


Abstract
Cisplatin resistant (CP-r) cells often show decreased uptake of cisplatin in association with reduced cell surface proteins and decreased endocytosis. In this report, two major [14C]carboplatin-binding proteins were identified as filamin and actin by photoaffinity labeling and mass spectrometry. Decreased expression of these two proteins was found in two different human CP-r cell lines (KB-CP20 and 7404-CP20), in comparison with their parental cell lines (KB-3-1 and BEL-7404), respectively. Disorganization of β-actin and filamin 250 and 90 was also detected in these CP-r cells by confocal microscopy. Transfection of a wild-type actin-enhanced green fluorescent protein (EGFP) expression vector into 7404-CP20 cells resulted in a nonfilamentous actin-EGFP distribution compared with a normal distribution in the cisplatin-sensitive BEL-7404 cells, suggesting that cytoskeletal organization is disturbed in the CP-r cells. The identification of actin and filamin as [14C]carboplatin-binding proteins and decreased expression and disorganization of several cytoskeletal proteins in CP-r cells provide a molecular and cellular basis for the known defect in endocytosis in these cells.
Cisplatin (cis-diamminedichloroplatinum II) has become a major chemotherapeutic agent for treatment of a wide spectrum of solid tumors; however, the ability of cancer cells to become resistant to the drug remains a significant impediment to successful chemotherapy. Intensive efforts have been made through biochemical characterization and cellular and molecular approaches to identify the basis of resistance and define genes that are involved in acquisition of cisplatin resistance (Fojo, 2001; Niedner et al., 2001; Deverman et al., 2002; Siddik, 2002; Gottesman et al., 2003).
One of the most consistent features observed in cisplatin-resistant (CP-r) cells both in vivo and in vitro is the reduced accumulation of the drug and other related chemicals (Shen et al., 2000). We have previously reported that a pleiotropic defect resulting in reduced accumulation of [3H]methotrexate, 73As3+ and 73As5+ in the CP-r cells was found to be associated with reduced expression of folate-binding protein and arsenic-binding proteins (Shen et al., 1998). We recently found defective fluid-phase endocytosis and mislocalization to the cytoplasm of several cell surface proteins, such as folate-binding protein and multidrug resistance-associated proteins in human CP-r cell lines (Chauhan et al., 2003; Liang et al., 2003); however, the cell biology and molecular bases of these features remain unclear.
To investigate the cellular and biochemical events involved in cisplatin resistance, photoaffinity labeling was applied in the current work to trace the pathway of [14C]carboplatin entering cells as a powerful tool to detect intracellular molecules with which platinum compounds might interact during the uptake process. Two [14C]carboplatin-binding proteins (actin and filamin) were identified as elements of the cytoskeleton involved in endocytosis, both of which were down-regulated in CP-r cell lines. In addition, reduced expression of β-tubulin, keratin, and dynamin 2 were also detected in the CP-r cells. Defects in the cytoskeleton, revealed initially by UV cross-linking of [14C]carboplatin, may be an important component of the pleiotropic phenotype found in CP-r cells.
Materials and Methods
Cell Lines and Cell Culture. Two populations of CP-r cell lines and their parental cell lines were studied: the human epidermoid carcinoma cell line KB-3-1 and its CP-r derivative, KB-CP20, and the human liver carcinoma cell line BEL-7404 and its CP-r derivative 7404-CP20. Both human CP-r cell lines were selected in several steps up to 20 μg of cisplatin/ml of medium as described previously (Shen et al., 2000). All cell lines were grown as monolayer cultures at 37°C in 5% CO2, using Dulbecco's modified Eagle's medium with 4.5 g/l glucose (Invitrogen, Carlsbad, CA), supplemented with l-glutamine, penicillin, streptomycin, and 10% fetal bovine serum (Cambrex Bio Science Walkersville Inc., Walkersville, MD).
