


Abstract
Three mRNA variants are generated from the mouse κ-opioid receptor (KOR) gene. The expression patterns of these KOR mRNA variants in adult animal tissues and during developmental stages are examined. Furthermore, the biological significance of generating these variants is demonstrated with respect to two post-transcriptional mechanisms, i.e., mRNA stability and translation efficiency. Variants A and B are both transcribed from promoter 1 of the KOR gene and expressed from early developmental stages through adult life. Although their sequences differ only at a 30-nucleotide insertion for variant B, these two variants are distinct with regard to their expression patterns, mRNA stability, and translation efficiency. Variant A is expressed ubiquitously in all the tissues examined and has a longert 1/2 (12 h), whereas variant B is more specific to the central nervous system both pre- and postnatally and has a t 1/2 of ∼8 h. Variant C is transcribed from promoter 2 of the KOR gene and is most specifically expressed, being detected only in the brain stem, spinal cord, and thalamic/hypothalamic areas of postnatal animals. With regard to protein translation, variants B and C are significantly more efficient than variant A. This study provides the evidence for multiple levels of KOR regulation. The biological implication of the generation of KOR mRNA variants is discussed.
Opiates exert a wide spectrum of pharmacologic effects, mediated by a family of G protein-coupled transmembrane receptors called opioid receptors. Three opioid receptor types, μ, δ, and κ, are present (Goldstein and Naidu, 1989; Loh and Smith, 1990). The genes of the three opioid receptors have been cloned, and their expression patterns in animals have been examined with in situ hybridization, immunohistochemistry, and ligand binding assays (Rius et al., 1991; Elde et al., 1995;Kieffer, 1995; Knapp et al., 1995; Mansour et al., 1995; Zhu et al., 1998). The physiologic roles have only begun to be uncovered from studying genetically altered animal models (Mattes et al., 1996;Sora et al., 1997; Tian et al., 1997; Loh et al., 1998; Simonin et al., 1998; Schuller et al., 1999).
Studies using antibodies, nucleic acid probes, and radioisotope-labeled ligands reveal different expression patterns of the three opioid receptors in the brain and spinal areas that are associated with pain sensation and behavior. Our recent studies in developing animals also show distinct patterns of expression of the three opioid receptor genes in embryonic stages, suggesting potential functions of opioid receptors in early animal development (Chen et al., 1999). Particularly, the κ-opioid receptor (KOR) appears very early during animal development [embryonic day (E) 9.5]. Consistent with the observation that KOR is expressed in developing embryos before the formation of the nervous system, the KOR gene is found to be active in the stem cell populations of an embryonal carcinoma cell line P19 (Chen et al., 1999).
The mouse KOR gene has been isolated in several laboratories, and its genomic structure has been determined (Liu et al., 1995). In the P19 cell line model, we have demonstrated the biologic activities of dual promoters of the mouse KOR gene (Lu et al., 1997). These two promoters can potentially generate transcript variants that either contain or lack exon 1 sequence (Lu et al., 1997). In addition, our own as well as other studies (Belkowske et al., 1995; Lu et al., 1997), have demonstrated the presence of another variant using an alternative, splicing acceptor site at intron 1. As a result of this alternative splicing, this mRNA variant encodes an additional 30 nucleotides between exons 1 and 2 (Lu et al., 1997). Despite the demonstration of these KOR mRNA variants in mouse brain and cell lines, it remains elusive as to whether these KOR mRNA variants represent biologically functional KOR mRNAs and whether they are expressed differentially in animal tissues. Furthermore, the biologic significance of generating KOR variants is unknown.
To address these questions, we set up experiments, first to examine the expression of these KOR mRNA variants in adult animals as well as during development. Second, we addressed the biological significance of these variants in two post-transcriptional events, mRNA stability and translation efficiency. The results of this study provide the evidence, for the first time, of differential expression of KOR mRNA variants as well as post-transcriptional regulation of KOR expression.
Materials and Methods
Analyses of KOR mRNA Variant Expression.
