


Abstract
We report the discovery, tissue distribution and pharmacological characterization of a novel receptor, which we have named H4. Like the three histamine receptors reported previously (H1, H2, and H3), the H4 receptor is a G protein-coupled receptor and is most closely related to the H3 receptor, sharing 58% identity in the transmembrane regions. The gene encoding the H4 receptor was discovered initially in a search of the GenBank databases as sequence fragments retrieved in a partially sequenced human genomic contig mapped to chromosome 18. These sequences were used to retrieve a partial cDNA clone and, in combination with genomic fragments, were used to determine the full-length open reading frame of 390 amino acids. Northern analysis revealed a 3.0-kb transcript in rat testis and intestine. Radioligand binding studies indicated that the H4 receptor has a unique pharmacology and binds [3H]histamine (K d = 44 nM) and [3H]pyrilamine (K d = 32 nM) and several psychoactive compounds (amitriptyline, chlorpromazine, cyproheptadine, mianserin) with moderate affinity (K i range of 33–750 nM). Additionally, histamine induced a rapid internalization of HA-tagged H4 receptors in transfected human embryonic kidney 293 cells.
Histamine is a monoamine neurotransmitter thus far known to activate three G protein-coupled receptors (GPCRs), the H1, H2, and H3 receptors (Hill et al., 1997). Although molecular cloning has made possible the identification, isolation, and characterization of the majority of known GPCRs, the histamine receptor subtypes have proven more difficult to identify. The first two histamine receptor genes cloned were H1 (Yamashita et al., 1991) and H2 (Gantz et al., 1991). The identification of the H3 receptor came nearly a decade later (Lovenberg et al., 1999). Collectively, the H1, H2, and H3 receptors share less than 35% identity with one another and each has greater sequence identities with other aminergic receptors. Thus, the histamine receptor gene family is significantly divergent and may have evolved from different ancestral genes (Leurs et al., 2000). In addition, there is evidence that multiple subtypes of the H3 receptor may exist. Pharmacological studies performed on membranes extracted from rat brain tissue revealed two classes of H3 binding sites (West et al., 1990;Leurs et al., 1996). However, efforts to clone a second H3 receptor subtype in the brain have thus far been unsuccessful.
Through molecular cloning techniques, we have identified numerous novel GPCRs, including many subtypes not suspected to exist on the basis of pharmacology. Over the past decade, our cloning efforts have identified a number of GPCR genes, including genes encoding such receptors as the cysteinyl leukotriene CysLT2 (Heise et al., 2000), galanin GalR2 and GalR3 (Kolakowski et al., 1998), thyrotropin-releasing hormone TRH-R2 (O'Dowd et al., 2000), uridine nucleotide receptor UNR (Nguyen et al., 1995), as well as a large cohort of orphan GPCRs for which the endogenous ligands remain to be elucidated (Marchese et al., 1999; Lee et al., 2001). In a scan of the GenBank high-throughput genomic sequence (HTGS) database, we identified a small DNA fragment that had greatest homology to the known histamine receptor genes. Here we report the discovery, tissue distribution, and pharmacological characterization of human DNA encoding a receptor, H4. H4 receptor mRNA had discrete and limited expression in rat testis and intestine. Although the H4 receptor was most closely related to the H3 receptor [58% identity in the transmembrane (TM) regions], it seemed to possess a unique pharmacology, with highest affinity for psychoactive drugs (amitriptyline, chlorpromazine) with a tricyclic structure.
Materials and Methods
Database Searching.
A customized search was used to query the GenBank HTGS database of high-throughput genomic sequences maintained by the National Center for Biotechnology Information (NCBI) with the sequences of various GPCRs, using the FAST_PAN program (Retief et al., 1999).
Screening and Construction of the H4 Receptor Gene.
