


Abstract
Glutathione-S-transferase-catalyzed conjugation of glutathione (GSH) to aflatoxin B1-8,9-epoxide plays an important role in preventing binding of this ultimate carcinogen to target macromolecules. Once formed, the aflatoxin B1-epoxide-GSH conjugates are actively extruded from the cell by an unidentified ATP-dependent export pump or pumps. Two possible candidates for this GSH conjugate pump are the 190-kDa multidrug resistance protein (MRP) and the 170-kDa P-glycoprotein. Both proteins belong to the ATP-binding cassette superfamily of transmembrane transport proteins and confer resistance to a similar spectrum of natural-product drugs. Using membrane vesicles from MRP-transfected cells, we found that MRP transports GSH conjugates of both the endo-isomers and exo-isomers of aflatoxin B1-8,9-epoxide in an ATP-dependent, osmotically sensitive manner (V max = 180 pmol/mg/min,K m = 189 nm). Membrane vesicles from P-glycoprotein-overexpressing cells showed very low levels of transport. MRP-mediated transport was inhibited by an MRP-specific monoclonal antibody and by a variety of GSH derivatives and cholestatic steroid glucuronides. ATP-dependent transport of unmodified aflatoxin B1 by MRP-enriched membrane vesicles was low but markedly enhanced in the presence of 5 mmGSH, even though GSH conjugates of aflatoxin B1 were not formed by the vesicles. These data demonstrate that MRP is capable of energy-dependent transport of aflatoxin B1 and its GSH conjugates and suggest a potential protective role for MRP in mammalian chemical carcinogenesis.
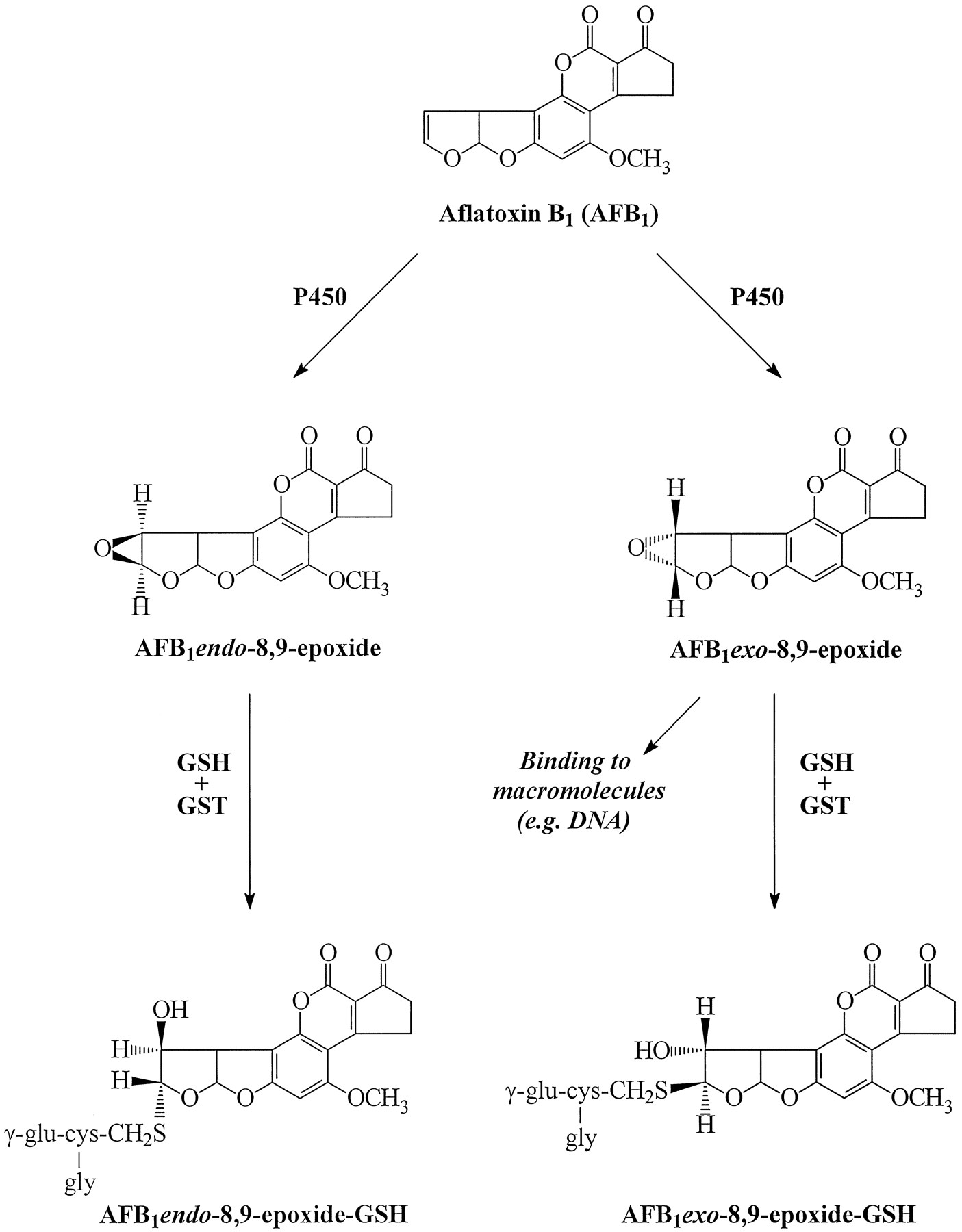
AFB1 is a mycotoxin produced by certain Aspergillus species. Although primarily characterized as a hepatotoxin and hepatocarcinogen in many vertebrate species, there is evidence that the lung is also a target for AFB1 carcinogenicity (1, 2). The mutagenicity and carcinogenicity of AFB1 are believed to result primarily from its cytochrome P450-mediated bioactivation to AFB1-8,9-epoxide, which binds to DNA (Fig.1). Both endo- and exo-stereoisomers of AFB1-8,9-epoxide exist, and although they are both produced in a variety of tissues, only exo-FB1-8,9-epoxide binds efficiently to DNA and is capable of inducing G-to-T transversionsin vitro (3). The more chemically stable endo-epoxide is 500-fold less mutagenic (4). Epoxide hydrolase and GST are both involved in hepatic detoxification of activated AFB1, but the GST-catalyzed conjugation of GSH to AFB1-8,9-epoxides is thought to play the more important role in preventing epoxide binding to target macromolecules (2, 3). Once generated, the AFB1 endo- and exo-epoxide-GSH conjugates are extruded from the cell. The transport of AFB1-GSH and other GSHS-conjugates is mediated by functionally defined ATP-dependent GSH S-conjugate export pumps (so-called GS-X pumps), which are part of the elimination system or phase III drug metabolism pathways (5). GS-X pump activity has been found to exist in many different organs and cell types (2, 6-8), but the protein or proteins responsible for AFB1-GSH transport have not yet been physically or biochemically identified.

