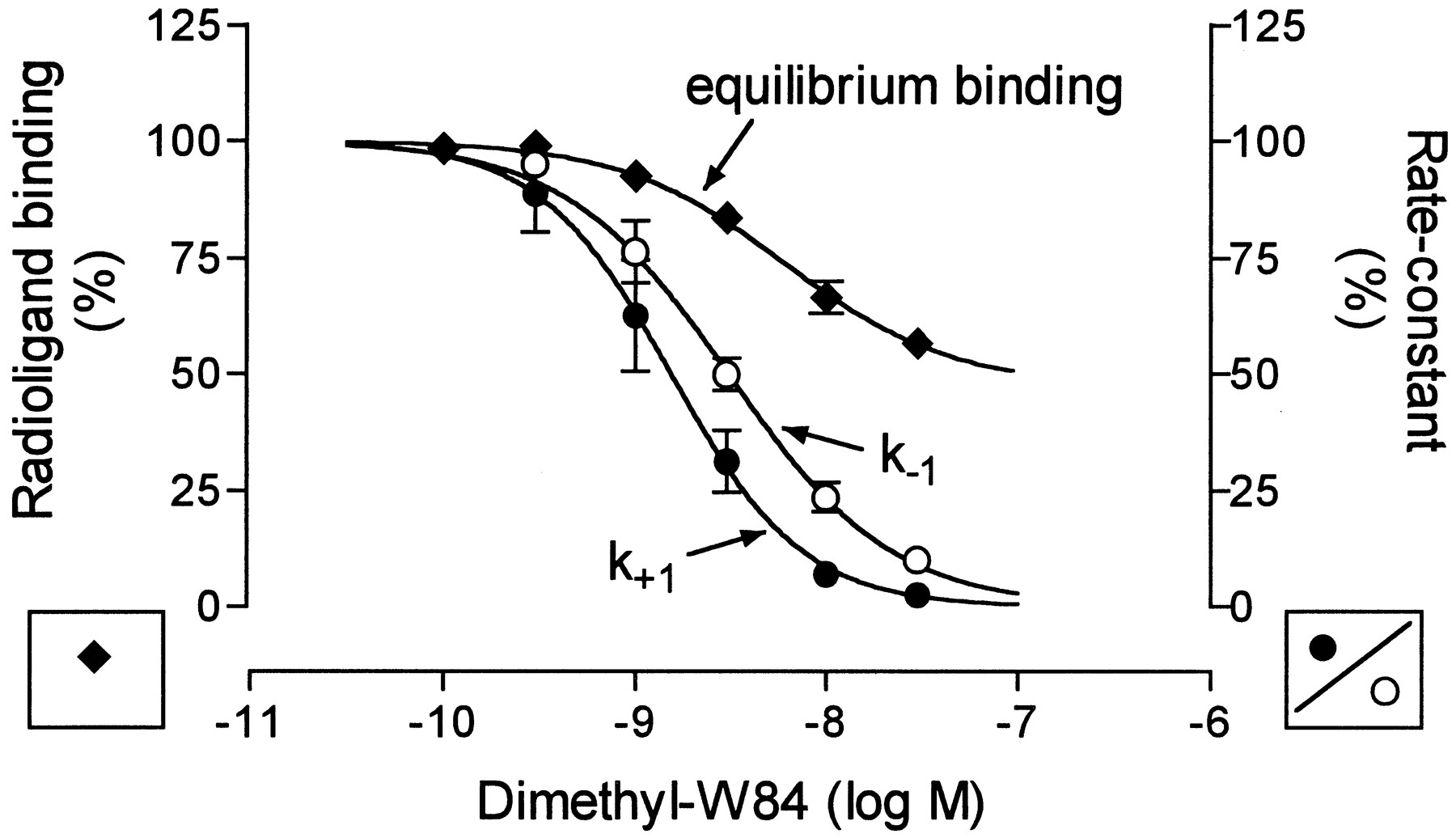
Dimethyl-W84 allosterically inhibits the dissociation of [3H]NMS from porcine cardiac M2 receptors. The concentration-effect curve is illustrated in Fig.2 using the apparent rate constant of [3H]NMS dissociation,k −1, as a measure of the dissociation rate. The curve has a slope not different from unity (partialF test, p > 0.05) and levels off atk −1 = 0, indicating that bound dimethyl-W84 may completely prevent the dissociation of [3H]NMS/receptor complexes. The concentration of dimethyl-W84 causing a 50% decrease in thek −1 of [3H]NMS dissociation served as a measure of potency and amounted to pEC50,diss = 8.51 ± 0.02 (mean ± standard error, eight experiments; Table1). This value should reflect the affinity of dimethyl-W84 at NMS-occupied M2receptors (Lazareno and Birdsall, 1995).