Preparation of Enriched Plasma Membrane Proteins and Whole Cell Lysates. Membrane proteins were purified according to the method of Cornwell et al. (1987). In brief, 1 × 109 cells from each cell line were disrupted on ice by nitrogen cavitation with constant stirring for 40 min. Two cycles of sucrose gradient ultracentrifugation were followed. The purified membrane pellets were re-suspended in 3 ml of TSNa buffer (10 mM Tris-HCl, pH 7.45, 250 mM sucrose, 50 mM NaCl, and 1% aprotinin) and stored at -80°C until use. For whole cell lysates, cells were lysed in a lysis buffer [50 mM Tris-HCl, pH 8.0, 140 mM NaCl, 1.5 mM MgCl2, and 0.5% (v/v) Nonidet P-40], then centrifuged at 2000 rpm for 5 min to discard the nucleus.
Radiochemical Photoaffinity Labeling and Mass Peptide Spectrometric Mapping. [14C]Carboplatin was purchased from Amersham Biosciences Inc. (Piscataway, NJ). Ultraviolet irradiation was applied as described previously (Shen et al., 1998). In brief, a 200 W mercury lamp was used as a light source at a distance of 6 to 8 cm from microtubes containing a mixture of 100 μg of enriched plasma membrane proteins in 15 mM HEPES, 150 mM mannitol, 1 mM MnCl2, and 1 mM phenylmethylsulfonyl fluoride with 4 μCi of [14C]carboplatin on ice for 15 min. After UV cross-linking, the proteins were separated on SDS-PAGE, and enhanced with Fluoro-Hance (Research Products International, Mount Prospect, IL) and then exposed to X-ray film (BioMax MR; Eastman Kodak, Rochester, NY) at -80°C for the desired period of time. [14C]Carboplatin-labeled protein bands of interest were excised from a dried gel. After reduction and alkylation with iodoacetamide, the proteins were digested with trypsin and then analyzed on a Voyager DE RP mass spectrometer in the linear mode.
Antibodies and Immunoblotting. Antibodies directed to actin, filamins, dynamin, tubulin, keratin, and EEA1 were purchased from BD Biosciences PharMingen (San Diego, CA) unless otherwise noted. SDS-PAGE immunoblotting was carried out as described previously (Shen et al., 1998). Minigels were run as recommended by the manufacturer (Invitrogen). After electrophoresis, the gels were transblotted onto nitrocellulose membranes (Schleicher & Schuell, Keene, NH) at 4°C. Immunoreaction was performed with desired primary antibodies and secondary horseradish peroxidase-conjugated antibodies (Jackson ImmunoResearch Laboratories, West Grove, PA). Pierce enhanced chemiluminescence reagents (Pierce Chemical, Rockford, IL) were used for developing signals as described by the manufacturer.
Confocal Image Analysis. Cells were cultured in a Lab-Tek Chamber Slide (Nalge Nunc International, Naperville, IL), and fixed with 70% ETOH at -20°C for 15 min. The fixed cells were reacted with the primary antibodies and then followed by rhodamine-labeled anti-mouse IgG secondary antibody. Fluorescein isothiocyanate-phalloidin (Molecular Probes, Eugene, OR) was used for direct staining of actin on fixed cells. Immunofluorescent images of cells were monitored under a laser-scanning confocal microscope (Bio-Rad, Hercules, CA) at 600× magnification.
Actin-pEGFP Expression Vector and Transient Transfection. The expression vector for actin-pEGFP (actin fused to enhanced green fluorescent protein) was purchased from BD Biosciences Clontech, (Palo Alto, CA). Transient transfection of this vector into BEL-7404 and 7404-CP20 cells was performed using a LipofectAMINE reagent kit (Invitrogen) according to the manufacturer's instructions.