P19 embryonal carcinoma cells were maintained as described (Wei and Chang, 1996). RNA was isolated from normal adult (body weight 25–30 g) CD1 mouse tissues and P19 cells using the method of Charron and Drouin (1986). To differentiate the expression of KOR mRNA variants in small tissue samples such as dissected brain areas, conventional mRNA detection methods such as Northern blot and RNase protection methods are not appropriate because of the limit in sensitivity and sample size. Another method, in situ hybridization, was not chosen, because the sequence that allowed these variants to be detected differentially are too short (i.e., 30 nucleotides long). Therefore, reverse transcription polymerase chain reaction (RT-PCR) was used. RT was conducted using SuperScript II RT (Life Technologies, Grand Island, NY) and primed with oligo(dT) primers on 2 μg of RNA samples in a total volume of 20 μl. To differentiate between the two variants generated from the first promoter with alternative splicing that occurred at intron 1, the variant-specific 5′ primers were designed according to specific exon 1/exon 2 junction sequences (Lu et al., 1997). These two KOR mRNA variants were called A and B. The 5′ primer for variant A is 5′-ATCAGCGATCTGGAGCT-3′ and that for variant B (containing an insertion of 30 nucleotides) is 5′-TCAGCGATCTGGAGCCCC-3′ (underlined residues showing the splicing junctions of each species of mRNA). The sequence of the 5′ primer for the third variant (C), transcribed from the second promoter located in intron 1, is 5′-ACAGGCAAAGTTTGTC-3′. A common 3′ primer, 5′-GCAAGGAGCATTCAATGAC-3′, was designed according to an exon 4 sequence that is present in all the three variants. Thus, KOR mRNA variants A, B, and C were amplified as 730 base pair (bp), 760-bp, and 800-bp fragments, respectively. To establish RT-PCR, the linear ranges of amplification cycles and the PCR inputs in relation to PCR products were first determined (seeResults). Appropriate dilutions of RT products were each amplified in a 20 μl PCR using commercially provided buffer with a reaction cycle of 94°C for 45 s, 55°C for 45 s, and 72°C for 1 min, for a total of 30 cycles. A pair of actin-specific primers (5′-TGGCCTTAGGGTGCAGGG-3′; 5′-GTGGGCCGCTCTAGGCACCA-3′) (Rappollee et al., 1989) were included for 23 cycles in each reaction for internal controls. For all the RT-PCR experiments, a negative control without RT was included. After PCR, 5 μl of each sample was analyzed on Southern blots using [α-32P]dCTP-labeled probes prepared from the mouse KOR cDNA spanning exon I to exon IV and from actin cDNA. Southern blot analyses were conducted according to the established protocols (Maniatis et al., 1990).
To quantify PCR products, the hybridizing bands were quantified in a phosphoimager by using Image Quant software (Molecular Dynamics, Sunnyvale, CA). The KOR-specific signal was normalized to the actin-specific signal in each reaction to obtain a value representing the specific KOR variant expression. The expression of each variant in all the samples (see Figs. 3 and 4) was represented as the relative level of expression by arbitrarily setting the expressed level of one variant at a specific stage as the value of 1. The mean and S.D. values were obtained from at least three experiments.

Differential expression of KOR mRNA variants during prenatal development. A, the products of RT-PCR were examined on Southern blot as described in the text. Abbreviations are B, brain; T, trunk. B, a quantitative presentation of KOR variant A expression. The variant A-specific signal was normalized to actin-specific signal to obtain a value representing specific variant A expression. The normalized expression level of variant A in E9.5 brain was set arbitrarily as the value of 1 to determine the relative expression level of variant A at each developmental time point. C, a quantitative presentation of KOR variant B expression. Quantitation was conducted as described for panel B. Three independent experiments were conducted to obtain the mean and S.D. values.
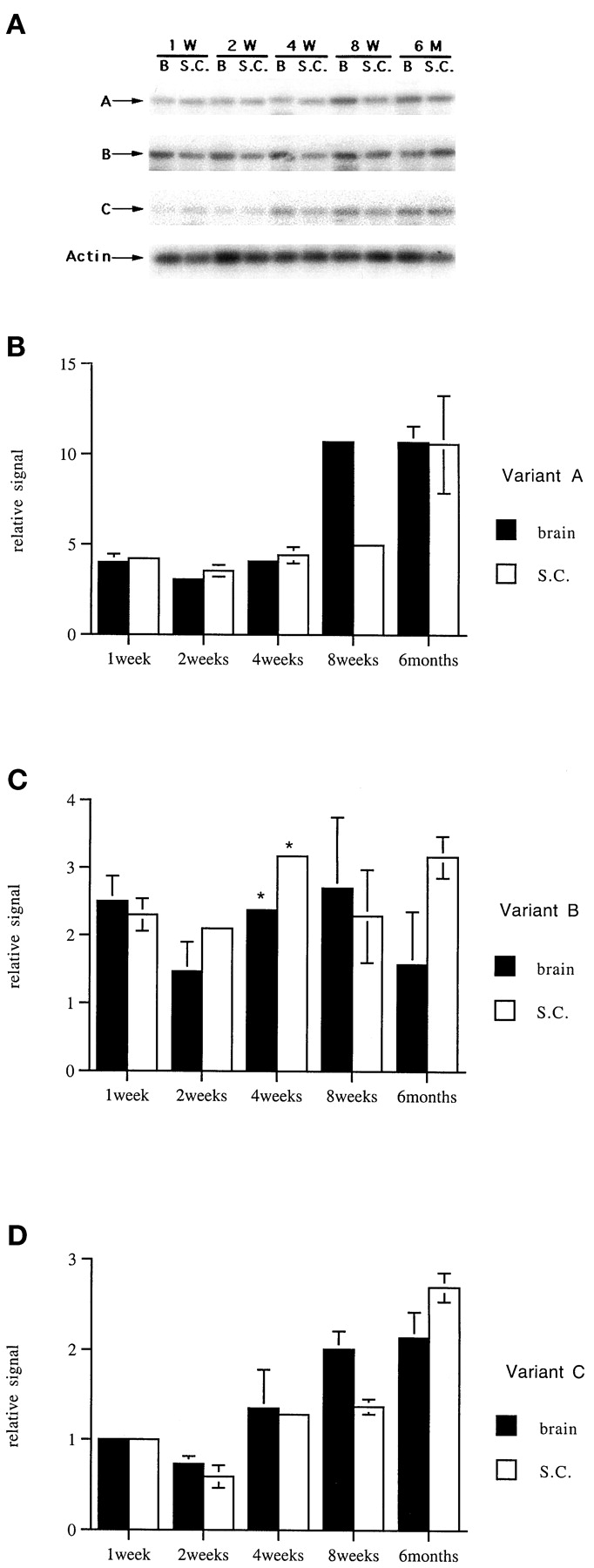
Differential expression of three KOR mRNA variants during postnatal development. A, Southern blot analyses of RT-PCR products. RT was diluted 20-fold to amplify variant A and 5-fold to amplify variants B and C. The developmental stages of brain (B) and spinal cord (SC) samples are indicated. B, a quantitative presentation of KOR variant A expression during postnatal development. Quantitation was conducted by using variant B expression (panel C) in postnatal 1-week-old brain as an arbitrary value of 1. C, a quantitative presentation of KOR variant B expression during postnatal development as described for panel B. D, a quantitative presentation of KOR variant C expression as described for panel B. Three independent experiments were conducted to obtain the mean and S.D. values.