Effects of dimethyl-W84 on the equilibrium binding of the muscarinic receptor antagonist [3H]NMS in porcine heart homogenates and on the association and dissociation of [3H]NMS. Left ordinate, specific binding of 0.2 nm [3H]NMS (♦) plotted as percentage of the control in the absence of dimethyl-W84. Indicated are mean ± standard error values of four experiments performed as triplicate determinations. Curve fitting was based on the ternary complex model of allosteric interactions (Ehlert, 1988). Right ordinate, apparent rate constants of association k +1(•) and dissociation k −1 (○) shown as percentage of the respective controls in the absence of dimethyl-W84. Given are mean ± standard error of one to four independent association and two to eight dissociation experiments.Scatterbars, not shown when they do not exceed the symbols.
Potency of the test compounds to retard allosterically the dissociation of [3H]NMS from M2 receptors in porcine heart homogenates and affinity of the test compounds to displace [3H]dimethyl-W84 binding (0.3 nm) measured in the presence of 1 μm NMS under otherwise identical conditions
Concentration-effect curves for the action of dimethyl-W84 on [3H]NMS association and on the equilibrium binding of [3H]NMS also are included in Fig. 2. The inhibitive action of dimethyl-W84 on [3H]NMS association results from occupation of free M2 receptors by the modulator. The concentration-effect curve approaches k +1 = 0, suggesting that the radioligand does not gain access to M2 receptors occupied by dimethyl-W84. The curve has a slope of unity and its inflection point at pEC50,ass = 8.81 ± 0.04 (mean ± standard error, four experiments). The curve for the action of dimethyl-W84 on [3H]NMS equilibrium binding does not attain the zero level of radioligand binding. At higher concentrations of dimethyl-W84, equilibrium binding of [3H]NMS was no longer attained during the incubation time of 120 min due to the retarding action of the modulator on [3H]NMS binding kinetics. The curve fit is based on the ternary complex model of allosteric interactions (Ehlert, 1988). According to this model, the −log equilibrium dissociation constant for dimethyl-W84 binding to the free M2receptor amounts to pKA = 8.32 ± 0.11, the cooperativity factor is α = 3.2 ± 0.12 (mean ± standard error, four experiments), and the equilibrium dissociation constant for dimethyl-W84 binding at the [3H]NMS-occupied receptor is −log(α·KA ) = 7.82. These values are in acceptable correspondence with the respective binding constants as derived from the kinetic measurements. Thus, independent of the experimental approach the affinity of dimethyl-W84 for the free receptor can be predicted to be only slightly higher than the affinity for the NMS-occupied receptor.
Interaction of dimethyl-W84 with the “allosteric antagonist” obidoxime.
One aim was to check whether the action of dimethyl-W84 may be attributed to an interaction with the proposed common allosteric site of M2 receptors; therefore, we measured whether obidoxime is capable of antagonizing the allosteric action of dimethyl-W84 on [3H]NMS dissociation. A complete concentration-effect curve for dimethyl-W84 was determined in the presence of a high concentration of obidoxime; this curve was shifted to the right in a parallel fashion compared with the control curve (Fig. 3). The antagonistic effect of intermediate concentrations of obidoxime was studied using a single fixed concentration of 30 nm dimethyl-W84. The obidoxime-induced inhibition of the effect of dimethyl-W84 is compatible with a competitive interplay between obidoxime and dimethyl-W84. The pKB of obidoxime amounts to 4.71 ± 0.19 (mean ± standard error, two or three experiments).

