


Abstract
The nuclear receptor peroxisome proliferator-activated receptor (PPAR) is activated by a diverse group of acidic ligands, including many peroxisome proliferator chemicals present in the environment. Janus tyrosine kinase-signal transducer and activator of transcription (JAK-STAT) signaling is activated by multiple cytokines and hormones and leads to the translocation of dimerized STAT proteins to the nucleus where they activate transcription of target genes. Previous studies have shown that growth hormone (GH)-activated STAT5b can inhibit PPAR-regulated transcription. Here, we show that this inhibitory cross-talk is mutual, and that GH-induced, STAT5b-dependent β-casein promoter-luciferase reporter gene transcription can be inhibited up to ∼80% by ligand-activated PPARα or PPARγ. Dose-response experiments showed a direct relationship between the extent of PPAR activation and the degree of inhibition of STAT5-regulated transcription. PPAR did not block STAT5b tyrosine phosphorylation or inhibit DNA-binding activity. Both PPARs inhibited the transcriptional activity of a constitutively active STAT5b mutant, indicating that inhibition occurs downstream of the GH-stimulated STAT5 activation step. Transcriptionally inactive, dominant-negative PPAR mutants did not block STAT5b inhibition by wild-type PPAR, indicating that PPAR target gene transcription is not required. PPARα retained its STAT5b inhibitory activity in the presence of the histone deacetylase inhibitor trichostatin, indicating that enhanced histone deacetylase recruitment does not contribute to STAT5b inhibition. PPARα lacking the ligand-independent AF-1 trans-activation domain failed to inhibit STAT5b, highlighting the importance of the AF-1 region in STAT5-PPAR inhibitory cross-talk. These findings demonstrate the bidirectionality of cross-talk between the PPAR and STAT pathways and provide a mechanism whereby exposure to environmental chemical activators of PPAR can suppress expression of GH target genes.
Peroxisome proliferator-activated receptors (PPARs) are nuclear receptors that control a variety of cellular processes in response to a diverse group of natural and synthetic ligands (Escher and Wahli, 2000). Like other nuclear receptors, PPARs have a conserved protein structure containing distinct DNA-binding, trans-activation, and ligand-binding domains. After ligand binding, PPAR heterodimerizes with the retinoid X receptor and binds upstream of, and activates target genes. The three identified mammalian PPAR subtypes (α, γ, and δ) have unique functions, tissue localizations, and ligand selectivities. PPARα regulates expression of genes involved in lipid metabolism, such as those encoding the peroxisomal enzymes acyl-CoA oxidase, bifunctional enzyme, and thiolase. PPARα has been implicated in rodent hepatocarcinogenesis (Corton et al., 2000), which reflects in part the inhibition of hepatocyte apoptosis (Roberts et al., 1998). Humans and several other species are resistant to the peroxisome proliferative and hepatocarcinogenic effects of PPARα activators, in part because of the significantly lower PPARα expression level in human liver (Palmer et al., 1998). In contrast, PPARγ is expressed at high levels in multiple human tissues, including adipose tissue, where it plays a key role in adipocyte differentiation (Tontonoz et al., 1994). PPARα is activated by hypolipidemic compounds of the fibrate class, such as clofibrate and Wy-14,643, and by naturally occurring long-chain fatty acids. Specific ligands and activators of PPARγ include antidiabetic thiazolidinedione drugs (Lehmann et al., 1995) and the prostaglandin metabolite 15-deoxy-Δ12,14 prostaglandin J2 (Escher and Wahli, 2000). Less is known about PPARδ, which is thought to play a role in development (Peters et al., 2000).
Previous studies have demonstrated the potential for cross-talk between STAT transcription factors and nuclear receptors such as PPARs. STATs are latent cytoplasmic signaling molecules activated by tyrosine-phosphorylation catalyzed by JAKs, tyrosine kinases associated with many cytokine and growth factor receptors, including growth hormone (GH) receptor (GHR) (Darnell, 1997). The tyrosine phosphorylated STATs form homo- and heterodimeric complexes that translocate to the nucleus where they bind to specific DNA response elements and stimulate target gene transcription (Kisseleva et al., 2002). Inhibition of STAT1-regulated transcription by PPARγ occurs in HeLa cells (Ricote et al., 1998), although not in COS-1 cells (this report). STAT1 can decrease PPARγ-regulated gene transcription indirectly, by binding upstream of, and repressing transcription of the PPARγ gene, leading to decreased PPARγ protein expression (Hogan and Stephens, 2001). STAT5 transcriptional activity is strongly inhibited by the estrogen receptor (ER) via mechanisms that involve a direct interaction between the receptor and STAT5 (Faulds et al., 2001) and via an indirect inhibitory effect of ER on STAT5 activation and nuclear localization (Sueyoshi et al., 1999). STAT5 inhibits transcription stimulated by glucocorticoid receptor, mineralocorticoid receptor, and progesterone receptor, but, conversely, these three steroid receptors synergize with STAT5 to enhance STAT5 target gene transcription (Stoecklin et al., 1999). STAT5 can also inhibit PPARα- and PPARγ-regulated transcription, by a mechanism that involves the AF-1 ligand-independent trans-activation domain of PPAR (Zhou and Waxman, 1999a,b). The possibility that PPAR may, in turn, inhibit STAT5 transcriptional activity is suggested by the finding that ligand activation of PPARα leads to down-regulation of several GH-regulated, sexually dimorphic liver genes (Corton et al., 1998), which are regulated, in part, by STAT5b (Udy et al., 1997; Park et al., 1999; Park and Waxman, 2001). PPAR inhibition of STAT5 transcriptional activity would provide a mechanism whereby peroxisome proliferator chemicals (PPCs) may down-regulate such GH-regulated genes.
STAT5 is coexpressed with PPAR in many tissues, including hepatocytes (PPARα) and preadipocytes (PPARγ). STAT5 increases in expression early during the course of adipogenesis (Stephens et al., 1999), becomes activated during differentiation, and contributes to the enhanced expression of proadipogenic transcription factors, including PPARγ (Nanbu-Wakao et al., 2002). PPARα, as well as PPARγ, can be activated by a broad range of environmental chemicals (Maloney and Waxman, 1999; Hurst and Waxman, 2003), and cross-talk with STATs is potentially an important route through which foreign chemical exposure may impact on endogenous pathways of metabolism and differentiation. The STAT and PPAR pathways are tightly regulated by an overlapping set of nuclear regulatory proteins, including coactivators (Chen and Li, 1998), and by post-translational modification, e.g., inhibitory serine phosphorylation of the NH2-terminal AF-1 domain (A/B domain) of PPARγ (Adams et al., 1997) and phosphorylation of several STATs, including STAT5a and STAT5b, at a conserved COOH-terminal serine, in some cases leading to stimulation and in other cases inhibition of transcriptional activity (Yamashita et al., 1998; Park et al., 2001). Given the multiple regulatory mechanisms controlling STAT and PPAR signaling pathways, there may be multiple mechanisms by which the activation of one pathway can lead to cross-talk with the other.