Biotransformation of aflatoxin B1(AFB1) to exo- and endo-epoxides and the corresponding GSHS-conjugates. P450, cytochrome P450.
The 170-kDa P-glycoprotein and the 190-kDa MRP are known to confer resistance to multiple natural-product xenobiotics and chemotherapeutic agents when transfected into previously sensitive cells (9-11). Overexpression of these N-glycosylated phosphoproteins in transfected mammalian cells is associated with reduced drug accumulation resulting from ATP-dependent enhanced drug efflux (12-14). Although both MRP and P-glycoprotein belong to the ATP-binding cassette superfamily of transport proteins, they share only 15% amino acid identity (10). They also seem to differ in the mechanism by which they transport xenobiotics, despite conferring similar cross-resistance profiles (13-15). P-glycoprotein-enriched membrane vesicles have been shown to transport directly a number of chemotherapeutic agents (16-18). In contrast, ATP-dependent transport of the Vinca alkaloid vincristine in MRP-enriched vesicles seems to require GSH (19-21), although some controversy exists (22). MRP can also function as a high affinity transporter of certain GSH and glucuronide conjugates. Potential physiological substrates of MRP include the GSH-conjugated LTC4 (19, 20, 23), GSSG (24), and 17β-estradiol 17-(β-d-glucuronide) (21, 25). In contrast, these conjugated organic anions are transported much less efficiently, if at all, by P-glycoprotein.
In the current study, we show that MRP, but not P-glycoprotein, mediates high affinity ATP-dependent, osmotically sensitive transport of AFB1-GSH that is inhibitable by several organic anions shown previously to compete with known MRP substrates and by an MRP-specific mAb. Furthermore, after separating the endo-isomers and exo-isomers of AFB1-GSH by chiral HPLC, we found that MRP-enriched membrane vesicles transport these two stereoisomers with similar efficiency. Finally, we show that MRP can mediate ATP-dependent transport of unmodified AFB1 and that this transport is markedly enhanced by the addition of GSH.
Experimental Procedures
Materials.
Generally labeled [3H]AFB1 (16.9 Ci/mmol) was obtained from Moravek Biochemicals (Brea, CA). Nucleotides, GSH, GSSG, LTC4, S-alkyl GSH derivatives, steroid glucuronides, glucose-6-phosphate, glucose-6-phosphate dehydrogenase, and NADP were purchased from Sigma Chemical (St. Louis, MO). Authentic AFB1-GSH used as an HPLC standard was generously provided by Drs. G. E. Neal and D. J. Judah (MRC Laboratories, Surrey, UK). AFB1 exo-epoxide-GSH standard used for chiral HPLC analysis was kindly supplied by Dr. T. M. Harris (Vanderbilt University, Nashville, TN). The murine MRP-specific mAbs QCRL-1 and QCRL-3 (Centocor, Malvern, PA) detect different epitopes of the human MRP molecule and have been described previously (26, 27).
