


Abstract
We have previously reported that the reducing agent dithiothreitol (DTT) strongly increases thermally induced activity of the transient receptor potential vanilloid receptor-1 (TRPV1) channel. Here, we show that exposure to oxidizing agents also enhances the heat-induced activation of TRPV1. The actions of sulfhydryl modifiers on heat-evoked whole-cell membrane currents were examined in TRPV1-transfected human embryonic kidney 293T cells. The sensitizing effects of the membrane-permeable oxidizing agents diamide (1 mM), chloramine-T (1 mM), and the copper-o-complex (100:400 μM) were not reversed by washout, consistent with the stable nature of covalently modified sulfhydryl groups. In contrast, the membrane-impermeable cysteine-specific oxidant 5,5′-dithio-bis-(2-nitrobenzoic acid) (0.5 mM) was ineffective. The alkylating agent N-ethylmaleimide (1 mM) strongly and irreversibly affected heat-evoked responses in a manner that depended on DTT pretreatment. Extracellular application of the membrane-impermeable reducing agent glutathione (10 mM) mimicked the effects of 10 mM DTT in potentiating the heat-induced and voltage-induced membrane currents. Using site-directed mutagenesis, we identified Cys621 as the residue responsible for the extracellular modulation of TRPV1 by reducing agents. These data suggest that the vanilloid receptor is targeted by redox-active substances that directly modulate channel activity at sites located extracellularly as well as within the cytoplasmic domains. The results obtained demonstrate that an optimal redox state is crucial for the proper functioning of the TRPV1 channel and both its reduced and oxidized states can result in an increase in responsiveness to thermal stimuli.
The capsaicin receptor transient receptor potential vanilloid receptor-1 (TRPV1) is a sensory neuron-specific ion channel that plays an important role in thermal nociception and inflammatory hyperalgesia (Caterina et al., 1997). This channel can be activated by vanilloid compounds, low pH, and noxious heat, and its function is modulated by a wide range of endogenous and exogenous agents (for review, see Planells-Cases et al., 2005). Among them, redox-active substances have recently been shown to modulate TRPV1 receptor activity (Vyklicky et al., 2002; Jin et al., 2004); however, it is not yet clear whether these effects are mediated through the direct modification of sulfhydryl groups present in the TRPV1 channel protein complex. As demonstrated by earlier radioligand binding studies, both the reducing and oxidizing agents diminished the apparent affinity and positive cooperativity characteristic of [3H]resiniferatoxin binding (Szallasi and Blumberg, 1993; Szallasi et al., 1993). From these studies, it has become apparent that cooperation between the TRPV1 subunits is, at least in part, subject to redox modulation. Also, our earlier studies demonstrated that the sulfhydryl redox agent dithiothreitol (DTT) strongly potentiates both the native and recombinant rat TRPV1 channel when applied at millimolar concentrations to the external solution under intact whole-cell conditions. Since the effects of DTT were almost immediate, dose-dependent, and reversible, the contribution of extracellular cysteine residues within the putative pore-loop region of TRPV1, Cys616, Cys621, and Cys634 has been proposed (Vyklicky et al., 2002). In an attempt to identify the mechanisms involved in the redox modulation of TRPV1 channel, the effects of an oxidizing agent, copper-o-phenantroline [Cu(II)-1,10-phenantroline; Cu:Phe], on heat-induced membrane currents were studied in human embryonic kidney (HEK) 293T cells expressing rat TRPV1. However, a detailed examination of these effects has shown that this oxidant exerts its strong inhibitory effect mainly through a steric block of the TRPV1 channel rather than by redox modulation (Tousova et al., 2004). The recent demonstration that oxidizing agents such as thimerosal decrease the capsaicin-induced TRPV1 activity by oxidation of extracellular sulfhydryl residues (Jin et al., 2004), namely, the cysteine Cys621, seemed to be in line with this idea. However, the results of the latter study were obtained with oxidizing agents coapplied with capsaicin, which may cast doubt as to whether these compounds may or may not affect the capsaicin molecule directly, since the alkyl side chain of capsaicin is considered to be susceptible to enzymatic oxidation (Boersch et al., 1991). Therefore, it seems that a more advantageous strategy could be to explore the effects of redox-active substances on the heat-dependent mode of TRPV1 receptor activation.
The goal of this investigation was to characterize the effects of redox-active substances on the TRPV1 receptor and to determine the mechanisms by which these reagents affect the heat-induced activity of this channel. The results presented here indicate that both reducing and oxidizing agents enhance the heat-evoked currents mediated by these channels. We also show that the effects of reducing agents are at least partially attributable to modifications taking place at specific thiol-sensitive sites on the extracellular side of the TRPV1 receptor pore-forming loop.
Materials and Methods
Cell Culture and Transfection. HEK293T cells (SD 3515; American Type Culture Collection, Manassas, VA) were cultured in OPTI-MEM I (Invitrogen, Paisley, Scotland) supplemented with 5% fetal bovine serum. Cells were plated on to dishes coated with collagen at a density ∼180,000 cells cm-2. 293T cells were transfected transiently with 300 to 400 ng/dish recombinant plasmid DNA encoding wild-type or mutant rat TRPV1 in pcDNA3 vector (wild type kindly provided by D. Julius, San Francisco, CA) using either the LipofectAMINE 2000 (Invitrogen) or Magnet-assisted transfection (IBA GmbH, Göttingen, Germany) method according to the manufacturers' protocols. To identify the transfected cells in the electrophysiological experiments, DNA plasmid encoding green fluorescent protein (GFP) in pQBI 25 vector (TaKaRa, Kyoto, Japan) was cotransfected at a concentration of 400 ng/dish. Transfected cells were replated onto glass coverslips (three 12-mm coverslips per 35-mm dish) coated with collagen and poly-l-lysine. Electrophysiological experiments were performed 24 to 48 h after transfection. For each experimental group, five to eight GFP-positive cells per coverslip were studied from at least three different (independent) transfections.
Construction of C616G, C621G, C634G, and 3Cys Mutants of TRPV1. Rat TRPV1 mutants were constructed by PCR amplification using rat TRPV1-specific primers synthesized to contain a point mutation converting the respective nucleotides at positions Cys616, Cys621, and Cys634 to Gly616, Gly621, and Gly634. The Quik-Change XL site-directed mutagenesis kit (Stratagene, La Jolla, CA) was used according to manufacturer's protocol to perform point mutations in TRPV1. The overlapping primer pairs were as follows: TRPV1-C616G, 5′-G TCC ACA CCA CAC AAG GGC CGG GGG TCT GCC TGC AAG CC-3 (sense) and 5′-GCA GGC AGA CCC CCG GCC CTT GTG TGG TGT GGA CTC C-3′ (antisense); TRPV1-C621G, 5′-GC CGG GGG TCT GCC GGC AAG CCA GGT AAC TCT TAC AAC-3′ (sense) and 5′-GA GTT ACC TGG CTT GCC GGC AGA CCC CCG GCA CTT GTG-3′ (antisense); TRPV1-C634G, 5′-C AAC AGC CTG TAT TCC ACA GGT CTG GAG CTG TTC AAG TTC-3′ (sense) and 5′-CTT GAA CAG CTC CAG ACC TGT GGA ATA CAG GCT GTT GTA AG-3′ (antisense). Using the TRPV1-C634G as a template, the following primer pair was designed to mutate the remaining two extracellularly located cysteines: TRPV1-3′CYS, 5′-G TCC ACA CCA CAC AAG GGC CGG GGG TCT GCC GGC AAG CCA GGT AAC TCT TAC-3′ (sense; 52-mer) and 5′-GTA AGA GTT ACC TGG CTT GCC GGC AGA CCC CCG GCC CTT GTG TGG TGT GGA C-3′ (antisense).
Construction of S502A/S800A and M644A Mutants of TRPV1. Double mutant S502A/S800A was constructed by two PCR reactions. In a first reaction, TRPV1-S800A, the following primer pair was used with TRPV1 as a template: 5′-CTT CTG AGG GAT GCA GCC ACT CGA GAT AGA CAT GC-3′ (sense) and 5′-GC ATG TCT ATC TCG AGT GGC TGC ATC CCT CAG AAG-3′ (antisense). In the second reaction, TRPV1-S800A was used as a template and the overlapping primer pair was as follows: TRPV1-S502A/S800A, 5′-CTG CAG AGG CGA CCA GCC CTC AAG AGT TTG TTT G-3′ (sense) and 5′-C AAA CAA ACT CTT GAG GGC TGG TCG CCT CTG CAG-3′ (antisense). To construct TRPV1-M644A mutant, the following primer pair was used: 5′-C AAG TTC ACC ATC GGC GCG GGC GAC CTG GAG TTC-3′ (sense) and 5′-GAA CTC CAG GTC GCC CGC GCC GAT GGT GAA CTT G-3′ (antisense).