In the present study, we investigate the effects that ligand-activated PPARα and PPARγ have on STAT5b-regulated reporter gene transcription in GH-stimulated cells. PPARα and PPARγ are shown to inhibit the transcriptional activity of STAT5b at a step downstream of GH activation, providing a mechanistic explanation for the previously observed down-regulation of GH-activated genes by PPCs (Corton et al., 1998). We evaluate the mechanism that underlies this inhibitory cross-talk and highlight the importance of the NH2-terminal AF-1 trans-activation domain of PPAR, a protein domain that was previously found to be a target of the inhibitory effects of STAT5b on PPARα (Zhou and Waxman, 1999b).
Materials and Methods
Plasmids. The PPAR-activated firefly luciferase reporter pHD(x3)Luc, obtained from Dr. J. Capone (McMaster University, Toronto, ON, Canada), contains three tandem copies of the peroxisome proliferator response element from the rat enoyl-CoA hydratase/3-hydroxyacyl-CoA dehydrogenase gene upstream of a minimal promoter cloned into the plasmid pCPS-luc. The reporter plasmid pZZ1, provided by Dr. B. Groner (Institute for Experimental Cancer Research, Freiburg, Germany), contains the β-casein milk protein gene promoter upstream of the firefly luciferase gene. Mouse PPARα cloned into the expression plasmid pCMV5 was obtained from Dr. E. Johnson (Scripps Research Institute, La Jolla, CA). The STAT5 reporter plasmid pT109-4Xntcp-Luc, which contains four copies of a STAT5 response element from the rat ntcp gene, was provided by Dr. M. Vore (University of Kentucky, Lexington, KY). STAT5b1*6 cDNA was excised from the pMX-puro-STAT5b1*6 plasmid, provided by Dr. Toshio Kitamura (University of Tokyo, Tokyo, Japan), and the EcoRI-NotI fragment was subcloned into the expression vector pCI (Promega, Madison, WI) by Dr. S. H. Park of this laboratory. The PPARγ expression plasmid pSV-Sport-mPPARγ was obtained from Dr. J. Reddy (Northwestern University, Chicago, IL). Rat GHR cloned into the expression plasmid pcDNAI was provided by Dr. N. Billestrup (Hagedorn Research Institute, Gentofte, Denmark). pME18S expression plasmid encoding mouse STAT5b was obtained from Dr. A. Mui (DNAX Research Institute of Molecular and Cellular Biology, Inc.). An expression plasmid encoding hPPARα-6/29, a naturally occurring dominant-negative inhibitory variant of human liver PPARα, was provided by Dr. Ruth Roberts (Zeneca Central Toxicology Lab, Brixham, UK) (Roberts et al., 1998). FLAG epitope-tagged wild-type human PPARγ and a dominant-negative human PPARγ, PPARγ-L466A/E469A, both subcloned into pcDNA, were provided by Dr. V.K.K. Chatterjee (University of Cambridge, Cambridge, UK) (Barroso et al., 1999). pNCMV-PPARα and PPARα lacking the A/B domain, pNCMV-PPARαΔA/B, were gifts of Dr. T. Osumi (Himeji Institute of Technology, Kamigori Hyogo, Japan) (Hi et al., 1999). The STAT1 luciferase reporter p36-8GAS-Luc, containing eight interferon γ-activated sites cloned upstream of a p36 minimal promoter, was provided by Dr. C. K. Glass (University of California San Diego, La Jolla, CA) (Ricote et al., 1998). Renilla reniformis luciferase expression plasmid pRL-CMV was purchased from Promega.
Cell Culture and Transfection. COS-1 cells were grown in Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal calf serum. Cells were plated in 48-well tissue culture plates at a density of 2.5 × 104 cells/well in 500 μl of medium. Twenty-four hours later the medium was replaced with 250 μl of DMEM + serum, and the cells were transfected using 0.3 μl of FuGENE 6 transfection reagent (Roche Diagnostics, Indianapolis, IN) and 250 ng of total DNA per well of a 48-well plate. Salmon sperm DNA was used as a carrier to adjust the total to 250 ng of DNA per well. The culture medium was changed to serum-free DMEM 24 h after addition of the DNA-FuGENE 6 mixture to the cells. Chemical hormone treatments (e.g., Wy-14,643, troglitazone, GH) were supplied to the cells in this medium change at the concentrations indicated in the figure legends. Cells were lysed 24 h later in 250 μl of passive lysis buffer (Promega), and firefly and R. reniformis luciferase activity was measured using a dual luciferase assay kit (Promega). Transfections were performed using the following amounts of plasmid DNA/well of a 48-well tissue culture plate: 90 ng of reporter plasmid (pHD(x3)-Luc, pT109-4Xntcp-Luc, p36-8GAS-Luc or pZZ1), 5 ng of PPARα or PPARγ, and 1 ng each of STAT5b, GHR, and pRL-CMV. Mouse PPAR expression plasmids were used, except as noted.
EMSA and Western Blot Analysis. EMSA analysis using probes for STAT5 and PPAR DNA-binding activity was performed as described previously (Zhou and Waxman, 1999a). For Western blotting, whole cell lysates from transfected COS-1 cells were subjected to 7.5% SDS-polyacrylamide gel electrophoresis and then transferred to nitrocellulose membranes. Western blotting was performed using anti-STAT5b, anti-hPPARγ (Santa Cruz Biotechnology, Inc., Santa Cruz, CA), and anti-phosphotyrosyl-694/699 STAT5 antibody (Cell Signaling Technology Inc., Beverly, MA) as described previously (Zhou and Waxman, 1999a).