Cells.
The origin and culture conditions of the small cell lung cancer cell line H69, its multidrug-resistant MRP-overexpressing variant H69AR, and the revertant H69PR cell line have been described previously (19). The doxorubicin-selected multidrug-resistant 8266/Dox40 human myeloma cell line overexpresses P-glycoprotein and was obtained from Dr. W. S. Dalton (Arizona Cancer Center, Tucson, AZ). It also contains very low levels of MRP, similar to those found in the revertant H69PR cells (26, 27). The production and maintenance of MRP transfected (T14) or vector-transfected (C6) HeLa cell populations were as previously described (13).
Synthesis and HPLC analysis of [3H]AFB1-GSH conjugates.
Liver microsomes and 100,000 × g supernatants (cytosolic fractions) from male New Zealand White rabbits (2.0–2.5 kg) and male CD-1 mice (25–30 g) were prepared and [3H]AFB1-GSH conjugates were generated as previously described (28). Briefly, mouse liver cytosolic protein (21 mg) and rabbit liver microsomal protein (2.5 mg) were added to a 2-ml reaction mixture containing 0.1 m potassium phosphate buffer, pH 7.4, 1.15% (w/v) KCl, 10 mmglucose-6-phosphate, 5 mm MgCl2, 1 mm EDTA, 1 mm GSH, 2 units of glucose-6-phosphate dehydrogenase, and 1.25 mm NADP. Reactions were initiated by the addition of [3H]AFB1 (125 μm; 12.5 Ci/mmol) in dimethylsulfoxide (25 μl). After 2 hr, reactions were terminated by the addition of 4 ml of ice-cold chloroform and centrifuged at 150 × g for 15 min, and the aqueous layer was stored at −20° until separation by HPLC. Total AFB1-GSH conjugates were isolated by reverse-phase HPLC according to the method of Raj et al. (29). Briefly, a 200-μl aliquot of aqueous phase was eluted isocratically with mobile phase (30% methanol and 0.03% acetic acid in water) at 1.5 ml/min using a Waters μBondapak C18 column (3.9 × 300 mm). Peaks were detected by absorbance at 365 nm, and fractions containing [3H]AFB1-GSH were collected. Identification of [3H]AFB1-GSH was established by coelution with authentic AFB1-GSH standard (28).
HPLC separation of [3H]AFB1-GSH endo-isomers and exo-isomers.
AFB1 endo- and exo-epoxide-GSH conjugates were separated according to a modification (28) of the HPLC method of Stresser et al.(30). A 200-μl aliquot of aqueous phase of the conjugation reaction mixture above was injected onto a Pirkle-concept chiral column packed with d-phenylglycine covalently bound to aminopropyl silica (4.6 × 250 mm) (Regis Technologies, Morton Grove, IL). A two-reservoir mobile-phase system was used consisting of 37% methanol in 10 mm KH2PO4 buffer, pH 4.0, pumped at 0.5 ml/min. Absorbance was measured at 365 nm, and 0.5-ml fractions were collected. Identities of [3H]AFB1 endo- and exo-8,9-epoxide-GSH conjugates were determined as previously described (28).
Membrane vesicle preparation.
Plasma membrane vesicles were prepared as described with modifications (19). Cells were homogenized in buffer containing 50 mm Tris·HCl, 250 mmsucrose, 0.25 mm CaCl2, pH 7.5, and protease inhibitors. Cell pellets were frozen at −70° for ≥1 hr, thawed, and then disrupted by N2 cavitation. EDTA was added to 1 mm, and after centrifugation at 500 × gfor 15 min, the supernatant was layered over 35% (w/w) sucrose in 10 mm Tris·HCl and 1 mm EDTA and centrifuged at 100,000 × g for 2 hr. The interface was collected and washed twice by centrifugation. The membrane pellet was resuspended in transport buffer (50 mm Tris·HCl, 250 mmsucrose, pH 7.5) and passed 20 times through a 27-gauge needle for vesicle formation.
Membrane vesicle transport studies.
ATP-dependent transport into the membrane vesicles was measured by a rapid filtration technique (19, 23). Standard transport assays were performed with membrane vesicles (40 μg of protein in a 125-μl reaction volume) that were incubated at 37° in the presence of 250 nm[3H]AFB1-GSH (12.5 Ci/mmol), 10 mm MgCl2, and 4 mm ATP or AMP. Preliminary experiments indicated that an ATP-regenerating system was not required. Transport assays of the individual stereoisomers of [3H]AFB1-GSH were carried out at an initial substrate concentration of 15 nm because of their limited availability. Uptake was stopped by filtration through glass-fiber (Type A/E) filters (Gelman Sciences, Dorval, Quebec, Canada). Transport of 250 nm unmodified [3H]AFB1(16.9 Ci/mmol) was measured in the presence or absence of 5 mm GSH.