Base changes introducing glycine or alanine are in bold. All site-directed mutated constructs were confirmed by DNA sequencing using an automated sequencer (ABI PRISM 3100; Applied Biosystems, Foster City, CA).
Electrophysiology. Whole-cell membrane currents were recorded by using an Axopatch-1D amplifier and the pCLAMP8 and pCLAMP9 software (Molecular Devices, Sunnyvale, CA). Electrodes were pulled from borosilicate glass and after filling had a resistance of 4 to 6 MΩ. The series resistance was usually less than 10 MΩ and was compensated to ∼80%. A system for fast superfusion of the cultured cells was used for drug and heat application. It consisted of a manifold of seven fused silica capillaries connected to a common outlet made from a glass capillary around which insulated copper wire (20 μm in thickness) was coiled to pass direct current for heating the solutions superfusing the cell under investigation (Dittert et al., 2006). The temperature of the superfusing solution was measured by a miniature thermocouple inserted into the outlet capillary near to its orifice that was placed less than 100 μm from the cell under investigation.
Reagents and Solutions. Before and after the test solutions, the cells were superfused with control extracellular solution of the following composition: 160 mM NaCl, 2.5 mM KCl, 1 mM CaCl2, 2 mM MgCl2, 10 mM HEPES, and 10 mM glucose; pH was adjusted to 7.3 with NaOH. The divalent-free (DVF) extracellular solution contained 160 mM NaCl, 10 mM HEPES, and 10 mM EDTA; pH was adjusted to pH 7.4 with NaOH. The intracellular pipette solution contained 125 mM Cs-gluconate, 15 mM CsCl, 5 mM EGTA, 10 mM HEPES, 0.5 mM CaCl2, and 2 mM MgATP, pH was adjusted to 7.3 with CsOH. The osmolarities of the extracellular and the intracellular solution were 320 and 290 mOsM. Unless otherwise specified, all chemicals and reagents were purchased from Sigma-Aldrich (St. Louis, MO). Capsaicin solution was prepared from a 0.1 M stock solution in ethanol, stored at -20°C. The final concentration of ethanol was <0.001%. DTT, H2O2, glutathione (GSH), and diamide were prepared from a stock solution of 1, 10, and 1 M, respectively, in distilled water; the final dilutions were used for about 8 h. N- Ethylmaleimide (NEM) was freshly prepared before each experiment from a stock solution of 2 M in ethanol. The final concentration of ethanol was <0.05%. DTNB was diluted directly into the bath solution to achieve the final concentration. The experimental extracellular solutions containing the redox reagents were prepared, and the pH was adjusted immediately before use and checked after the experiments.
Chromatographic Separation of Capsaicin. Chromatographic separation of capsaicin was done on an HPLC-mass spectrometer system (Agilent 1100; Agilent Technologies, Palo Alto, CA) equipped with a mass spectrometric detector (quadrupole MSD). Separation was done on Extend C18 (2.1 × 150 mm; 5 μm) column at 46°C. Elution was made by gradient between mobile phase A (water with 0.1% heptafluorbutyric acid) and B (acetonitrile with 0.085% heptafluorbutyric acid). Gradient started with 5% B and reached 50% B at 10 min, after next 30 min 100% B was achieved. Flow rate was 0.25 ml/min. Condition for mass spectrometric detection was set to drying gas (N2) flow, 10 l/min; nebulizer pressure, 20 psig (138 kPa); drying gas temperature, 350°C; capillary voltage, 4000 V; and fragmentor, 70 V (positive polarity, selective ion monitoring at 306 m/z). Under these conditions, the retention time of capsaicin was 17.8 min. Capsaicin concentration changes were quantified as the peak area ratios between the analyte and the internal standards measured at selective ion monitoring at 306 m/z.
Statistical Analysis. Heat-evoked currents sampled at the rising phase of the temperature ramp were pooled every 0.5°C. Data were routinely discarded if the leak exceeded 5% of the maximum heat induced currents, but for all the data in the DTT experiments, the leak currents were less than 1% of the maximum current; therefore, no leak subtraction was used. Voltage dependence of the open probability, Po, was estimated according to the Boltzmann equation (Nilius et al., 2005): Itail = Imax/(1 + exp(-zF(V - V1/2)/RT)); Po = Itail/Imax, where z is valence of the gating charge; V1/2 is the halfactivation voltage; Imax is the extrapolated maximum tail current; and F, R, and T have their usual thermodynamic meaning. Boltzmann fits were achieved with Clampfit 9 (Axon Instruments) software. All data are expressed as the mean ± S.E.M. Overall statistical significance was determined by analysis of variance, if not stated otherwise. In case of significance (*p < 0.05 or ***p < 0.001), statistical comparisons were performed by Student's t test for individual groups.
Results
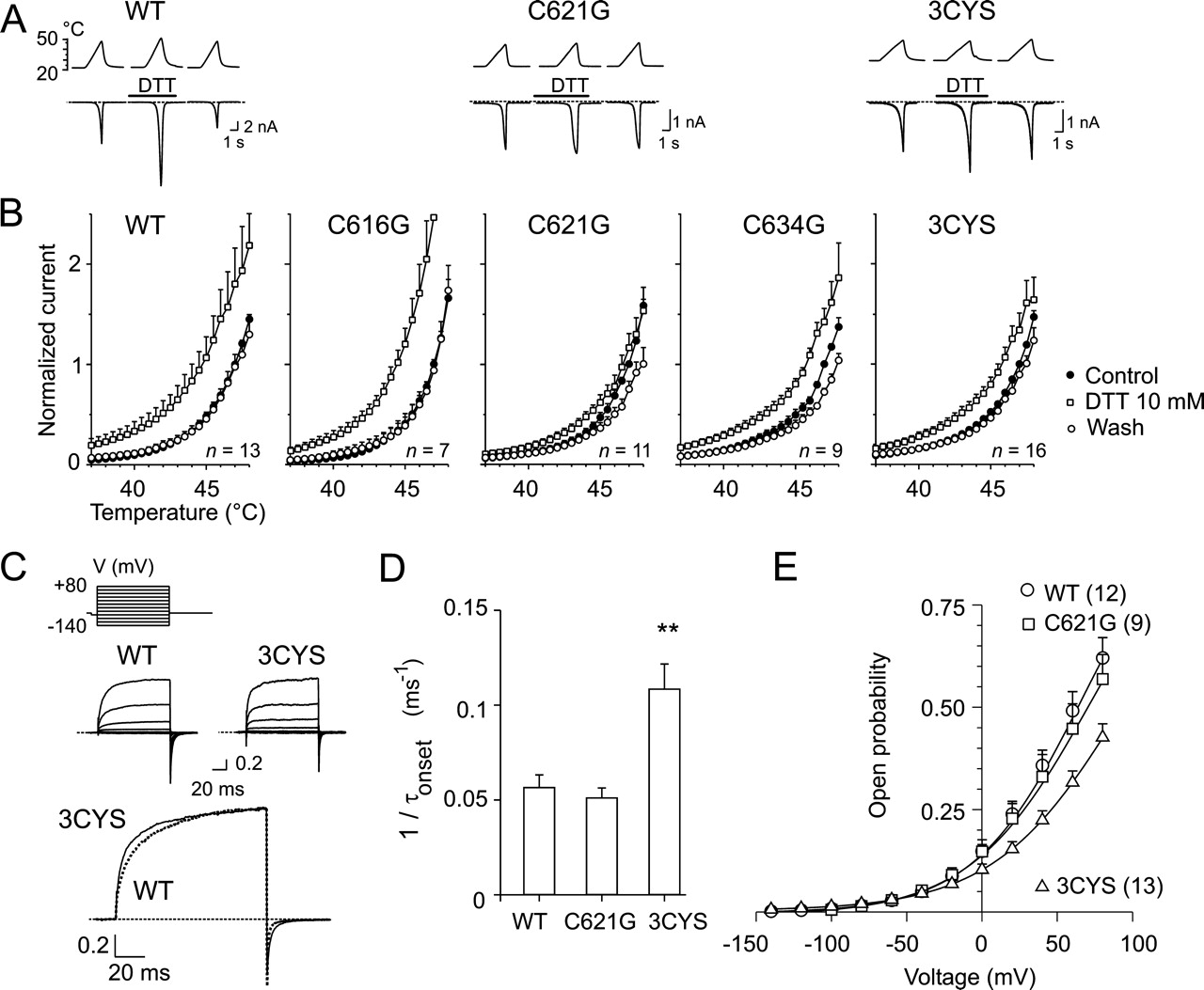
Mutation of Cys621 in TRPV1 Reduces DTT-Induced Potentiation of Heat-Activated Ionic Currents. We have recently demonstrated that the external application of 10 mM DTT to native and recombinant rat TRPV1 receptors markedly potentiates the heat and capsaicin-induced membrane currents (Vyklicky et al., 2002). Because the TRPV1 receptor contains three extracellular cysteine residues (Cys616, Cys621, and Cys634) that could potentially be involved in redox modulation, we substituted these residues with glycine either individually or as a triple mutant (3CYS) and tested the effects of redox-active substances on the whole-cell membrane currents induced by heat. Heat-activated inward currents were induced by 3-s ramps of increasing temperature from 24-48°C at -70 mV in an extracellular control solution and in the presence of 10 mM DTT (10 s). The cells were then washed with bath solution, and the reversibility of the observed effects was determined 1 min later (Fig. 1A). The heat-induced responses of the mutant channels were indistinguishable from those of the wild-type controls. In the presence of DTT, heat-activated currents were potentiated in all constructs over the temperature range of 37-47°C; however, this effect was significantly reduced in the C621G and 3CYS mutants (*, p < 0.05). As shown in Fig. 1B, wild-type TRPV1, C616G, and C634G were potentiated by 2.2 ± 0.4, 2.8 ± 0.6, and 1.8 ± 0.1 at 45°C (n = 13, 7, and 9, respectively), which was greater than the 1.2 ± 0.1 and 1.4 ± 0.1 potentiation produced by DTT in the C621G and TRPV1-3CYS mutants (*, p < 0.05; n = 11 and 16).