Obidoxime-induced attenuation of the effect of dimethyl-W84 on [3H]NMS dissociation.Ordinate, k −1 (normalized) represents the apparent rate constant of [3H]NMS dissociation normalized to compensate for the delay of [3H]NMS dissociation induced by obidoxime alone (for details, see Tränkle and Mohr, 1997) [i.e.,k −1 observed in the presence of obidoxime alone was set at k −1 (normalized) = 1.0]. At the highest obidoxime concentration applied, various concentrations of dimethyl-W84 were applied to check for a parallel curve shift. The effect of dimethyl-W84 in the absence of obidoxime (○, see Fig. 2) is included. Otherwise, the action of dimethyl-W84 at a fixed concentration of 30 nm was measured in the presence of the indicated obidoxime concentrations. Given are mean ± standard error values of two to three data points, each derived from complete dissociation curves. Simultaneous nonlinear fitting of both the data in the presence of obidoxime and the control data was done according toLazareno and Birdsall (1993) as described previously (Tränkleet al., 1998). The data could sufficiently be described by a competitive model (dashed lines).
Binding of [3H]dimethyl-W84 to NMS-occupied receptors.
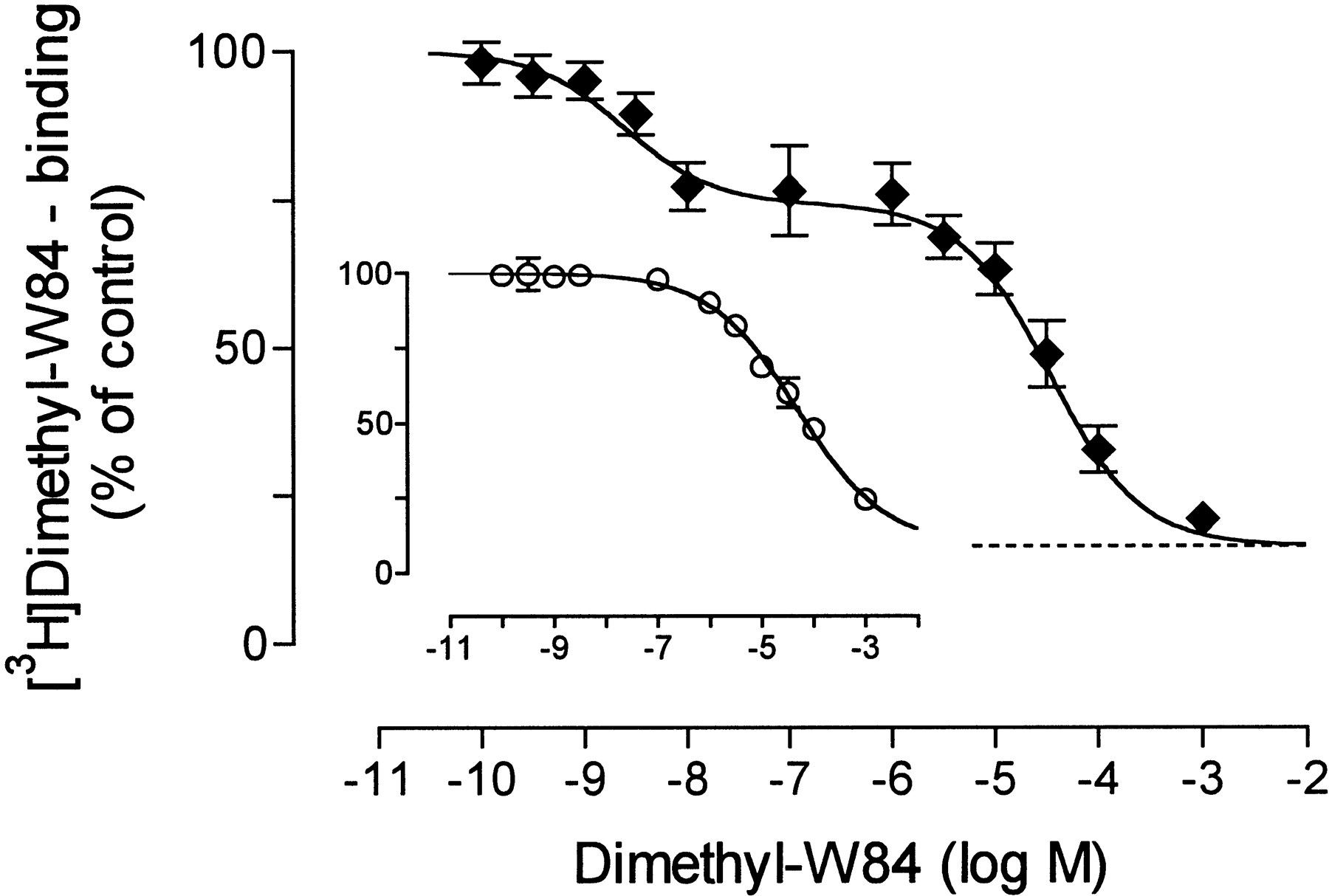
[3H]Dimethyl-W84, at a concentration of 0.3 nm, was incubated with increasing concentrations of unlabeled dimethyl-W84 in the presence of 1 μm NMS to occupy the orthosteric site. As shown in Fig.4, binding of the radioligand was diminished, indicating a saturable binding of [3H]dimethyl-W84. The curve plateaus above the zero level of [3H]dimethyl-W84 binding, thus revealing a small component of nonsaturable radioligand binding. The curve for inhibition of saturable [3H]dimethyl-W84 binding was biphasic. The high affinity component of [3H]dimethyl-W84 binding is characterized by a capacity of 31% of the total saturable binding and by an inflection point at pIC50,high = 8.61. As for a homologous competition curve, the equilibrium dissociation constant for [3H]dimethyl-W84 binding to