Partial sequences encoding the H4 receptor were used to design primers to PCR-amplify H4-encoding fragments from human genomic DNA. A fragment encoding the start methionine to TM2 was amplified using primers P1 (5′-ATGCCAGATACTAATAGCACAATC-3′) and P2 (5′-CACAAAGAAGTCAGAGATGG-3′) and another fragment encoding from TM5 to TM6 using primers P3 (5′-TGGTACATCCTTGCCATC-3′) and P4 (5′-TATGGAGCCCAGCAAACAG-3′). PCR products were extracted with phenol/chloroform, precipitated with ethanol, and electrophoresed on a low-melting agarose gel. DNA in the expected size range were excised from the gel, ligated into theEcoRV site of the pBluescript vector (Stratagene, La Jolla, CA), and then the sequence was determined. H4 receptor-encoding fragments were used to screen human and rat genomic libraries (CLONTECH, Palo Alto, CA) and a human testis cDNA library (CLONTECH), as described previously (Marchese et al., 1994). Isolated phages from the human and rat genomic libraries were purified and subcloned, and the sequence was determined as described previously (Marchese et al., 1994). Isolated phages from the human cDNA library were subjected to PCR amplification using primers specific for regions flanking the insert of the λgt10 library vector, subcloned, and sequenced.
A DNA fragment encoding the full-length human H4 receptor was amplified by PCR in three stages using isolated human genomic and cDNA library phage as templates. In stage 1, three overlapping fragments (A, B, and C), together encoding the full-length H4 receptor, were PCR-amplified as follows. Fragment A was obtained using primers P5 (5′-CATCATTTGATGTGATGCCA-3′) and P6 (5′-CAAAGGAATGGAGATCACACCCACAAAGAAGTCAGA-3′) from an isolated genomic library phage obtained with the Met-TM2 encoding DNA probe. Fragment B was obtained using primers P7 (5′-GTGATCTCCATTCCTTTG-3′) and P8 (5′-TCCAATAAATATTCAT-3′) from an isolated testis cDNA library phage. Fragment C was obtained using primers P9 (5′-TGGTACATCCTTGCCATC-3′) and P10 (5′-GAGGTGAGAAAATTGTC-3′) from an isolated genomic library phage obtained with the TM5–6 encoding DNA probe. In stage 2, fragments B and C were joined by PCR using primers P7 and P10. In stage 3, fragments A and B/C were joined together using primer P5 and P10 to obtain a fragment with a length of ∼1200 base pairs encoding the full-length H4 receptor. DNA encoding a hemagglutinin (HA) epitope tag (YPYDVPDYA) was inserted after the start methionine codon using PCR mutagenesis. This DNA was ligated into the EcoRV site of the mammalian expression vector pcDNA3 (Invitrogen, Carlsbad, CA) and its sequence was determined.
Northern Expression Analyses.
mRNA from various human and rat tissues were extracted as described previously (Marchese et al., 1994). Briefly, poly(A)+ RNA was isolated using oligo(dT) cellulose spin columns (Pharmacia, Uppsala, Sweden), denatured and size fractionated on a 1% formaldehyde agarose gel, transferred onto nylon membrane, and immobilized by UV irradiation. The blots were hybridized with 32P-labeled DNA fragments encoding the human and rat H4 receptor, washed with 2× standard saline/phosphate/EDTA and 0.1% SDS at 50°C for 20 min and with 0.1× standard saline/phosphate/EDTA and 0.1% SDS at 50°C for 2 h and exposed to X-ray film at −70°C in the presence of an intensifying screen.
Radioligand Binding Studies.
For these studies, the HA-tagged H4 receptor was transiently transfected into human embryonic kidney 293 cells into 100-mm plates using FUGENE-6 (Roche Molecular Biochemicals, Rotkreuz, Switzerland) at a 6:1 ratio of FUGENE/DNA as detailed previously (Shapiro et al., 2000). At 72 h after transfection, cells were harvested and membranes prepared as described previously (Shapiro et al., 2000); membranes were stored as pellets at −80°C until use. Radioligand binding assays were performed in a total volume of 500 μl in 50 mM Tris-Cl, 0.5 mM EDTA, pH 7.4, with 15 nM [3H]pyrilamine in the 96-well format. After a 1-h incubation at room temperature, membranes were harvested by rapid filtration with a Brandel Harvester followed by two quick washes of ice-cold binding buffer. After drying, filters were placed into scintillation fluid and quantified by liquid scintillation spectrometry. Data were analyzed using GraphPad Prism (GraphPad Software, San Diego, CA) and data presented represents the mean ± S.E.M. of computer-derived estimates of at least three separate experiments, each done in duplicate. For inhibition studies, 12 concentrations of unlabeled ligand spanning 6 log units of test drug were used; for saturation studies, six concentrations of labeled ligand spanning 3 log units were used.
