


Abstract
The structurally divergent agents gallamine and hexamethylene-bis-[dimethyl-(3-phthalimidopropyl)ammonium]dibromide (W84) are known to interact competitively at a common allosteric site on muscarinic receptors. Previous studies reported that the M2 selectivity of gallamine depended largely on the EDGE (172-175) sequence in the second outer loop (o2) and on 419Asn near the junction of o3 and the seventh transmembrane domain (TM7), whereas the selectivity of W84 depended on nearby residues 177Tyr and 423Thr. However, it has so far proven difficult to confer the high sensitivity for allosteric modulation of the M2 subtype onto the weakly sensitive M5 subtype by substituting these key residues. We now have found that M2 423Thr, not 419Asn, is the dominant residue in the o3/TM7 region for gallamine's high potency, although 419Asn can substitute for 423Thr in some contexts; in contrast, the presence of 419Asn reduces the potency of W84 in every context we have studied. In addition, the orientation of 177Tyr is crucial to high sensitivity toward W84, and it seems that the proline residue at position 179 in M5 (corresponding to M2 172Glu) may interfere with that orientation. Consistent with these observations, a mutant M5 receptor with these three key mutations, M5P179E, Q184Y, and H478T, showed dramatically increased sensitivity for W84 (>100-fold), compared with the wild-type M5 receptor. This same mutant receptor approached M2 sensitivity toward gallamine. Thus, gallamine and W84 derive high potency from the same receptor domains (epitopes in o2 and near the junction between o3 and TM7), even though these allosteric agents have quite different structures.
Muscarinic acetylcholine receptors (mAChRs) belong to the super family of G protein-coupled receptors and possess a highly conserved binding site (orthosteric site) for the endogenous agonist acetylcholine and for other traditional agonists or competitive antagonists (orthosteric ligands); this site is formed by the transmembrane domains (TM). The mAChRs are among the best known of a growing number of G protein-coupled receptors that possess another site (allosteric site) at which a second small ligand can bind, allowing ligand-ligand allosteric interactions at the external surface of the receptor (Christopoulos and Kenakin, 2002; Ellis, 2002). For ligands with significant selectivity, the M2 subtype is the most sensitive of the five subtypes of mAChRs to allosteric modulation (Ellis et al., 1991; Lee and el-Fakahany, 1991; Trankle et al., 1998; Ellis and Seidenberg, 2000). Although there is evidence for the existence of multiple muscarinic allosteric sites (Ellis and Seidenberg, 1989; Potter et al., 1989; Trankle and Mohr, 1997; Birdsall et al., 2001, Lazareno et al., 2002; Trankle et al., 2003), many muscarinic allosteric modulators seem to act at a “common allosteric site” (Ellis and Seidenberg, 1992; Trankle and Mohr, 1997).
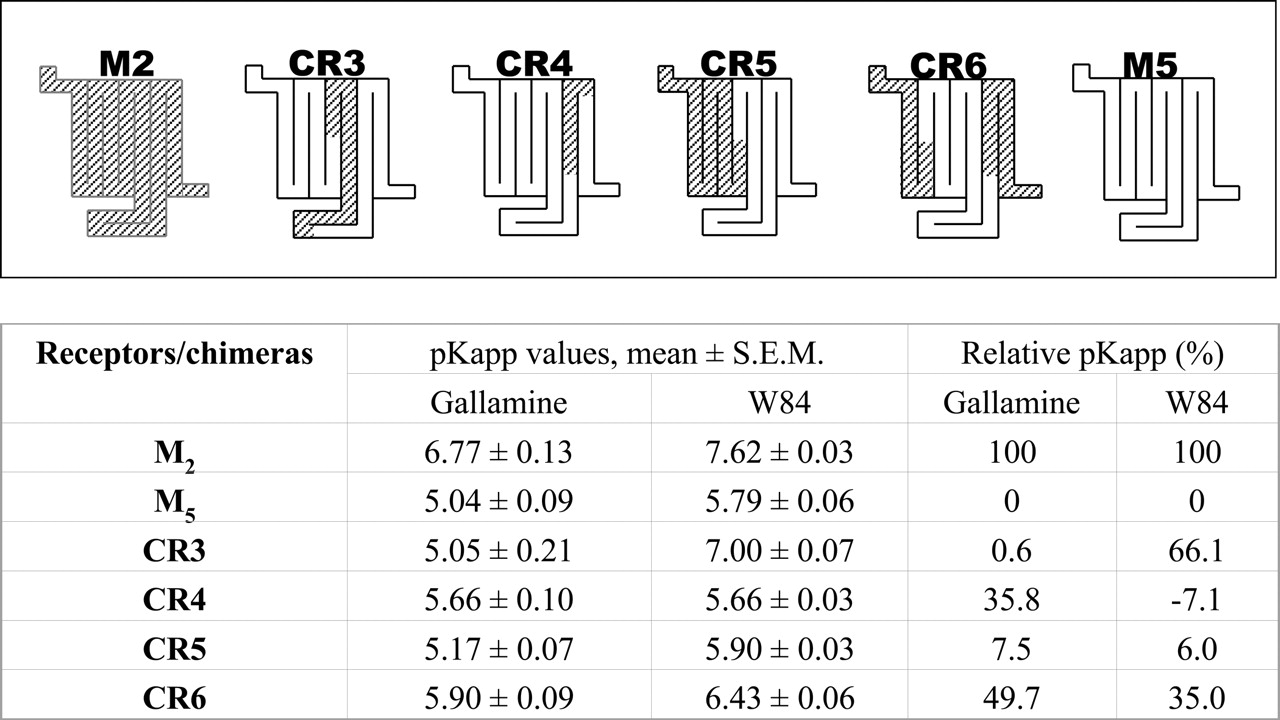
A number of investigations have attempted to identify the residues that comprise this common allosteric site. Initial studies in this area mutated either conserved residues or subtype-specific residues, and both approaches suggested that the binding site for allosteric ligands lies extracellular to the orthosteric site (Ellis et al., 1993; Leppik et al., 1994; Matsui et al., 1995). Most subsequent mutagenic studies have continued the subtype-specific approach after initial findings derived from M2/M1 and especially M2/M5 chimeric receptors. The structures of two allosteric ligands that have been investigated intensively, gallamine and W84, are shown in Fig. 1. The saturable type of antagonism seen with these agents in isolated organ preparations was the first evidence for a sensitivity of mAChRs for allosteric modulation (Lullmann et al., 1969; Clark and Mitchelson, 1976). Schematic diagrams of some M2/M5 chimeras are shown at the top of Fig. 2. The first chimeric studies investigated gallamine and implicated only a short segment of sequence in TM6 or o3 (i.e., CR4 in Fig. 2). However, subsequent studies with a number of other ligands (including W84) have implicated epitopes within a broad region containing o2 (CR3) and within TM7 (CR6) but not the TM6/o3 region (CR4) found for gallamine (Fig. 2). Furthermore, despite the lack of influence of the o2 loop on gallamine's potency in M2/M5 chimeras, studies in which M1 and M2 sequences were interchanged identified a role for an acidic region of o2 in gallamine's preference for M2 (the “EDGE motif” at M2172-175); it seemed that the essential acidic epitope in this region was shared by M5 (Leppik et al., 1994; Gnagey et al., 1999). Mutations of individual amino acids have suggested that the essential residue for gallamine in the CR4 chimera is M2 419Asn in o3, whereas the essential residues for W84 are M2 177Tyr in o2 and M2 423Thr in TM7 (Gnagey et al., 1999; Buller et al., 2002; Voigtlander et al., 2003). Thus, the binding selectivities of these two ligands are dependent on different but nearby residues, consistent with the finding that they seem to bind to a common site (Trankle and Mohr, 1997). However, despite this rather detailed knowledge concerning the residues that seem to be responsible for the subtype selectivities of these allosteric ligands, it has so far not been possible to replicate their high M2-like potency by the substitution of a few amino acids into the low-potency M5 structure.