Results and Discussion
In this study, our goal was to trace the uptake pathway of a platinum compound in cells by comparing binding in wild-type cells and cells with a defect in uptake. [14C]Carboplatin was used in combination with UV cross-linking to determine whether there are any cellular proteins that differentially bind this radioactively labeled compound in sensitive and resistant cells. Figure 1 shows that two bands at Mr 230,000 (P230) and Mr 45,000 (P45) that are labeled with [14C]carboplatin are markedly reduced in both CP-r KB-CP20 and 7404-CP20 cells compared with their parental KB-3-1 and BEL-7404 cell lines, respectively. To identify these two [14C]carboplatin-binding proteins, these protein bands were excised and then analyzed by mass peptide spectrometric mapping as described under Materials and Methods. Figure 2, A and B, shows the mass spectra generated from tryptic digestion of these two peptides (P230 and P45) by MALDI-MS. The polypeptides were identified as filamin for P230 and actin for P45, respectively (Table 1).

[14C]Carboplatin labeling of proteins associated with a membrane pellet. Preparation of membrane proteins, UV cross-linking, and mass peptide spectrometric mapping were performed as described under Materials and Methods. Relative molecular mass is indicted to the left of the gel. KB-3-1, human epidermoid carcinoma cells; KB-CP20, CP-r cells selected in 20 μg/ml cisplatin from the KB-3-1 cells; 7404 (BEL-7404), human liver carcinoma cells; 7404-CP20, CP-r cells selected in 20 μg/ml cisplatin from the BEL-7404 cells.