Immunoblot Analysis.
The HA-tagged H4 receptor was transiently transfected into COS-7 monkey kidney cells (American Type Culture Collection, Manassas, VA) and membranes prepared from these cells as described previously (Lee et al., 2000). In brief, tissues were solubilized in sample buffer consisting of 50 mM Tris-HCl, pH 6.5, 1% SDS, 10% glycerol, 0.003% bromphenol blue, and 10% 2-mercaptoethanol. The samples were subjected to polyacrylamide gel electrophoresis with 12% acrylamide gels and electroblotted onto nitrocellulose as described previously (Ng et al., 1996). HA-tagged H4 immunoreactivity was revealed with the 3F10 rat monoclonal antibody (Roche, Laval, Quebec).
Internalization Studies.
For these studies, HA-tagged H4 receptors were transfected into HEK-293 cells as described above into 100-mm plates. At 24 h after transfection, cells were split into 24-well plates containing poly-lysine-coated glass cover slips using Dulbecco's modified Eagle's medium containing 10% dialyzed fetal calf serum. The medium was replaced 24 h later with serum-free Dulbecco's modified Eagle's medium. The next day, cells were exposed to 100 μM histamine for 0, 2, 5, or 15 min and then fixed with freshly prepared 4% paraformaldehyde in PBS for 30 min at room temperature. Cells were then permeabilized on ice (0.2% Triton X-100 in ice-cold PBS) for 20 min and then incubated with blocking buffer (5% nonfat dry milk, 2% bovine serum albumin in PBS) for 1 h and then incubated with a 1:2000 dilution of monoclonal anti-HA antibody in blocking buffer overnight at 4°C. The next day, after extensive washing in room temperature PBS, cells were incubated with secondary antibody (BODIPY-labeled goat anti-mouse; 1:250 dilution in blocking buffer) for 1 h, washed extensively with PBS, and prepared for confocal microscopy as detailed previously (Berry et al., 1996;Kristiansen et al., 2000). Internalization was quantified as described previously (Berry et al., 1996; Willins et al., 1999).
Results and Discussion
As part of our ongoing search of novel genes encoding GPCRs, we queried the GenBank sequence databases maintained by NCBI with known GPCR sequences. A search with the histamine H3 receptor sequence retrieved partial sequences of a novel GPCR-encoding gene in an unfinished sequence of a human contig mapped to chromosome 18 (GenBank accession number AC007922). The retrieved sequence was obtained in three separate fragments, including one fragment that seemed to encode the receptor from the start methionine to TM2, another fragment encoding TM3, and a third fragment encoding TM5 through TM7 of a novel GPCR-like gene. These partial sequences were used to design primers for PCR amplification of human genomic DNA. Two fragments of DNA, encoding the start methionine to TM2 and TM5 to TM6, respectively, were obtained and used as probes to screen a human genomic library. The Met-TM2 probe retrieved two phages encoding the 5′ end of the novel gene from the start methionine to TM2, whereas the TM5-TM6 probe retrieved four phages encoding the 3′ end of the gene from TM5 to the stop codon.
Using the TM5-TM6 probe, we screened several tissues by Northern analyses, which revealed a 3.0-kb signal in rat testis. A human testis cDNA library was screened with both probes, which isolated two identical phages encoding the sequence from TM2 to TM6 of the gene. The sequences were identical in regions of overlap, confirming them to be partial sequences of the same gene. A comparison of the cDNA fragment to the GenBank genomic sequence revealed two introns interrupting the H4 receptor sequence. The first intron was ∼8 kb in length and located within the TM2-encoding region (interrupting the translated sequence “LNLAISDFFVG… VISIPLYIPH”). The second intron was downstream in the region encoding the second intracellular loop (interrupting the translated sequence “DRYLSVSNA… VSYRTQHTGV”). The length of this intron could not be determined, because the GenBank genomic sequence was incomplete.