In the wild type, C616G, and C634G, DTT shifted the threshold for heat activation so that 38°C was sufficient to induce 23 ± 7, 17 ± 7, and 20 ± 3%, respectively, of the maximal response induced at 47°C in the control extracellular solution. Unexpectedly, the reduction in DTT-induced potentiation of the heat-induced responses was highest in the C621G mutant, not in 3CYS-TRPV1 (*, p < 0.05). These findings suggest that the cysteine residue at position 621 is indeed involved in the modulation of the TRPV1 channel by DTT on the extracellular side. Furthermore, the data indicate that the triple mutation of all extracellular cysteine residues (3CYS) has a noticeable impact on TRPV1 functionality, and this could account for the observation that DTT affects the 3CYS channel slightly more than the C621G mutant.
Voltage-Dependent Activation of Mutant Channels C616G, C621G, C634G, and 3CYS-TRPV1. Since the interpretation of mutagenesis studies may be confounded by functional changes induced by the mutations themselves, we investigated the overall function of the cysteine mutant constructs. The expression levels of the mutants were assessed by comparison of current densities pA/pF of the initial heat response measured at a holding potential of -70 mV and at 44-45°C. Although quite variable, the median inward current densities obtained in cells overexpressing the cysteine mutants (-83 pA/pF, n = 13; -48 pA/pF, n = 10; -61 pA/pF, n = 19; -69 pA/pF, n = 10 in C616G, C621G, C634G, and 3CYS, respectively) were not significantly different from those seen in cells transfected with wild-type TRPV1 (-50 pA/pF; n = 22; *, p = 0.75). Cells transfected with either of the mutant receptors gave responses to heat stimuli that were indistinguishable from those of the wild-type channels (Fig. 1A). Given the high degree of cell-to-cell variability in TRPV1 expression and in the magnitudes of capsaicinevoked currents within each experimental group, we were also unable to detect any obvious changes in the kinetics of the membrane currents induced by capsaicin (1 μM; data not shown). At room temperature and normal pH (7.3), the ratcloned TRPV1 can be activated by depolarizing voltages in the absence of any agonists (Vlachova et al., 2003; Voets et al., 2004), and this mode of activation is linked to conformational changes associated with the temperature-dependent gating of the TRPV1 channel (Voets et al., 2004; Nilius et al., 2005). We therefore examined the voltage-dependent gating properties of the mutant channels by recording whole-cell membrane currents induced by a sequentially applied series of 100-ms voltage steps ranging from -140 to +80 mV, in +20-mV increments.
In the wild-type and 3CYS-TRPV1 channels, depolarizing voltage steps elicited large outward membrane currents with a maximum amplitude of 4.4 ± 0.9 nA (n = 13) and 3.9 ± 0.7 nA (n = 16) at +80 mV (*, p = 0.653). The activation kinetics of wild-type TRPV1-mediated currents elicited by a depolarizing step from -140 to +80 mV was described by a single exponential function (τ = 21.0 ± 2.0 ms). As shown in Fig. 1, C and D, the 3CYS mutation produced a channel with significantly faster voltage-dependent gating kinetics (τ = 11.6 ± 1.2 ms; ***, p < 0.001).
Repolarization to the holding membrane potential (-70mV) produced pronounced inward “tail currents”. Detailed analysis of wild-type TRPV1 deactivation kinetics revealed a fast (2.5 ± 0.2 ms) and a slow (15.8 ± 2.7 ms) time constant with a relative contribution of 74 ± 5% for the fast deactivating component. Again, 3CYS-TRPV1 exhibited significantly faster deactivation rates than those of the wild type channels (1.7 ± 0.1 ms; 76 ± 3%; ***, p < 0.001), whereas the slow time constant was not significantly changed (14.6 ± 1.2 ms; *, p = 0.677). We further examined to what extent the individual mutations could be responsible for the functional changes observed in the 3CYS-TRPV1 construct. A comparison of the outwardly rectifying currents induced by the depolarizing voltage steps to +80 mV in the wild type and in the C616G, C621G, and C634G constructs showed that none of the cysteine mutants exhibited significantly different activation and relaxation kinetics from the wild type at the 0.05 level (one-way analysis of variance). Neither were any significant differences found among the wild-type, C616G, C621G, and C634G constructs in the time constants of tail currents induced by repolarization from +80 to -70 mV. To further substantiate these findings, we quantified the open probability of the wild-type, C621G, and 3CYS mutants from the tail currents plotted against activation voltage (Fig. 1E). These plots were fitted to Boltzmann relationships, and the measured tail current amplitudes were normalized to the estimated maximal current amplitude to obtain the voltage dependence of the open probability. By comparing the averaged data, it seemed that substitution of all three cysteines caused a shift in the midpoint of voltage activation (V1/2) from 64.8 ± 7.2 mV (wild-type TRPV1; n = 12) to 95.5 ± 7.5 mV (n = 13) in the 3CYS mutant, whereas V1/2 was unchanged in C621G (69.7 ± 9.5 mV; n = 9; *, p = 0.686). These findings indicate that mutating all three extracellular cysteines together results in measurable functional changes in the voltage-induced activation of TRPV1 channel.
Does DTT Act on TRPV1 Channel Molecule Directly? During whole-cell recordings, extracellular application of the membrane-impermeable physiological reducing agent GSH (10 mM) mimicked the effects of 10 mM DTT in potentiating the heat-induced and voltage-induced membrane currents (Fig. 2, A-C). The effects of GSH on the heat-evoked responses seemed weaker than those of DTT, which could be explained by its smaller redox potential (Scott et al., 1963). These data indicate that sulfhydryl groups facing toward the extracellular side of the membrane are indeed involved in regulating the activity of the TRPV1 channel.

