


Abstract
Cytochrome P450 enzymes catalyze the first step of the metabolism and subsequent elimination of hydrophobic xenobiotics. However, the activity of some isoforms, among them CYP1A1 and CYP2E1, may result in cellular insults such as oxidative stress and activation of procarcinogen compounds into reactive metabolites. The regulation of the expression of these enzymes is therefore important. We have previously shown that the CYP1A1 gene promoter was repressed by oxidative stress. We show here that theCYP2E1 gene promoter is down-regulated by exogenous H2O2 addition and glutathione depletion. It is also repressed by the transfection of a CYP2E1 expression vector, which elicits an intracellular H2O2 generation. This autoregulation is limited by catalase (which catalyzes the catabolism of H2O2), thus implying H2O2 as a mediator of the negative feedback mechanism. Furthermore, we observed that the activity of CYP1A1 resulting either from the stimulation of the endogenous gene by benzo[a]pyrene treatment or from the transfection of an expression vector, repressed the activity of theCYP2E1 gene promoter. Conversely, CYP2E1 overexpression repressed the activity of the CYP1A1 gene promoter. In both cases, catalase and a specific inhibitor of one enzyme prevented the repression of the other. This suggests that the generation of H2O2 during the catalytic cycle of these enzymes is a mediator of the cross-regulatory mechanisms. These novel repressive mechanisms of autoregulation and cross-regulation using H2O2 as a common mediator may limit the potential toxicity resulting from high cytochrome P450 activity within the cell.
Cytochrome P450 monooxygenases constitute a multigenic superfamily of enzymes that metabolize both endogenous and exogenous compounds. They play an important role in the metabolism of hydrophobic xenobiotics by initiating a process that leads to their solubilization and elimination. In some cases, however, they activate their substrates into reactive metabolites that can form adducts with proteins or DNA. The activity of cytochromes P450 can thus be implicated in cytotoxic processes. In this respect, it is important to control their expression. Most isoforms involved in the metabolism of xenobiotics are inducible, often by their own substrates. Conversely, several pathophysiological conditions, such as infection and inflammation, repress the expression of several isoforms (Morgan et al., 1998). Extensive studies have been published on the transcriptional regulation of CYP1A1 (Whitlock, 1999), CYP3A4 (reviewed in Guengerich, 1999), and other isoforms including CYP2B and CYP4A (Waxman, 1999).
The expression of the human CYP1A1 gene is mainly controlled by the regulation of its promoter. Its induction by polycyclic aromatic compounds such as 2,3,7,8 tetrachlorodibenzo-p-dioxin or benzo[a]pyrene (BP) is particularly potent. In HepG2 cells, a 100-fold increase was observed (Kress et al., 1998). The mechanism involves the stimulation of the Ah receptor (for a recent review, see Whitlock, 1999). On the contrary, CYP1A1 mRNA levels are depressed by inflammatory cytokines and growth factors (Barker et al., 1992; Muntane-Relat et al., 1995). A common mechanism for these repressions could involve reactive oxygen species (ROS). Indeed, theCYP1A1 gene promoter is inhibited by oxidative stress (Morel and Barouki, 1998).
The human CYP2E1 gene is induced by ethanol ingestion (Wrighton et al., 1986) and has therefore been widely studied. Contrary to other major isoforms, this induction mainly involves post-transcriptional mechanisms. Several potent CYP2E1 inducers, including ethanol, do not induce mRNA synthesis (Carroccio et al., 1994); rather, they increase the translation efficiency (Kim et al., 1990) and the protein half-life. Compared with other isoforms, CYP2E1 normally has a shorter half-life of about 6 h (Yang and Cederbaum, 1997) because it is degraded after ubiquitination by a specific rapid pathway involving the proteasome complex (Tierney et al., 1992; Yang and Cederbaum, 1997). On binding to CYP2E1, some of its substrates stabilize the protein, which then displays a half-life of about 37 h, similar to that of other P450 isoforms (Roberts et al., 1995). For these reasons, the transcriptional regulation of the CYP2E1gene was less studied than that of CYP1A1. However, because inflammatory cytokines (Abdel-Razzak et al., 1993), cause a decrease in CYP2E1 mRNA, we asked whether oxidative stress could regulate theCYP2E1 gene promoter.