A BLASTX search of the GenBank database with this novel DNA sequence revealed the greatest overall identity with the histamine receptor H3 (∼40%) and, more distantly, with other amine-type receptors of the GPCR family (<30%). As also reported with the H3 receptor (Lovenberg et al., 1999), this novel receptor shared greater identity with other amine receptors including the serotonin, adrenergic, dopamine, and muscarinic receptors (∼ 25%) than the histamine receptors H1 or H2 (∼20%). An alignment with the known histamine receptors (Fig.1) revealed a higher sequence similarity to the H3 receptor (58% in the TM regions) than to the H1 and H2 receptors (26 and 27% in the TM regions, respectively). It has been determined from their sequence similarities and phylogenetic analyses that the members of the histamine receptor family may have evolved from different ancestral genes and, through convergent evolution, acquired the residues to recognize and bind histamine (Leurs et al., 2000). This novel receptor, which we have named H4, seems to be a novel member of the histamine receptor subfamily, with closest relation to the H3 receptor. The amino acid sequence of the H4 receptor revealed many conserved residues and motifs found within the GPCR family (Fig.2). Among these is an aspartic acid residue in TM 3 that is conserved in all cationic amine receptors and has been shown to be important in binding various amines to GPCRs (Savarese and Fraser, 1992). In addition, the human Met-TM2 probe was used to screen a rat genomic library, which retrieved a phage encoding the 5′ end of the novel gene from the start methionine to TM2. This fragment shared 71% identity (83% in the TM regions) to the human H4 sequence, revealing a rat H4 ortholog.

Alignment of four human histamine receptors H1, H2, H3 and H4. Residues identical among the receptors are shaded. Numeric amino acid positions are indicated on the right. The presence of introns interrupting the H4 sequence are shown as “▴”. H4 sequence data has been deposited in GenBank (accession numberAY008280).
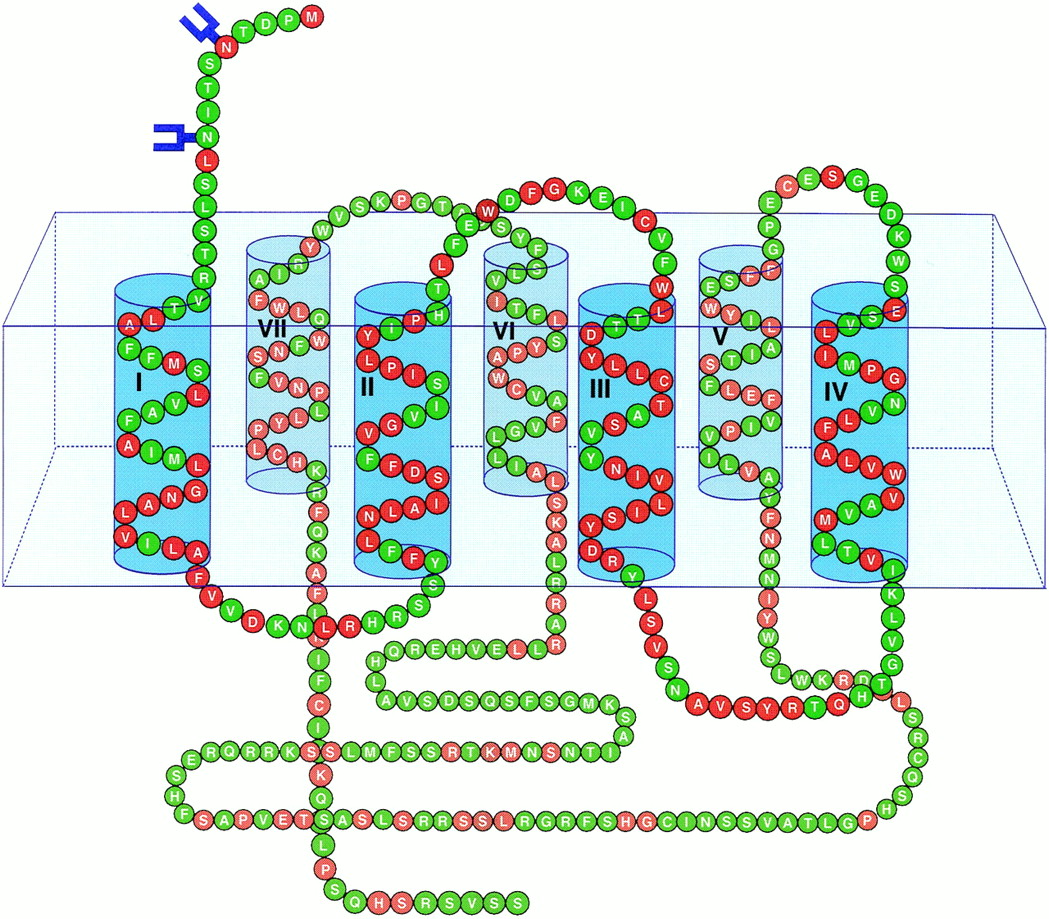
Representative schematic of the human H4 receptor embedded in a cell membrane (box). Transmembrane regions are numbered and depicted with a top (extracellular)/bottom (intracellular) orientation. N-linked glycosylation sites are indicated in blue. Amino acids in red represent residues shared with the histamine receptor H3.