DTT-induced potentiation of heat-evoked currents reduced in C621G and 3CYS mutants of heterologously expressed rat TRPV1. A, representative families of whole-cell currents evoked in wild-type, C621G, and 3CYS mutants by 3-s ramps of heated solutions from 24-48°C in extracellular control solution, in the presence of 10 mM DTT, and recovery 1 min later. Heat-activated currents recovered completely upon washout. Temperatures of superfusing solutions measured by a thermocouple inserted into common outlet capillary of drug application system are indicated above each trace. Dashed lines represent zero current level. Bars above the records indicate duration of drug application. B, averaged currents normalized to currents evoked at 47°C in extracellular control solution shown with error bars representing S.E.M. Heat-activated inward currents were induced by 3-s ramps of increasing temperature from 24-48°C. Holding potential, -70 mV. One-way analysis of variance did not reveal significant differences among wild type and mutants in the extent of recovery over the temperature range of 44-47°C (*, p = 0.399 at 44°C; *, p = 0.192 at 47°C). C, effects of mutation of all three extracellularly located cysteines (3CYS; dotted line) on currents elicited by voltage steps (protocol shown, top) in extracellular control solution. Currents were normalized to maximal current produced at +80 mV. D, activation rates calculated for WT (n = 11), C621G (n = 7), and 3CYS (n = 13) from activation time constants of current traces elicited at +80 mV. E, voltage dependence of open probability of wild-type, C621G, and 3CYS-TRPV1 channels at 24°C. Solid lines are fits with the Boltzmann equation: V1/2 = 64.8 mV, z = 0.71 (wild type); V1/2 = 69.7 mV, z = 0.65 (C621G); and V1/2 = 95.5 mV, z = 0.63 (3CYS). Note shift in V1/2 toward more positive potentials in 3CYS mutant.
That the redox-active substance DTT slightly (but still significantly) potentiates the mutant lacking the three extracellular cysteine residues indicates that DTT may have remaining “nonspecific” effects unrelated to thiol-disulfide exchange at TRPV1 (Alliegro, 2000). Previous studies on NR1/NR2A (Choi et al., 2001) and GABAA (Wilkins and Smart, 2002) receptors demonstrated that the reversible effects of DTT might be caused by the chelation of trace amounts of divalent cations rather than by redox-based mechanisms. We therefore further examined the extent to which 10 mM DTT, applied in the absence of divalent cations, affects the heatinduced currents mediated through wild-type and cysteinemutated TRPV1 channels. As illustrated in Fig. 3, A and B, the presence of DVF extracellular solution elicited significant inward currents in both wild-type and 3CYS-TRPV1-transfected HEK293T cells at room temperature. In both cases, the extracellular application of 10 mM DTT in DVF solution resulted in a marked potentiation of the heat-induced membrane currents, which indicated that the chelation of divalent cations is not the underlying cause of the remaining DTT-induced potentiation of the heat-evoked membrane currents in TRPV1.
Another mechanism that contributes to the potentiation of heat-induced currents might involve the second messenger pathways associated with protein kinases A, C, or calcium/calmodulin-activated protein kinase II (Humphries et al., 2005). Recent functional studies have identified several serine/threonine residues that are substrates for the direct phosphorylation of TRPV1. Among them, S502 seems to be involved in the potentiation of TRPV1 activity by all three types of kinases (Bhave et al., 2002; Rathee et al., 2002; Jung et al., 2004), and together with another serine, S800, is critically involved in protein kinase C-mediated phosphorylation (Numazaki et al., 2002). To explore the possibility that DTT partially affects some of the protein kinase cascades and thus phosphorylation of TRPV1, we mutated two serine residues, S502 and S800, to alanine and measured the effects of 10 mM DTT on the heat-induced membrane currents. Similarly to wild-type TRPV1, the mutant S502A/S800A was potentiated by the extracellular application of 10 mM DTT as illustrated in Fig. 3C. Together, the results obtained with DTT imply that its dominant effect on the TRPV1 channel is caused by a thiol-disulfide interchange process that involves the extracellular cysteine residue Cys621. The minor nonspecific effects of DTT do not seem to involve chelation of divalent cations or protein kinase C (and perhaps also PKA or calcium/calmodulin-activated protein kinase II) signaling pathways.
Thiol Alkylating Agent N-Ethylmaleimide Modulates TRPV1 Channel in a Manner That Depends on the Preceding Treatment with DTT. To further substantiate the notion that the actions of reducing agents are mediated through the modulation of redox-sensitive site(s) and not by some other nonredox-related mechanism, we used the irreversible thiol alkylating agent NEM. NEM forms covalent bonds with cysteine residues and therefore should prevent further chemical oxidation or reduction of these sites. In the first series of experiments, illustrated in Fig. 4, A and B, HEK293T cells expressing wild-type TRPV1 were stimulated with superfusing extracellular solution heated up to 48°C before, during, and 1 to 2 min after a 40-s application of 1 mM NEM. We found that the amplitude and temperature course of the heat-evoked currents were markedly reduced in the presence of NEM and that the threshold for heat activation was shifted toward lower temperatures (less than ∼30°C). The NEM-induced changes were irreversible after >3 min of washout and not reversed by DTT as expected for an alkylation of thiol groups.

Reducing agent GSH mimics effects of DTT. A, GSH-potentiated whole-cell currents evoked by temperature ramps (final temperature, 46-48°C) in wild-type TRPV1. Current values normalized to currents evoked at 46°C in extracellular control solution. Averaged currents shown with error bars representing S.E.M. B, GSH failed to affect 3CYS mutant. Averaged currents from three 3CYS-TRPV1-transfected HEK293T cells exposed to GSH normalized with respect to control response evoked at 46°C. C, effects of DTT and GSH on currents elicited by voltage steps in wild-type TRPV1 (protocol as in Fig. 1C; holding potential, -70 mV; voltage steps from -140 to +80 mV, increments 20 mV). Current-voltage relationships constructed from steady-state currents obtained in control conditions (closed symbols) and in the presence of DTT or GSH (open symbols).
In a separate set of experiments, we sought to generate the maximum number of free sulfhydryl groups that can be alkylated with NEM. Cells were therefore initially exposed to 10 mM DTT together with 1 mM NEM followed by the application of NEM alone, each for a duration of 15 s. We were unable to increase either exposure time or NEM concentration without causing cell damage. In contrast to the results obtained in the experiments shown in Fig. 4A, NEM markedly increased the amplitude of the heat-evoked currents, whereas the slope of the temperature-response relationship remained unaltered. Unexpectedly, NEM-treated cells were substantially more susceptible to the subsequent application of DTT compared with the effects of DTT before the addition of NEM (Fig. 4C). The threshold for heat activation was shifted toward lower temperatures (less than ∼30°C; Fig.4D) in the presence of DTT, and this effect was irreversible after >3 min of washout. NEM and DTT further increased the heat-evoked responses, although less effectively than before.
Together, the data strongly suggest that NEM specifically reacts with free sulfhydryl groups, which, in a manner that depends on the preceding treatment with DTT, results in irreversible changes in the gating of the TRPV1 channel. NEM is an alkylating agent of lower molecular mass and with substantial membrane permeability; it thus seems highly probable that the TRPV1 receptor is modified on the intracellular side. Regardless, the irreversible shift in the activation threshold indicates that the increased channel activity and susceptibility to DTT may result from allosteric and/or conformational changes in TRPV1 induced by NEM.

Effects of DTT do not seem to involve chelation of divalent cations or protein kinase signaling pathways. A, left, WT-TRPV1 channels activated by ramps of heated solutions in extracellular control solution (a), in the presence of DVF extracellular solution (b), and during 8-s exposure to 10 mM DTT in DVF solution (c). The presence of DTT resulted in a potentiation of heat-induced membrane currents. Right, current-temperature plot for currents shown in the left panel. B, effect of DTT in DVF extracellular solution on heat-evoked responses in HEK293T cell expressing 3CYS-TRPV1. C, left, representative traces of DTT-induced increase of heat-activated currents in transfected HEK293T cells expressing double mutant S502A/S800A. Right, temperature-response profiles of heat-activated currents obtained from traces a and b. The traces shown in A, B, and C are representative of three to five similar experiments.