ROS are important regulators of cellular functions. For example, they can either induce (Dalton et al., 1999) or repress (Morel and Barouki, 1999) gene expression. ROS generation can be triggered by several cellular stresses, such as mitochondrial dysfunction, inflammation, or UV radiations (Morel and Barouki, 1999). The metabolism of endogenous or xenobiotic compounds is also an important generator of ROS. Indeed, several microsomal cytochromes P450 have been shown to produce ROS in vitro, especially when metabolizing uncoupled substrates. This phenomenon was first observed in intact hepatoma cells with CYP2E1 (Dai et al., 1993). In lymphocytes, the overexpression of several isoforms was also shown to increase ROS production (Puntarulo and Cederbaum, 1998). Moreover, 2,3,7,8 tetrachlorodibenzo-p-dioxin (a potent CYP1A1 inducer) generates an oxidative stress in vivo (Shertzer et al., 1998). We have shown recently that CYP1A1 activity triggers an intracellular H2O2production (Morel et al., 1999). This ROS release by CYP1A1 is involved in negative feedback that limits CYP1A1 induction and the related toxicity.
In this study, we asked whether the catalytic activity of one cytochrome P450 isoform could regulate the gene expression of another isoform. We first showed that both exogenous H2O2 and the catalytic activity of either CYP2E1 or CYP1A1 repress the CYP2E1 gene promoter. We also showed that the activity of CYP2E1 could inhibit theCYP1A1 gene promoter. These regulations constitute a novel cross-regulatory mechanism.
Materials and Methods
Chemicals.
H2O2was used from a 30% stock obtained from Merck (Darmstadt, Germany), Other chemicals were obtained from Sigma (Saint-Quentin Fallavier, France) and oligonucleotides from Genset (Paris, France).
Cell Culture.
The human hepatoma cell line HepG2 was used because the CYP1A1 gene promoter is regulated by oxidative stress (Morel and Barouki, 1998) and because of its good transfection efficiency (Morel et al., 1999). It was maintained at 37°C in an incubator under an atmosphere containing 5% CO2. The medium used in cell cultures was half Dulbecco's modified essential medium and half Ham's F12 (Life Technologies, Paisley, UK), supplemented with 10% fetal calf serum (Life Technologies), 0.5 mg/ml fungizone (Bristol-Myers Squibb, Paris La Défense, France), 100 U/ml penicillin G (Diamant, Puteaux, France), and 100 U/ml streptomycin (Life Technologies).
Plasmids.
The construction of the Firefly luciferase expression plasmid p1A1-FL driven by 1.6 kilobase pairs of the humanCYP1A1 gene promoter has been described previously (Morel and Barouki, 1998). The p2E1-FL vector expresses Firefly luciferase under the control of 1.4 kilobase pairs of the human CYP2E1promoter [[mimus]1342; +32]. This promoter was cloned from human lymphocyte DNA using the 5′CATTGTCAGTTCTCACCTC3′ and 5′GGACACCAGCAGGAGGAAG3′ oligonucleotides as polymerase chain reaction primers and was inserted into the pGL3 vector (Promega, St. Quentin Fallavier, France) between the XhoI andHindIII cloning sites. The Renilla reniformisluciferase expression plasmid pαglob-RL, which contains the proximal promoter of the human α-globin gene, is not sensitive to oxidative stress (Morel and Barouki, 1998) and was used as an internal control of the transfection efficiency. The pcDNA 1.1 AmpR (Invitrogen Gröningen, The Netherlands; also named pCMV/MCS in this study) plasmid was used to express the human CYP1A1 and the rat CYP2E1 cDNAs. The construction of the resulting pCMV-1A1 vector has been reported elsewhere (Morel et al., 1999). As for pCMV-2E1, partial CYP2E1 cDNA lacking the 5′ untranslated region (UTR) was a generous gift of Dr. F. J. Gonzalez (National Cancer Institute, Bethesda, MD). To obtain the whole CYP2E1 cDNA, this partial cDNA was reamplified by polymerase chain reaction using a primer containing the 5′ UTR.EcoRI restriction sites were added in both primers for cloning into pcDNA 1.1 AmpR. The addition of the 5′ UTR allows correct protein expression in HepG2 cells.
Transfection Experiments.