Results
PPARα and PPARγ Inhibit STAT5b-Regulated Transcription. GH-activated STAT5b inhibits PPARα-dependent gene transcription by ∼80% when evaluated in a reporter gene trans-activation assay (Zhou and Waxman, 1999a). To determine whether the cross-talk between the PPAR and STAT5b signaling pathways is bidirectional, we examined the effect of PPAR on GH-regulated STAT5 transcriptional activity. GH-induced STAT5 signaling and PPAR transcriptional activity were reconstituted in COS-1 cells by cotransfection of key components: PPARα or PPARγ for PPAR signaling; and GHR, STAT5b, and a STAT5b-activated reporter plasmid for GH signal transduction (pT109-4Xntcp-Luc or pZZ1). The transfected cells were stimulated for 24 h with GH either in the presence or absence of the PPAR form-specific ligands troglitazone (PPARγ) and Wy-14,643 (PPARα). Ligand-activated PPARα (Fig. 1A) and PPARγ (Fig. 1B) effected 65 to 80% inhibition of GH-stimulated reporter activity driven by four tandem copies of an isolated STAT5 response element (reporter plasmid pT109-4Xntcp-Luc). The inhibitory action of PPARγ was also manifest in the context of the native promoter sequence of β-casein, a STAT5 target gene in the mammary gland (pZZ1 reporter; Fig. 1C). This inhibition was seen using either mouse (Fig. 1) or human PPARs (data not shown). Dose-response experiments revealed a direct relationship between the extent of PPARγ activation, monitored with a PPAR reporter plasmid (pHD(x3)Luc), and the extent to which PPARγ inhibits STAT5 reporter gene activity as a function of troglitazone concentration (Fig. 1D). A close correlation was also seen between the degree of PPARα activation in cells treated with various concentrations of the PPARα activator Wy-14,643 and the extent of STAT5b inhibition (Fig. 1E). The PPAR inhibitory effect is specific to STAT5b, insofar as troglitazone-activated PPARγ did not inhibit interferon γ-activated STAT1 reporter activity (Fig. 1F), which is probably mediated by endogenous COS-1 cell STAT1 protein (Zhou and Waxman, 1999b). This finding is consistent with the absence of an effect of STAT1 on PPARα-stimulated transcription (Zhou and Waxman, 1999b). Thus, the inhibitory cross-talk between PPARs and STATs is bidirectional and is restricted to a subset of STAT subtypes.

PPARα and PPARγ inhibit STAT5b, but not STAT1-regulated reporter gene activity. A, transcription of the STAT5b luciferase reporter plasmids pT109-4Xntcp-Luc (A and B) and pZZ1 (C) regulated by GH-activated STAT5b is inhibited by ligand-activated PPARα (A) and PPARγ (B and C). COS-1 cells were transfected for 24 h with the indicated STAT5b reporter plasmid, together with pRL-CMV as an internal control and expression plasmids encoding STAT5b and GHR, in the absence (▪) or presence of PPAR (□). Twenty-four hours after transfection, cells were stimulated for 24 h with GH (500 ng/ml) and the indicated PPAR ligands (3 μM troglitazone for PPARγ, 5 μM Wy-14,643 for PPARα). Cell lysates from triplicate wells were then prepared and assayed for luciferase activity. Activities are expressed as firefly luciferase normalized by the reporter activity of a R. reniformis luciferase internal standard, mean ± S.D. values. D and E, COS-1 cells were transfected as described in A in the presence of the STAT5b reporter pZZ1 (▪) or the PPAR reporter pHD3(x3)Luc (▵). Cells were treated with GH (250 ng/ml) and the indicated concentrations of troglitazone (PPARγ-cotransfected cells; D) or Wy-14,643 (PPARα-cotransfected cells; E). An 11-fold activation of PPARγ (EC50 = 220 nM) and a 72% inhibition of STAT5b (EC50 = 160 nM) by troglitazone was observed (D). A 3.5-fold activation of PPARα (EC50 = 31 nM) and a 62% inhibition of STAT5b (EC50 = 46 nM) by Wy-14,643 was observed (E). Normalized luciferase activities are graphed as a percentage of the observed maximal inhibition (▪) or as a percentage of the observed maximal stimulation (▵). F, troglitazone-activated PPARγ failed to inhibit an interferon γ-activated luciferase reporter. COS-1 cells were cotransfected for 24 h with the interferon γ-activated reporter plasmid p36-8GASluc, pRL-CMV as an internal control, in the absence (▪) or presence of an expression plasmid for PPARγ (□). Beginning 24 h after transfection, cells were treated with interferon γ (10 ng/ml) and troglitazone (3 μM) for 24 h. Data were analyzed as in A-C. Data presented is representative of three independent experiments (A–C) or two independent experiments (D–F).
PPARα Suppresses Transcriptional Activity of a Constitutively Active STAT5b Mutant. STAT5b1*6 is a constitutively active STAT5b that contains two site-specific mutations, H299R and S711F, which render it constitutively phosphorylated on tyrosine 699 and transcriptionally active in the absence of hormone or cytokine stimulation (Onishi et al., 1998). We used this mutant to determine whether the inhibitory effects of PPAR on STAT5b transcriptional activity occur at the level of STAT activation. When expressed in COS-1 cells, in the absence of GHR or GH stimulation, STAT5b1*6 strongly activates transcription of the STAT5 reporter pZZ1 (Fig. 2, third bar). This transcriptional activity was inhibited by PPARα and by PPARγ when activated by their respective ligands, Wy-14,643 and troglitazone (Fig. 2, last four bars). This PPAR-dependent inhibition of constitutively active STAT5b transcriptional activity suggests that the PPARs act downstream of the GHR-dependent STAT5 activation step.

PPARs inhibit STAT5b-regulated transcription at a step downstream of
GH-stimulated STAT5b activation. COS-1 cells were cotransfected for 24 h with
the STAT5 luciferase reporter pT109-4Xntcp-Luc, pRL-CMV, expression plasmids
encoding STAT5b and GHR or the constitutively active STAT5b1*6, in
the absence ( ) or presence of PPARα (□) or PPARγ (
) or presence of PPARα (□) or PPARγ ( ).
Beginning 24 h after transfection, cells were treated with GH (500 ng/ml),
Wy-14,643 (5 μM) or troglitazone (3 μM) for 24 h, as indicated. Cell
lysates from triplicate wells were then prepared and assayed for luciferase
activity. Activities are expressed as firefly luciferase normalized by the
reporter activity of the R. reniformis luciferase internal standard,
mean ± S.D. values. Data presented is representative of two independent
experiments.
).
Beginning 24 h after transfection, cells were treated with GH (500 ng/ml),
Wy-14,643 (5 μM) or troglitazone (3 μM) for 24 h, as indicated. Cell
lysates from triplicate wells were then prepared and assayed for luciferase
activity. Activities are expressed as firefly luciferase normalized by the
reporter activity of the R. reniformis luciferase internal standard,
mean ± S.D. values. Data presented is representative of two independent
experiments.