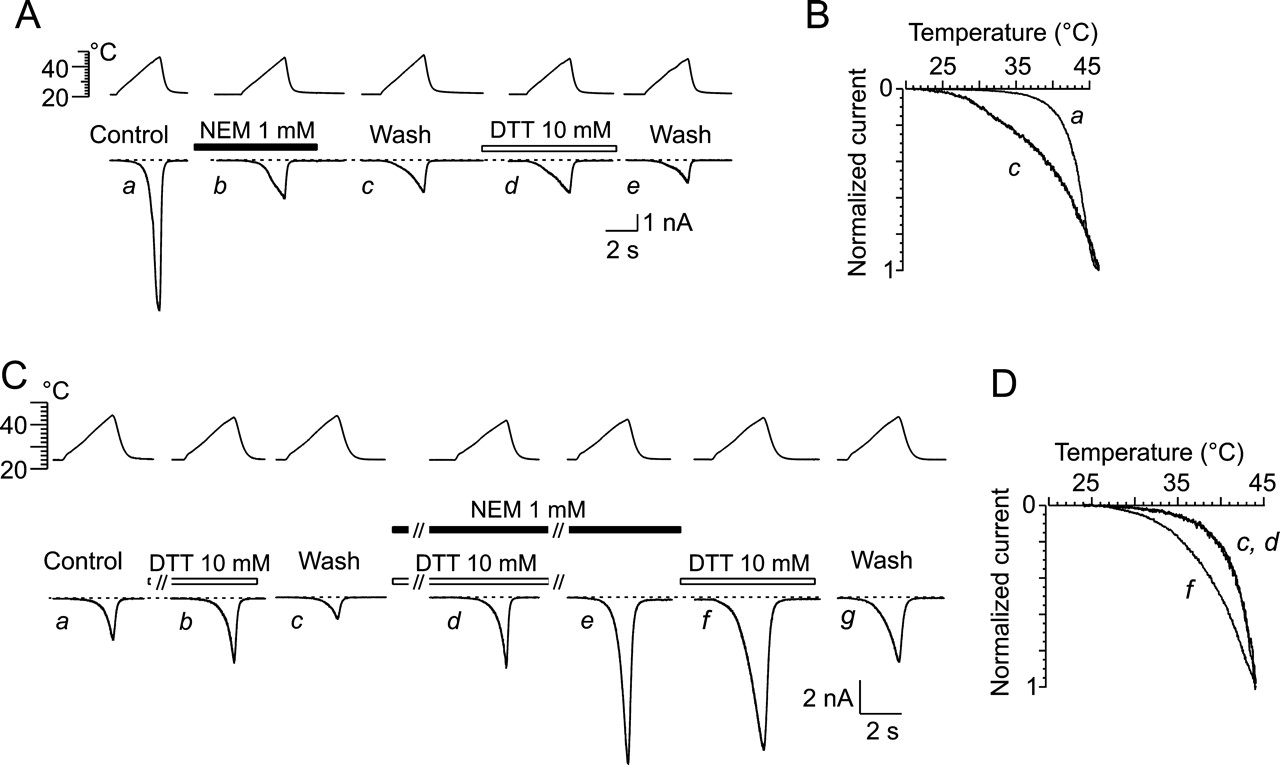
Thiol-alkylating reagent NEM (1 mM) irreversibly and state dependently modifies heat-induced currents in HEK293T cells expressing WT-TRPV1 channels. A, TRPV1 channels activated by superfusing extracellular solution heated up to 48°C before (a), during (b), and 3 min after 40-s application of NEM (c). DTT (10 mM applied for 30 s) exerted no effect when applied ∼2 min after NEM washout. Holding potential, -70 mV. B, current-temperature plot for currents, shown in A, before (a) and after (c) 1 mM NEM treatment. Amplitudes normalized to maximal response obtained at 46°C. C, representative traces of heat-induced responses in TRPV1-expressing HEK cell in control extracellular solution (a), during application of DTT (10 mM) (b), and wash (c), subsequently exposed to DTT together with NEM (1 mM) (d) followed by application of NEM alone (e), each for duration of 15 s. Effects of 10 mM DTT (f) and wash (g) 1 min later. D, current-temperature plot for currents shown in C, recorded in extracellular control solution (c), in the presence of 1 mM NEM together with 10 mM DTT (d), and in the presence of DTT applied after NEM (f). Amplitudes were normalized to maximal response obtained at 44°C. Similar results were obtained in five independent experiments for each type of application protocol.
Oxidizing Agent Diamide Irreversibly Sensitizes TRPV1-Mediated Membrane Currents Induced by Voltage and Noxious Heat. Diamide is a small thiol oxidant that has been shown to rapidly react with GSH, oxidizing it to glutathione disulfide (Kosower and Kosower, 1995). It can cross the membrane readily by diffusion and perturbs the redox balance of cells by oxidizing intracellular GSH. To study the effects of diamide on the heat-dependent and voltage-dependent mode of TRPV1 activation, either inward currents were induced by 3-s ramps of increasing temperature from 24-48°C, or outward currents were elicited with depolarizing voltage steps (60-ms duration; from -140 to +80 mV in 20-mV increments) in control solution and in the presence of 1 mM diamide. The cells were then washed with bath solution for 1 min, and the reversibility of the observed effects was determined (Fig. 5). A short application of diamide (<10 s) was sufficient to induce marked alterations in the TRPV1-mediated currents: diamide produced a shift in the threshold of the heat-evoked responses to lower temperatures (Fig. 5B) and increased the onset rate (from 16.7 ± 2.6 to 9.8 ± 1.4 ms) and the maximum amplitude of outward currents at +80 mV (2.9 ± 0.5-fold; n = 4; Fig. 5C). The effects of diamide were partially reversed by the application of 10 mM DTT for a period of 1 min (data not shown). Frequently, significant (0.4 ± 0.2 nA; n = 5), slowly rising, and irreversible inward currents developed upon 8- to 10-s application of 1 mM diamide at room temperature. The heat-induced currents through the 3CYS-TRPV1, C616G, C621G, and C634G mutant channels were sensitized by diamide to a similar extent to the wild type (Fig. 5D), indicating that neither of the three extracellular cysteine residues are involved in diamide-induced sensitization.
Effects of Oxidizing Agent Cu:Phe on TRPV1-Mediated Heat-Induced Currents. We have previously reported that the oxidizing reagent Cu:Phe (100:400 μM) blocks membrane currents induced by noxious heat in native and recombinant TRPV1 channels (Tousova et al., 2004). Although the amplitudes of the heat-evoked responses substantially recovered after Cu:Phe blockage by washing the cells for 1 min in extracellular control solution, a more detailed quantitative reexamination of the recovered heat-evoked currents indicated that after the application of Cu: Phe (8 s), the threshold for heat activation was frequently shifted to lower temperatures in the triple mutant 3CYS (Fig. 6A). Moreover, subsequent extracellular application of 10 mM DTT (twice for 8 s) restored the threshold for heat activation (Fig. 6B). These data provide additional evidence that membrane-permeable oxidizing agents can modulate the thermal sensitivity of TRPV1 channel, probably from the intracellular side.

Oxidizing agent diamide irreversibly potentiates TRPV1-mediated heat-induced membrane currents. A, effect of diamide on heat-evoked responses in HEK293T cell expressing WT-TRPV1. Temperature protocol shown above. Dashed lines indicate zero membrane current in all records. B, summary of effects of diamide on heat-evoked currents in five HEK293T cells expressing wild-type TRPV1. Currents normalized to current obtained at 45°C before application of diamide, and normalized values represent means ± S.E.M. C, effect of 1 mM diamide on currents elicited by voltage steps (protocol shown in b, inset). b, time course of onset of current traces, shown in a, elicited at +80 mV before (○), in the presence of diamide (▪) and 1 (•) and 2 min (▴) after washing out diamide. Currents were normalized to maximal current produced at +80mV. c, instantaneous current-voltage plots of recordings shown in a measured at the end of each pulse to +80 mV. D, representative heat-induced responses recorded from HEK293T cell transiently transfected with 3CYS mutant of TRPV1. Similar results obtained in five cells. Exposure of cell to diamide resulted in increase in resting membrane current in this cell.