To determine the t 1/2 of mRNA, experiments were conducted in P19 stem cells that expressed endogenous KOR variants A and B. Actinomycin D was added at a concentration of 2 μg/ml. At different time points, RNA was collected and subjected to RT-PCR analyses and phosphoimager quantification as described previously.
In Vitro Transcription/Translation (TNT).
The cDNA of each KOR mRNA variant was cloned into the PvuII site of pSP73 (Promega, Madison, WI) following the T7 promoter. A control vector was generated by fusing the 5′-untranslated sequence of luciferase (Luc) reporter from pGL3 (Promega) to the upstream of the ATG codon of the KOR coding region, which also was cloned into pSP73 vector at the samePvuII site. TNT reactions were conducted using the TNT-coupled reticulocyte lysate reaction system (Promega) at 30°C for 60 min. In each reaction, 0.5 μg of specific KOR cDNA vector and 50 ng of an internal control vector (a cytochrome enzyme CYP26 expression vector also cloned in pSP73) (Haque and Anreola, 1998) were used as the starting material, and [35S]methionine was added during translation. The condition was established to synthesize KOR and CYP26 proteins in a linear range. The transcription efficiency was monitored by examining RNA products in Northern blot analyses. The translated products from the same amount of KOR mRNA variants were analyzed on 12% SDS polyacrylamide gels and quantified using a phosphoimager as described previously. The signal of the KOR protein was normalized to the internal control CYP signal to obtain a value representing specific KOR protein expression in each TNT reaction.
Transfection Experiments to Determine Reporter Activities Representing the Translation Efficiency of 5′-Untranslated Region (5′-UTR) of KOR Variants.
The 5′-UTR of KOR variants was each fused to the Luc coding sequence of pGL3 (Promega), followed by the addition of a thymidine kinase promoter (Wagner et al., 1981) to its upstream and an SV40 poly(A) to its downstream, generating reporters for the different 5′-UTRs of KOR mRNA variants. COS-1 cells were maintained and transfected with each Luc reporter, together with an internal control β-galactosidase (lacZ) reporter, as described (Lu et al., 1997). Thirty six hours after transfection, Luc activity was determined and normalized to control and lacZ activity to obtain the relative Luc unit for each transfected culture.
Results
Differential Expression of KOR mRNA Variants in Animal Tissues.
Previously, we have demonstrated three KOR mRNA variants by using RNase protection assay and sequence analyses (Lu et al., 1997). Variant A is transcribed from promoter 1 and spliced at the −14 position (14 bp upstream from the translation initiation codon ATG). Variant B is also transcribed from promoter 1, but is spliced at the −44 position, resulting in a mRNA species with a 30-nucleotide insertion. Variant C is transcribed from promoter 2 located in intron 1 and is initiated at the −93 position (79 bp from intron 1 and 14 bp from exon 2). To detect the expression of these mRNA species in different tissues, brain regions, and mouse embryo parts, we have designed a more sensitive RT-PCR assay that allows small amounts of tissue samples and low levels of expression to be examined. Using this RT-PCR approach, all three KOR mRNA variants can be differentiated as described in Materials and Methods. By using different 5′ primers that span the alternative splicing junctions and a common 3′ primer derived from an exon 4 sequence, variants A and B can be differentially detected. By using a 5′ primer specific to an intron 1 sequence transcribed from promoter 2 and the same 3′ primer, variant C can be detected specifically and differentiated from the other two. Oligo(dT) primers were used in RT and an exon 4-specific, 3′ primer was used in PCR; therefore, only mature mRNA with polyadenylation and proper splicing can be amplified in the expected sizes of 730, 760, and 800 bp for variants A, B, and C, respectively. For qualitative and quantitative control of each reaction, actin-specific primers were included for each sample, and the cycle numbers and PCR inputs for a linear amplification were determined. Figure1A shows specific primers and expected fragments in these RT-PCRs. Figure 1B shows the results of experiments that determine amplification cycles in a linear range for each KOR variant and for actin. Figure 1C shows the results of experiments that demonstrate amplified products of each variant in relation to the amounts of PCR input. Based on these results, the following experiments were conducted for a total of 30 cycles for KOR variants and 23 cycles for actin. One tenth of each RT product was used for PCR. These conditions allowed each mRNA species to be amplified in a linear range.