Transfection experiments were performed in HepG2 cells. Briefly, one day before the transfection, cells (0.5 × 106 cells/5 cm dish) were seeded into the usual culture medium. The vectors expressing the Firefly and R. reniformis luciferase genes and the P450 expression vectors were introduced into the cells by the calcium phosphate coprecipitation technique followed 4 h later by a 2-min glycerol shock. Five hours later, cells were treated or not by chemicals added to the culture medium. After incubation, cells were homogenized for enzymatic assays. Dual luciferase assay (Firefly andR. reniformis) was performed with a Promega kit according to the manufacturer's instructions. R. reniformis luciferase activity was used to normalize the transfection efficiency in all culture dishes. Blanks were obtained by assaying luciferase activity in mock-transfected cells. Results were expressed as the ratio [(Firefly luciferase activity) − blank] / [(R. reniformisluciferase activity) − blank].
Intracellular H2O2 Generation Assay.
The oxidation-sensitive probe 2′,7′-dichlorodihydrofluorescein-diacetate is a nonpolar compound that readily diffuses into cells, where it is hydrolyzed by endogenous esterases (Royall and Ischiropoulos, 1993). The resulting compound is not fluorescent but yields the fluorescent 2′,7′-dichlorofluorescein (DCF) when oxidized. Cells were cultured in 6-well plates (Costar, Corning, NY). 2′,7′-Dichlorodihydrofluorescein-diacetate (200 μM) was added directly to the culture medium and cells were cultured in standard conditions for 1 h. The fluorescence of DCF was then measured with a Bio-Tek FL-600 fluorometer (Fisher, Elancourt, France) using 485 nm and 530 nm as excitation and emission wavelengths, respectively. In each well (diameter, 3.5 cm), 109 measurements were made with a 3 mm-diameter optic so as to cover the whole well surface. The result given for each well was expressed as the addition of the 109 values obtained.
CYP2E1 Activity Assay.
HepG2 cells were transfected in 10-cm dishes with 10 μg of either the pCMV/MCS (control) or the pCMV-2E1 plasmid as described above. Transfected cells were homogenized in a Potter apparatus in 0.25 M sucrose, 10 mM Tris, 1 mM EDTA, pH 7.4, and a protease inhibitor cocktail (Roche Diagnostics, Neylan, France). The suspension was centrifuged for 20 min at 9,000g (4°C) then the supernatant