Chloramine-T Irreversibly Sensitizes TRPV1-Mediated Membrane Currents Induced by Noxious Heat. To further elucidate the mechanisms underlying the modulation of TRPV1 by oxidizing agents, we explored the effects of chloramine-T (Ch-T) on the TRPV1-mediated, heat-evoked currents. Ch-T is an oxidizing agent that, apart from cysteines, preferentially oxidizes methionine to methionine sulfoxide (Vogt, 1995; Schlief et al., 1996). Thus, other possible targets for redox modulation are the methionine residues in TRPV1, of which Met644, a critical residue for TRPV1 channel permeability (Garcia-Martinez et al., 2000; Ferrer-Montiel et al., 2004) located at the center of a putative selectivity filter, seems to be the most exposed to the extracellular environment. The responses induced by 3-s ramps of increasing temperature from 24-48°C in wild-type TRPV1 were markedly and irreversibly sensitized by the presence of 1 mM Ch-T (Fig. 7, A-D). Ch-T irreversibly sensitized the heatinduced currents through 3CYS and M644A mutant channels to a similar extent to the wild type, indicating that neither of the three extracellular cysteine residues nor methionine at position 644 is involved in the Ch-T-induced sensitization of TRPV1. Similar to the effects of diamide, significant inward currents developed in most of the cells examined upon an 8- to 10-s application of 1 mM Ch-T at room temperature (182 ± 80 pA, n = 9; 314 ± 74 pA, n = 5; and 190 ± 70 pA, n = 5 for WT, 3CYS, and M644A, respectively). These currents had a slow onset (τ ∼ 2 s) and were irreversible over the time of recording, consistent with the hypothesis that Ch-T has to cross the plasma membrane before reaching the target of its activity. After several heat applications, the Ch-T-induced currents frequently developed into a deteriorating membrane leak.

Oxidizing agent Cu:Phe irreversibly reduces threshold for heat activation of TRPV1. A, representative effects of Cu:Phe and DTT on heat-evoked currents obtained from 3CYS-TRPV1-transfected HEK cell in extracellular control solution (a), in the presence of Cu:Phe (b), upon wash (c and d), followed by two applications of 10 mM DTT (e and f), and after 30-s washing with control extracellular solution (g). Holding potential, -70 mV. Data are representative traces, typical of n = 7. B, superimposed current-temperature plots for responses shown in A, recorded in extracellular control solution (a), after Cu:Phe (c), and after DTT (g). Currents normalized to the maximal response obtained at 48°C.
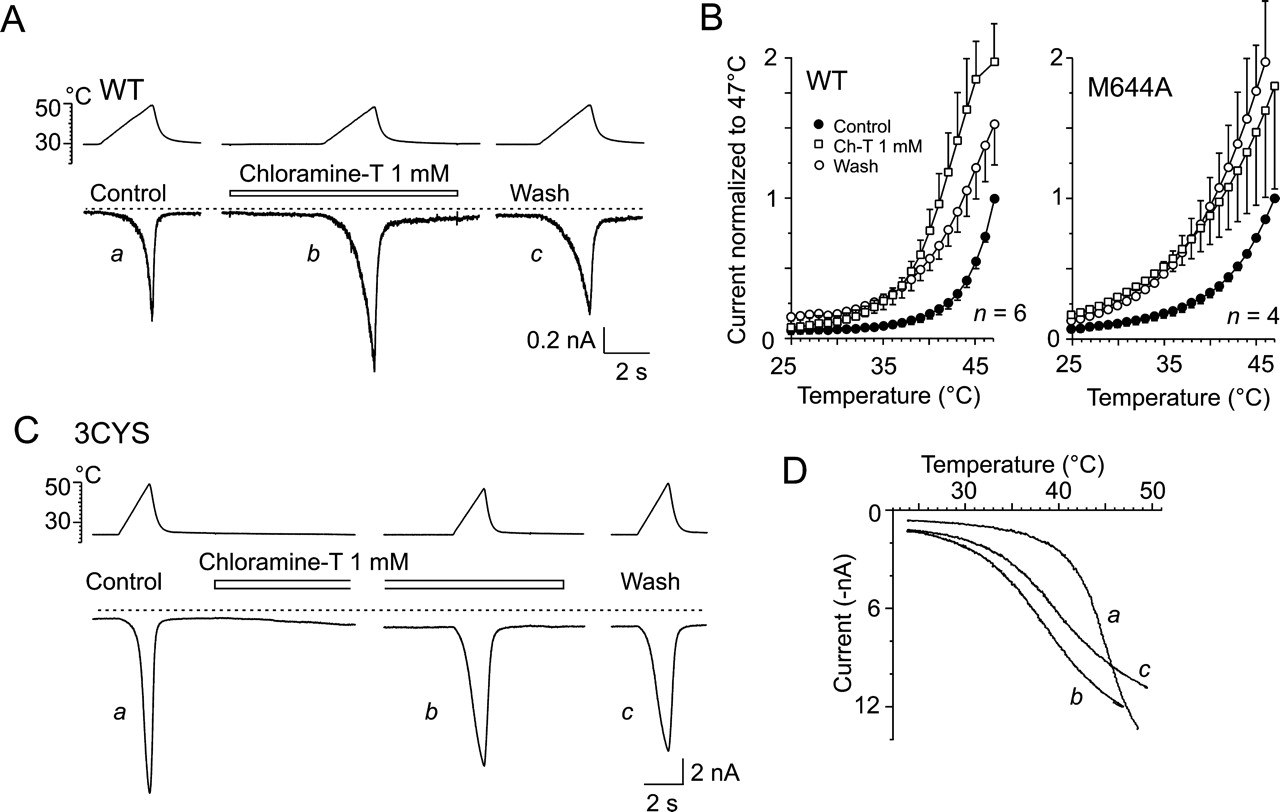
Ch-T irreversibly reduces threshold for heat activation of TRPV1. A and C, representative traces of heat-induced responses obtained from WT-TRPV1-transfected and 3CYS-TRPV1-transfected HEK293T cells in extracellular control solution (a), in the presence of 1 mM Ch-T (b), followed by 30-s washing with extracellular control solution (c). B, summary of effects of Ch-T on heat-evoked currents in six HEK293T cells expressing wild-type TRPV1 and in four cells expressing M644A mutant. Currents normalized to current obtained at 47°C before application of Ch-T, and normalized values represent means ± S.E.M. D, superimposed current-temperature plots for responses shown in C recorded in 3CYS mutant in extracellular control solution (a), in the presence of Ch-T (b), and upon wash (c).
Effects of Sulfhydryl-Specific Oxidizing Agents Hydrogen Peroxide and DTNB on TRPV1-Mediated HeatInduced Currents. To explore further to what extent the effects of oxidizing agents are specific to cysteines, we used the cysteine-selective oxidants H2O2 and DTNB (0.5 mM) and tested their effects on the heat-induced membrane currents mediated by wild-type TRPV1 channels (Fig. 8). The membrane currents evoked by heat were potentiated by 10 mM H2O2 (by 28% at 45°C). Notably, this potentiation was fully reversed within 50 s. In this respect, the potentiating effect of H2O2 was qualitatively different from that of other oxidizing agents (Fig. 8, A and B). The membrane-impermeable oxidant DTNB was completely ineffective (Fig. 8, C and D) in all cells tested (n = 6), which suggests that surface-exposed cysteine residues either do not contribute to TRPV1 modulation by oxidizing agents or are closer to the fully oxidized than the fully reduced state under steady-state conditions. In contrast, H2O2 is freely membrane permeant and the sulfhydryl groups available to react with H2O2 include both hydrophilic and hydrophobic sites. Thus, these data suggest that H2O2 can interact with sulfhydryl groups distinct from those exposed to the surface.