STAT5b Tyrosine Phosphorylation Is Unaffected by PPAR Inhibitory Cross-Talk. STAT5b tyrosine phosphorylation and STAT5b transcriptional activity are strongly inhibited by the liver nuclear factor hepatic nuclear factor 3β (Park and Waxman, 2001). To test whether STAT5b tyrosine phosphorylation can also be inhibited by PPAR, COS-1 cells transfected with PPARγ, STAT5b, GHR, and the STAT5b reporter pZZ1 were serum-starved and then treated with GH and troglitazone for 4 h. This time period is sufficient for detection of the transient, GH-dependent phosphorylation of STAT5b protein by Western blotting and for analysis of firefly luciferase reporter activity in the same extracts. Under these conditions, troglitazone-activated PPARγ inhibited STAT5b reporter activity by ∼50% (data not shown). Transfection of PPARγ did not affect STAT5b protein levels as monitored by Western blotting (Fig. 3A, top). Moreover, STAT5b protein levels were unchanged after stimulation of the cells with GH, troglitazone, or both ligands in combination. Finally, the GH-dependent increase in tyrosine phosphorylated STAT5b was unaffected by troglitazone-activated PPARγ, as revealed by Western blot analysis using STAT5b-phosphotyrosine-699-specific antibody (Fig. 3A, middle, lanes 16–18 versus 13–15; also note the low mobility phosphotyrosyl-STAT5b band seen at top).

STAT5b and phospho-STAT5 protein levels are not altered by PPAR. COS-1 cells were cotransfected with hPPARγ1, STAT5b, and GHR expression plasmids for 24 h and then stimulated for 30 min with GH (250 ng/ml) and troglitazone (3 μM). Cell lysates were analyzed on Western blots probed with antibodies to STAT5b (top), phosphotyrosyl-699-STAT5b (middle), and PPARγ1 (bottom). Data presented is representative of three independent experiments. B, COS-1 cells transfected with expression plasmids encoding PPARα, STAT5b, and GHR were assayed for STAT5 EMSA activity. Cells were treated with GH (250 ng/ml) and Wy-14,643 (5 μM) for 30 min, as indicated, followed by preparation of whole cell extracts. EMSA assays were carried out with a 32P-labeled β-casein promoter DNA probe, which contains a STAT5 binding site. Anti-STAT5b-specific antibody was used to supershift a STAT5b-containing DNA-protein complex (supershift; lane 12). Data shown are for two or three independent samples in each treatment group.
STAT5b DNA-Binding Activity Is Not the Target of PPAR Inhibitory Cross-Talk. We next investigated whether PPARα inhibits STAT5b DNA-binding activity, which was assayed by EMSA using a STAT5 binding site DNA probe. No decrease in STAT5b DNA-binding was seen in extracts prepared from cells cotransfected with PPARα, independent of whether PPARα was activated by Wy-14,643 (Fig. 3B, lanes 8–11 versus lanes 5–7). Therefore, PPARα does not inhibit STAT5b-regulated transcription by blocking the binding of STAT5b to its DNA binding sites. Together, these findings establish that PPARs inhibit STAT5 signaling at a step downstream of initial, cell surface receptor-dependent activation step.
Pretreatment of Cells with PPAR Ligand Does Not Increase STAT5b Inhibition. We investigated the possibility that PPAR may activate transcription of a gene that codes for a STAT5 inhibitory protein, such as PIAS3 (Rycyzyn and Clevenger, 2002). In such a case, treatment of the cells with a PPAR activator several hours before the activation of STAT5b by GH would increase cellular levels of the inhibitory protein factor, thereby enhancing the PPAR inhibitory effect. This hypothesis was tested by treating PPAR- and STAT5b-signaling component-transfected COS-1 cells with PPAR ligand either 1) simultaneously with GH, followed by 24-h incubation, as was done in the experiments shown in Figs. 1 and 2 (Fig. 4, a–c); 2) 8 h before GH, followed by costimulation of the cells with Wy-14,643 and GH for 16 h (Fig. 4, d–f); or 3) 16 h before GH, followed by an 8-h period of ligand costimulation (Fig. 4, g–i). Although STAT5b reporter activity was somewhat reduced in cells stimulated by GH for 8 h (Fig. 4h) or 16 h (Fig. 4e) compared with 24 h (Fig. 4b), the extent to which PPARα inhibited STAT5b did not increase with Wy-14,643 pretreatment (Fig. 4, i versus h, compared with Fig. 4, f versus e and c versus b).