The human Met-TM2 and TM5-TM6 probes were used in Northern analyses of various human and rat tissues. In the rat, the TM5-TM6 probe revealed a single transcript of 3 kb in the testis (Fig.3). The rat DNA fragment encoding from the start methionine to TM2 was used in Northern analyses of various rat tissues, revealing a 3-kb transcript in intestine (data not shown). The H3 and H4 receptors had significantly different mRNA expression distributions. H4 mRNA was detected in two peripheral tissues (with no detectable levels in brain or various peripheral tissues, including heart, stomach, small intestine, kidney, or liver). In contrast, H3 mRNA has been reported to be abundant in the brain (Lovenberg et al., 1999). Thus, H4 is not likely to be the H3-subtype characterized previously in brain tissue (West et al., 1990; Leurs et al., 1996), which suggests that yet another histamine receptor subtype remains unidentified.

Northern blot analysis of the distribution of H4 mRNA in various rat tissues. Each lane contained 10 μg of poly(A)+ RNA.
To test the H4 receptor for pharmacological characterization, we constructed a full-length open reading frame by individually amplifying and joining the three H4-encoding fragments by PCR. The receptor was expressed in HEK-293 cells, and a variety of tritiated ligands including [3H]histamine (histamine receptor nonselective), [3H]pyrilamine (H1 receptor-selective), and [3H]tiotidine (H2 receptor-selective) were tested against unlabeled ligands including mianserin, cyproheptadine, histamine, and clozapine. Specific binding obtained with [3H]histamine (44 nM) or [3H]pyrilamine (5–20 nM) and mianserin (100 μM) represented between 40 and 75% of total binding. Saturation binding studies performed with [3H]histamine and [3H]pyrilamine indicated respectiveK d values of 44 and 32 nM and respectiveB max values of 235 and 437 fmol/mg of membrane protein (Fig. 4). No specific [3H]histamine or [3H]pyrilamine binding was observed with untransfected HEK-293 cells.

Saturation binding studies of [3H]histamine (A) and [3H]pyrilamine (B) specific binding in membranes prepared from HEK-293 cells transiently transfected with H4 cDNA. Shown are typical results from experiment replicated three (A) and two times (B), with data representing mean of duplicate determinations. The curve represents the theoretical fit of the data A) (K d = 44 nM;B max = 235 fmol/mg) and B) (K d = 32 nM;B max = 437 fmol/mg). Nonspecific binding was determined with 100 μM mianserin.
Competition binding studies with various selective and nonselective histaminergic compounds were demonstrated. Typical data are shown in Fig. 5 and summarized in Table1. The highest affinities were for amitriptyline and chlorpromazine, which are tricyclic compounds that have high affinity for the H1 histamine receptor (see on-line database at: http://pdsp.cwru.edu/pdsp.asp ). Doxepin, cinnarizine, and promethazine (H1-selective antagonists) also displayed high affinity for the H4 receptor. Imetit (H3-selective agonist) and dimaprit (H2-selective agonist) had weak affinities for the H4 receptor, whereas mianserin (an H1 and H2 antagonist), cyproheptadine (a nonselective histamine/serotonin antagonist), and clozapine (an atypical antipsychotic drug with high affinity for a large number of receptors) had moderate affinities. The pharmacological profile of the H4 receptor is distinct from the histamine receptors (Table 1).

Two sets (A and B) of competition binding of [3H]pyrilamine with various ligands in membranes prepared from HEK-293 cells transiently transfected with H4 cDNA. Shown are typical results from competition binding experiments using 20 nM [3H]pyrilamine and various concentrations of unlabeled ligands. Also shown are typical results obtained with untransfected HEK-293 cells studied in parallel (A).