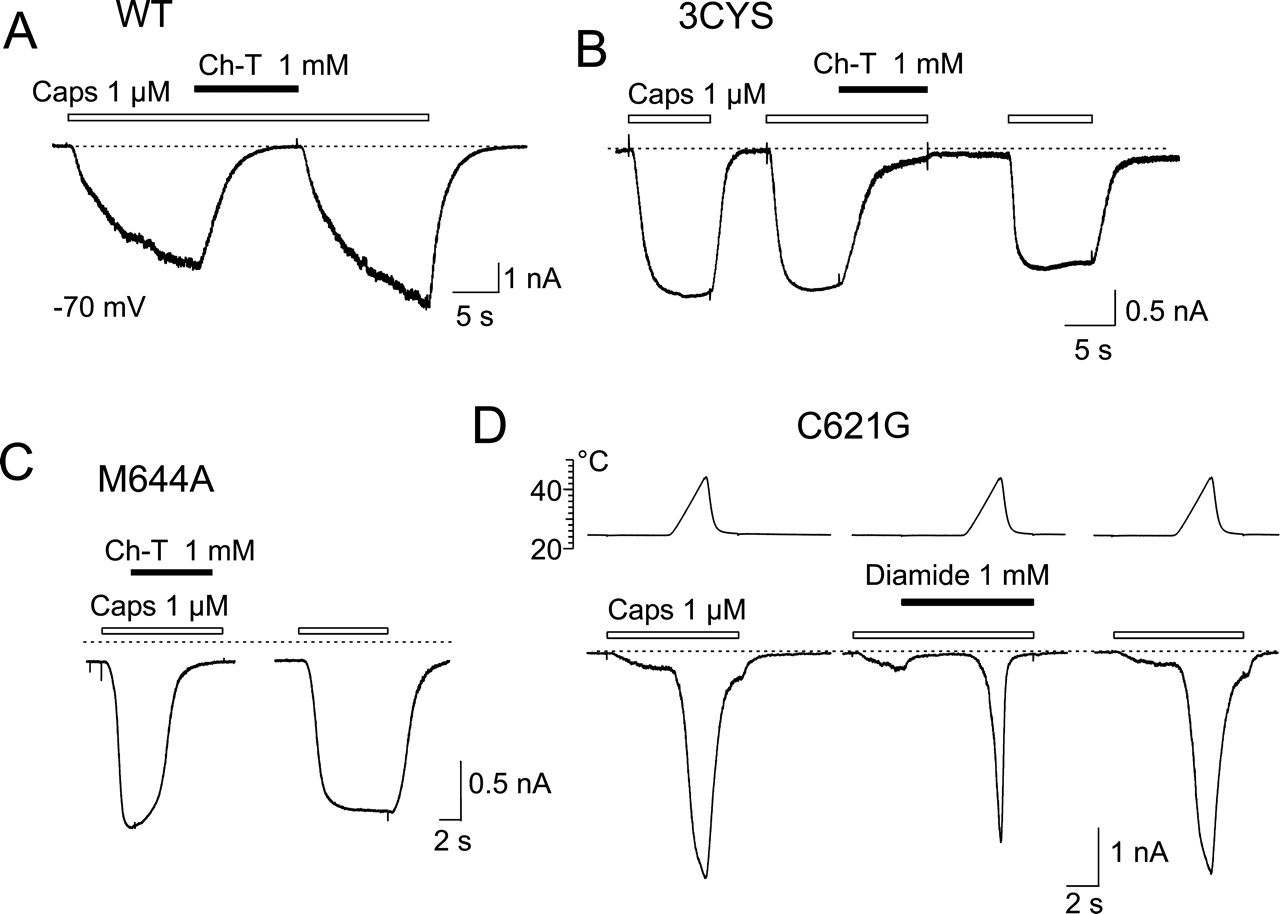
Concentration of Capsaicin Reduced in the Presence of Oxidizing Compounds. The sulfhydryl-oxidizing agents Hg2+, DTNB, GSSH, H2O2, and thimerosal have been reported to reduce the capsaicin-evoked currents in TRPV1-transfected HEK293T cells (Jin et al., 2004). Consistent with these findings, we found that the coapplication of capsaicin (1 μM) and Ch-T (1 mM) reversibly inhibited the capsaicinevoked responses in wild-type TRPV1 at room temperature (by 95 ± 4%; n = 5; Fig. 9A). However, we subsequently found that Ch-T equally inhibits the capsaicin-evoked currents in the TRPV1 mutant lacking extracellular cysteines, 3CYS (Fig. 9B). Moreover, since Ch-T is known to oxidize both cysteine and methionine residues, we evaluated the effects of this oxidizing agent on the mutant TRPV1-M644A and found that the capsaicin-evoked responses were again inhibited to the same extent as in the wild type (Fig. 9C). To a lesser degree (by 50 ± 15%; n = 5), Ch-T also inhibited the inward currents induced by 100 μM 2-aminoethoxydiphenyl borate (data not shown), a compound that is a common activator of TRPV1, TRPV2, and TRPV3 channels (Hu et al., 2004).

Effects of hydrogen peroxide and DTNB on TRPV1-mediated heat-induced currents. A, representative traces showing whole-cell patch-clamp recordings from HEK293T cell transiently transfected with wild-type TRPV1. Bar above the records indicate 40-s period of drug application. Holding potential, -70 mV. B, summary of effects of H2O2 on heat-evoked currents derived from four cells expressing WT-TRPV1. Currents normalized to current obtained at 45°C before application of H2O2, and normalized values represent means ± S.E.M. C, membrane-impermeable, cysteine-specific oxidizing agent DTNB had no effect on TRPV1-mediated heat-evoked currents. D, superimposed current-temperature plots for responses shown in C, recorded in extracellular control solution (a), in the presence of DTNB (b), and 1 min after washing out DTNB reagent (c).

Oxidizing agents influence capsaicininduced activity of TRPV1 by decreasing its concentration. A to C, representative traces of capsaicin-induced whole-cell responses recorded from HEK293T cell transfected with WT-TRPV1, 3CYS-TRPV1 mutant, and M644A-TRPV1 mutant. Capsaicin-induced currents abolished by coapplication of capsaicin with 1 mM chloramine-T. Bars above the records indicate duration of drug application. Holding potential, -70 mV. D, effect of diamide on capsaicin-evoked responses in HEK293T cell expressing C621G-TRPV1. Temperature protocol shown above. Dashed lines indicate zero membrane current in all records in this figure.
These observations led us to suspect that the effects of oxidizing agents on capsaicin-evoked currents did not reflect the thiol-disulfide exchange at TRPV1 since the above-mentioned effects were determined by coapplication with capsaicin. Based on the results from the study of Jin et al. (2004) mentioned above, in which an extracellular cysteine at position 621 has been identified as the site for redox modulation, we next examined the effects of diamide on the capsaicininduced currents in the C621G mutant of TRPV1. Also, the coapplication of 1 mM diamide on this construct reversibly inhibited the currents evoked by 1 μM capsaicin over the temperature range 23-48°C as illustrated in Fig. 9D. Therefore, we were interested to learn whether the oxidizers might influence the effective concentrations of capsaicin in general. Chromatographic separation of the equilibrium mixture of 1 μM capsaicin and 1 mM diamide or 1 mM chloramine-T on an HPLC-mass spectrometer system revealed that the capsaicin concentration decreased by 35% in the presence of diamide, and, in the case of chloramine-T, it was below the detection level of the system. These data strongly suggest that the effects of oxidizing agents on capsaicin-activated TRPV1 channels are caused by a reduction in the concentration of capsaicin rather than by a redox-based mechanism.
Discussion
In this study, we demonstrate that the chemical modification of TRPV1 by both reducing and oxidizing agents leads to an increased response to heat and involves both extracellular and intracellular mechanisms. In addition, we show that the extracellularly located cysteine at position 621 contributes to the DTT-induced potentiation of heat-activated ionic currents mediated by TRPV1. On the intracellular side, the heat-induced activity of TRPV1 is modulated by the membrane-permeable oxidizing agents diamide, Cu:Phe, and chloramine-T.
Potentiation of TRPV1 Channel by Reducing Agents. The tetrameric TRPV1 channel possesses 18 cysteine and 20 methionine residues in each subunit. Of these residues, three cysteines (Cys616, Cys621, and Cys634) and three methionines (Met541, Met609, and Met644) can theoretically undergo reduction or oxidation on the extracellular side of the plasma membrane. Additional state-dependent sites of redox modulation may be represented by the methionine residues located at pore-lining segments S5 (Met581) and S6 (Met677 and Met682), but in this case, further studies may actually be complicated by mutations within this critical region frequently giving rise to nonfunctional channels (Kuzhikandathil et al., 2001).
In our present study, the mutation of the three extracellular cysteines (3CYS) fully abrogated the effects of the membrane-impermeable reducing agent GSH (10 mM) on heatevoked responses. However, the 3CYS mutation did not fully abolish the potentiating effects induced by DTT, a compound that readily permeates cell membranes (Lauriault and O'Brien, 1991). We excluded the possibility that some effects of DTT on 3CYS might be caused by chelation of trace amounts of divalent cations and we also made an attempt to exclude the involvement of protein kinase activation by DTT. The prevailing view is that, on the cytoplasmic side, high concentrations of the most abundant thiol GSH (above 10 mM in neuronal cells; Slivka et al., 1987) keep proteins in the reduced state. Therefore, one would expect that DTT, once passed through the cell membrane, should not affect protein sulfhydryls on the intracellular side. The remaining sensitivity of 3CYS to DTT observed in our experiments could thus be unrelated to thiol-disulfide exchange (Alliegro, 2000).