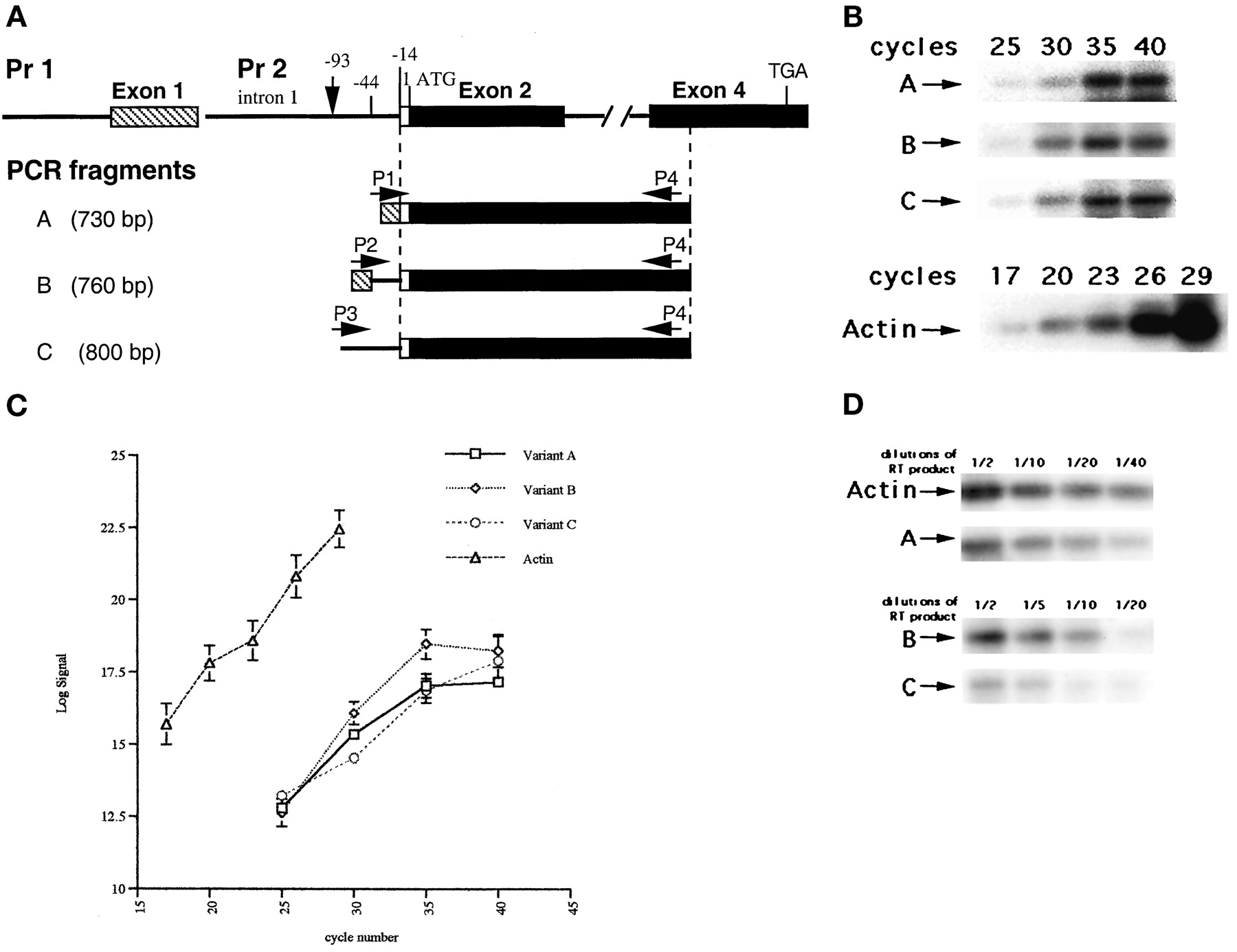
The RT-PCR strategy to differentially detect the expression of KOR mRNA variants. A, primers and PCR products. Two KOR mRNA variants (A and B) are initiated from promoter 1, one spliced at the −14 position relative to the initiation codon ATG in exon 2 (variant A), the second spliced at the −44 position (variant B). The third variant (C) is transcribed from promoter 2 located in intron 1, at the −93 position. A 3′ primer (P4) is designed according to exon 4 sequence, which is common to all three variants. Primer 1 (P1) spans the specific splicing junction of exons 1/2 at the −14 position; thus only variant A can be amplified in the size of 730 bp. Primer 2 (P2) spans the specific splicing junction of exons 1/2 at the −44 position; thus only variant B can be amplified in the size of 760 bp. Primer 3 (P3) is specific to the variant transcribed from promoter 2; thus only variant C can be amplified in the size of 800 bp. Dashed lines indicate the transcribed region common to all three variants. B, determination of amplification cycles in a linear range. One tenth of RT products from adult brain was used in PCRs for various cycles and analyzed on Southern blots. C, quantification of PCR yields versus amplification cycles. Log values of PCR products were plotted against cycle numbers used to amplify variants A, B, and C, and for actin. Two independent experiments were conducted to obtain the average readings and S.D. values. D, demonstration of PCR yields versus inputs. Half to 1/40 dilutions of RT products from adult brain were amplified for 30 cycles for KOR variants, and 23 cycles for actin, and analyzed on Southern blots.