Chemical structures of muscarinic allosteric modulators, gallamine (top) and W84 (bottom), used in this study. Note that although both agents are positively charged and interact competitively at an allosteric site on mAChRs, they have quite different structures. W84 is an elongated dicationic molecule, whereas the tricationic gallamine is rather small and compact.
Because the known essential residues are in such close proximity in the receptor structure, we believed that it would be useful to compare the effects of a detailed series of mutations in these regions on the allosteric affinities of gallamine and W84. We have found that the substitution of just three residues of the M2 sequence into the M5 receptor (M5P179E, Q184Y, and H478T) is sufficient to increase its sensitivity toward W84 by more than 100-fold, slightly exceeding the sensitivity of the M2 receptor for this ligand. This triply mutated M5 receptor also has greatly increased sensitivity for gallamine, although in this case, it did not reach the level of the wild-type M2 receptor.
Materials and Methods
Materials. Atropine sulfate, gallamine triethiodide, and polyethyleneimine were purchased from Sigma Chemical Co. (St. Louis, MO). W84 is commercially available from Tocris Cookson Inc. (Ellisville, MO). The orthosteric radioligand [3H]N-methylscopolamine chloride ([3H]NMS; 81 Ci/mmol) was obtained from PerkinElmer Life and Analytical Sciences (Boston, MA). Glass fiber filters and all other inorganic chemicals were from VWR International, Inc. (Bridgeport, NJ).

Schematic presentations of chimeric receptors used or mentioned in this study (top) and their sensitivity profiles for gallamine and W84 (bottom, table). Details of the constructions and the experimental data are given in Ellis et al. (1993) and Buller et al. (2002). Potencies of the allosteric agents are expressed as pKapp. To facilitate comparisons, these potency data have also been transformed to a scale of relative difference between the pKapp values of the M2 and M5 mAChR (see Materials and Methods). The diagrams are intended to illustrate the regions of the receptor in which human M5 sequence has been replaced by the homologous human M2 sequence. In brief, the M2 residues that have been inserted are (M2 numbering) the following: CR3, 156-300; CR4, 391-421; CR5, 1-155; and CR6, 1-69 and 391-466.
Mutagenesis, Receptor Expression, and Membrane Preparation. Human mAChRs were used throughout. Some receptor constructs have been described previously: chimera CR3 (Ellis et al., 1993); mutants M2T423H, M5H478T, and CR3H478T (Buller et al., 2002); and mutants M2EDGE-LAGQ, M2N419V, M2N419K, and M5V474N (Gnagey et al., 1999). All mutation primers and sequencing primers used in this study were synthesized in the Core Facility at Hershey Medical Center. Site-directed mutagenesis was carried out using the QuikChange kit from Stratagene (La Jolla, CA), as reported previously (Buller et al., 2002). Mutations were confirmed by sequencing in the Core Facility at Hershey Medical Center. Plasmids containing wild-type or mutant receptor genes were purified using the QIAGEN plasmid purification kit (Valencia, CA). Purified plasmid was transiently transfected into COS-7 cells using the Poly-Fect Transfection Reagent (QIAGEN). Approximately 48 h after transfection, cells were scraped into 5 mM sodium/potassium/phosphate buffer, pH 7.4 (4 mM Na2HPO4 and 1 mM KH2PO4; PB) and homogenized in the PB buffer on ice. After centrifugation at 50,000g for 30 min, membrane pellets were resuspended with a glass homogenizer in 5 mM PB and stored in aliquots at -70°C. Protein concentrations were determined using the Advanced Protein Assay reagent from Cytoskeleton Inc. (Denver, CO).
[3H]NMS Saturation Binding Assays. All binding assays, including dissociation assays (below), were carried out in 5 mM PB, pH 7.4, at 25°C. To determine receptor expression levels and binding affinities for [3H]NMS, membranes were incubated with 6 concentrations of [3H]NMS (ranging from 3 to 1000 pM) in duplicate in a final volume of 1 ml for 30 min. The reactions were terminated by filtration onto 32 glass-fiber filters (S & S, Keen, NH) pretreated with ice-cold 0.1% polyethyleneimine solution and followed with two washes with ice-cold 40 mM PB. Radioactivity from membranes trapped on filter discs was determined by liquid scintillation counting. Data were fitted to a one-site hyperbolic binding curve with the Prism 4.0 software (GraphPad Software, Inc., San Diego, CA). Non-specific binding was determined in the presence of 3 μM atropine at each concentration point.
[3H]NMS Dissociation Assays. Dissociation assays were set up and conducted as described previously (Ellis and Seidenberg, 2000; Buller et al., 2002). In brief, receptors (quantity of membrane protein estimated to produce approximately 2000 dpm in total binding) were first labeled with a saturating concentration of [3H]NMS (1 nM) for 30 min in 1 ml. Dissociation of [3H]NMS was initiated by the addition of atropine (3 μM, in a final volume of 2 ml), with or without the indicated concentrations of allosteric modulators in duplicate and was terminated by filtration after a period of time, which is typically set between 2 and 3 times the standard half-time of [3H]NMS dissociation (determined in the presence of atropine but in the absence of any allosteric modulator), up to a maximum of 120 min for M5 receptor and some of its mutants.
Dissociation assays were set up such that the delay of [3H]NMS dissociation was mediated by the binding of the allosteric modulator to an allosteric site, separate from the orthosteric site at which [3H]NMS binds. The true dissociation rate constant (k0) was determined in the presence of 3 μM atropine without the allosteric modulator, and the apparent dissociation rate constant (kobs) was determined in the presence of both atropine and the allosteric modulator. The ratios of kobs/k0 were then plotted against the logarithms of the concentrations of allosteric modulator (X), and the resulting curve was fitted (using Prism 4.0 software) to the three-parameter logistic function kobs/k0 = [Bottom + (Top - Bottom)/(1 + 10(X - logEC50)), where Top (constrained to 1.0 in curve-fitting) and Bottom refer to the upper and lower plateaus of the sigmoidal curve. The curve represents the effect of allosteric delay of [3H]NMS dissociation and corresponds to the occupancy curve of the tested allosteric modulator at the [3H]NMS-occupied receptor (Ellis and Seidenberg, 1992; Lazareno and Birdsall, 1995). Curve-fitting yielded EC50 values, which correspond to equilibrium dissociation constants of the allosteric modulator on the [3H]NMS-occupied receptor, Kapp. For convenient comparison of effects of a given mutation on the binding of gallamine versus W84 on the same scale, we transformed pKapp values (negative logarithm of Kapp values) into a percentage degree of the difference between M2 and M5 receptors: relative pKapp (%) = 100% × [(pKapp - pKapp M5)/(pKapp M2 - pKapp M5)]. On this scale, M2 would score 100% and M5 would score 0%; a mutation that reduced the pKapp of M2 by one third of the M5/M2 span would score 67%, and a mutation that increased the pKapp of M5 by one third of that span would score 33% (Table 1).