Analysis of vesicle-associated [3H]AFB1.
T14 membrane vesicles (300 μg) were incubated at 37o with 250 nm[3H]AFB1 (1 μCi) in a 200-μl reaction mixture containing 4 mm ATP, 10 mmMgCl2, 10 mm dithiothreitol, and 5 mm GSH in transport buffer. After 10 min, the mixture was rapidly diluted in ice-cold transport buffer, and vesicles were pelleted by centrifugation at 100 000 × g. Vesicles were washed and solubilized in 1% sodium dodecyl sulfate and then extracted with CHCl3. The organic layer was dried under nitrogen and resuspended in methanol followed by HPLC analysis on a Waters μBondapak C18 column with 15% acetonitrile/6% ethyl acetate as the mobile phase. Fractions were collected, and radioactivity was determined. The aqueous layer was analyzed by HPLC analysis as described above for AFB1-GSH.
Results and Discussion
The molecular basis of the broad substrate specificities of P-glycoprotein and MRP is not well understood. In 1990, it was suggested that rather than recognizing and transporting unmodified xenobiotics, P-glycoprotein might recognize its substrates after their conjugation to glutathione or another small molecule by the phase II biotransformation enzymes (2, 31, 32). Although there is no evidence to support a requirement for phase II conjugation in P-glycoprotein-mediated transport of xenobiotics, MRP has recently been demonstrated to be a rapid and high affinity transporter of several GSH and glucuronide conjugates in a variety of membrane vesicle systems (19-21, 25). These studies suggested to us that MRP might be a more likely candidate than P-glycoprotein as a physiological mediator of ATP-dependent AFB1-GSH transport. The results of the current study demonstrate that this is indeed the case.
The time course of [3H]AFB1-GSH uptake by vesicles prepared from MRP-transfected T14 HeLa and control transfected C6 HeLa cells is shown in Fig. 2A. Accumulation was measured at 37° at an initial concentration of 250 nm[3H]AFB1-GSH in the presence of 4 mm ATP or AMP. ATP-dependent uptake of [3H]AFB1-GSH in T14 vesicles was rapid and linear up to 5 min. Steady state was approached by 10 min and remained constant for ≤20 min at ∼650 pmol/mg (not shown). During the linear phase, ATP-dependent uptake was ∼120 pmol/mg/min for T14 vesicles. The very low levels of [3H]AFB1-GSH uptake by T14 vesicles in the presence of AMP were similar to those observed for vesicles from C6 cells transfected with vector alone, with or without ATP. Both the initial rate and steady state levels of MRP-mediated [3H]AFB1-GSH uptake were osmotically sensitive (Table 1) and dependent on the presence of hydrolyzable nucleotide (not shown). Initial rates of uptake were determined at several [3H]AFB1-GSH concentrations. Double-reciprocal plots of the data yielded aKm value of 189 nm and a V max value of 180 pmol/mg/min for MRP from T14 cells (average of two independent determinations not shown). The Km value for AFB1-GSH, although not as low as that for LTC4 (Km = 105 nm) (19, 23), is considerably lower than that for 17β-estradiol 17-(β-d-glucuronide) (Km = 2.5 μm) (25) or GSSG (Km = 93 μm) (24). TheKm value for ATP was 330 μm.

Time course of [3H]AFB1-GSH uptake by membrane vesicles from MRP-transfected HeLa cells (T14), control HeLa (C6) cells, drug-sensitive H69 cells, multidrug-resistant H69AR cells, revertant H69PR cells, and P-glycoprotein-overexpressing 8226/Dox40 cells. Membrane vesicles were incubated at 37° in transport buffer containing [3H]AFB1-GSH (250 nm; 12.5 Ci/mmol) and ATP (4 mm) (filled symbols) or AMP (4 mm) (open symbols) for the times indicated. Vesicles were derived as described in Experimental Procedures from HeLa C6 (A, ○, •), T14 (A, ▿, ▾), H69AR (B, ▵, ▴), H69 (C, ○, •), H69PR (C, ⋄, ♦), and 8226/Dox 40 (D, □, ▪). Data points,mean ± range of duplicate determinations.