was centrifuged for 60 min at 100,000g (4°C). The pellet containing microsomes was resuspended with 100 mM phosphate buffer, pH 7.4, containing 20% glycerol, 10 mM MgCl2, and protease inhibitors. The microsomal protein concentrations were determined by the bicinchoninic acid assay (Pierce Chemical Co., Beigerland, The Netherlands) using serum albumin as standard. The 6-hydoxylation of chlorzoxazone in microsomal preparations of transfected cells was measured according to Carrière et al. (1993).
Statistics.
Student's two-tailed t tests were performed using a Statview software (Abacus Concepts, Inc., Berkeley, CA).
Results
Oxidative Repression of the CYP2E1 Gene Promoter.
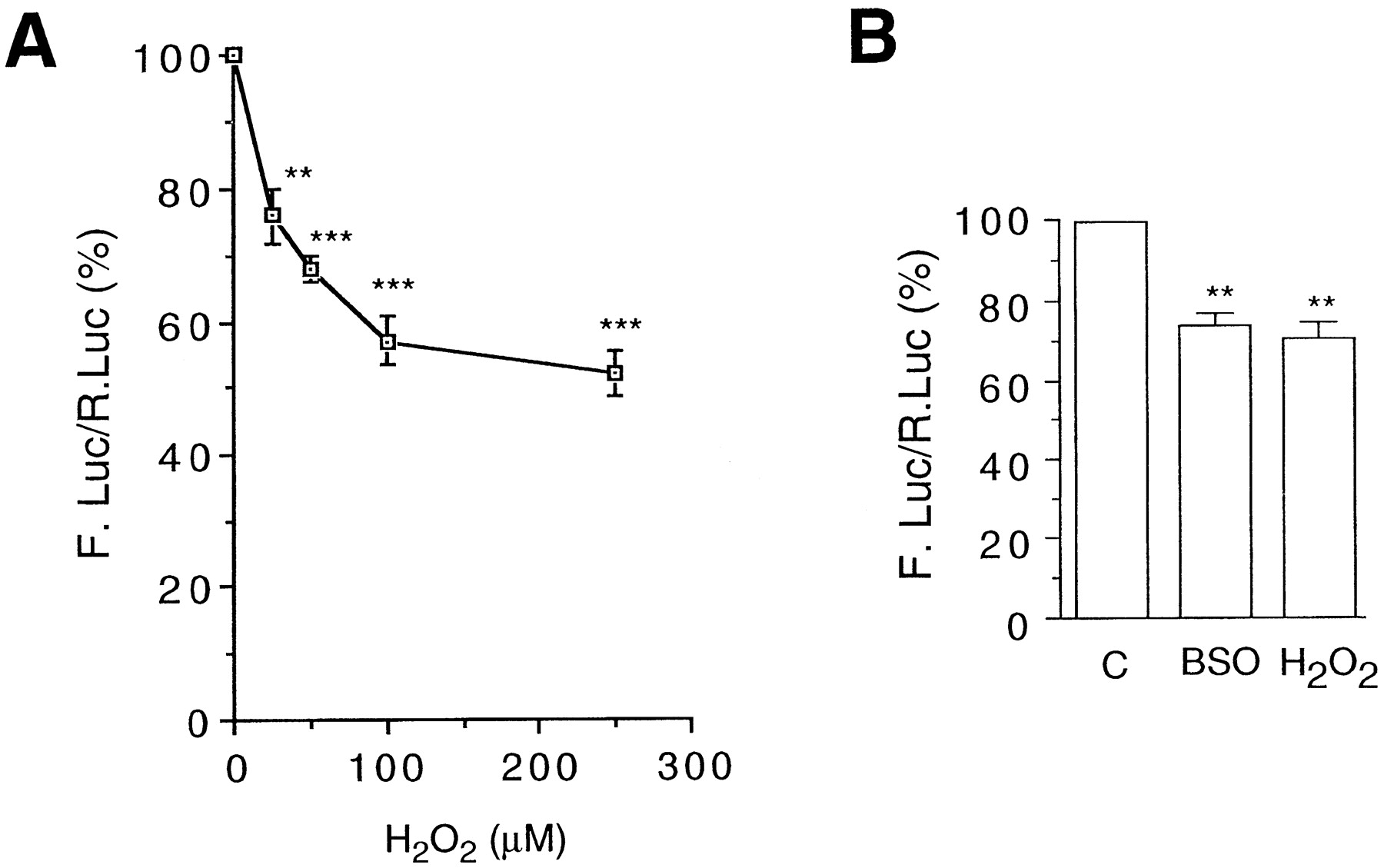
The p2E1-FL vector containing the Firefly luciferase reporter gene driven by the human CYP2E1 gene promoter (seeMaterials and Methods) was used to assay the sensitivity of this promoter to oxidative stress. This plasmid was transfected into HepG2 cells and cultures were treated with hydrogen peroxide as shown in Fig. 1. In a previous study, we have shown that the H2O2concentrations used here do not affect the growth or viability of this cell line (Morel and Barouki, 1998). In addition, these concentrations do not elicit a general modulation of transcription. In particular, the control vector pαglob-RL (see Materials and Methods) used in this study is not affected. Under these conditions, a dose-dependent inhibition of the reporter gene expression was observed (Fig. 1A). The repression was significant at 25 μM and reached 50% at 250 μM H2O2. These results are consistent with previous studies in our laboratory showing the repression of endogenous CYP2E1 mRNA levels following H2O2 treatment of the Fao hepatoma cell line (data not shown).

The CYP2E1 gene promoter is repressed by oxidative stress. HepG2 cells were transfected with p2E1-FL (3.5 μg) and pαglob-RL (1 μg) as a control. The reporter gene activities were assayed as described under Materials and Methods. The results (mean ± S.E., n> 8) are expressed as percent of the activity of untreated cells. Statistical differences with the control values are indicated by **P < .01 or ***P < .0001. A, cell cultures were treated with the indicated amounts of H2O2 and harvested 16 h later. B, cell cultures were treated with 50 μM BSO for 24 h or 50 μM H2O2 for 16 h before harvest.