the NMS-occupied receptors was calculated according to DeBlasi et al. (1989) as a pKD ,high value of 8.74 ± 0.09 (mean ± standard error, seven experiments); the binding site density was a B max value of 68 ± 11 fmol/mg protein. The low affinity component has its inflection point at a pIC50,low value of 4.43 corresponding to pKD , low = 4.43 ± 0.08 (B max = 3.9 ± 0.8 nmol/mg protein). It was checked whether the high affinity component of [3H]dimethyl-W84 vanished in membranes pre-exposed to 80° for 15 min, under which condition the specific binding of [3H]NMS was abolished. The total binding of [3H]dimethyl-W84 was not affected considerably, yet the shape of the inhibition curve changed. Notably, as shown in Fig. 4 (inset), the heat-pretreated membranes did not reveal a high affinity component of saturable [3H]dimethyl-W84 binding. A low affinity component of saturable [3H]dimethyl-W84 binding was still present. The slope factor of the curve amounted to annH value of 0.48, which may indicate that the low affinity sites became heterogeneous on heat treatment. The formation of sites with somewhat higher affinity may explain that total [3H]dimethyl-W84 binding in the absence of unlabeled dimethyl-W84 was not altered under heat treatment.

Inhibition of total binding of [3H]dimethyl-W84 (0.3 nm) in porcine heart homogenates by unlabeled dimethyl-W84. Binding of [3H]dimethyl-W84 in the absence of competitor was set at 100% (corresponding to bound radioactivity of ∼13,000 dpm/1.5 ml of assay volume). Given are mean ± standard error values of seven experiments with each data point determined in quadruplicate. Error bars are not shown when they do not exceed the symbols. The curve fit is based on a two-site model. Horizontal dashed line, bottom level of the curve. Inset, inhibition curve obtained in heat-pretreated membranes; for details, see text. Indicated are mean ± standard error values of five experiments performed as quadruplicate determinations. Curve fitting was based on the four-parameter logistic function.