To examine how these KOR variants are expressed differentially in adult tissues, RT-PCR was conducted on RNA samples prepared from dissected adult mouse tissues. Figure 2 shows a representative Southern blot analysis from two independent experiments, demonstrating differential expression of KOR mRNA variants in mouse tissues and brain regions. Variant A is expressed in all the tissues examined, including lung (L), heart (H), stomach (S), ovary (O), cortex (Co), thalamus/hypothalamus (T), cerebellum (Ce), brain stem (BS), and spinal cord (SC). Interestingly, variants B and C are expressed only in limited tissues. Variant B is clearly detected in most brain areas (except in cerebellum, where a very low level of expression is detected) but not other tissues, whereas variant C is detected only in the brain stem, spinal cord, and thalamic/hypothalamic regions. Therefore, variant A seems to be expressed ubiquitously in adult animal tissues and variant B is expressed in most parts of the central nervous system (CNS), whereas variant C is most specific, being restricted to the brain stem, spinal cord, and thalamic/hypothalamic regions.

Differential expression of three KOR mRNA variants in adult tissues and dissected brain regions. The products of RT-PCR were analyzed on agarose gels, followed by Southern blot hybridization to KOR- (top three panels) or actin- (bottom) specific probes to examine the expression patterns of KOR mRNA variants A, B, and C. Samples in each lane are lung (L), stomach (S), heart (H), ovary (O), cortex (Co), thalamus/hypothalamus (T), brain stem (BS), cerebellum (Ce), and spinal cord (SC).
Expression Profile of Each KOR mRNA Variant during Developmental Stages
The total KOR mRNA expression has been detected in early embryos (E9.5) and in undifferentiated P19 cells in our previous studies (Chen et al., 1999). To gain insight into how these KOR mRNA variants are expressed differentially during prenatal stages, RT-PCR was conducted using RNA prepared from mouse embryos. Older embryos were dissected into two portions, the brain (B) and the trunk (T). Figure3A illustrates a representative Southern blot analysis from three independent experiments, showing KOR variants and actin expression in embryonic stages. Figure 3, B and C, shows the results of phosphoimager analysis of variant A and B expression, respectively, normalized to actin signal. The relative expression level of each sample is determined by arbitrarily setting the level of variant A expression in E9.5 embryos as the value of 1. Although variants A and B both are expressed in E9.5 embryos and the brain (B) and the trunk (T) of E12.5 embryos, their expression patterns begin to divert in older embryos. Variant A is detected in both brain and trunk of all embryos examined, but variant B is restricted to the brain of older embryos. Interestingly, variant C, which is initiated from the second promoter, is not detected in any of the embryo samples.
The postnatal profiles of KOR variant expression also was examined in RT-PCR experiments. Because KOR is expressed primarily in the brain and spinal cord in adults (Fig. 2), only the whole brain and spinal cord samples were examined in these experiments. Furthermore, because variant A is expressed at a much higher level compared with variants B and C (Fig. 2), the RT products were diluted 5× for amplifying variants B and C and 20× for amplifying variant A to obtain PCR products that can be quantified reliably. The data then were corrected for the dilution factors. The expression level of variant B in 1-week-old postnatal brain was arbitrarily set as the value of 1 to determine the relative expression levels of all the variants in all the samples. Three independent experiments were conducted, and one representative pattern was shown in Fig.4A. It appears that all three KOR variants are expressed in both brain and spinal cord at all ages, starting from postnatal week 1 to 6 months of age. Figure 4, B–D, shows the results of phosphoimager analyses of relative expression levels of variants A, B, and C, respectively, during postnatal development. From these analyses, it is concluded that the expression of variants A and C is slightly elevated (for ∼2-fold) in older animals; whereas the expression of variant B is relatively constant. Additionally, variant A is expressed at the highest level in all the developmental stages; variant C is expressed at the lowest level in young animals (1–2 weeks old) and gradually levels with variant B in mature animals.
Therefore, KOR mRNA variants are expressed differentially during both pre- and postnatal development. Variant C is detected only postnatally, specifically in the brain stem, spinal cord, and thalamic/hypothalamic areas, whereas variants A and B are detected both pre- and postnatally. In mid- to late gestation stages of embryos (older than E12.5), when the CNS is being formed, variant B is detected only in the brain (Fig.3), consistent with the observation that this variant is detected only in the CNS of adult animals (Fig. 2). Variant A appears to be the most ubiquitous and highly expressed species. It can be detected in all embryo samples and all tissues of adult animals.
mRNA stability of KOR Variants.
To gain insight into the biologic significance of generating KOR mRNA variants, we first examined mRNA stability of the two alternatively spliced variants A and B, which are expressed in cultured cells and are assessable. The undifferentiated P19 cells expressed both variants constitutively. Actinomycin D was added to the culture to stop RNA transcription. At different time points after actinomycin D treatment, RNA was collected from the cultures and the relative levels of variants A and B were determined by RT-PCR. Figure 5A shows a representative Southern blot analysis of these experiments, and Fig. 5B shows the determination of the t 1/2 of variants A and B from two experiments. At later time points, the message level was too low to be quantified; nonetheless, data could be quantified reliably within 20 h. From these graphs, clearly the decay of variant B is faster than that of variant A, and it is estimated that variant A has a t 1/2 of ∼12 h, whereas variant B has a t 1/2 of ∼8 h. Therefore, it is concluded that KOR variant A is more stable than variant B in P19 cells.