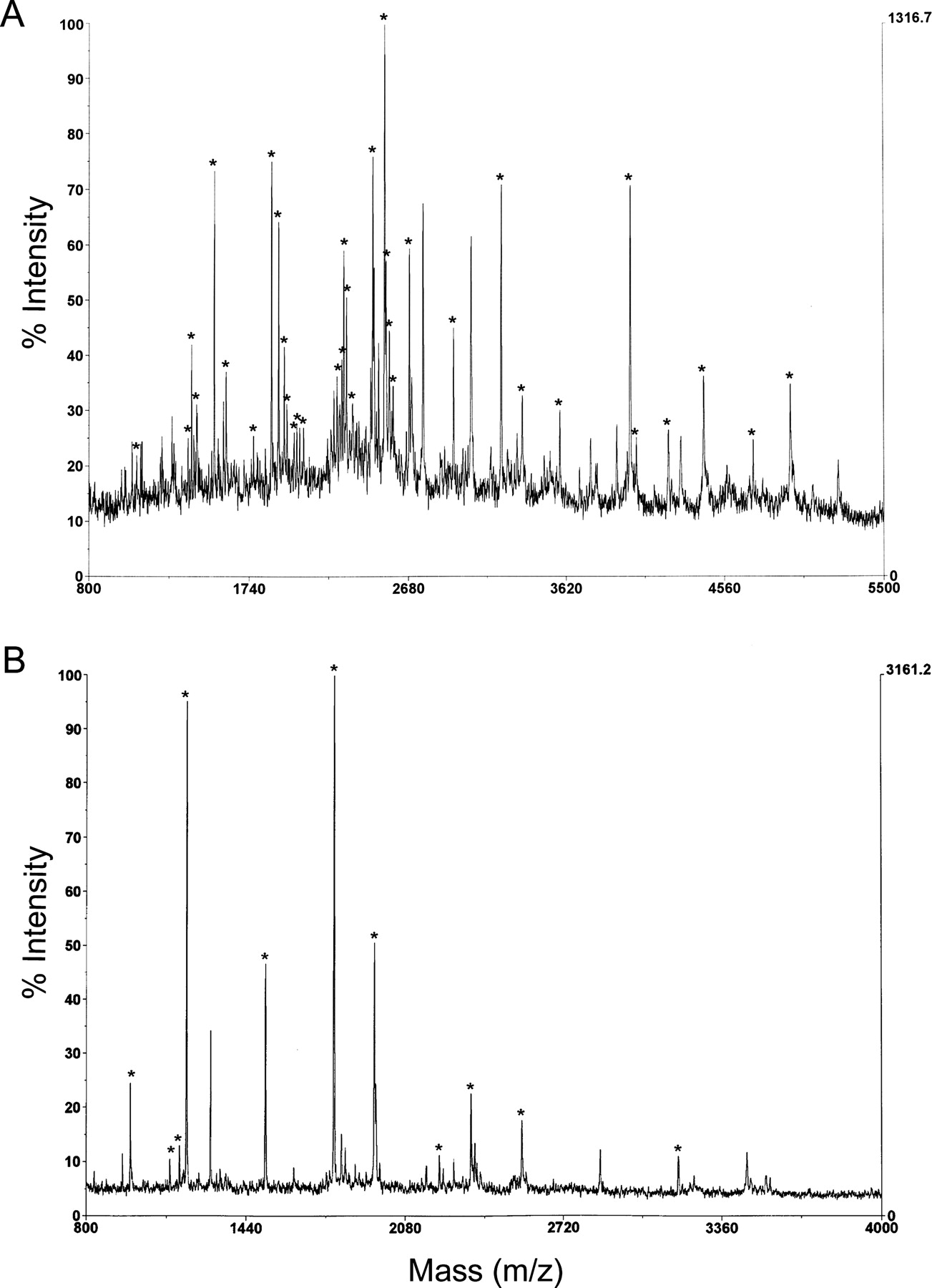
Mass spectra of the tryptic digests derived from P230 and P45. A, the mass spectrum of P230; B, the mass spectrum of P45. Peaks marked with an asterisk were assigned to the protein identified and are listed in Table 1. The average masses of the peaks in the mass spectra were matched within 200 ppm or better to the theoretical masses of filamin and β-actin, respectively. MALDI-MS analysis was performed as described under Materials and Methods.
Peptides identified in enzymatic digests of the P230 and P45 proteins by mass spectrometry
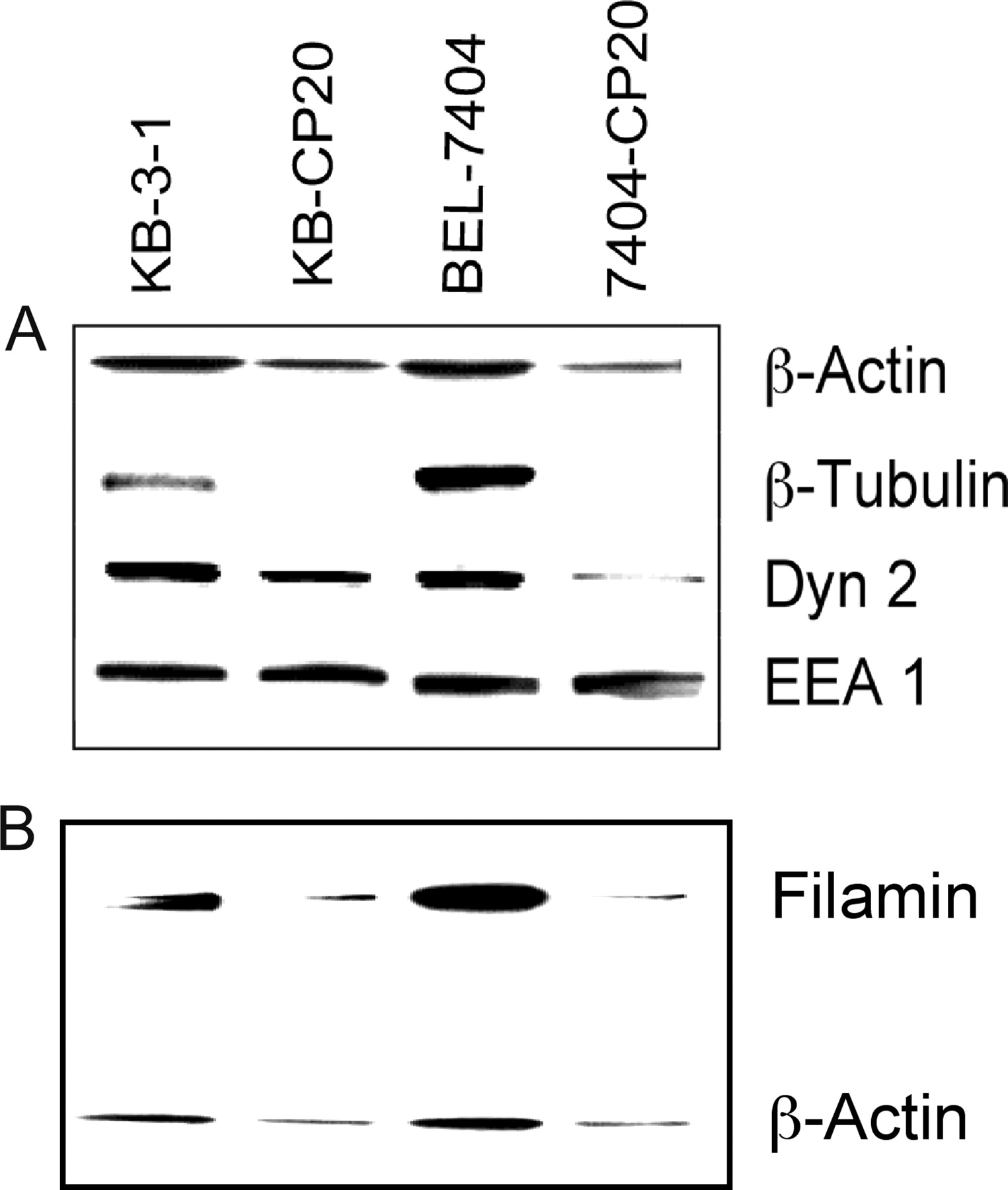
To further confirm the identification of these two [14C]carboplatin-binding proteins, immunofluorescence confocal microscopic observation was performed using fluorescent phalloidin for actin and monoclonal antibodies directed to filamin 90 and filamin 250. Figure 3 shows that expression of actin and filamins (Mr 90,000 and Mr 250,000) was decreased in the CP-r cells compared with their parental cisplatin-sensitive (CP-s) cells. Furthermore, the distribution and arrangement of β-actin, filamin 90, and filamin 250 were significantly changed in the CP-r cells as shown in Fig. 3 (bottom). A well defined intracellular microfilament fiber network was displayed in the CP-s cells (Fig. 3, top), whereas fewer structural microfilaments occurred