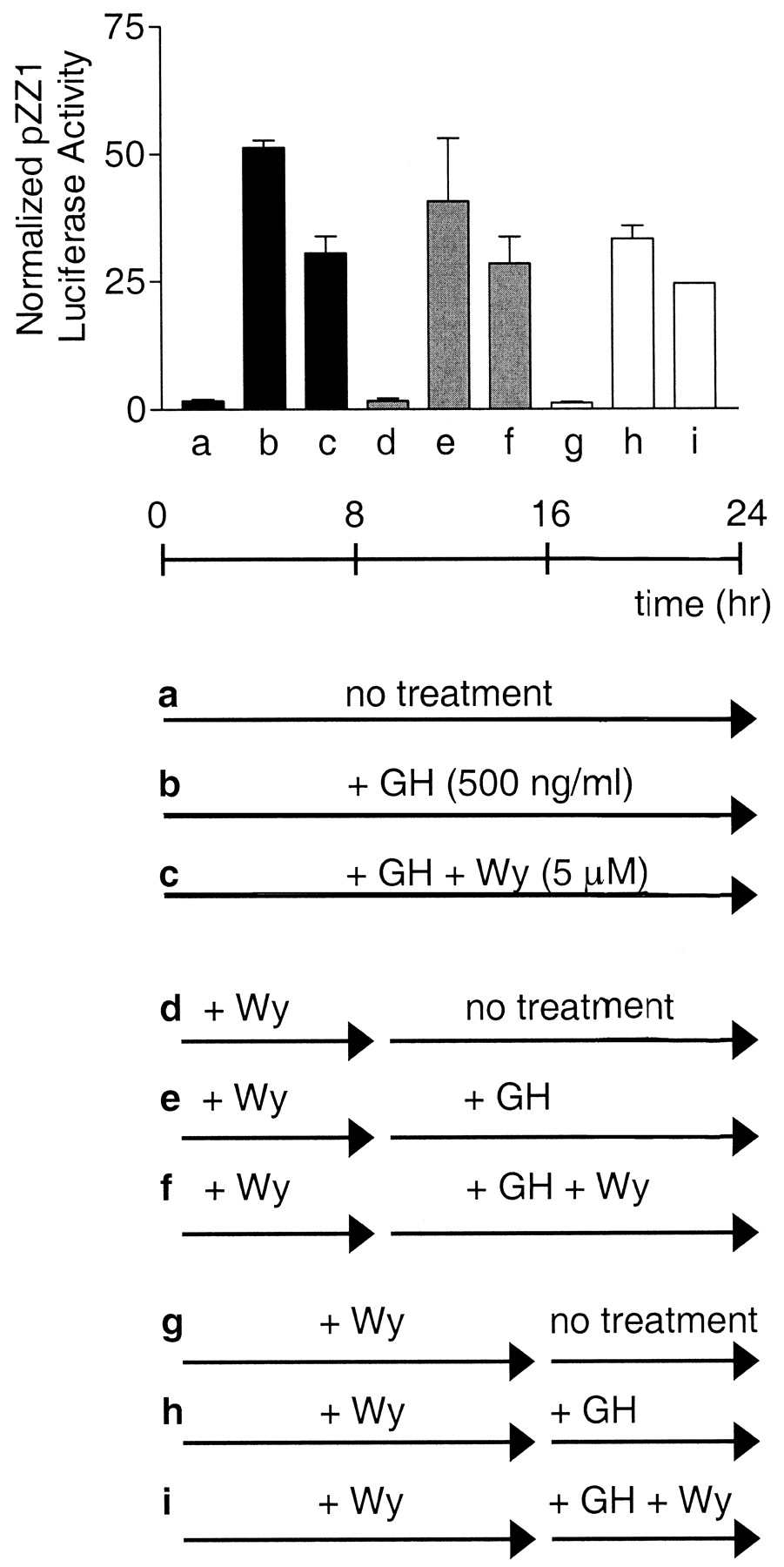
Pretreatment with PPARα ligand does not increase the extent to which
STAT5b-regulated transcription is inhibited by PPARα. COS-1 cells were
cotransfected for 24 h with the STAT5b luciferase reporter pZZ1, pRL-CMV as an
internal control, and expression plasmids encoding STAT5b, GHR, and
PPARα. Beginning 24 h after transfection, cells were treated with GH
(500 ng/ml) and Wy-14,643 (5 μM). Cell lysates from triplicate wells were
prepared and assayed for luciferase activity 24 h later. Activities are
expressed as firefly luciferase normalized by the reporter activity of a
R. reniformis luciferase internal standard. Cells in experiments a to
c were treated for 24 h as is routine (▪). Cells in experiments d to f
were treated for 16 h, with a pretreatment of Wy for 8 h ( ). Cells in
experiments g to i were treated for 8 h, with a pretreatment of Wy-14,643 for
16 h (□).
). Cells in
experiments g to i were treated for 8 h, with a pretreatment of Wy-14,643 for
16 h (□).
Dominant-Negative PPAR Mutants Do Not Reverse the Inhibition of STAT5b Activity by Wild-Type PPARs. The hypothesis that PPAR target gene transcription is required for STAT5b inhibition was further tested using dominant-negative inhibitors of PPARα and PPARγ. hPPARα-6/29 is a naturally occurring variant of human PPARα that heterodimerizes with retinoid X receptor and binds to peroxisome proliferator response element sequences, but is unable to activate transcription after ligand stimulation. hPPARα-6/29 acts as a dominant-negative inhibitor of PPARα-induced gene transcription (Roberts et al., 1998). The PPARγ double mutant L468A/E471A is a potent dominant-negative inhibitor of wild-type PPARγ. It contains mutations in the ligand-binding AF-2 domain, resulting in a receptor that retains ligand-binding and DNA-binding activities, but exhibits reduced coactivator recruitment and delayed corepressor release (Gurnell et al., 2000). We first verified the dominant-negative activities of these PPAR mutants toward the corresponding wild-type PPARs. Transfection of increasing amounts of dominant-negative PPAR plasmid led to a dose-dependent inhibition of both basal and ligand-induced PPAR activity, as shown for PPARγ (Fig. 5A) and PPARα (Fig. 5B) using the PPAR reporter pHD(x3)luc. However, cotransfection of the dominant-negative PPARs failed to block the suppression of STAT5b transcriptional activity by wild-type PPARγ (Fig. 5C) or PPARα (Fig. 5D). These experiments confirm that PPAR transcriptional activity is not required for STAT5b inhibition. Thus, PPAR does not inhibit STAT5b by stimulating transcription of a STAT5 inhibitor.

Impact of dominant-negative PPARs on PPAR-STAT5 cross-talk. A and B,
transcriptionally inactive dominant-negative mutant human PPARs inhibit
wild-type PPAR-regulated transcription. COS-1 cells were transfected for 24 h
with the PPAR reporter plasmid pHD(x3)luc and pRL-CMV as an internal control.
A, cells were cotransfected with 0.5 ng of pcDNAFlag-γ1 in the absence
(▪) or the presence of either 0.5 ng (□) or 5 ng ( ) of the
PPARγ dominant-negative construct pcDNAFlag-γ1 L466A/E469A. Data
presented are representative of three independent experiments. B, cells were
cotransfected with 5 ng of mPPARα in the absence (black columns) or the
presence of either 5 ng (□) or 25 ng (
) of the
PPARγ dominant-negative construct pcDNAFlag-γ1 L466A/E469A. Data
presented are representative of three independent experiments. B, cells were
cotransfected with 5 ng of mPPARα in the absence (black columns) or the
presence of either 5 ng (□) or 25 ng ( ) of the PPARα
dominant-negative construct hPPARα6/29. C and D, transcriptionally
inactive dominant-negative PPARs do not block the inhibition of
STAT5b-stimulated transcription by wild-type PPAR. COS-1 cells were
transfected for 24 h with STAT5b reporter plasmid (pZZ1 in C, pT109-4Xntcp-Luc
in D) and pRL-CMV as an internal control. C, cells were cotransfected with 0.5
ng of pcDNAFlag-γ1 in the absence (▪) or the presence of 0.5 ng
(□) or 5 ng (
) of the PPARα
dominant-negative construct hPPARα6/29. C and D, transcriptionally
inactive dominant-negative PPARs do not block the inhibition of
STAT5b-stimulated transcription by wild-type PPAR. COS-1 cells were
transfected for 24 h with STAT5b reporter plasmid (pZZ1 in C, pT109-4Xntcp-Luc
in D) and pRL-CMV as an internal control. C, cells were cotransfected with 0.5
ng of pcDNAFlag-γ1 in the absence (▪) or the presence of 0.5 ng
(□) or 5 ng ( ) of pcDNAFlag-γ1 L466A/E469A. Beginning 24 h
after transfection, cells were treated with GH (500 ng/ml) and troglitazone (3
μM). The enhanced reporter activity upon cotransfection of
dominant-negative PPARγ reflects an overall decrease in R.
reniformis luciferase rather than a true increase in reporter activity
(C). Data presented are representative of two independent experiments. D,
cells were cotransfected with mPPARα (10 ng) in the absence (▪) or
the presence of hPPARα6/29 (100 ng) (□). Beginning 24 h after
transfection, cells were treated with GH (200 ng/ml) and Wy-14,643 (10 μM).