The stability of KOR mRNA variants A and B. A, Southern blot analyses of variants A and B at different time points after the addition of actinomycin D. B, determination of thet 1/2 of each variant. The highest value (at time 0) of each variant expression was set arbitrarily at the value of 1. The relative values of each variant at different time points were plotted against the time after actinomycin D treatment. Thet 1/2 for variant A is ∼12 h, and thet 1/2 for variant B is ∼8 h. Two independent experiments were conducted to obtain the mean and S.D. values.
Translation Efficiency of KOR Variants.
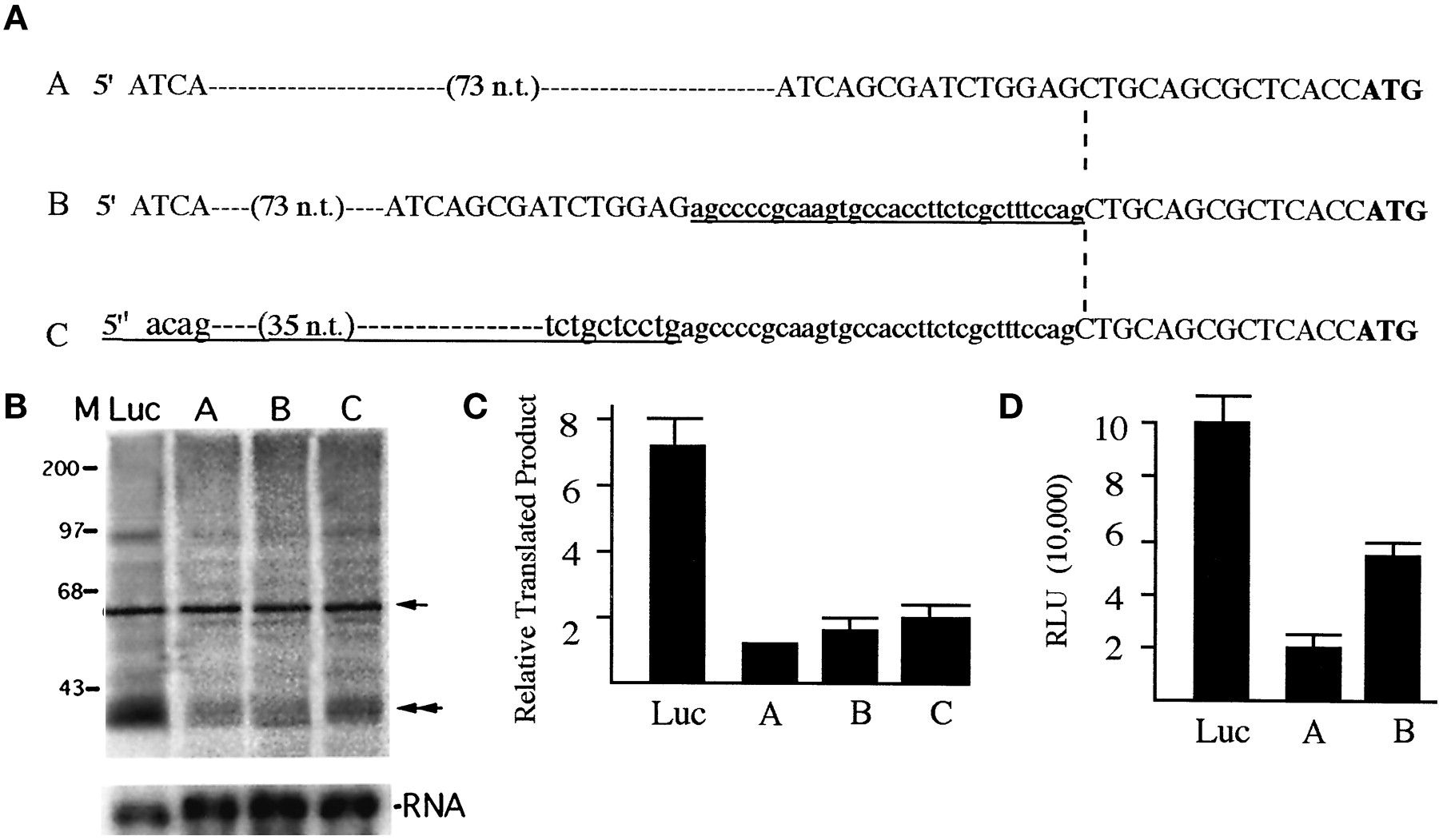
The three KOR mRNA variants share the identical protein coding region, yet they differ dramatically within a short range in their 5′-UTR, starting at the −14 position (from the initiating codon ATG). To compare their translation efficiency, in vitro TNT and reporter analyses were conducted. We first generated KOR cDNAs encompassing both the common and unique sequences of the 5′-UTR of the three mRNA variants. The cloned cDNAs of variants A and B share the same 5′-UTR derived from exon 1 (−477 position relative to the ATG codon according to the genomic sequence, which encodes a 92-bp sequence from exon 1) and a common 14-bp sequence immediately upstream of ATG codon located in exon 2. The only difference between variants A and B is a 30-bp insertion at the −14 position of variant B. The construct of variant C starts at the −93 position relative to the ATG codon, which contains a unique 79-bp sequence derived from intron 2 (following promoter 2) and the common 14-bp sequence upstream of ATG located in the 5′ end of exon 2. Therefore, variant B is identical with A, except for a 30-nucleotide insertion at the −14 position and variant C is identical with B except for a unique 49-bp sequence upstream of the alternative splicing site within intron 1. All the sequences have been confirmed by DNA sequencing. The three cDNAs were each inserted into the same site of pSP73 in vitro expression vector following the T7 promoter. Therefore, transcription of the three cDNAs is under the control of the identical T7 promoter. For a comparison on the efficiency of the Kozak sequence, a control vector was generated by fusing the 5′-UTR of a commercially available Luc reporter, in frame, upstream of the ATG codon of the KOR coding region. In addition, for all the reactions, two control steps were taken, one to include an internal translation control using a well-characterized Cyp26 expression vector (Haque and Anreola, 1998) and the second to monitor the quantity and quality of RNA produced in all the reactions by performing Northern blot analyses on RNAs used in each in vitro translation reaction.