in the CP-r cells (Fig. 3, bottom). The reduced expression of β-actin was also detected in the enriched membrane protein fractions from both KB-CP20 and 7404-CP20 cells by immunoblotting analysis using a specific monoclonal antibody as seen in Fig. 4A (top). Because actin and filamin are known to be involved in endocytosis and protein trafficking, we also determined whether there were changes in other cytoskeleton proteins that are involved in endocytosis. Figure 4 shows that expression of dynamin 2 was decreased in both KB-CP20 and 7404-CP20 cells compared with their parental CP-s cell lines, whereas expression of dynamin 3 was unchanged (data not shown). The expression of β-tubulin was dramatically reduced in both KB-CP20 and 7404-CP20 cells, as was β-actin, as seen in Fig. 4A; however, an early endosomal protein, EEA1, showed no significant changes in both CP-r cell lines compared with their parental CP-s cell lines. Decreased expression levels of β-actin and filamin were also detected in the whole cell lysates of these two pairs of CP-r cells (Fig. 4B) in agreement with observations by confocal microscopy (Fig. 3). Figure 5, A and B, shows the confocal analysis of keratin 5/8 in CP-s BEL-7404 cells (A), and CP-r 7404-CP20 cells (B). These keratin species were almost undetectable in the CP-r cells; therefore, expression of the cytoskeletal elements β-actin, filamins, β-tubulin, keratin 5/8, and dynamin 2 were down-regulated in CP-r cells.