Effects of mutations in o3/TM7 region on allosteric interactions
Affinities (expressed as pKapp) for gallamine and W84 are shown, along with half-times of dissociation of [3H]NMS in the absence of allosteric modulators. To facilitate comparisons, the affinit y data have also been transformed to a scale of relative difference between the p Kapp values of the human M2 and M5 receptors (see Materials and Methods). Data shown represent mean ± S.E.M. from three or more assays for pKapp values and six or more assays for half-times. Receptors were expressed in COS-7 cell membranes.
Results
In this study, we systematically mutated a series of amino acid residues in o2 and o3/TM7 regions of M2 or M5 receptors, carried out [3H]NMS dissociation assays to measure allosteric modulator potencies (apparent binding affinity, Kapp), and compared their individual roles in the binding and subtype selectivities of gallamine and W84. We used dissociation assays for reasons that have been presented previously (Ellis, 1997; Ellis and Seidenberg, 2000; Buller et al., 2002). In brief, a change in the dissociation rate of the labeled ligand is a purely allosteric phenomenon that is not confounded by any concomitant competitive interaction. In addition, the marked slowing of the kinetics of the labeled ligand that is caused by these allosteric modulators does not alter the rapid equilibration of the allosteric modulator, whereas the slow kinetics of the labeled orthosteric ligand may be problematic in equilibrium (or pseudoequilibrium) experiments (M. Seidenberg and J. Ellis, unpublished data).
The half-times of [3H]NMS dissociation from each receptor construct are reported in Tables 1 to 3 and are consistent with previous data (Ellis et al., 1993) that M2 receptors have short half times (fast dissociation) and M5 receptors have long half-times (slow dissociation). Buller et al. (2002) reported initially that the M2T423H mutant exhibits a significantly slower rate of [3H]NMS dissociation compared with the wild-type M2 receptor. We also have observed that our M2 mutants, which included histidine at that position (and only these M2 mutants), were more than 3-fold slower than the wild-type receptor. Furthermore, the converse mutation in any of the M5 backgrounds, or in the CR3 chimera, induced significantly faster dissociation compared with the parent receptor.
Receptor constructs with combined mutations in o2 and o3/TM7 regions
Affinities (expressed as pKapp) for gallamine and W84 are shown, along with half-times of dissociation of [3H]NMS in the absence of allosteric modulators. To facilitate comparisons, the affinity data have also been transformed to a scale of relative difference between the p Kapp values of the human M2 and M5 receptors (see Materials and Methods). Data shown represent mean ± S.E.M. from three or more assays for pKapp values and six or more assays for half-times. Receptors were expressed in COS-7 cell membranes.
Residue M2423Thr Is Equally Important for both Gallamine and W84 Binding. As illustrated in Fig. 2, gallamine has been found to be unique among many tested allosteric ligands in that its potency is sensitive to epitopes found in both CR4 and CR6 but not in CR3; W84, like many other ligands, is sensitive to epitopes in CR3 and CR6 but not in CR4 (Ellis and Seidenberg, 2000; Buller et al., 2002). The residue in CR4 that confers specificity toward gallamine seems to be 419Asn, whereas 423Thr, which is included in CR6 but not in CR4, has been identified as the critical residue for the selectivity of W84 (Gnagey et al., 1999; Buller et al., 2002). The proximity of residues 419 and 423 (Fig. 3A) led us to examine gallamine's potency at the M2T423H mutant. This substitution of the M5 residue reduced gallamine's potency significantly; additionally, the converse mutation, M5H478T, significantly enhanced gallamine's potency (Fig. 4). To facilitate comparison of the changes in the affinities of gallamine and W84 that are caused by receptor modification, we expressed these changes as percentages of the M2-M5 span of potency for each ligand on a logarithmic scale (as described under Materials and Methods). On this scale, the M2T423H mutation reduces gallamine's potency by 31% and reduces W84's potency by 45%; the converse mutation in M5 enhances gallamine's potency by 41% and enhances W84's potency by 31% (Table 1). Similar degrees of enhancement and reduction in affinities have been reported previously for W84, dimethyl-W84, and diallylcaracurine V (Buller et al., 2002), indicating the importance of residue 423Thr to these structurally different muscarinic allosteric modulators.
The Presence of Asparagine at M2419 or M5474 Leads to Complex Effects on Gallamine's Potency but Always Reduces the Potency of W84. As stated above, residue 419Asn in the o3 loop has been found to be important for gallamine's subtype selectivity (Gnagey et al., 1999). In brief, residues 523Lys in M3 receptors and 474Val in M5 receptors are associated with significantly lower potency of gallamine, and acidic residues in M1 and M4 receptors are associated with slightly higher potency of gallamine. Furthermore, the K523N mutation in M3 increased both the pKapp value toward gallamine and also the negative cooperativity between gallamine and NMS (Krejci and Tucek, 2001). All of these data have led to the conclusion that residue 419Asn is very important in the binding of gallamine and in its ability to modulate the orthosteric site.
In light of the impact and proximity of 419Asn and 423Thr in M2, we examined the effects of mutating each of these residues to their M5 counterparts. The reverse mutations were also studied in the M5 receptor, both individually and in concert. In Fig. 5A, it can be seen that the mutations M5H478T and M5V474N produce similar effects, because each construct exhibits significantly higher sensitivity for gallamine than does the wild-type M5 receptor. However, the enhancements are clearly not additive; indeed, the sensitivity of the double mutant is slightly lower than that of the M5H478T single mutant. Figure 5B shows a different pattern of effects toward W84. As reported previously (Voigtlander et al., 2003), the M5H478T mutation significantly increases sensitivity toward this ligand; however, the M5V474N mutation actually reduces sensitivity toward W84. These results are summarized in Table 1, along with data from additional single mutations at the M2419Asn site. Inserting a positively charged lysine residue produces almost as much decrement in sensitivity toward W84 as toward gallamine. However, W84 has significantly higher potency at the M2N419V mutant than it has at the wild-type M2 receptor, which is in agreement with the converse result in M5. Thus, the potency of W84 is attenuated by the presence of asparagine, whether at M2419 or at M5474. Perhaps more surprising is the finding that gallamine also has moderately higher potency at the M2N419V mutant; nonetheless, this effect is consistent with the attenuation by asparagine of the beneficial effect of the threonine mutation in the M5 receptor (compare M5V474N, H478T to M5H478T in Table 1).

Schematic presentations of o2 and o3 and their adjacent TM domains of the M2 mAChR, shown with partial sequences of the outer loops and the adjacent TM domains of the mAChR family. The boundaries of transmembrane domains of the human mAChR sequences were estimated by manually aligning the sequences to bovine rhodopsin, for which a high-resolution crystal structure has been obtained (Palczewski et al., 2000). A, o3; B, o2.

Gallamine's potency is reduced by the T423H mutation in the o3/TM7 region of the M2 receptor and increased by the reverse mutation H478T in the M5 receptor. The modulation of the rate of dissociation of [3H]NMS from the receptors was determined as described under Materials and Methods, and the data were fitted to the model given under Materials and Methods. Points represent the mean ± S.E.M. from four to eight experiments.