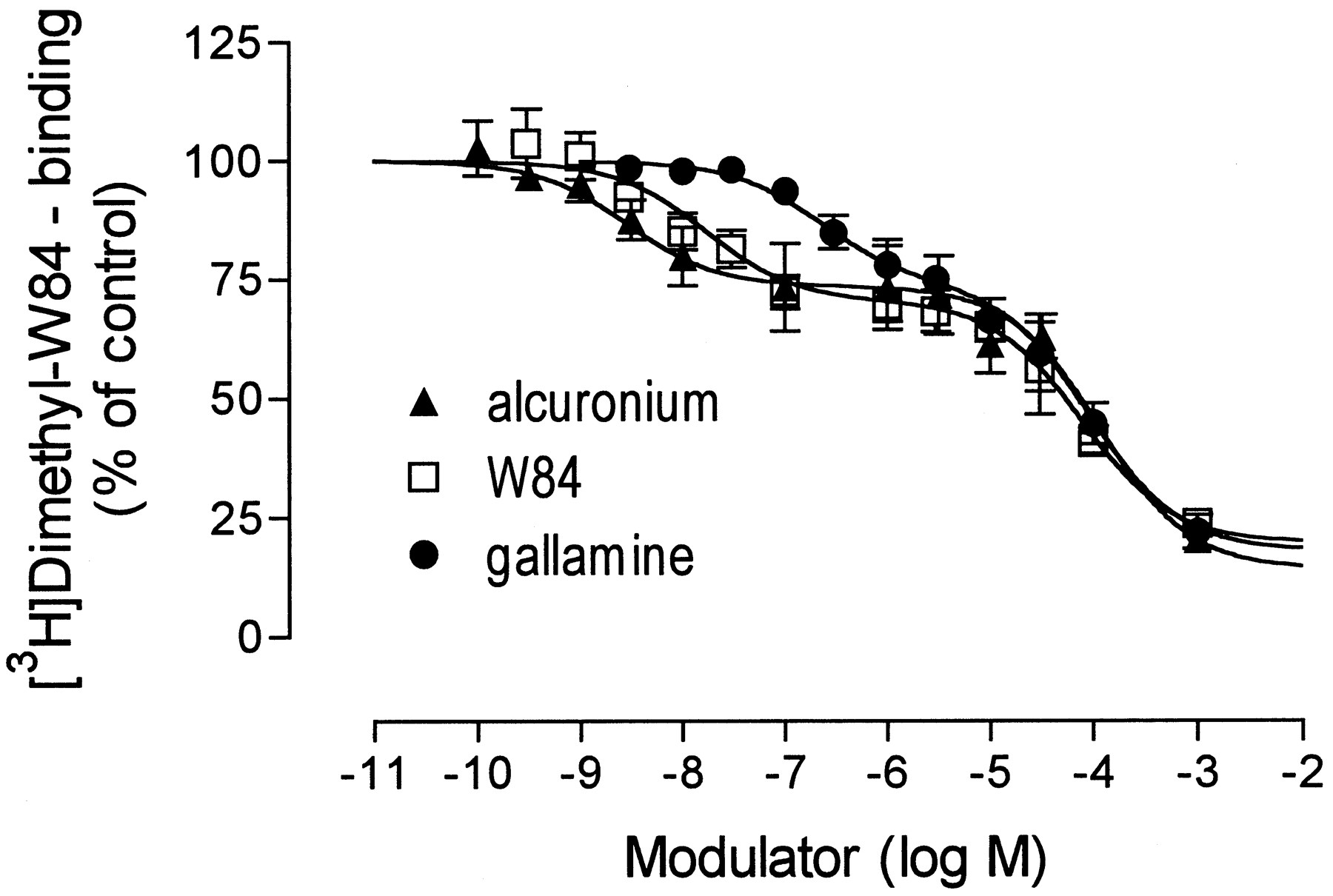
It was tested whether the allosteric model compounds alcuronium, W84, and gallamine compete with the binding of 0.3 nm[3H]dimethyl-W84. As shown in Fig.5, all compounds reduced [3H]dimethyl-W84 binding in a biphasic manner. The curves level off at very similar minimum values. In addition, the intermediate plateaus of the curves are found at the same level of ∼33% of saturable [3H]dimethyl-W84 binding. pKi values were derived from the IC50 values according to Cheng-Prusoff (1973)using the pKD value of 8.74 for [3H]dimethyl-W84 binding to the NMS-occupied receptor. The low affinity components of the inhibition curves are almost identical (pKi , low values in Table 1). In contrast, with regard to the high affinity inhibition of [3H]dimethyl-W84 binding, the compounds differ considerably. The pKi , highvalues are listed in Table 1.