Ligand affinities for the H4 receptor
An HA epitope tag-encoding sequence was inserted after the start methionine for Western blot visualization. This plasmid was transiently transfected into COS-7 cells. Immunoblot analyses of membranes from these cells revealed high expression of the H4 receptor (Fig.6A), with bands at 44 kDa, 85 kDa, and higher molecular mass species. The 44-kDa band matched the expected mass of the unglycosylated receptor and the 85-kDa band matched the glycosylated form. The bands >250 kDa represent oligomeric receptor species (Lee et al., 2000). The functional activity of the HA-tagged H4 receptor was examined by measuring histamine-induced internalization. As shown in Fig. 6B, exposure to 100 μM histamine caused an internalization of H4 receptors from the plasma membrane to intracellular sites. Quantification revealed a rapid internalization of H4 receptors with significant internalization at 2 min after agonist exposure (Fig. 6C). These results indicated that the surface expression of the H4 receptor is functionally regulated by histamine exposure in a time-dependent fashion.
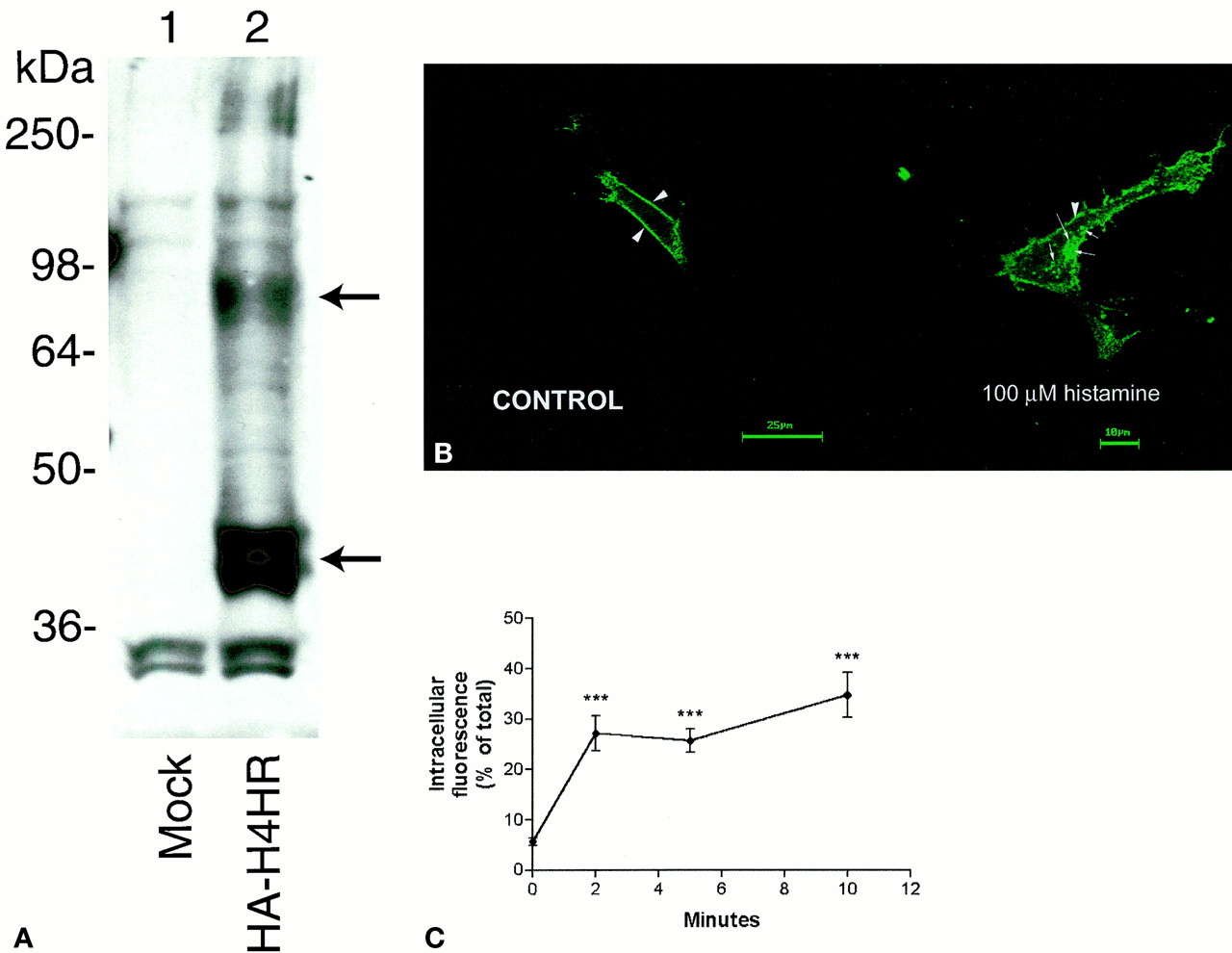
A, immunoblot analysis of membranes from mock-transfected COS-7 cells (lane 1) and COS-7 cells expressing the HA-H4 receptor (lane 2). Membrane protein (25 μg) was used in each lane. Arrows indicate the unglycosylated (bottom) and glycosylated (top) receptor. B) histamine-induced internalization of the H4 receptor in HEK-293 cells. Shown are representative confocal micrographs in which HA-tagged H4 receptors were examined in cells exposed to vehicle (PBS) or 100 μM histamine for 5 min and then prepared for microscopy as described previously (Berry et al., 1996; Willins et al., 1999;Kristiansen et al., 2000). Arrowheads and arrows indicate cell-surface and internalized receptors, respectively. C, histamine-induced time-dependent internalization of the H4 receptor in HEK-293 cells. Shown are the mean ± S.E.M. of the percentage internalization of HA-tagged H4 receptors in HEK-293 cells (n = 20–30 cells/time point) after exposure to histamine (100 μM) for various time periods. ***significantly different compared with 0-min value (P < 0.0001).