Osmotic sensitivity of [3H]AFB1-GSH transport by MRP
Initial rates of AFB1-GSH transport were proportional to the relative levels of MRP in different membrane vesicle preparations (Fig. 2), as we have shown previously for LTC4uptake (19). Thus, at a substrate concentration of 250 nm, the initial [3H]AFB1-GSH transport rate (320 pmol/mg/min) in vesicles prepared from the doxorubicin-selected multidrug-resistant H69AR cells (Fig. 2B), which express 4–8-fold more MRP than T14 cells, was significantly higher than in T14 vesicles (Fig.2A). [3H]AFB1-GSH transport rates in vesicles from the drug-sensitive parental small cell lung cancer cell line (H69) and the revertant cell line (H69PR) were low and consistent with their relatively low levels of MRP (Fig. 2C) (26, 27). Rates of transport were somewhat higher in H69PR vesicles than in H69 vesicles, which is in keeping with the slightly higher levels of MRP in the revertant cells (10). The multidrug-resistant 8226/Dox40 myeloma cell line overexpresses moderate-to-high levels of P-glycoprotein (27), and the low levels of transport in vesicles from these cells indicate that AFB1-GSH is at best a poor substrate for P-glycoprotein (Fig. 2D). Because these cells are also known to contain levels of MRP somewhat less than those found in the revertant H69PR cells (27), it is likely that the low level AFB1-GSH transport by 8226/Dox40 vesicles is attributable to MRP rather than to P-glycoprotein. As shown in Fig. 3, MRP-specific mAb QCRL-3 inhibited [3H]AFB1-GSH uptake into T14 vesicles with an IC50 value of ∼0.6 μg/ml, whereas MRP-specific mAb QCRL-1 had no effect at ≤82 μg/ml. These results are in agreement with our previous findings that mAb QCRL-3, which recognizes a conformation-dependent epitope (26), but not mAb QCRL-1, which recognizes a linear epitope (26, 27), inhibited transport of the GSH-conjugated LTC4 and the conjugated estrogen 17β-estradiol 17-(β-d-glucuronide) in membrane vesicles from MRP-transfected cells (19, 25).

Inhibition of [3H]AFB1-GSH transport by MRP-specific mAb QCRL-3. The initial rate of ATP-dependent [3H]AFB1-GSH uptake by T14 membrane vesicles was measured at 120 sec in the presence of affinity purified MRP-specific mAbs QCRL-1 (▪) and QCRL-3 (•). Data were calculated and plotted as percentage of control uptake in the absence of antibody.Points, mean ± standard error of triplicate determinations. The control rate of [3H]AFB1-GSH uptake for this experiment was 93 ± 8 pmol/mg/min.
To further compare AFB1-GSH transport with that of other MRP substrates such as LTC4 and 17β-estradiol 17-(β-d-glucuronide), we examined the ability of several GSH derivatives and steroid glucuronides to inhibit AFB1-GSH transport in MRP-enriched T14 vesicles (Table2). GSSG, which unlike GSH is a substrate of MRP (24), was found to be an effective inhibitor of [3H]AFB1-GSH transport at a concentration of 100 μm, as observed previously for LTC4 and 17β-estradiol 17-(β-d-glucuronide) transport (19-21,23, 25). LTC4 and 17β-estradiol 17-(β-d-glucuronide) compete reciprocally for transport by MRP (25), and despite their lack of structural similarity, both of these substrates also inhibited [3H]AFB1-GSH transport by >85% at concentrations of 10 and 100 μm, respectively. The inhibitory potency of alkylated GSH derivatives on LTC4 transport in hepatocanalicular (33), sarcolemmal (33), and MRP-enriched membranes from transfected and drug-selected cells (19, 20) increases proportionately with the length of the alkyl chain of the derivative. Such was also the case with respect to inhibition of MRP-mediated AFB1-GSH transport by these compounds (Table2). For example, S-methyl GSH (10 μm) did not inhibit transport, whereas S-hexyl GSH andS-decyl GSH at the same concentration inhibited AFB1-GSH transport by 72% and 96%, respectively. Finally, our previous studies with a series of A-ring and D-ring conjugated estrogen glucuronides showed that their ability to inhibit MRP-mediated 17β-estradiol 17-(β-d-glucuronide) transport correlates well with their cholestatic potential (25). In the current study, a similar relative ability of cholestatic D-ring conjugates [17β-estradiol 17-(β-d-glucuronide), 16α,17β-estriol 17-(β-d-glucuronide), and 17β-estradiol 3-sulfato-17-(β-d-glucuronide)] and inability of noncholestatic A-ring conjugates [17β-estradiol 3-(β-d-glucuronide) and 16α,17β-estriol-3-(β-d-glucuronide)] to inhibit [3H]AFB1-GSH transport was observed (Table2). Thus, the profile of transport inhibition is comparable to that described previously with LTC4 and 17β-estradiol 17-(β-d-glucuronide). Overall, our findings demonstrate that MRP is a specific and high affinity transporter of AFB1-GSH, and that it is a likely candidate for a GS-X pump responsible for elimination of this conjugate from mammalian cells, at least in certain tissues.