In addition, we investigated the effect of GSH depletion. It was achieved usingl-buthionine-S,R-sulfoximine (BSO), an inhibitor of γ-glutamylcysteine synthase (the rate-limiting enzyme of GSH synthesis). We showed previously that treatment of HepG2 cells with BSO indeed led to decrease of the intracellular GSH pool (Morel and Barouki, 1998). GSH is an important endogenous antioxidant and its depletion causes an increase in intracellular ROS (data not shown). We observed that BSO treatment caused an average 30% decrease of theCYP2E1 gene promoter activity, reaching an amplitude similar to that obtained with 50 μM H2O2 (Fig. 1B)
Transfection of a CYP2E1 Expression Vector Leads to Microsomal CYP2E1 Activity and Intracellular ROS Generation.
Transfection of HepG2 cells with the pCMV-2E1 plasmid led to the expression of a functional CYP2E1 enzyme as assayed by chlorzoxazone hydroxylase activity [which allows a specific assay of the activity of this cytochrome P450 isoform (Lucas et al., 1999)]. The microsomal fraction of pCMV-2E1-transfected cells displayed such an activity (25.6 ± 16.0 pmol/min/mg of protein, n = 4), whereas that of control pCMV/MCS-transfected cell displayed no detectable activity.
Because some cytochromes P450 can produce H2O2 during their catalytic cycle (see introduction), we next investigated whether the transfection of the CYP2E1 expression vector caused an increased intracellular ROS generation in our system. For this purpose, we used the ROS-sensitive probe DCF (see Material and Methods). As shown in Fig.2, the transfection of the CYP2E1 expression vector in HepG2 cells led to a significant 50% increase in ROS generation (compare bars 1 and 5). As a positive control, we observed in the same experiment that the exogenous addition of 50 μM H2O2 to the culture medium of pCMV/MCS-transfected control cells led to a 60 ± 18% (n = 9) increase in DCF fluorescence after 1 h of treatment (data not shown). This increase is thus similar to that obtained with the transfection of the CYP2E1 expression vector. In pCMV/MCS-transfected cells, ethanol caused a slight induction of DCF fluorescence, which was not significant. On the contrary, the increase of DCF fluorescence after pCMV-2E1 transfection was significantly enhanced in the presence of ethanol (compare bars 5 and 8). In addition, the increase in intracellular ROS production resulting from pCMV-2E1 transfection was not observed when cells were treated with either the H2O2-scavenging enzyme catalase or the CYP2E1 inhibitor 4-methylpyrrazole (4-MP) (Feierman and Cederbaum, 1986), either in the absence of ethanol (compare bars 6 and 7 with bar 5) or in the presence of ethanol (compare bars 9 and 10 with bar 8).