During the preparation of this article, other researchers (Oda et al., 2000) also reported the identification of a cDNA encoding a novel histamine receptor. This cDNA sequence varied from our sequence at three nucleotide positions, which translated into differences in three amino acids. Specifically, Ala138, His206, and Gln253 (as found in our sequence) were replaced by valine and two arginines, respectively. In the study by Oda et al., expression analyses revealed signals in peripheral blood leukocytes, small intestine, spleen and colon, and no expression in the brain. In addition, they reported histamine signaling through the novel receptor to be pertussis toxin-sensitive, suggesting a Gi/o pathway of activation. For our receptor, we examined several second messenger-effector systems. We were not able to demonstrate H4 receptor-mediated inhibition of forskolin-stimulated adenylyl cyclase, alterations in phosphoinositide hydrolysis, or mitogen-activated protein kinase phosphorylation (extracellular signal-regulated kinase 1/2 phosphorylation) in HEK-293 cells.
In conclusion, we report the discovery of a novel histamine receptor, H4. Previously, it was observed that histamine receptors shared greater sequence similarities with other biogenic amine-binding GPCRs than with one another. H4 shared highest sequence similarity with the previously reported histamine H3 receptor. In combination with the H1, H2, and H3 receptors, this receptor, with its unique distribution and pharmacology, will undoubtedly provide further insight into the physiological functions and therapeutic applications of this receptor family.
Footnotes
- Received October 4, 2000.
- Accepted January 5, 2001.
-
Send reprint requests to: Dr. Brian F. O'Dowd, Department of Pharmacology, University of Toronto, Medical Science Building, 8 Taddle Creek Rd. Rm 4353, Toronto, Ontario, Canada M5S 1A8. E-mail:brian.odowd{at}utoronto.ca
-
This research was funded by the Canadian Institutes of Health Research (B.F.O. and S.R.G.), the National Institute on Drug Abuse (B.F.O. and S.R.G.), and in part by K02-MH01366 and N01–80005 to B.L.R.
-
T.N. and D.A.S. contributed equally to this work.
Abbreviations
- GPCR(s)
- G protein-coupled receptor(s)
- HTGS
- high-throughput genomic sequence
- TM
- transmembrane
- NCBI
- National Center for Biotechnology Information
- PCR
- polymerase chain reaction
- HA
- hemagglutinin
- HEK
- human embryonic kidney cells
- kb
- kilobase pair
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}