Confocal microscopic fluorescence images of actin, filamin 90, and filamin 250. Actin was directly stained with fluorescein isothiocyanate-phalloidin. Monoclonal antibodies directed to filamin 90 and 250 were applied. Decreased expression and disorganization were observed in the CP-r cells as described in the text. BEL-7404 and 7404-CP20 cells are described in the legend to Fig. 1. Immunofluorescence images of cells were monitored under a laser-scanning confocal microscope (Bio-Rad) at 600× magnification.

Protein profiles by immunoblot analysis as visualized by enhanced chemiluminescence. Enriched plasma membranes (A) and whole cell lysates (B) were transblotted on nitrocellulose membranes after 10% SDS-PAGE as described under Materials and Methods. Monoclonal antibodies directed to β-actin, β-tubulin, dynamin 2, EEA1, and filamin 250 were applied. A, reduced expression of β-actin, β-tubulin, and dynamin 2 was seen in the CP-r cells; however, expression of EEA1 was not affected. B, two antibodies directed to β-actin and filamin 250 were immunoreacted on the same blot, showing reduction of these two proteins in the whole cell lysates of the CP-r cells. KB-3-1, KB-CP20, BEL-7404, and 7404-CP20 cells are described in the legend to Fig. 1.
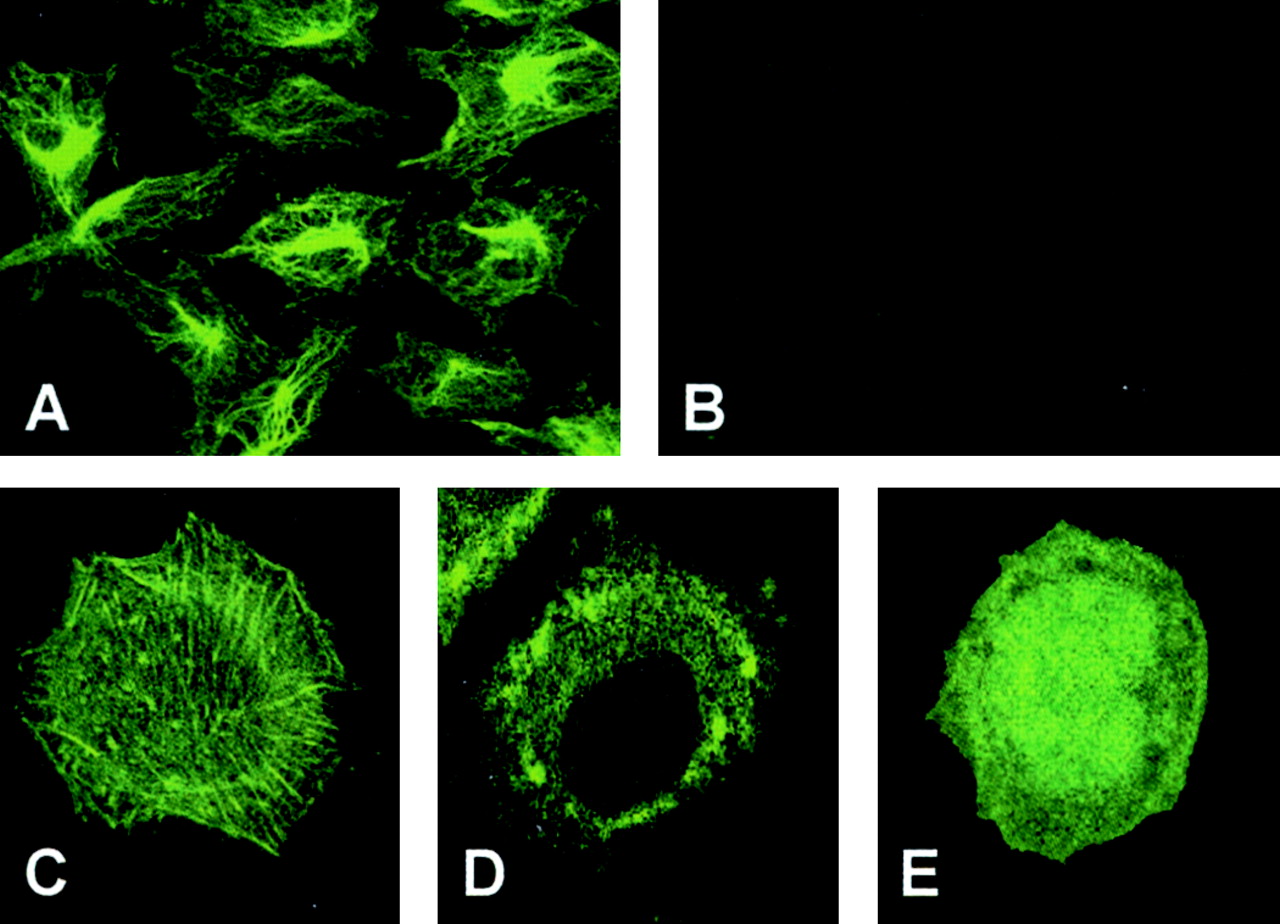
Confocal microscopic fluorescence images: A, BEL-7404; B, 7404-CP20, keratin 5/8. Monoclonal antibodies directed to keratin 5/8 were applied. The expression and structure of the keratin were almost undetectable in 7404-CP20 cells. C–E, expression and distribution of actin-EGFP in BEL-7404 (CP-s) cells, showing a well organized fiber network pattern and normal expression (C); 7404-CP20 (CP-r) cells, showing a cluster type of actin structure (D); and EGFP expression only in CP-s cells with a diffuse pattern (E). BEL-7404 and 7404-CP20 cells are described in the legend to Fig. 1. Confocal immunofluorescence imaging conditions were described in the legend to Fig. 3.