Inhibition of the total binding of [3H]dimethyl-W84 (0.3 nm) by the allosteric modulators alcuronium, W84, and gallamine. Binding of [3H]dimethyl-W84 in the absence of inhibitor was set 100%. Given are mean ± standard error values of four or five experiments with triplicate determinations. Error bars are not shown when they do not exceed the symbols. The curve fit is based on a two-site model.
Discussion
Three features of the allosteric action of dimethyl-W84 were pivotal for choosing this compound for radiolabeling. First, with a half-maximally effective concentration for the inhibition of [3H]NMS dissociation of an EC50 value of 3 nm, dimethyl-W84 has a top position among various modulators in this respect. Previously, we determined in a comparative study the potency of a wide spectrum of compounds to interact allosterically with NMS-occupied receptors (Tränkle et al., 1996). In that study, alcuronium was found to be most potent with an EC50 value of 4 nm. We concluded that dimethyl-W84 should belong to the compounds with highest affinity to NMS-occupied receptors. Second, dimethyl-W84 is rather easily available by chemical synthesis compared with alcuronium. Third, the results obtained with obidoxime as an “allosteric antagonist” against dimethyl-W84 (pKb = 4.71) suggest that dimethyl-W84 interacts with the postulated common allosteric site of M2 receptors (Ellis and Seidenberg, 1992); recently, it was shown (Tränkle and Mohr, 1997) that obidoxime antagonized the effects of the allosteric model compounds gallamine, W84, and alcuronium on [3H]NMS dissociation with very similar potency (pKb = 4.70, 4.61, and 4.30, respectively). In line with the concept of a common site are the findings of Proška and Tuc̆ek (1995), who could predict the effect of combinations of alcuronium and gallamine on the equilibrium binding of [3H]NMS in cardiac membranes by applying a model of a competitive interaction.Lanzafame et al. (1997) provided evidence for a competition among alcuronium, gallamine, and C7/3′-phth (the heptane homologue of W84) in contracting guinea pig atria. Thus, under various experimental conditions, the concept of a common allosteric site on M2 receptors has been found to be appropriate. The sensitivity of the allosteric action of dimethyl-W84 toward the “common-site antagonist” obidoxime was essential for the decision to develop the tritiated compound as a potential common-site radioligand.