Inhibition of [3H]AFB1-GSH transport by various organic anions
Although MRP is clearly able to transport molecules with very different structures, the ability of steroid glucuronides to inhibit MRP-mediated 17β-estradiol 17-(β-d-glucuronide) transport was found to exhibit some dependence on the site of conjugation of the steroid nucleus (25). Thus, the 17-(β-d-glucuronide) of 16α,17β-estriol was a very good transport inhibitor, whereas the 3- and 16- β-d-glucuronides of 16α,17β-estriol were poor inhibitors (25). The endo- and exo-AFB1-8,9-epoxides have vastly different DNA-binding and mutagenic potencies (4). To determine whether there were also stereospecific requirements in AFB1-GSH transport, we separated the exo- and endo-[3H]AFB1-GSH conjugates by chiral HPLC and then examined the transport of the individual stereoisomers. The results presented in Fig. 4 show that MRP-mediated AFB1-GSH transport does not display stereospecificity, at least not at the site of GSH conjugation, because the exo- and endo-AFB1-GSH conjugates are transported with similar efficiency.

Time course of [3H]AFB1-exo-8,9-epoxide-GSH and [3H]AFB1-endo-8,9-epoxide-GSH uptake by T14 HeLa cell membrane vesicles. A mixture of [3H]AFB1-exo-8,9-epoxide-GSH (•, ○) and [3H]AFB1-endo-8,9-epoxide-GSH (▪, □) conjugates were separated by HPLC fractionation overd-phenylglycine-aminopropyl silica as described in Experimental Procedures. ATP-dependent uptake of the individual stereoisomers into T14 membrane vesicles was measured at an initial concentration of 15 nm and for the times indicated.Filled symbols, uptake in the presence of 4 mm ATP; open symbols, uptake in the presence of 4 mm AMP; data points, mean ± range of duplicate determinations in a single experiment.
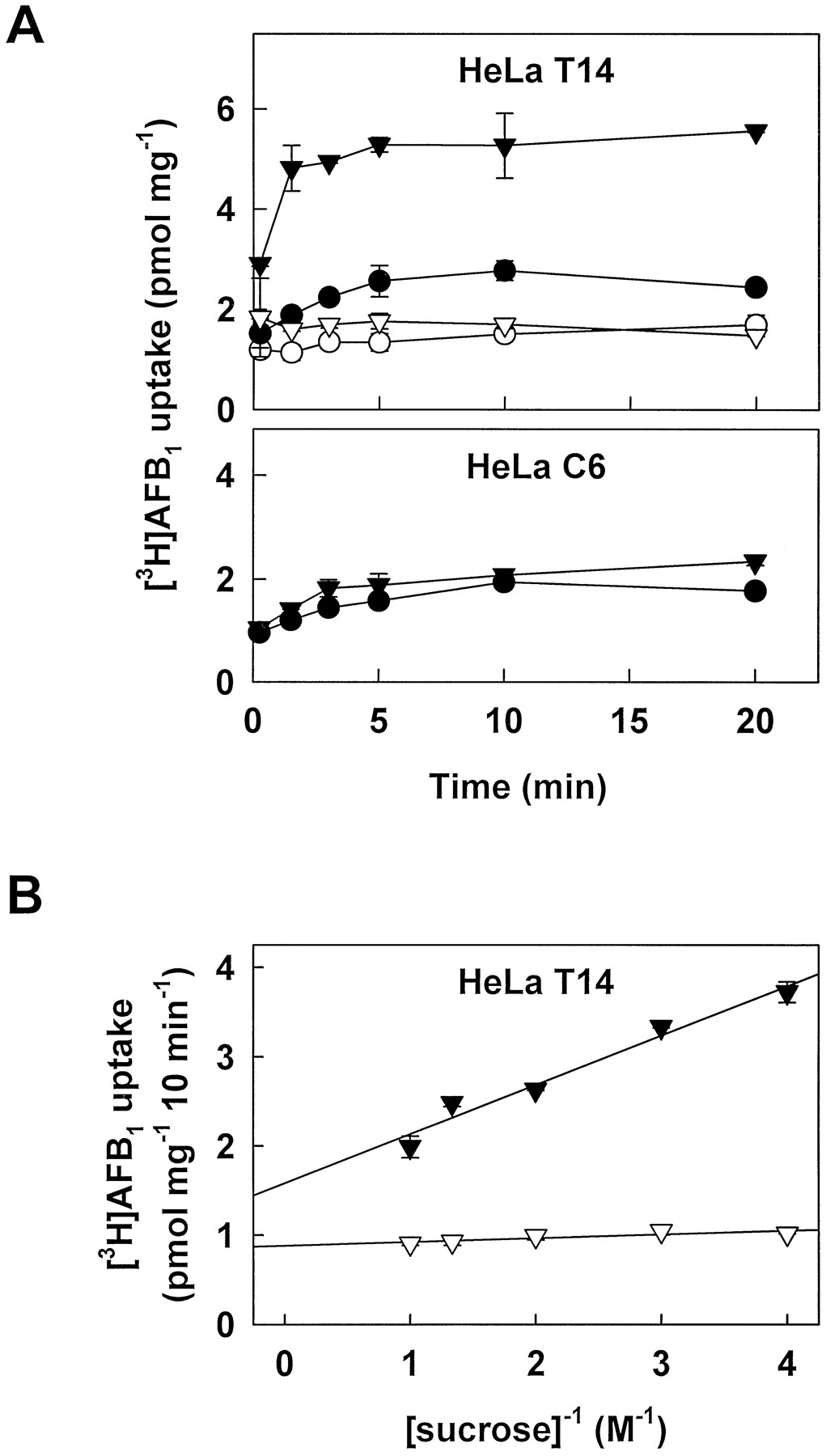
Low levels of ATP-dependent uptake of unmodified [3H]AFB1 were also observed in membrane vesicles from MRP-transfected T14 cells (Fig. 5A) and drug-selected H69AR cells (not shown). Steady state levels of [3H]AFB1 in T14 vesicles in the presence of ATP were ∼2.6 pmol/mg compared with <1.8 pmol/mg in the presence of AMP or in control C6 membrane vesicles with or without ATP (Fig. 5A). Although the steady state levels of vesicle-associated unmodified [3H]AFB1 are considerably lower than those of its GSH conjugates, our data suggest that the presence of the glutathione moiety of AFB1-GSH is not absolutely essential for its transport by MRP. More strikingly, as we reported previously with MRP-mediated transport of unmodified vincristine (19), transport of [3H]AFB1 in T14 vesicles was greatly enhanced by the addition of physiological concentrations of GSH (Fig.5A). In the presence of 5 mm GSH, steady state levels of [3H]AFB1 in T14 vesicles were increased ∼2-fold to 5.5 pmol/mg from 2.6 pmol/mg in the absence of GSH (Fig.5A). This enhancement was not observed in control C6 vesicles. ATP- and GSH-dependent [3H]AFB1 uptake was osmotically sensitive (Fig. 5B) and was completely inhibited by mAb QCRL-3 (not shown). Unlabeled AFB1 (1000-fold molar excess) completely inhibited GSH-dependent [3H]AFB1 uptake in the presence of ATP. However, excess AFB1 did not affect uptake