To determine whether the down-regulation of β-actin was the primary cause for disruption of the cytoskeleton, an expression vector for an actin-EGFP fusion protein was used in this study to trace the expression and distribution of exogenous β-actin in CP-s and CP-r cells. The fusion protein actin-EGFP was well expressed in CP-s BEL-7404 cells, and showed a fine fibrillar pattern as seen in Fig. 5C. Expression of EGFP alone in BEL-7404 cells gave a diffuse localization without the appearance of actin filaments (Fig. 5E); however, expression of the actin-EGFP fusion protein in the CP-r 7404-CP20 cells exhibited an entirely different picture, with clumps of fluorescence in the cytoplasm (Fig. 5D). These results indicate that disorganization and mislocalization of the cytoskeleton is caused not by poor expression of β-actin per se in the CP-r cells but by a more global defect in regulation of the cytoskeleton.
This study demonstrates that [14C]carboplatin binds to elements of the cytoskeleton, and points to a major defect in cytoskeletal regulation in at least two different CP-r cell lines. Because actin, filamin, and dynamin all play important roles in the endocytosis machinery (Qualmann et al., 2000; Sorkin, 2000; Pelkmans et al., 2002; Schafer, 2002), it is reasonable to speculate that the defective cytoskeletal system should contribute, at least in part, to reduced uptake of cisplatin, carboplatin, and methotrexate in CP-r cells. The exact mechanism by which cytoskeletal defects occur and what role these play in the development of cisplatin resistance in CP-r cells in patients still remain to be clarified.
Acknowledgments
We thank Dr. Susan Garfield for assistance with confocal microscopy and valuable discussions, and George Leiman and Joyce Sharrar for excellent editorial assistance.
Footnotes
-
This work was supported by the National Institutes of Health.
-
ABBREVIATIONS: CP-r, cisplatin-resistant; CP-s, cisplatin-sensitive; PAGE, polyacrylamide gel electrophoresis; EGFP, enhanced green fluorescent protein; MALDI-MS, matrix-assisted laser desorption ionization-mass spectrometry.
- Received January 27, 2004.
- Accepted June 10, 2004.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}