Effects of mutations near the junction of o3/TM7 in the M5 receptor on the potencies of gallamine (A) and W84 (B). The M2 (far left) and M5 (far right) curves from Fig. 4 are shown as broken lines in A and in subsequent figures for convenient comparison. Experiments were conducted and analyzed as in Fig. 4. Points represent the mean ± S.E.M. from three to five experiments.
Residue M2177Tyr Is More Important for W84 than for Gallamine. The mutation M2Y177Q (Fig. 3B) has been found to reduce the affinities for the NMS-occupied receptor of bis(ammonio)alkane-type (such as W84) and caracurine V-type allosteric modulators; affinities are enhanced by the reverse mutation M5Q184Y (Voigtlander et al., 2003). Our experiments with W84 in receptors with mutations at this residue were in good agreement with these previous data. The mutation in M2 reduced the potency of W84 precipitously, decreasing more than 80% on our M2/M5 scale; the converse mutation in M5 was less dramatic but still increased the potency of W84 by 28% (Table 2). In contrast, gallamine's potency is reduced much less (15%) by the M2Y177Q mutation and is increased even less (7%) by the M5Q184Y mutation (Fig. 6 and Table 2).
Mutations in o2 region
Affinities (expressed as p Kapp) for gallamine and W84 are shown, along with half-times of dissociation of [3H]NMS in the absence of allosteric modulators. To facilitate comparisons, the affinit y data have also been transformed to a scale of relative difference between the p Kapp values of the human M2 and M5 receptors (see Materials and Methods). Data shown represent mean ± S.E.M. from three or more assays for p Kapp values and six or more assays for half-times. Receptors were expressed in COS-7 cell membranes.

Gallamine's potency is somewhat reduced by the Y177Q mutation in the M2 receptor and is not increased by the reverse mutation Q184Y in the o2 loop of the M5 receptor. M2 and M5 curves are included for comparison. Experiments were conducted and analyzed as in Fig. 4. Points represent the mean ± S.E.M. from six to nine experiments.
It was noted previously that the potency of W84 is greatly enhanced in a chimeric receptor that includes M2 sequence in the o2 region (CR3; Fig. 2), compared with its potency at the wild-type M5 receptor. Figure 7 shows that this greatly increased potency is predominantly caused by the presence of the tyrosine residue in that chimeric receptor, because the construct that contains the other 144 residues of M2 sequence but lacks the tyrosine actually has lower sensitivity for W84 than the wild-type M5 receptor. In studies with W84, the substitution of glutamine for tyrosine at this position, whether in the wild-type M2 receptor or in the CR3 chimeric receptor, produces a greater decrease in potency than any other single-residue mutation in this study (Table 2).

The 177Tyr residue is critical to the enhanced action of W84 in the chimera CR3 relative to the M5 receptor. M2 and M5 curves are included for comparison. Experiments were conducted and analyzed as in Fig. 4. Points represent the mean ± S.E.M. from three to four experiments.
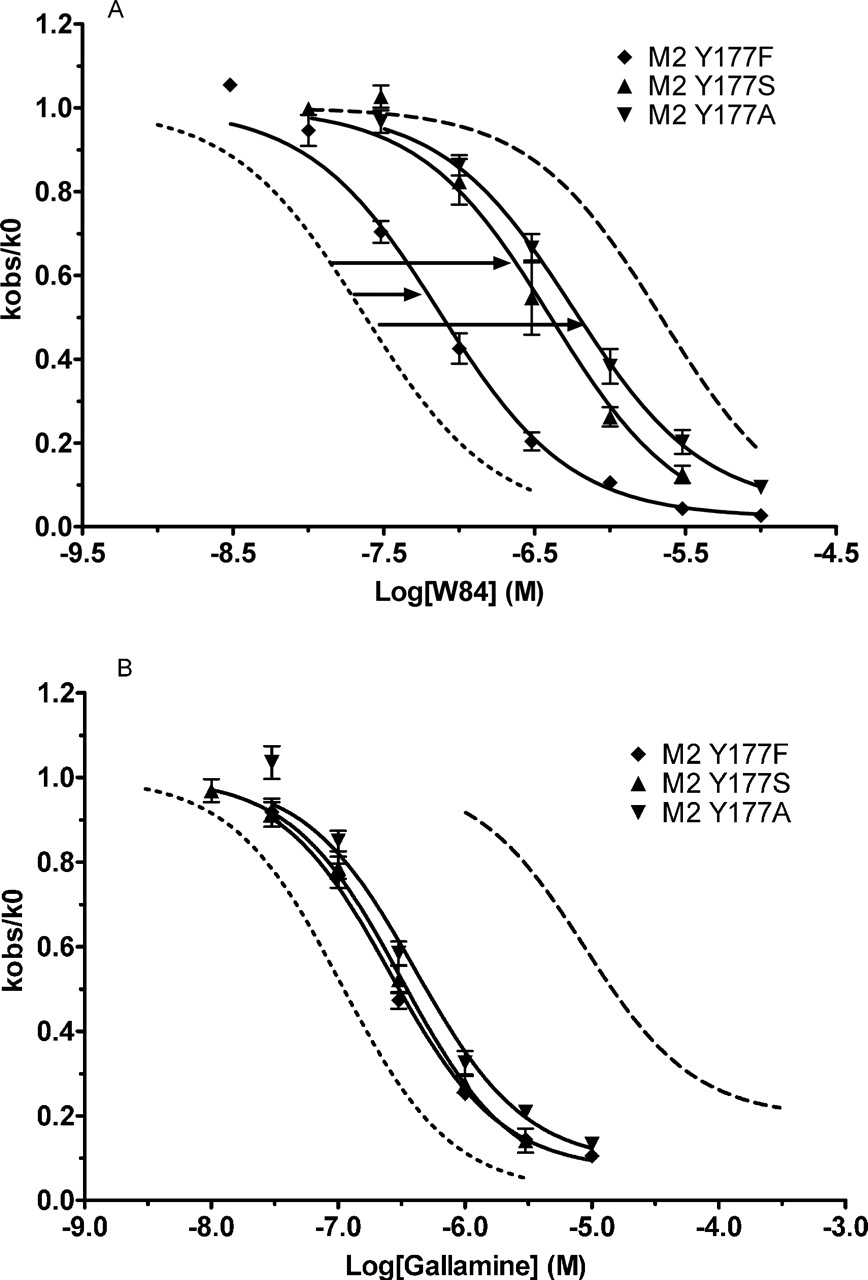
The Impact of M2177Tyr on W84 Binding Is Caused by the Residue's Aromatic Nature. Molecular modeling studies have suggested that aromatic rings in diallylcaracurine V or W84 interact with M2177Tyr via π-π interactions (Voigtlander et al., 2003). To test this suggestion experimentally, we systematically replaced M2177Tyr with residues containing different functional side chains. Thus, in the M2 receptor, we replaced the tyrosine with phenylalanine to retain the aromatic ring, with serine to retain the hydroxyl group, or with alanine to retain neither characteristic. Figure 8 shows that the M2Y177A and M2Y177S mutations reduced the receptor's sensitivity toward W84 almost as much as the M2Y177Q mutation did (Table 2). However, the M2Y177F mutation caused a much smaller reduction in sensitivity toward W84, as would be expected if a π-π interaction is the important feature at that residue. It is interesting that all of the mAChR subtypes except for M5 have an aromatic residue at this position—tyrosine in M1 and M2 and phenylalanine in M3 and M4 (Fig. 3B).