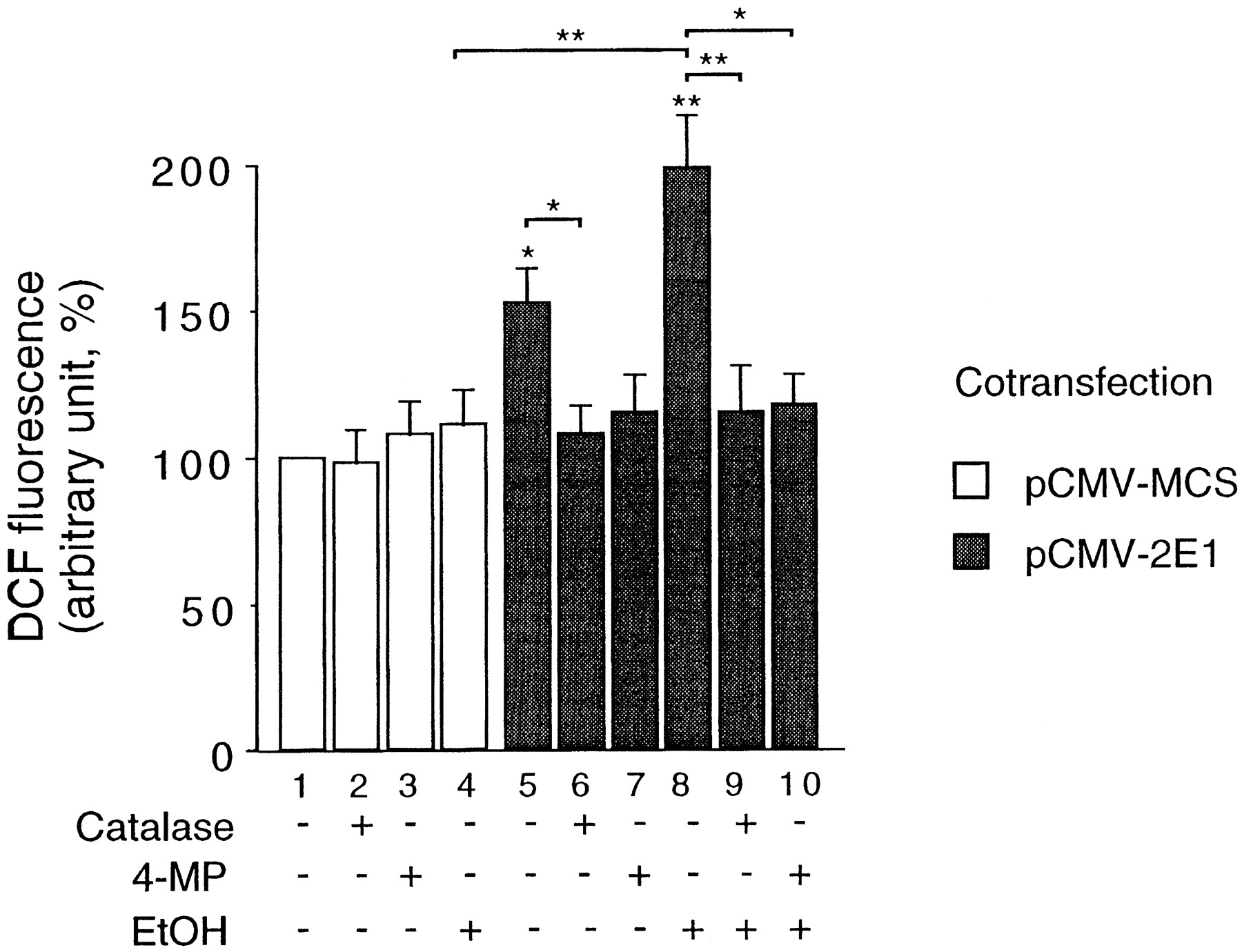
CYP2E1 expression results in intracellular ROS generation. H2O2 levels within HepG2 cells were assayed as described under Materials and Methods. Cells were transfected with either the CYP2E1 expression vector pCMV-2E1 (gray bars) or with the control vector pCMV/MCS (open bars) and cultured for 48 h in the presence or absence of catalase (200 U/ml), 4-MP (5 mM), or ethanol (40 mM) as indicated. Plates were then read in a fluorometer and fluorescence expressed in arbitrary units. Results were expressed as mean ± S.E. (n > 9) from at least three independent transfection experiments realized in triplicates, normalized to 100% for pCMV/MCS-transfected untreated control cells. Statistical differences to this control are spotted above the bars by *P < .05 and **P < 0.005. In addition, statistical differences between certain mean values are shown with brackets and the same *P < .05 and **P < .005 symbols.
Autoregulation of CYP2E1.
Because the activity of CYP2E1 was shown to generate ROS in our cellular model, we asked whether this activity could repress the CYP2E1 gene promoter, as was the case with exogenous H2O2addition. To increase CYP2E1 activity within the cells, we transfected the pCMV-2E1 vector, which expresses the CYP2E1 cDNA. The activity of the CYP2E1 gene promoter was assessed using the p2E1-FL reporter vector. As shown in Fig. 3, the transfection of pCMV-2E1 resulted in a 30% inhibition of the reporter gene activity. This decrease is similar to that observed with 50 μM H2O2 (see Fig. 1A). Interestingly, both procedures (i.e., CYP2E1 expression or 50 μM H2O2 exogenous addition) led to a similar increase in intracellular ROS (see results described above). These data suggest that CYP2E1 activity could repress the promoter of its own gene. The addition of catalase to the culture medium abolished the effect of CYP2E1 overexpression. Therefore, the intracellular generation of H2O2 after CYP2E1 expression seems to be required for the repressive autoregulation.