In the [3H]dimethyl-W84 binding experiments, we applied 1 μm NMS to occupy the orthosteric site of the M2 receptor protein for the reasons mentioned above. There is a negative cooperativity between dimethyl-W84 and NMS (see Fig. 2), which, however, is rather weak. Nevertheless, the question arises of whether high concentrations of dimethyl-W84 might reduce occupation of the orthosteric site by NMS to a relevant extent. In the absence of dimethyl-W84, 1 μm NMS (pKD = 9.56) leads to a fractional receptor occupancy of 0.9997. Dimethyl-W84 was applied in the [3H]dimethyl-W84 binding experiments at a maximum concentration of 10−3 m (see Fig. 3); from the characteristics of the inhibition of [3H]NMS equilibrium binding by dimethyl-W84 (see Fig. 2; pKD = 8.32, α = 3.2), it can be derived using the cooperativity model that the fractional receptor occupancy by NMS in the presence of the highest applied concentration of dimethyl-W84 (10−3 m) amounts to 0.9991. This implies that occupancy of the orthosteric site by NMS is maintained even at high concentrations of dimethyl-W84. Thus, for the analysis of the [3H]dimethyl-W84 binding data, it seems appropriate to consider the NMS/receptor complex as the binding site of the allosteric agent.
The high affinity component of saturable [3H]dimethyl-W84 binding probably represents occupation of the allosteric site. This conclusion is derived from the following findings. The pKD , high value of 8.74 of high affinity binding of [3H]dimethyl-W84 to NMS-occupied receptors matches the pEC50,diss value of 8.51 for the allosteric effect of dimethyl-W84 on the dissociation of [3H]NMS/receptor complexes. The shape of the dimethyl-W84 allosteric effect curve (nH = 1.1, not significantly different from unity) corresponds with the shape of the [3H]dimethyl-W84 binding curve (nH = 1.0 by definition of the two-site model). Furthermore, the number of high affinity binding sites for [3H]dimethyl-W84 (B max = 68 ± 11 fmol/mg protein) is not significantly different from the number of [3H]NMS-binding sites (B max = 79 ± 21 fmol/mg protein), which supports the notion that high affinity [3H]dimethyl-W84 binding takes place at muscarinic receptors. Heat treatment, which abolishes specific binding of [3H]NMS, also eliminates the high affinity component of [3H]dimethyl-W84 binding. The other putative ligands for the allosteric site (i.e., alcuronium, W84, and gallamine) displace [3H]dimethyl-W84 binding at concentrations that match the concentrations for the allosteric effect of these agents on NMS dissociation. There is an excellent linear correlation between the pEC50,diss values and the binding constants pK of the four applied modulators, pEC50,diss = 0.87·pK + 0.90,r 2 = 1.00. Thus, high affinity [3H]dimethyl-W84 binding apparently occurs at the site that mediates the allosteric effect of the modulators on NMS/receptor complexes.
The low affinity component of [3H]dimethyl-W84 binding probably represents nonspecific, yet saturable, binding of the radioligand to the cardiac membranes. Because there is evidence that modulators have fast binding kinetics at their site of action (Tränkle and Mohr, 1997), membranes were separated in the current experiments by centrifugation instead of filtration. Thus, there was no intensive washing of the membranes to reduce nonspecific radioligand binding; it is common in filtration assays. The low affinity component has a very large capacity, suggesting that this binding is rather nonselective. The test compounds gallamine, alcuronium, and W84 inhibit [3H]dimethyl-W84 binding in the low affinity component, yet the binding affinity is low (Ki =∼100 μm) and hardly differs between the test compounds. Using phosphatidylserine monolayers as an interphase model and the displacement of45Ca2+ as an indicator for cationic drug binding (Girke et al., 1989), we found that dimethyl-W84, gallamine, alcuronium, and W84 bind with similar affinity to this negatively charged phospholipid (Mieskes A, unpublished observations). In 5 mmNa,K,Pi buffer at 23°, as applied in the current study, the half-maximally effective concentrations ranged between 3 and 30 μm. Remarkably, the affinity of NMS to the phospholipid is ∼1000-fold lower (Mieskes A, unpublished observations). The comparably high propensity of dimethyl-W84 to bind to a negatively charged interphase offers an explanation of why the nonspecific binding of [3H]dimethyl-W84 to the heart homogenate is considerably higher than the nonspecific binding of [3H]NMS.