Nonaromatic substitutions at residue 177Tyr of the M2 receptor strongly reduce potency of W84. The wild-type tyrosine was replaced by phenylalanine, serine, or alanine. The pKapp values for these mutant curves were 7.13, 6.39, and 6.25, respectively. M2 (pKapp value, 7.66) and M5 (pKapp value, 5.66) curves are included for comparison. Experiments were conducted and analyzed as in Fig. 4. Points represent the mean ± S.E.M. from three to seven experiments.
Glutamic Acid Residues within the EDGE Motif in M2-o2 Exert Differential and Complex Effects on the Potencies of Gallamine and W84. Leppik et al. (1994) first reported that mutation of EDGE to LAGQ (the corresponding M1 sequence) significantly reduced gallamine's potency at the M2 receptor. This was somewhat surprising, because the chimera CR3 includes the EDGE motif from the M2 receptor but did not increase gallamine's potency relative to M5 (Ellis et al., 1993). The explanation for these divergent results seemed to be that the M5 receptor also has acidic residues in the corresponding motif (PLDE) that were also important (perhaps equally important) for gallamine binding (Gnagey et al., 1999; sequence alignments are shown in Fig. 3B). However, the roles of individual residues in the EDGE motif have not been investigated; therefore, in this study, we have investigated single amino acid mutations at strategic sites in this region. In particular, we mutated residues within the EDGE motif to the corresponding M1 residues and/or M5 residues to evaluate their roles in the binding and subtype selectivities of gallamine (and W84). The rationale for using M1 as well as M5 residues is that the M1 receptor is the only subtype with no acidic residues in the corresponding motif (LAGQ); the M5 receptor maintains two acidic residues in the corresponding motif (PLDE), even though M5 has the lowest sensitivity toward either gallamine or W84 among the mAChR subtypes.

Effects of mutations at the glutamate residues of the EDGE motif on potencies of gallamine (A) and W84 (B). The EDGE motif in the M2-o2 region was replaced by M1 sequence in whole (EDGE-LAGQ) or in part, as indicated. M2 and M5 curves are included for comparison. Experiments were conducted and analyzed as in Fig. 4. Points represent the mean ± S.E.M. from three to nine experiments.
As expected from previous studies (Gnagey et al., 1999), the mutation EDGE-LAGQ, which removed all three acidic residues, reduced gallamine's pKapp by 42%; the same mutation also reduced W84's potency, albeit to a lesser extent (26%; Fig. 9 and Table 2). In the EDGE-LAGQ (M2/M1) mutation, residue M2174Gly is retained, but the other three individual mutations remove three negatively charged residues. Previous studies have found that gallamine's potency at the M4 receptor is unaffected by the PDNQ-LAGQ mutation, suggesting that the aspartate residue (corresponding to M2173Asp) does not play a major role (Gnagey et al., 1999), leaving the glutamic acid residues (M2172Glu and 175Glu) for further investigation. The mutations M2E172L and M2E175Q reduced gallamine's pKapp by 10 and 17% and W84's pKapp by 7 and 12%, respectively. Furthermore, the combined mutations (M2E172L, E175Q) produced reductions in potency of gallamine and W84 that were similar to the EDGE-LAGQ mutation (Fig. 9 and Table 2). In the M5 receptor, the residue corresponding to M2175Glu is conserved (as M5182Glu), but the residue corresponding to M2172Glu is a proline (M5179Pro). The mutation M2E172P reduced the potencies of gallamine and W84 to similar extents, approximately 17% (Table 2).
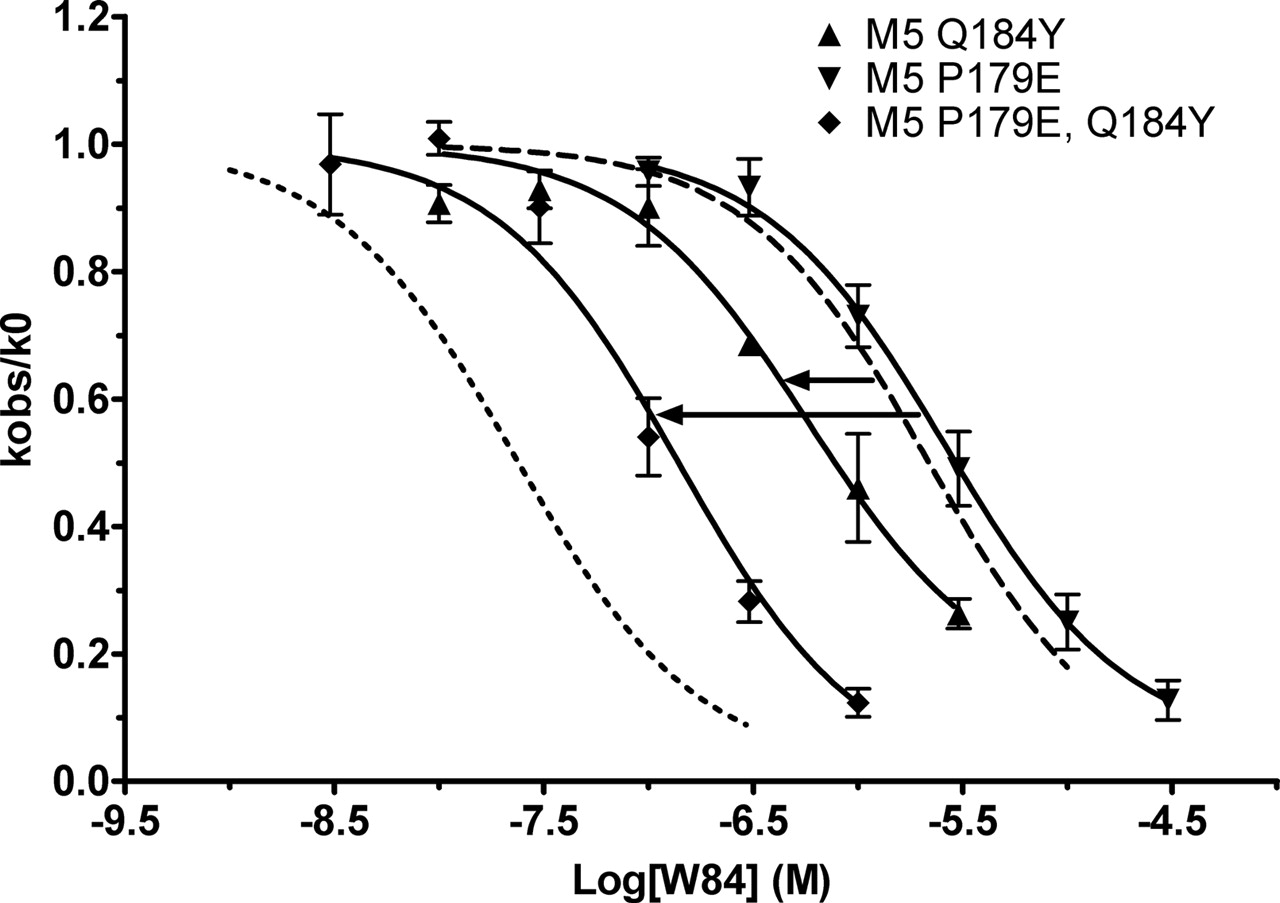
M2E172P Mutation Alters the Conformation of M2177Tyr. One mechanism by which the M2E172P mutation could exert a reduction in the potency of W84 would be via a change in the conformation of nearby residue 177Tyr that might disrupt the π-π interaction between 177Tyr and W84. This mechanism would also explain why the introduction of the tyrosine residue in M5Q184Y might be under an unfavorable conformation for W84 binding, which in turn would explain why the enhancement caused by this mutation is so much less than the decrement observed with the M2Y177Q mutation (Table 2). If this explanation is correct, then additional mutations upstream of the inserted tyrosine might facilitate the interaction with W84. To test this possibility, we introduced the M2 glutamate into M5 in place of the proline. The M5P179E mutation had virtually no effect by itself on W84 binding, but it significantly potentiated the ability of Q184Y to increase the potency of W84 from 28 to 60% greater than its potency at M5 (Fig. 10 and Table 2). This mechanism is unlikely to apply to gallamine, however, because gallamine's potency is so much less sensitive to M2177Tyr. Indeed, although the M5P179E mutation significantly enhanced the potency of gallamine, there was no potentiation when the Q184Y mutation was added (compare gallamine at M5P179E versus M5P179E, Q184Y in Table 2).