Figure 6A shows sequence comparison of the 5′-UTR of three KOR variants, and Fig. 6B shows a representative autoradiograph of KOR protein expression (top) using various 5′-UTRs of the KOR as well as that of pGL3. The quality and quantity of KOR RNA variants made in each reaction are shown at the bottom. Figure 6C shows the results of phosphoimager analyses of KOR protein products normalized to the internal control Cyp26 products from four independent experiments. All three KOR mRNA variants and the control vector generate the same KOR protein product with a molecular weight of ∼40 kD. Interestingly, all three KOR 5′-UTRs are much less efficient than pGL3, suggesting that the natural Kozak sequence of all three KOR RNA variants is significantly less efficient compared with the optimized Kozak used in the commercial vector. Among the three variants, variants B and C are significantly more efficient (∼2-fold of variant A) in terms of in vitro translation efficiency.

The translation efficiency of KOR mRNA variants. A, DNA sequences of the 5′-UTRs of three constructs for expressing three variants. Small letters show sequence identified as the first intron in our previous study (Lu et al., 1997). Underlined sequences indicate differences between variants B and A, and between variants C and B. Dashed line points at the beginning of exon 2 followed by the translation initiation codon ATG (bold), which are identical in all three variants. B, TNT reactions (top panel) were conducted using the T7 expression vectors of three KOR variants (lanes A, B, and C) and the control using the 5′-untranslated sequence of Luc reporter (lane Luc). An internal control (Cyp26) was included in each reaction, and the products are indicated with an arrow on the right. A double arrowhead indicates the translated KOR protein. The transcribed RNA was monitored on a Northern blot, as shown on the bottom panel. C, a quantitative presentation of KOR protein expression from mRNA variants A, B, and C. The KOR-specific signal was normalized to the Cyp26-specific signal to determine the yield of specific KOR protein from each reaction. The lowest value (generated from the template using the 5′-UTR of variant A) was set as the value of 1 to determine the relative yield of KOR protein produced from variants B and C and the pGL3. Four independent experiments were conducted to obtain the mean and S.D. values. D, relative reporter activities of 5′-UTR of each KOR variant and pGL3. The Luc reporters using the 5′-UTR of KOR variants were determined in transient transfection experiments and represented as the relative Luc unit by normalizing to internal control lacZ activities, as described in the text. Three independent experiments were conducted to obtain the mean and the S.D. values.
The translation efficiency of each 5′-UTR was also examined in reporter assays. The 5′-UTR of each variant was used to generate reporters using the same promoter (thymidine kinase), coding sequence (Luc), and poly(A) (SV40) as described in Materials and Methods. These reporter activities were determined in three transient transfection experiments as shown in Fig. 6D. Consistent with the results of TNT reactions (Fig. 6, B and C), the 5′-UTR of variants B and C are more efficient (∼3-fold of variant A) in directing reporter translation. Similarly, the 5′-UTRs of KOR mRNA are much less efficient than the optimized 5′-UTR derived from the pGL3 reporter (Luc) in the reporter assays.
Discussion
This is the first study to demonstrate differential expression of three KOR mRNA variants in animal tissues as well as during developmental stages. Furthermore, the biologic significance of generating mouse KOR mRNA variants is demonstrated, for the first time, at the level of controlling RNA stability and protein translation. Variant A is expressed most ubiquitously in all the tissues examined in both pre- and postnatal developmental stages, whereas variant B is more specific to the CNS both pre- and postnatally. Variant C is most restricted, detected only in brain stem, spinal cord, and thalamic/hypothalamic areas of postnatal animals. The RNA stability is different between variants A and B. Variant A is more stable, with at 1/2 of ∼12 h, whereas variant B has at 1/2 of only 8 h. In terms of protein translation, variants B and C are more efficient, approximately twice as efficient as variant A. Table 1summarizes the potential regulatory mechanisms of the three KOR mRNA variants, as compiled from this and our previous studies.