The enhanced reporter activity observed upon cotransfection of
dominant-negative PPARα (D) may reflect prevention of ligand-independent
inhibition by wild-type PPARα, although this has not been tested. In A
and C, n = 3, and in B and D, n = 2, mean ± S.D.
Firefly luciferase values are normalized to R. reniformis luciferase
values (A, C, and D). Relative firefly luciferase activities are shown in
B.
) of pcDNAFlag-γ1 L466A/E469A. Beginning 24 h
after transfection, cells were treated with GH (500 ng/ml) and troglitazone (3
μM). The enhanced reporter activity upon cotransfection of
dominant-negative PPARγ reflects an overall decrease in R.
reniformis luciferase rather than a true increase in reporter activity
(C). Data presented are representative of two independent experiments. D,
cells were cotransfected with mPPARα (10 ng) in the absence (▪) or
the presence of hPPARα6/29 (100 ng) (□). Beginning 24 h after
transfection, cells were treated with GH (200 ng/ml) and Wy-14,643 (10 μM).
The enhanced reporter activity observed upon cotransfection of
dominant-negative PPARα (D) may reflect prevention of ligand-independent
inhibition by wild-type PPARα, although this has not been tested. In A
and C, n = 3, and in B and D, n = 2, mean ± S.D.
Firefly luciferase values are normalized to R. reniformis luciferase
values (A, C, and D). Relative firefly luciferase activities are shown in
B.
PPARs Do Not Inhibit STAT5b-Regulated Transcription by Recruitment of Histone Deacetylases (HDACs). The transcriptional activity of a promoter DNA template is strongly influenced by the association of acetylated histones, which render the DNA more accessible to the cellular transcriptional machinery (Xu et al., 1999). One potential mechanism for inhibitory cross-talk between PPAR and STAT5b could therefore involve changes in the levels of bound histone acetylases and histone deacetylases, with the latter factors decreasing the extent of histone acetylation, leading to a decrease in gene transcription. To investigate whether PPARs modulate the acetylation status of histones or other factors associated with STAT5 target genes, COS-1 cells reconstituted with the PPARα and STAT5b pathways were treated with GH + Wy-14,643 in the presence or absence of the HDAC inhibitor trichostatin A (TSA). If PPARs inhibit STAT5 activity by increasing HDAC recruitment, the observed PPAR inhibition should be abolished in cells where TSA is used to block HDAC activity. Figure 6, however, shows that the inhibitory activity of PPARα is fully retained, even at 3 μM TSA, corresponding to a 10-fold higher TSA concentration than is required for HDAC inhibition (Minucci et al., 1997). The effectiveness of TSA was evidenced by its stimulation of firefly and R. reniformis luciferase activity (∼4-fold increase at 3 μM TSA; data not shown). This TSA-stimulated increase is not directly evident from the data shown in Fig. 6, where normalized firefly/R. reniformis luciferase activity ratios are presented. This finding rules out enhanced recruitment of HDACs as the mechanism of PPAR inhibition, but does not eliminate the possibility that PPAR increases the recruitment of other inhibitory factors to the STAT5-activated promoter.

Impact of TSA on PPAR-STAT5b cross-inhibition. COS-1 cells were cotransfected for 24 h with the STAT5b reporter pZZ1, pRL-CMV as an internal control, and expression plasmids encoding for STAT5b, GHR, and PPARα. Beginning 24 h after transfection, cells were treated with GH (500 ng/ml), Wy-14,643 (5 μM), and TSA at the indicated concentrations. Cell lysates from triplicate samples were prepared and assayed for luciferase activity 24 h later. Activities are expressed as firefly luciferase normalized by the reporter activity of a R. reniformis luciferase internal standard, mean ± S.D. Data presented are representative of two independent experiments.
PPARα Lacking the NH2-Terminal, Ligand-Independent AF-1 Trans-Activation Domain Does Not Inhibit STAT5b-Regulated Transcription. STAT5b inhibits transcription driven by the NH2-terminal AF-1 trans-activation domain of PPARα (Zhou and Waxman, 1999b). We therefore investigated whether this ligand-independent trans-activation domain is similarly required for PPAR to inhibit STAT5b. Figure 7A shows that full-length PPARα is capable of inhibiting the STAT5b reporter pZZ1, but that PPARαΔA/B, corresponding to PPARα with a deletion of the AF-1 region (also known as the A/B domain), does not. Ligand activation of PPARαΔA/B was confirmed in transfection studies using the PPAR reporter plasmid pHD(x3)luc (Fig. 7B). Thus, the AF-1 domain of PPAR is essential for the bidirectional inhibitory cross-talk between the PPAR and STAT5b signaling pathways.

Requirement of PPARα AF-1 domain for inhibition of STAT5b-regulated transcription. A, COS-1 cells were cotransfected for 24 h with the STAT5b reporter pZZ1, pRL-CMV as an internal control and expression plasmids for STAT5b, GHR, and mPPARα or the AF-1 deletion construct mPPARαΔA/B. Beginning 24 h after transfection, cells were treated with GH (500 ng/ml) and Wy-14,643 (5 μM). B, ligand-dependent activation of mPPARα and mPPARαΔA/B was demonstrated by cotransfection of the PPAR reporter p(HD)x3luc and stimulation with Wy-14,643 (5 μM) for 24 h. Cell lysates from triplicate wells were prepared and assayed for luciferase activity 24 h later. The apparent overall increase in reporter activity upon PPARαA/B cotransfection seen in this experiment was not observed in replicates of this experiment (data not shown). Activities are expressed as firefly luciferase normalized by the reporter activity of a R. reniformis luciferase internal standard, mean ± S.D., n = 3. Data presented are representative of three independent experiments.