Modulation of TRPV1 by Oxidizing Agents. Whereas extracellular Cys621 seems to be essential for the redoxmediated modulation of TRPV1, the precise structural components that contribute to this type of modulation on the intracellular side remain to be identified. Our results demonstrate that oxidizing compounds such as diamide, Cu:Phe, chloramine-T, and H2O2 strongly potentiate the heat-evoked activity of TRPV1. The 3CYS mutation did not influence the effects of membrane-permeable diamide, Cu:Phe, or Ch-T, whereas the membrane-impermeable cysteine-specific oxidizing agent DTNB had no effect on wild-type TRPV1-mediated heat-induced currents. These results together strongly suggest that there are no oxidizable residues exposed to the surface that may contribute to TRPV1 function. We cannot rule out that the effects of the membrane-permeable oxidizing agents might be a result of alterations in the activity of a protein putatively associated with TRPV1 or the involvement of second messenger signaling cascades. However, in the present study, we demonstrate that there are state-dependent irreversible changes in TRPV1 functionality after NEM treatment. We found that a 40-s application of NEM, an alkylating agent, rendered TRPV1 channel insensitive to subsequent DTT modulation. In contrast, TRPV1-mediated heat-evoked responses were strongly sensitized by DTT following the coapplication of NEM and DTT (Fig. 4). Thus, thiol-reactive agents seem to modulate the gating of TRPV1 channels by an action directly on the TRPV1 protein or associated protein.
Redox-Active Substances Reduce Effective Concentration of Capsaicin. In this study, we examined the effects of redox-active substances on the heat-induced membrane currents, because increasing the temperature above ∼42°C activates the channels directly (Tominaga et al., 1998). This experimental approach has the advantage that no ligand binding to the receptor is needed to gate the channels; thus, potential ambiguities arising from direct modification of the ligand molecule by redox-active substances can be avoided. That ligands themselves can be modified by redox reagents should be taken into consideration when defining the role of endogenous reducing and oxidizing agents on TRPV1 functionality. Our data indicate that capsaicin concentration is radically altered by the presence of oxidizing agents. More generally, the redox-active substances can substantially affect the activity of TRPV1 channels by influencing their modulators or coactivators.
Thermal Threshold for TRPV1 Activation Depends on Cellular Redox State. It is widely thought that disulfide bonds have been added during evolution to enhance the thermal stability of those proteins that play their biological roles in an oxidizing extracellular environment. The formation of disulfide bonds, however, does not necessarily lead to enhanced protein stability. On the contrary, a decreased thermodynamic stability has been observed in some of the proteins in which novel disulfides have been introduced (Matsumura et al., 1989; Betz, 1993; Petersen et al., 1999). That both reducing and oxidizing conditions lead to a shift in the temperature activation threshold toward cooler temperatures in TRPV1 channels suggests that their functionality may require an optimal redox state. This is consistent with the hypothesis that the thiol redox state may serve as a critical tuner for the thermal threshold of TRPV1 ion channels and thus sensory neurons.
In spite of the uncertainties in our present knowledge concerning the specific molecular mechanisms involved, it seems likely that changes in the redox state of TRPV1 channels could play an important role in the pathogenesis of acute or chronic pain states. We demonstrate that in the presence of redox compounds, the temperature threshold for TRPV1 activation is substantially reduced such that normal body temperature is capable of activating TRPV1. Therefore, disturbances in the redox state in peripheral tissues under a variety of pathophysiological conditions, including tissue damage, metabolic stress, ischemia, and inflammation, may contribute directly to regulating activity of TRPV1 channel and thus to nociceptor sensitization. A potential physiological role for redox agents in the modulation of pain sensation has also been recently proposed by Todorovic et al. (Todorovic et al., 2001; Nelson et al., 2005). Their results provide evidence that redox agents selectively modulate T-type Ca2+ channels in rat peripheral nociceptors, whereas the currents through other voltage-gated (Na+, K+, high-voltage-activated, or Ca2+) and ligand-gated channels (capsaicin, low pH, or ATP) are not influenced. Although these authors indicate that DTT is ineffective in modulating capsaicin-gated and heat-gated currents in sensory neurones even at severalfold higher concentrations than those affecting T-type Ca2+ channels, in the present report, we demonstrate that redox-active substances regulate the heat-induced activity of recombinant TRPV1 receptor channels. The changes induced by membrane-permeable oxidizing agents are irreversible and robust enough to significantly lower the threshold for heat activation.
Structural Implications of Our Findings. Our studies using various sulfhydryl reagents may help to clarify the multiple mechanisms involved in the modulation of TRPV1 by changes in redox potential. By using the alkylating agent NEM, we demonstrated that the threshold for heat activation can be irreversibly modified when NEM is concurrently applied with DTT during heat stimulation. This is consistent with the possibility that the target residues of the redox modulation lie directly within the TRPV1 receptor channel protein and undergo state-dependent changes in accessibility (Fig. 4). Recently, structural information on the architecture of numerous channels has been gathered by substituted cysteine scanning mutagenesis studies (for review, see Karlin and Akabas, 1998). Thus, precise knowledge of the properties of endogenous cysteines is essential.
To obtain a more complete picture of the molecular basis of redox modulation, we submitted the TRPV1 protein amino acid sequence from V596 to D654 to the predictive software program DiANNA (DiAminoacid Neural Network Application) that incorporates a neural network-based predictor trained to distinguish the bonding states of cysteine in proteins (Ferre and Clote, 2005). The results from the cysteine oxidation state prediction module yielded a half-cystine for Cys621, which was in agreement with our observation that Cys621 is involved in the modulation of the TRPV1 channel by extracellular DTT. Based on this prediction software, it is also likely that there are no intramolecular disulfide bonds within the TRPV1 receptor protein. The process of reduction and oxidation of Cys621, therefore, may involve an interaction of two adjacent TRPV1 subunits. Of 13 cysteine residues in the cytoplasmic N-terminal (10) and C-terminal regions (three) of TRPV1, two are predicted to be oxidized at the N-tail outside of the ankyrin repeat domain (Cys21 and Cys63). It can be imagined that these intracellular cysteines can participate in the formation of interior intramolecular disulfide cross-links, similar to those reported for homotetrameric cyclic nucleotide-gated channels (Gordon et al., 1997) and voltage-gated Shaker potassium channels (Schulteis et al., 1996).
Earlier radioligand binding studies (Szallasi et al., 1993) showed that reducing and oxidizing agents diminish both the positive cooperativity and apparent binding affinity of [3H]resiniferatoxin to the vanilloid receptor. Our present findings extend these results to the temperature-dependent mode of TRPV1 activation and support the interpretation that cooperation between TRPV1 subunits may be, at least in part, subject to redox modulation.
Conclusions
In summary, our results provide evidence that at least some of the actions of the redox-active substances are likely to be mediated by the reduction or oxidation of specific sites on the TRPV1 channel molecule. This specific oxidation and chemical reduction may have substantial consequences for TRPV1 channel functionality and may constitute a mechanism for the regulation of TRPV1 activity under both physiological and pathological conditions.
Acknowledgments
We thank Dr. I. Miksik for HPLC analysis.
Footnotes
-
This work was supported by the Czech Science Foundation grants 305/06/0319 and 309/04/0496, the Research Project Fund of the Academy of Sciences on the Czech Republic grant AV0Z 5011922, and by the Ministry of Education, Youth and Sports of the Czech Republic grants 1M0517 and LC 554.
-
ABBREVIATIONS: TRPV1, transient receptor potential vanilloid receptor-1; DTT, dithiothreitol; Cu:Phe, copper-o-phenantroline [Cu(II)-1,10-phenantroline]; HEK, human embryonic kidney; PCR, polymerase chain reaction; DVF, divalent-free; GSH, reduced glutathione; DTNB, 5,5′-dithio-bis-(2-nitrobenzoic acid); ECS, extracellular solution; NEM, N-ethylmaleimide; HPLC, high-performance liquid chromatography; Ch-T, chloramine-T; WT, wild-type/wild type.
- Received February 1, 2006.
- Accepted April 13, 2006.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}