in the presence of AMP or in C6 control membrane vesicles (not shown). Unlabeled AFB1-GSH conjugate (16-fold molar excess) also inhibited [3H]AFB1 uptake by 59 ± 7%. Taken together, these results suggest that the conjugate and the parent compound (in the presence of GSH) may interact with similar or overlapping binding sites in MRP. Consistent with this conclusion, we found that coadministration of AFB1 (100 μm) and GSH (5 mm) inhibited the initial rate of ATP-dependent [3H]AFB1-GSH uptake by 76 ± 1%. In contrast, AFB1 or GSH alone were poor inhibitors at these concentrations (20 ± 4% and 27 ± 3% inhibition, respectively). We also observed that under conditions in which GSH- (and ATP-)dependent [3H]AFB1 uptake by MRP-enriched membrane vesicles was demonstrated, no uptake was observed in P-glycoprotein-enriched membrane vesicles from 8226/Dox40 cells (not shown). Our results thus suggest that P-glycoprotein does not play a significant role in the transport of either unmodified or GSH-conjugated AFB1. Enhancement of MRP-mediated [3H]AFB1 transport by GSH was specific and not simply the consequence of altering the redox state of the protein because it was not observed in the presence of other reducing agents such as 2-mercaptoethanol, dithiothreitol, or l-cysteine at concentrations of ≤5 mm (not shown). The increased transport is also extremely unlikely to be attributable to nonenzymatic formation of [3H]AFB1-GSH because although spontaneous conjugation of GSH is known to take place with certain xenobiotics (6, 34), we found that it does not occur with AFB1 under the experimental conditions used in the current study (not shown). No other AFB1 metabolites were detectable by HPLC analysis in either the aqueous or organic phases of extracts from control incubations with T14 membrane vesicles and [3H]AFB1. Virtually all vesicle-associated radioactivity (>99%) in the organic phase eluted at the identical retention time as the authentic AFB1 standard, confirming that the vast majority of the [3H]AFB1remains unmodified.

Time course and osmotic sensitivity of unmodified [3H]AFB1 uptake by membrane vesicles derived from transfected HeLa cells. A, Top, MRP-transfected T14 HeLa cell membrane vesicles were incubated at 37° with 250 nm [3H]AFB1 (16.9 Ci/mmol) in the presence of 4 mm AMP (○, ▿) or ATP (•, ▾) and in the presence (▿, ▾) or absence (○, •) of 5 mm GSH as described in Experimental Procedures. A, Bottom, control transfected C6 HeLa cell membrane vesicles were incubated with [3H]AFB1 in the presence of 4 mmATP (•) or 4 mm ATP and 5 mm GSH (▾).Data points, mean ± standard error of triplicate determinations in a single experiment. Similar results were obtained in one additional time course experiment; similar steady state levels of vesicle-associated [3H]AFB1 in the presence of GSH were observed in four additional independent experiments. B, T14 HeLa cell membrane vesicles were incubated with [3H]AFB1 as in A in the presence of 5 mm GSH and 4 mm AMP (▿) or ATP (▾) for 10 min in transport buffer containing the indicated concentrations of sucrose. Results are mean ± standard error of triplicate determinations in a single experiment.