Discussion
PPARs activate transcription of genes involved in essential cellular processes, such as adipocyte differentiation and fatty acid metabolism, in response to a variety of naturally occurring and synthetic ligands. PPAR activators include the hypolipidemic compound Wy-14,643, the insulin-sensitizer troglitazone, and a large number of industrial chemicals and environmental pollutants known as PPCs.
Previous studies have established that GH-activated STAT5b can inhibit PPARα-regulated transcription via the AF-1, ligand-independent trans-activation domain of PPARα (Zhou and Waxman, 1999a,b). The present study demonstrates that the cross-talk between these two signaling pathways is mutual, with ligand-activated PPAR capable of inhibiting transcription of a STAT5b-regulated reporter gene by up to ∼80%. This mutual inhibition provides a mechanistic explanation for the previous finding that several GH-regulated, sex-dependent liver proteins are down-regulated in rats treated with PPCs (Corton et al., 1998). STAT5b inhibitory cross-talk was demonstrated for both PPARα and PPARγ, indicating that both PPAR isoforms share common features required for STAT5b inhibition. STAT5b is a key intracellular mediator of the transcriptional effects of multiple cytokines, growth factors, and hormones (Kisseleva et al., 2002), including interleukins 2, 3, 5, and 7, erythropoietin and GH, which is a major stimulator of STAT5b activity in liver (Waxman et al., 1995). The inhibition of STAT5b by ligand-activated PPARα, outlined herein, also provides an explanation for the finding that Wy-14,643, a PPC and PPARα ligand, suppresses expression of the GH-regulated MUP-1 mRNA in wild-type mice, but not in PPARα-null mice (Corton et al., 1998). Consistent with a model of mutually inhibitory cross-talk, basal expression of PPARα-regulated peroxisomal and microsomal enzymes is elevated in livers of STAT5b-null mice, providing evidence in an in vivo model for the potential of STAT5b for inhibitory cross-talk toward PPARα target genes (Zhou et al., 2002). GH can also inhibit PPARα function by decreasing PPARα mRNA expression (Yamada et al., 1995). This latter inhibitory effect is STAT5b-dependent, as evidenced by the up-regulation of liver PPARα mRNA levels in STAT5b-null mice (Zhou et al., 2002).
PPARs inhibit the expression of a number of inflammatory genes, including those regulated by the transcription factors STAT1, activator protein-1, and nuclear factor-κB (Ricote et al., 1998). PPARγ inhibits transcription from a reporter containing eight isolated STAT1 binding sites in HeLa cells (Ricote et al., 1998), although interferon γ-activation of the same reporter, p36-8GASluc, was not inhibited by PPARγ in the present COS-1 cell studies (Fig. 1F). The apparent cell specificity of this inhibition suggests a requirement for a cell-specific factor, such as a coactivator. Shu et al. (2000) failed to observe an inhibitory effect of PPARγ agonists on the expression of tumor necrosis factor and interleukin-6, genes known to be controlled by activator protein-1, STAT, and nuclear factor-κB. However, another STAT1-regulated gene, matrix metalloproteinase 9, was inhibited, leading to the conclusion that PPARs may inhibit a subset of STAT1-regulated genes (Shu et al., 2000). We have observed PPAR inhibition of transcription from an isolated, multimerized STAT5 response element linked to a luciferase reporter gene, as well as transcription of a STAT5 response element in the context of an intact β-casein promoter (Fig. 1), suggesting that the promoter context of the STAT5b binding site is not critical to the inhibition by PPAR.
Nuclear receptor–STAT5 inhibitory cross-talk is not limited to PPAR, insofar as ligand-activated thyroid hormone receptor can inhibit prolactin-stimulated STAT5-dependent reporter gene activity (Favre-Young et al., 2000), whereas STAT5b can inhibit the transcriptional activity of thyroid hormone receptor (Zhou and Waxman, 1999b). Moreover, STAT5b inhibits ER-dependent activation of an estrogen-responsive gene promoter, whereas STAT5 induction of the β-casein promoter is repressed by ER (Stoecklin et al., 1999; Faulds et al., 2001). The mutually inhibitory STAT5b-PPAR cross-talk described here may thus serve as a more general example of how nuclear receptors cannot only regulate expression of their target genes but also may modulate the function of other, apparently distinct, signal transduction pathways. The precise mechanisms of inhibition may differ, however, depending on the receptor. In the case of ER, a direct physical interaction between ER and STAT5, mediated by the ER DNA-binding domain, may underlie the cross-talk (Faulds et al., 2001). An alternative inhibitory mechanism involves ER induction of cytokine signaling inhibitor SOCS2, which inhibits the tyrosine kinase JAK2, thereby inhibiting STAT5b tyrosine phosphorylation (Leung et al., 2003). This latter finding is consistent with the increase in nuclear STAT5b signaling seen previously in ERα-deficient female mouse liver (Sueyoshi et al., 1999). In contrast, in the case of PPAR, cotransfection of a dominant-negative PPARα or PPARγ did not reduce the STAT5b inhibitory effect of the corresponding wild-type PPAR. Moreover, prior exposure of the cells to a PPAR activator did not enhance the extent of STAT5b inhibition. Thus, in contrast to ER-STAT5 inhibition, the inhibition of STAT5b by PPAR does not involve a PPAR-inducible protein. Finally, the inhibitory cross-talk between STAT5 and PPAR can also be distinguished from the cross-talk between STAT5 and glucocortocoid receptor, which is inhibitory toward glucocorticoid receptor, but is synergistic toward STAT5 transcription (Stoecklin et al., 1999).
The AF-1 trans-activation domain of PPARα was found to be essential for the observed inhibition of STAT5b (Fig. 7). Previously, STAT5b was shown to inhibit transcription driven by the NH2-terminal ligand-independent AF-1 trans- activation domain of PPARα in a GAL4-linked chimera by approximately 80% (Zhou and Waxman, 1999b). Conceivably, the AF-1 domain may contain a binding site for a coactivator that is required for both STAT5b and PPAR-regulated transcription. A coactivator may become limiting to STAT5b as it is recruited by ligand-activated PPAR, and vice versa, it would be limiting to PPAR as it is used by STAT5b. Experiments using the well characterized coactivators SRC-1, p300 (Zhou and Waxman, 1999b), and GRIP1 (data not shown) do not, however, support a role for these particular factors in the inhibitory cross-talk. In the case of ER, a ternary complex of the coactivators GRIP1, CARM1, and p300 can synergistically coactivate ER when that nuclear receptor is expressed at very low levels (Lee et al., 2002). In unpublished experiments, we observed a 2- to 3-fold activation of PPAR reporter activity when the latter three coactivators were coexpressed with PPARγ. However, GH-activated STAT5b was still able to inhibit PPARγ transcriptional activity under these conditions. Moreover, STAT5b transcriptional activity was not affected by cotransfection of GRIP1, CARM1, or p300, either alone or in combination (data not shown). Together, these findings indicate that p300, GRIP1, and CARM1 are not limiting cofactors responsible for mutually inhibitory cross-talk between STAT5b and PPAR.