The mutations P179E and Q184Y in the o2 loop of the M5 receptor synergistically enhance potency of W84. M2 and M5 curves are and analyzed included for comparison. Experiments were conducted as in Fig. 4. Points represent the mean ± S.E.M. from three to four experiments.
Effects of Combining Mutations in the o2 and o3/TM7 Regions. The preceding studies have identified four epitopes that distinguish the M2 receptor from the M5 receptor and are involved in the binding and/or subtype selectivities of gallamine and W84. These are the M2 residues 419Asn in the o3 loop, 423Thr at the top of TM7, and, within the o2 loop, 172Glu of the EDGE motif and 177Tyr. The chimera CR3 possesses the entire o2 loop from the M2 receptor (Fig. 2) but does not contain the 419Asn or 423Thr of M2. Therefore, we examined the allosteric effects of gallamine and W84 on CR3 receptors with the single or combined mutations V474N and H478T. For W84, the effects of the additional mutations were similar in CR3 and M5. The histidine-to-threonine mutation enhanced potency, although in a somewhat less than additive manner, and resulted in a potency that was close to that of W84 at M2 (compare CR3 H478T in Table 3 with M5H478T and CR3 in Tables 1 and 3, respectively). This result is similar to that of a previous study (Buller et al., 2002). The valine-to-asparagine mutation had a negative effect in every case, and, again, the effect was similar to the results obtained in M5; the reductions in potency of W84 were approximately additive, in that the presence of the asparagine reduced the gains in potency caused by the CR3 or H478T manipulations by approximately the same amount that it reduced the potency of W84 in M5 itself (Tables 1 and 3). The effects of these mutations on the potency of gallamine were more complex. The M2 sequence in CR3 did not enhance gallamine's potency very much, but it acted synergistically with the histidine-to-threonine mutation to yield a supra-additive gain in potency (compare Figs. 5A and 11, and Tables 1 and 3). The valine-to-asparagine mutation, by contrast, was less than additive with the effect of CR3; however, the inclusion of this asparagine did not reduce the CR3-threonine synergy.
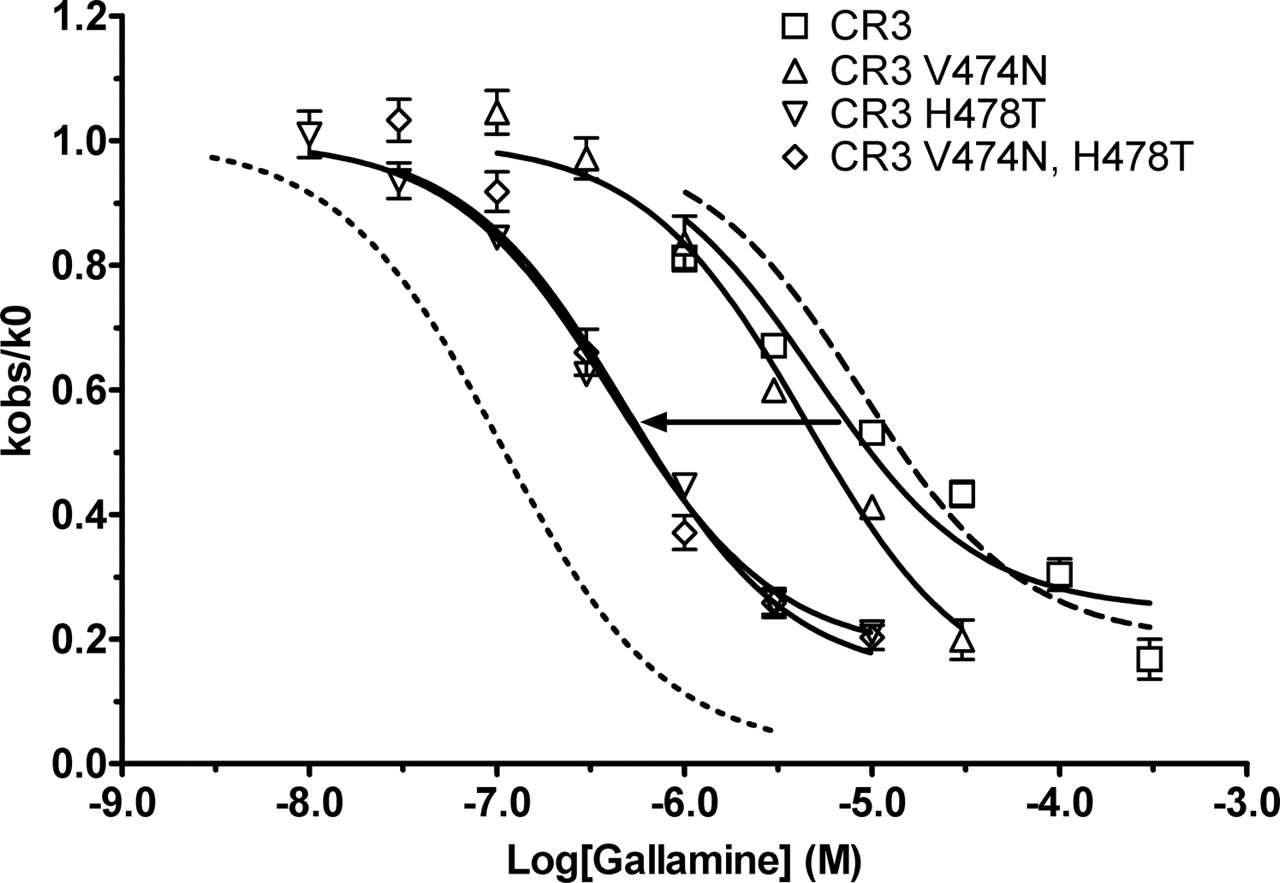
Effects of mutations in the o3/TM7 region of the CR3 chimera on potency of gallamine. M2 and M5 curves are included for comparison. Experiments were conducted and analyzed as in Fig. 4. Points represent the mean ± S.E.M. from three to four experiments.