Regulation of κ-opioid receptor mRNA variant expression
Differential KOR mRNA variant expression indicates that in addition to transcriptional control, post-transcriptional regulation also is required for proper expression of KOR proteins in different cell types and at different developmental stages. By using pharmacologic methods, different KOR subtypes have been detected in various animal tissues (Kitchen et al., 1990; Loh and Smith, 1990; Lawrence and Bidlack, 1992; Gurwell et al., 1996; Vonkeman et al., 1996); however, only one KOR gene has been identified. It is tempting to speculate that differential distribution of these KOR variants in different cell types may contribute to the distinct pharmacologic characteristics of KOR subtypes. For example, differences in translation efficiency and mRNA stability could result in very different kinetics of KOR protein expression in different cell types. This would explain the discrepancy between the results of detecting opioid receptor expression at the level of RNA (such as in situ hybridization) versus protein (immunohistochemistry or ligand-binding assays). Regulation at the level of RNA stability allows cells with a less-active transcription machinery to efficiently maintain a certain level of messages. For cells expressing messages with a shortert 1/2, a tighter transcriptional control may be required. Therefore, variant A, which is expressed most ubiquitously and has a longer t 1/2, is probably for a more common function, which is widely needed and less regulated. In contrast, variant B is primarily expressed in the CNS and has a shortert 1/2, suggesting a specific function of this variant in the nervous system, which requires an additional level of regulation. The most specific variant, C, is expressed only in certain brain regions and is active only postnatally, suggesting that this variant functions in certain neurons that are born postnatally. Our previous study (Lu et al., 1997) detected KOR variant C expression in early passages of P19 cells. Lately, in later passages of the culture, variant C could no longer be detected reliably. It is known that P19 culture represents a heterogeneous population of embryonal carcinoma cells for which differentiation potential often changes as cultures are passed. The t 1/2 of endogenous KOR variant C in P19 cells cannot be determined reliably; however, translation of variants B and C (both CNS-specific) are more efficient than is variant A (ubiquitous), suggesting that neurons expressing variants B and C requires a better translation efficiency of KOR mRNAs and κ-receptors may play a more specific role in certain types of neurons. It will be interesting in the future to examine what kind of neurons express these variants and how the translation is regulated.
From the expression pattern of the three KOR mRNA variants, it can be concluded that the two KOR promoters are activated in very different patterns, both temporally and spatially. Promoter 1, which controls the transcription of variants A and B, is readily active in early embryos in which neurons are not yet in place. In contrast, promoter 2 becomes active only postnatally and only in restricted brain regions. It is highly possible that promoter 1 is more of a house-keeping promoter and that promoter 2 is neuron-specific. Although the dissected promoter 2 is active in transfection experiments conducted in undifferentiated P19 cells, the endogenous promoter 2, in its natural genomic context, appears to be active only in the mature nervous system, most likely in specific types of neurons, or neuron stem cells, which are born later. Furthermore, in culture systems, variant C is detectable only in early passages of P19 cultures, indicating that promoter 2 is normally suppressed in most cell types. Because promoter 1 is active more ubiquitously, the two variants (A and B), which are transcriptionally controlled by the same promoter (promoter 1), then use the mechanism of alternative splicing for differential regulation. Splicing of variant A occurs widely, whereas splicing of variant B occurs mostly in the CNS. Within the CNS, certain neurons in specific brain stem regions use promoter 2 to generate variant C. In terms of translation, variants B and C, both specific to CNS, are more efficient than is variant A, which is ubiquitous. Therefore, by using three levels of regulation for KOR gene expression, i.e., alternative promoter use, alternative splicing and differential control for translation, animals will be able to very effectively control proper KOR protein expression in different cells. This represents the most efficient way of using a limited amount of genetic material to regulate gene expression. Exactly how the machinery for splicing variants A and B is assembled and what types of cells are capable of generating a specific splicing variant remain interesting questions to be answered.
Because the products of the three KOR mRNA variants appear to be the same (Fig. 6), the reason for generating these KOR mRNA variants is probably for differential controls of a proper level of KOR protein expression in specific cell types. Additionally or alternatively, the information for the control of RNA transport may be encoded in these KOR mRNA variants. Recent studies in mRNA transport suggest that specific information for RNA localization may reside in the primary sequence of mRNA, recognized by the transport motor containing specific RNA-binding proteins (Arn and MacDonald, 1998). In the future, it will be interesting to examine the localization of these KOR mRNA variants in specific neurons and whether the unique exon1/2 junction, or the 3′-UTR of the mouse KOR gene contains information for its specific transport.
Acknowledgment
We thank Dr. M. W. Wessendorf for help in dissecting mouse brain.
Footnotes
-
Send reprint requests to: Dr. Li-Na Wei, Department of Pharmacology, University of Minnesota Medical School, 6-120 Jackson Hall, 321 Church Street S.E., Minneapolis, MN 55455-0217. E-mail:weixx009{at}maroon.tc.umn.edu
-
This work was supported by Grants DA11190, DA11806, DA70554, and DA00564, and from the National Institute on Drug Abuse, National Institutes of Health.
- Abbreviations:
- KOR
- κ-opioid receptor
- RT-PCR
- reverse transcription polymerase chain reaction
- bp
- base pair
- TNT
- transcription/translation
- Luc
- luciferase
- UTR
- untranslated region
- CNS
- central nervous system
- Received June 3, 1999.
- Accepted October 19, 1999.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}