PPARγ and certain STATs are found at high levels in adipocytes and are up-regulated upon induction of differentiation of murine 3T3-L1 preadipocytes into adipocytes (Stephens et al., 1999; Harp et al., 2001; Waite et al., 2001). Adipocyte model studies have shown that STATs and PPARs can regulate each other either positively or negatively, depending on the cell-type and the STAT form. In primary rat preadipocytes, GH, potentially acting via STAT5b, inhibits differentiation by causing a 50% reduction in PPARγ protein levels (Hansen et al., 1998). STAT5 positively regulates expression of PPARγ during the initial phase of 3T3-L1 cell differentiation (Nanbu-Wakao et al., 2002). In contrast, STAT1 binds to regulatory sequences upstream of the PPARγ gene in 3T3-L1 cells, negatively regulating PPARγ protein expression, leading to a decrease in the activation in PPARγ-regulated genes (Hogan and Stephens, 2001). STAT1 is positively regulated by PPARγ in NIH-3T3 fibroblasts, with a differentiation-dependent up-regulation of STAT1 protein occurring downstream of PPARγ in a ligand-dependent manner (Stephens et al., 1999). A decrease in protein expression levels could potentially contribute to the mutually inhibitory cross-talk; however, under conditions of the inhibitory cross-talk, PPARγ, STAT5b, and tyrosine phosphorylated STAT5b protein levels remained constant (Fig. 3). The inhibitory cross-talk described here, together with the finding that STAT5 induces PPARγ expression early during the course of adipocyte differentiation (Nanbu-Wakao et al., 2002), suggests a mechanism whereby STAT5-induced PPARγ protein exerts feedback inhibition on STAT5 activity, thereby inhibiting further STAT5 stimulation of PPARγ expression.
HDACs remove acetyl groups from DNA-associated histones leading to DNA condensation and a consequent decrease in transcription. Thyroid hormone receptor induces a 60% decrease in STAT5-regulated transcription by direct interaction with STAT5 and by a mechanism proposed to alter recruitment of HDACs (Favre-Young et al., 2000). The experiments presented here used a transiently transfected COS-1 cell model, and therefore the transcription being studied is that of plasmid DNA that does not associate with histones in a native chromatin state. Nevertheless, up-regulation of STAT reporter gene transcription was seen in cells treated with the HDAC inhibitor TSA, suggesting that histone acetylation/deacetylation may indeed regulate STAT5b-dependent transcription in these cells. However, in contrast to the relief of thyroid receptor-STAT5 inhibitory cross-talk seen under conditions of TSA treatment (Favre-Young et al., 2000), no such effect on PPAR-STAT5b inhibition was seen (Fig. 6).
PPARα and PPARγ were shown to inhibit the constitutively active STAT5b1*6 in a manner indistinguishable from that of wild-type STAT5b (Fig. 2). Precisely how the H299R and S711F site-specific mutations of STAT5b1*6 (Onishi et al., 1998) lead to GH-independent activation of STAT5b is still undetermined. These mutations may enable STAT5b1*6 to become tyrosine phosphorylated, and thereby activated, via an unidentified tyrosine kinase distinct from JAK2. We can conclude that the inhibition of STAT5b by PPAR is not a GH-dependent process, although the mechanism of inhibition could still involve JAK2 in the case of GH-stimulated cells, or an unidentified tyrosine kinase in its absence. The inhibitory mechanism could also involve protein inhibitors of activated STATs, which display direct inhibitory interactions with STAT proteins (Shuai, 2000) and may also play a regulatory role in nuclear receptor function (Kotaja et al., 2000; Tan et al., 2002).
In conclusion, PPAR-STAT5b inhibitory cross-talk is mutual and has the potential to affect a broad range of STAT-dependent signaling pathways. This inhibition may be effected by environmental chemicals that activate PPARα and/or PPARγ, such as the chlorinated hydrocarbon trichloroethylene and the plasticizer hydrolysis product mono-2-ethylhexylphthalate, given their strong potential for PPAR activation (Maloney and Waxman, 1999; Hurst and Waxman, 2003). The observed inhibition of STAT5-regulated transcription by PPARα provides a mechanism for the previously observed PPARα-dependent decrease in expression of GH-activated genes in PPC-treated rats (Corton et al., 1998). Furthermore, the observation that PPCs inhibit GH-activated genes in rat liver validates our present in vitro findings based on the transiently transfected COS-1 cell model and exemplifies the cross-talk in a physiologically relevant system. Finally, the observation that PPARγ as well as PPARα inhibits STAT5b-regulated transcription raises the possibility that PPCs may inhibit STAT5 target genes in tissues other than liver. Further studies are required to fully understand the impact of this inhibitory cross-talk in vivo, under conditions of environmental or pharmacological exposure to these, and other PPAR activators.
Acknowledgments
We thank Dr. Yuan-Chun Zhou (Boston University) for preliminary studies, as well as the PPARα dominant-negative experiment included in Fig. 5. We also thank the many investigators who generously shared plasmid DNAs.
Footnotes
-
This research was supported in part by National Institutes of Health grant 5-P42-ES07381 and the Superfund Basic Research Center at Boston University.
-
ABBREVIATIONS: PPAR, peroxisome proliferator-activated receptor; STAT, signal transducer and activator of transcription; JAK, Janus tyrosine kinase; GH, growth hormone; GHR, growth hormone receptor; ER, estrogen receptor; PPC, peroxisome proliferator chemical; h, human; m, mouse; DMEM, Dulbecco's modified Eagle's medium; EMSA, electrophoretic mobility shift assay; HDAC, histone deacetylase; TSA, trichostatin A; CARM, coactivator-associated arginine methyltransferase; GRIP, glucocorticoid receptor interacting protein; Wy-14,643, pirinixic acid; CMV, cytomegalovirus.
- Received February 14, 2003.
- Accepted April 24, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}