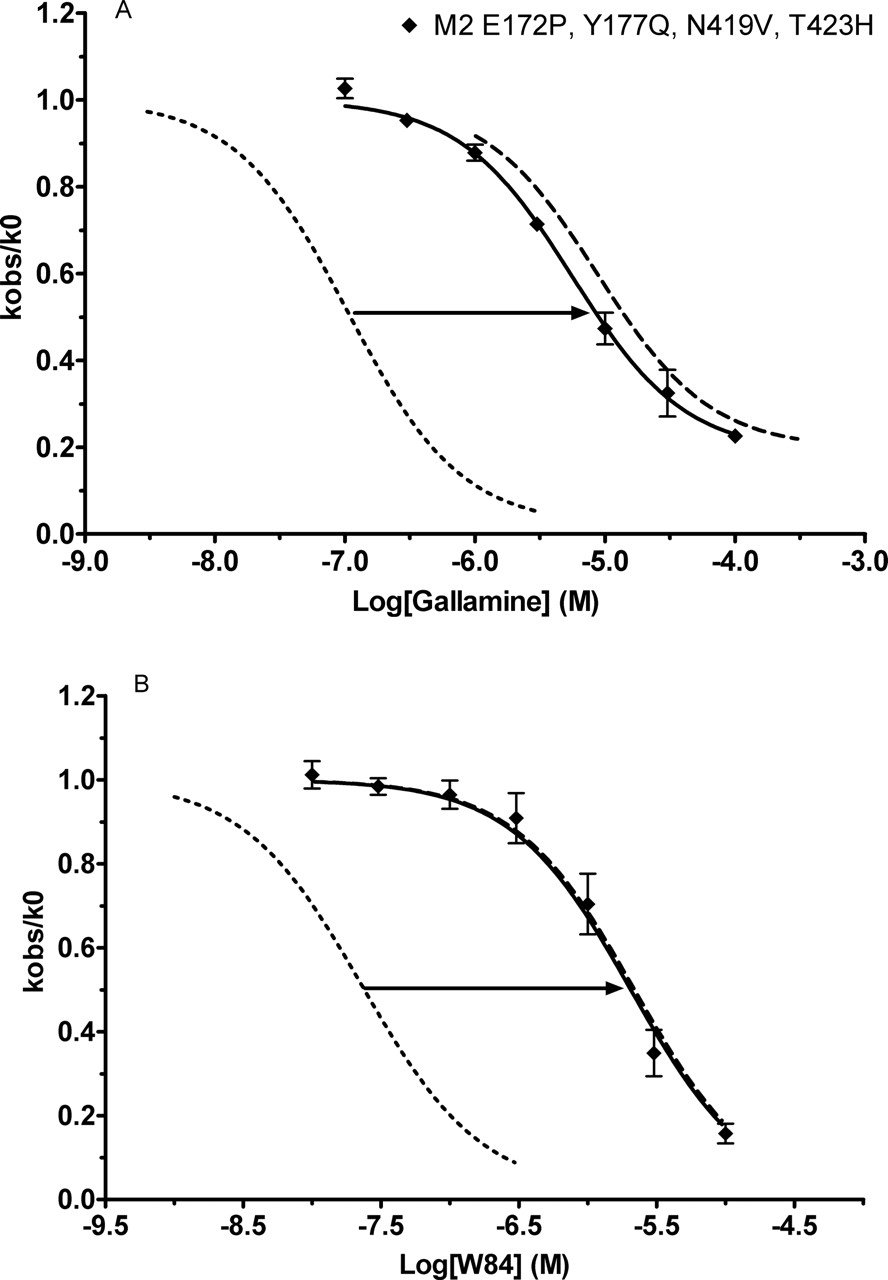
Compared with the M5V474N, H478T mutant (Table 1), the CR3 V474N, H478T mutant (Table 3) increased gallamine's pKapp by 0.66 log unit and that of W84 by 0.95 log unit. A major difference between these two mutants is the o2 loop of the M2 receptor. From the studies presented above, we expected that the P179E and Q184Y mutations were dominant in producing the effects of the o2 loop in the CR3 chimeric constructs. Therefore, we created two quadruple mutants, the M5P179E, Q184Y, V474N, H478T mutant and the converse construct in M2, in which the corresponding four residues were replaced by their M5 counterparts. These mutations in the M2 receptor reduced its sensitivities toward gallamine and W84 dramatically, nearly to M5 levels (Fig. 12 and Table 3). Confirming the importance of these residues, the M5 quadruple mutant showed significantly higher sensitivity toward gallamine than M5 (Fig. 13A and Table 3). Remembering the frequently deleterious effects of the o3 419Asn, we also tested the triple mutant M5P179E, Q184Y, H478T; it showed slightly higher sensitivity toward gallamine than the quadruple mutant, even higher than the CR3H478T mutant, increasing 74% on the relative scale between M5 and M2 pKapp. The potency of W84 was enhanced to an even greater extent than that of gallamine in these triple and quadruple M5 mutants; indeed, the sensitivity of the triple mutant for W84 actually surpassed that of M2 (Fig. 13B and Table 3).

Combined mutations in the o2 loop and the o3/TM7 region of the M2 receptor dramatically reduce potencies of gallamine (A) and W84 (B). M2 and M5 curves are included for comparison. Experiments were conducted and analyzed as in Fig. 4. Points represent the mean ± S.E.M. from three to six experiments.
Discussion
The aim of this investigation was to examine the differences between the epitopes involved in the subtype selectivity of gallamine and W84, which represent structurally divergent muscarinic allosteric agents. Before this study, the existing data suggested that although these ligands interacted at a common allosteric site (Trankle and Mohr 1997), the residues responsible for their subtype selectivities were distinct. W84 has recently been thoroughly characterized and shown to require 177Tyr in the o2 loop and 423Thr at the top of TM7 for its high potency at the M2 receptor, relative to M5 (Voigtlander et al., 2003). The M2/M5 selectivity of gallamine has been attributed to M2419Asn in the o3 loop and to a negatively charged sequence (EDGE) in the M2-o2 loop (Leppik et al., 1994; Gnagey et al., 1999), but individual residues in this region of the o2 loop had not been thoroughly characterized. Here, we have found that gallamine and W84 are approximately equally dependent on M2423Thr for high potency, that the orientation of M2177Tyr is crucial to the selectivity of W84 and can be modulated by nearby upstream residues, that gallamine's high potency apparently relies more on the negative charge in o2 than on the aromatic nature of M2177Tyr, and that the role of M2419Asn is more complex than earlier studies had suggested.