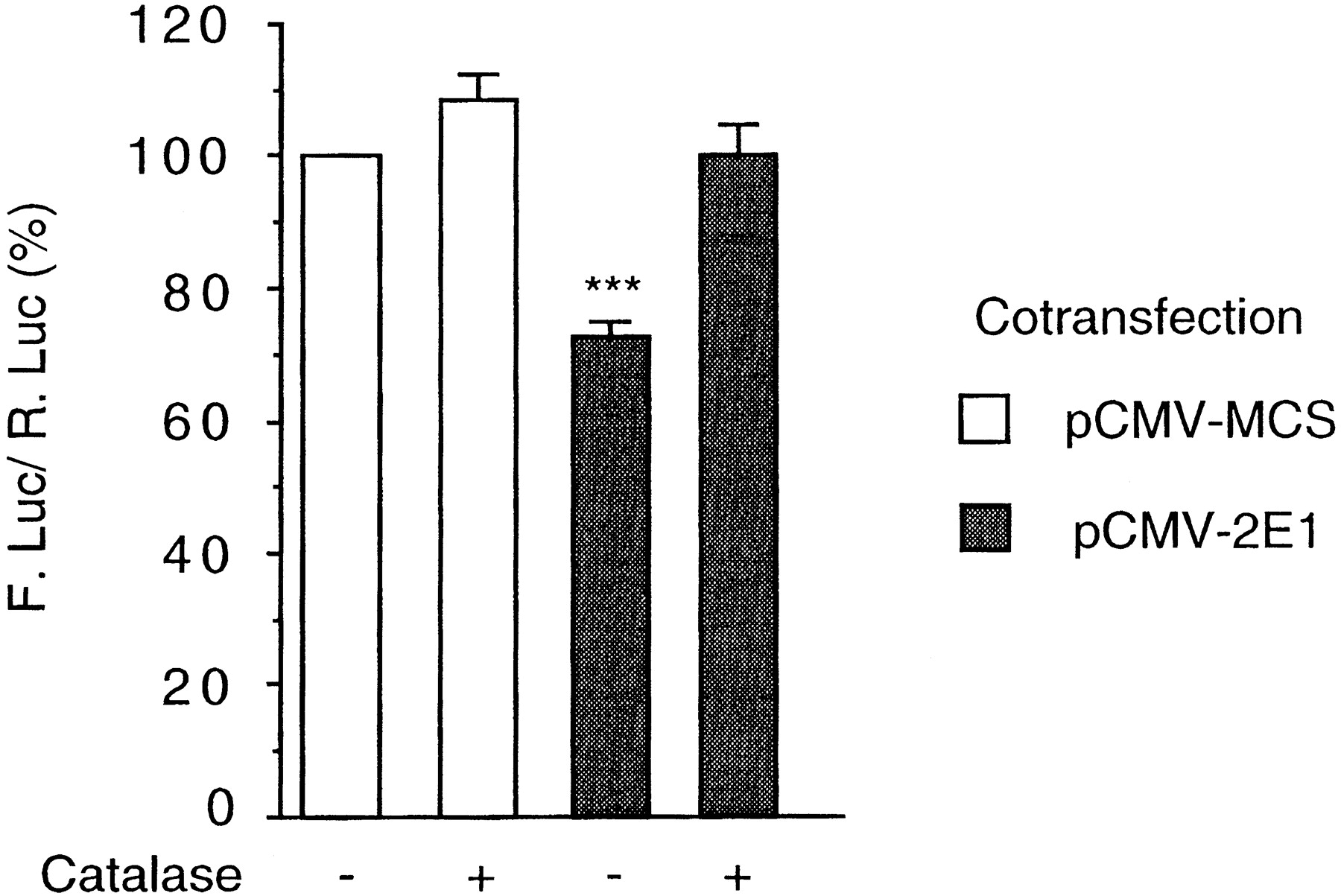
Autoregulation of the CYP2E1 gene. HepG2 cells were transfected with p2E1-FL (2.5 μg) and pαglob-RL (0.75 μg) as a control. Either pCMV/MCS (3.5 μg; open bars) or pCMV-1A1 (3.5 μg; gray bars) was also transfected as mentioned. Cell cultures were treated or not with catalase (200 U/ml) and harvested 40 h later. The reporter gene activities were assayed as described under Material and Methods. The results (mean ± S.E.; n = 8) are expressed as percent of the activity of untreated cells. Statistical differences with the control values are indicated by ***P < .0001.
CYP2E1 Activity Represses the CYP1A1 Gene Promoter.