Compared with the nonspecific binding, the specific binding of [3H]dimethyl-W84 at the allosteric site of M2 receptors has a ∼20,000-fold higher affinity (pKD , high = 8.74 versus pKD , low = 4.43). This large difference in affinity demonstrates that occupation of the allosteric site is a preferential event and not an aspect of a nonspecific interaction with membrane interphases. This finding will encourage to pursue in the development of allosteric modulators as potential therapeutics. It may be assumed that this preference does not vanish under physiological incubation conditions because higher cation concentrations, including divalent cations, not only reduce the potency of allosteric modulators but also can diminish the interaction of positively charged drugs with phospholipid interphases (Schlieper and Steiner, 1983; Ohki, 1984).
[3H]Dimethyl-W84 exemplifies that binding of allosteric agents to the common site can be measured directly in genuine M2 receptors of cardiac tissue. This opens a new methodological perspective in the study of allosteric phenomena. For instance, the effect on radioligand binding can be investigated for drugs that have been supposed to interact with the allosteric site such as the muscarinic agonist oxotremorine (Jakubı́k et al., 1997) and for allosteric agents assumed not to attach to the common site (Tränkle and Mohr, 1997).
Furthermore, the interaction of allosteric compounds with liganded and with free M2 receptors may be compared as an alternative approach to determine cooperativity factors. Preliminary [3H]dimethyl-W84 experiments carried out in the absence of unlabeled NMS gave a pKD value of 8.89 ± 0.18 (mean ± standard error, four experiments) for radioalloster binding at free M2receptors. This value matches the pEC50,ass value of 8.81 for inhibition of [3H]NMS association induced by dimethyl-W84 binding to free M2receptors. The ratio between the KD values of [3H]dimethyl-W84 binding to NMS-occupied receptors (pKD = 8.74,KD = 1.82 nm) and to free receptors (pKD = 8.89,KD = 1.26 nm), respectively, amounts to 1.4. This ratio favorably corresponds with the shift factor of 1.5 between the curves (Fig. 2) for the effects of dimethyl-W84 on the association of [3H]NMS and on its dissociation (pEC50,diss = 8.51). Furthermore, the [3H]dimethyl-W84 affinity ratio is close to the cooperativity factor α value of 3.2 (Fig. 2) as a measure derived from [3H]NMS equilibrium experiments for the difference in affinity of dimethyl-W84 at free and NMS-occupied receptors. Yet, for a detailed study of cooperativity phenomena involving such small affinity shifts, it is desirable to refine the binding assay in that nonspecific radioligand binding is reduced, for instance, by changing the ionic composition of the assay or by using cloned receptors at elevated concentrations.
In conclusion, a radiolabeled allosteric modulator of muscarinic receptors with high allosteric potency was developed. Evidence is presented for the first time that radioligand-binding measurements at the allosteric site of ligand-occupied muscarinic M2 receptors can be carried out. This finding opens a new methodological perspective in the study of allosteric interactions at muscarinic receptors.
Acknowledgments
We thank Iris Witten and Frauke Mörschel for their expert technical assistance. The active support of the project by Prof. Dr. K. H. Jakobs and Dr. A. Moritz (University of Essen, Essen, Germany) is gratefully indicated.
Footnotes
- Received December 8, 1997.
- Accepted March 12, 1998.
-
Send reprint requests to: Klaus Mohr, Department of Pharmacology & Toxicology, Institute of Pharmacy, University of Bonn, An der Immenburg 4, D-53121 Bonn, Germany. E-mail:k.mohr{at}uni-bonn.de
-
This work was supported by the Deutsche Forschungsgemeinschaft und the Fonds der Chemischen Industrie, Germany. H.M.B.C. acknowledges a grant given by the Katholischer Akademischer Austauschdienst.
Abbreviations
- NMS
- N-methylscopolamine
- dimethyl-W84
- N,N′-bis[3-(1,3-dihydro-1,3-dioxo-4-methyl-2H-isoindol-2-yl)propyl]-N,N,N′,N′-tetramethyl-1,6-hexanediaminium diiodide
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}