The evidence pointing to M2419Asn in gallamine's subtype selectivity originated from studies in which a smaller or larger segment of the M5 receptor was replaced with M2 sequence (see CR4 and CR6 in Fig. 2). The chimera CR6 includes both M2419Asn and M2423Thr, whereas CR4 contains only M2419Asn. Because the two chimeric constructs showed similar sensitivity toward gallamine and the larger segment included the smaller one completely, it was assumed that the essential residue resided within the smaller segment. Subsequent investigations found that the sensitivity of the M5 receptor (and M3 as well) toward gallamine was significantly enhanced when they contained an asparagine residue at the position corresponding to M2419 (Gnagey et al., 1999). However, inspection of Fig. 2 reveals that the larger substitution of CR6 leads to higher sensitivity toward gallamine than does the smaller substitution of CR4, in much the same way that the double mutant M5V474N, H478T compares with M5V474N (Fig. 5A), probably indicating that the slightly higher gallamine potency at CR6 (compared with CR4) is significant and that it represents the nonadditive effects of V474N and H478T on gallamine's potency. Overall, it seems that this asparagine residue is beneficial to sensitivity toward gallamine when the threonine is absent but not when that threonine is present. In other words, M2419Asn plays a minor role or even a negative role in M2 wild-type receptors; however, when M2423Thr is removed or replaced with histidine, then M2419Asn becomes important for gallamine binding. These data indicate that the role of 419Asn is more complicated and less important in the binding of gallamine to the M2 subtype than earlier studies had suggested. In the case of W84, the data are not so complex, because the presence of that asparagine residue reduces W84's potency whether it is in the M2 or M5 context.

Combined mutations in o2 loop and the o3/TM7 region of the M5 receptor dramatically enhance potencies toward gallamine and W84. The quadruple mutation M5P179E, Q184Y, V474N, H478T (and the triple mutation without V474N) were constructed and assessed for allosteric actions of gallamine (A) and W84 (B). M2 and M5 were curves are included for comparison. Experiments conducted and analyzed as in Fig. 4. Points represent the mean ± S.E.M. from three to four experiments.
Similar to the situation at M2419Asn, the residues at the position corresponding to M2423Thr are unique in each subtype of mAChRs. As noted above and in previous studies, the histidine in the M5 subtype is associated with low sensitivity for caracurine derivatives and dimethyl-W84 (as well as gallamine and W84), whereas the serine in the M4 subtype seems to be functionally equivalent to the threonine residue (Buller et al., 2002). Thus, it seems that both M2419Asn and M2423Thr (and their corresponding residues in other subtypes) are important in generating subtype selectivity, but the threonine residue is the dominant feature of the o3/TM7 region for the subtype selectivity of both gallamine and W84. The importance of these two positions in the receptor may be related to the fact that, if the region containing these residues is helical, their side groups will be adjacent on the same face of the helix. In addition, both 419Asn and 423Thr seem to be involved in mechanisms of cooperativity between NMS and several allosteric ligands, especially when comparisons are drawn between the M2 and M3 receptor subtypes (Krejci and Tucek, 2001; Jakubik et al., 2005).
In the o2 region, there seems to be no doubt that M2177Tyr is the most important residue in the subtype-selective binding of W84, as reported previously (Voigtlander et al., 2003). In the present study, we have shown that the dramatic increase in sensitivity toward W84 that is seen in the chimeric receptor named CR3, relative to M5 sensitivity, is completely lost if just that tyrosine is excluded from the 145 amino acid exchange (Fig. 7 and Table 2). Subsequent mutations at position 177 also supported the suggestion that there is a π-π interaction between W84 and the tyrosine phenol ring, because phenylalanine was a much better substitute for that tyrosine than serine or alanine (or glutamine).
With regard to the potency of W84, we have suggested previously that the proline at position 179 in M5 might interfere with the proper alignment of the tyrosine residue in M5 mutants that include the Q184Y mutation (Voigtlander et al., 2003), and that now seems to be the case. That is, whereas the P179E mutation leaves the sensitivity toward W84 almost unaffected, relative to M5, it greatly potentiates the effect of the tyrosine insertion at position 184 (Fig. 10 and Table 2). For gallamine, it seems that the negative charge within the EDGE motif is the most important factor in o2 and that the glutamates at positions 172 and 175 are the most important residues. However, the chimera CR3 contains the glutamate corresponding to M2172Glu instead of proline yet does not have much better sensitivity toward gallamine than M5 itself, suggesting that there must also be features within the M2 sequence of CR3 that neutralize the expected gain in sensitivity; indeed, gallamine has significantly higher potency for M5P179E, Q184Y than it has for CR3.
When mutations in the o2 and o3/TM7 regions are combined, some additional small synergies may be noted. For gallamine, the substitution of threonine for histidine within the CR3 context produces a greater gain in potency than the corresponding mutation in the M5 receptor itself (i.e., H478T). On the other hand, insertion of that threonine into the M5P179E, Q184Y produces just an additive effect (Table 3). It is as though the threonine residue relieves the unidentified negative influence in CR3 (referred to above). For W84, the situation is reversed, with the histidine-to-threonine mutation and the CR3 substitution being essentially additive, but the insertion of the threonine into M5P179E, Q184Y yielding a greater than additive effect; indeed, this triple mutant has slightly higher sensitivity than M2 itself toward W84. The source of these small inter-regional synergies is not immediately apparent. In any event, this is the first report of high potency of a muscarinic allosteric ligand (i.e., W84) being so fully recreated by three discrete mutations in the low-potency background of the M5 subtype.
A number of studies have suggested that the o2 and o3 loops (plus several residues predicted to lie in the extracellular ends of TMs 6 and 7) can modulate the conformation of mAChR structure. Liu et al. (1995) found that a threonine at the position corresponding to M2423 interfered with proper helix-helix packing in some recombinant receptor constructs (Buller et al., 2002). We have noted that the presence or absence of a threonine at this same position modulates the kinetics with which NMS binds to the receptor (see Results and half-times in Tables 1 and 3). Extending the observations of Spalding et al. (1995, 1997) at the M5 receptor, Ford et al. (2002) found that homologous mutations near the o3/TM6 junction constitutively activate all of the five mAChR subtypes. In o2, endogenous antibodies that bind to this loop region can activate the M2 receptor (Goin et al., 1999; Baba et al., 2004), and a monoclonal antibody fragment generated against a smaller peptide that still includes the EDGE motif displays inverse agonist activity (Peter et al., 2004).
In summary, we now have a more detailed explanation of the source of the relative affinities of the allosteric agents W84 and gallamine for the NMS-bound forms of the M2 and M5 mAChRs. The affinities of these two ligands benefit from a common feature, namely the threonine residue found near the junction of the o3 and TM7 in the M2 receptor. In addition, W84 interacts with a tyrosine residue in the o2 loop; the orientation of this residue is crucial to allow for an optimal π-π interaction with the ligand and may be modulated by nearby residues. The potency of gallamine seems to be influenced more by nearby negative charges than by the aromatic tyrosine residue. Thus, the subtype-selective features of the common allosteric site seem to reside in these two extracellular regions of the receptor.
Footnotes
-
This work was supported by Public Health Service grant R01-AG05214 (to J.E.) and by a grant of the Deutsche Forschungsgemeinschaft (to K.M.)
-
ABBREVIATIONS: mAChR, muscarinic acetylcholine receptor; CR, chimeric receptor construct; Kapp, apparent binding affinity (i.e., the affinity for [3H]NMS-occupied receptors in this study); ko, true dissociation rate constant; kobs, observed (apparent) dissociation rate constant; NMS, N-methylscopolamine chloride; o2, the second outer (extracellular) loop; o3, the third outer loop; PB, sodium-potassium phosphate buffer; TM, transmembrane domain; W84, hexamethylene-bis-[dimethyl-(3-phthalimidopropyl)ammonium]dibromide.
- Received April 24, 2005.
- Accepted June 3, 2005.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}