How GSH enhances transport of the parent compounds AFB1 and vincristine is unclear. It has been suggested that GSH may be actively cotransported with the unmodified xenobiotic; however, GSH by itself does not seem to be a good MRP substrate (24), nor does it compete for MRP-mediated transport of known substrates LTC4, 17β-estradiol 17-(β-d-glucuronide), or AFB1-GSH. On the other hand, depletion of intracellular GSH has been reported to enhance the toxicity of both vincristine (35, 36) and AFB1 (37). With respect to AFB1, this may simply reflect reduced formation of AFB1-GSH conjugates. However, in addition to GSH, efficient conjugate formation requires GST isoforms with specificity for AFB1-epoxides. In tissues or cell types in which the activity of these isoforms is low, detoxification by MRP-mediated GSH-dependent export of AFB1may be relatively more important. With respect to vincristine, there is no evidence that GSH conjugates of this drug are formed in any tissue. Thus, GSH may interact with MRP in some unknown fashion that favors binding of certain hydrophobic compounds such as vincristine and possibly AFB1 before transport (19).
Elimination of GSH S-conjugates from cells is important because their accumulation can lead to inhibition of GSTs and GSH reductase, which may have deleterious consequences with respect to cellular function and viability. Moreover, certain GSH conjugates can themselves be cytotoxic (2, 6). Multiple transport systems exist for GSH S-conjugates, each with its own distinct but often overlapping substrate and transport characteristics (5, 6). Mammalian ATP-dependent GS-X transporters that have been identified on a molecular level include MRP and the so-called MOAT (38, 39). These two ATP-binding cassette transporter proteins are highly related (48% identical), and although not formally shown, studies with mutant rats in which MOAT is not functional suggest that the two proteins have similar substrate specificities. However, the tissue distributions of MRP and MOAT differ markedly. MOAT is expressed at much higher levels in the liver than is MRP and is present in canalicular membranes, suggesting that MOAT may be more important in hepatic GSHS-conjugate elimination (38). In contrast, MRP is expressed at very low levels in the liver but at relatively high levels in the lung (10, 40, 41), which may indicate a more significant role for MRP in pulmonary GSH S-conjugate export. The lung is composed of many different cell types that vary in their ability to metabolize AFB1 and in their susceptibility to AFB1-mediated mutagenicity and toxicity. This has largely been attributed to differences in levels and isoforms of phase I and phase II biotransformation enzymes (1, 3). However, the different pulmonary cell types contain different levels of MRP, and it is possible that phase III elimination mediated by MRP may also contribute to their differing susceptibilities to AFB1 toxicity.
The phenomenon of multidrug resistance associated with overexpression of P-glycoprotein or MRP is usually thought of in the context of limiting the effectiveness of antineoplastic agents. However, it has long been proposed that one physiological role of P-glycoprotein may be to protect normal tissues from the toxicity of other natural-product xenobiotics, including certain chemical carcinogens (42, 43). The results of the current study demonstrating MRP-mediated high affinity ATP-dependent transport of AFB1-GSH as well as GSH-enhanced transport of unmodified AFB1 in membrane vesicles provide the first direct evidence to suggest that a chemoprotective function may also exist for MRP.
Acknowledgments
We wish to thank Dr. Masayo Yamazaki for helpful discussions and critical reading of the manuscript and Maureen Rogers for secretarial assistance.
Footnotes
- Received December 26, 1996.
- Accepted March 4, 1997.
-
Send reprint requests to: S. P. C. Cole, Ph.D., Cancer Research Laboratories, Queen’s University, Kingston, Ontario, Canada K7L 3N6.
-
This work was supported by grants MT-10519 (S.P.C.C., R.G.D.) and MT-10382 (T.E.M.) from the Medical Research Council of Canada. R.G.D. is the Stauffer Research Professor of Queen’s University, and S.P.C.C. is a Senior Scientist of the Ontario Cancer Foundation.
Abbreviations
- AFB1
- aflatoxin B1
- MRP
- multidrug resistance protein
- mAb
- monoclonal antibody
- GSH
- reduced glutathione
- GST
- GSH-S-transferase
- HPLC
- high performance liquid chromatography
- GSSG
- glutathione disulfide, or oxidized glutathione
- MOAT
- multispecific organic anion transporter
- LTC4
- leukotriene C4
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}