In a previous study, we have shown that oxidative stress could repress CYP1A1 gene expression at the transcriptional level (Morel and Barouki, 1998). We therefore asked whether the activity of CYP2E1 could repress the promoter of the CYP1A1gene. For this purpose, we used the p1A1-FL reporter vector in which the luciferase gene is driven by the promoter of the humanCYP1A1 gene. As shown in Fig.4, the transfection of the CYP2E1 expression vector caused an almost 40% decrease in CYP1A1 promoter activity (compare bars 1 and 3). This negative effect was totally prevented in the presence of catalase (compare bars 5 and 7), thus implying H2O2 as an intermediate in this regulation. In our experiments, catalase had a slight inducing effect on the activity of the CYP1A1 gene promoter. The scavenging of endogenous H2O2 that is produced during the basal cellular metabolism might cause this effect, but there are no data to support such a hypothesis. In the same experiment, the addition of 4-MP, an inhibitor of CYP2E1, limited the repression of this promoter (compare the decrease between bars 9 and 11 and bars 1 and 3, respectively). These data are consistent with the data presented in Fig. 2 and suggest that CYP2E1 is the actual generator of H2O2.

CYP2E1 activity represses the CYP1A1gene promoter. HepG2 cells were transfected with p1A1-FL (2.5 μg) and pαglob-RL (0.75 μg) as a control. Either pCMV/MCS (2.5 μg; open bars) or pCMV-2E1 (2.5 μg; gray bars) was also transfected. Cell cultures were treated or not with ethanol (40 mM) and with catalase (200 U/ml) or 4-MP (5 mM) as mentioned. Cells were harvested 40 h later. The reporter gene activities were assayed as described underMaterial and Methods. The results (mean ± S.E.;n > 6) are expressed as percent of the activity of untreated pCMV/MCS-transfected cells. In each group of results (bars 1 to 4, 5 to 8, and 9 to 12), statistical differences with the pCMV/MCS-transfected control values (i.e., bars 1, 5, and 9, respectively) are indicated by *P < .05, **P < .01, and ***P < .0001.
The addition of ethanol to pCMV/MCS-transfected cells caused a decrease in CYP1A1 gene promoter activity (compare bars 1 and 2) despite the absence of CYP2E1 activity. Thus, it seems that in the absence of pCMV-2E1 transfection, a CYP2E1-independant ethanol metabolism can result in a limited CYP1A1 gene promoter repression. The repressive effect of ethanol was clearly enhanced by pCMV-2E1 transfection (compare bars 2 and 4). Conversely, ethanol enhanced the pCMV-2E1-mediated reporter gene repression (compare bars 3 and 4). This seems to result from an increased ROS generation caused either by ethanol metabolism and/or CYP2E1 protein stabilization and subsequent increased activity (Roberts et al., 1995). 4-MP also limited the repressive effect of ethanol (compare bars 12 and 4).
In conclusion, the results shown in Fig. 4, support the hypothesis that the intracellular H2O2production resulting from CYP2E1 activity can repress the expression of CYP1A1. The data support the hypothesis that CYP2E1 is the actual ROS-producer and that this production can occur with or without exogenous addition of a CYP2E1 substrate such as ethanol.
CYP1A1 Activity Represses the CYP2E1 Gene Promoter.
Because CYP1A1 was shown to produce H2O2 during its catalytic cycle within HepG2 cells (Morel et al., 1999), we hypothesized that its activity could inhibit the CYP2E1 gene promoter. Increased CYP1A1 activity was achieved either by the transfection of an expression vector or by the induction of the endogenous gene after BP treatment. As shown in Fig. 5, the addition of BP to cell cultures had a repressive effect on theCYP2E1 gene promoter activity (bar 2). The solvent vehicle (dimethyl sulfoxide) had no effect on the reporter gene expression (data not shown). In HepG2 cells, we have previously shown that classical ligands of the Ah receptor such as BP induce the endogenousCYP1A1 gene (Morel and Barouki, 1998; Morel et al., 1999). The transfection of the pCMV-1A1 plasmid expressing the human CYP1A1 cDNA also resulted in a strong decrease of the reporter gene expression (bar 3).

CYP1A1 activity represses the CYP2E1gene promoter. HepG2 cells were transfected with p2E1-FL (2.5 μg) and pαglob-RL (0.75 μg) as a control. Either pCMV/MCS (3.5 μg; open bars) or pCMV-1A1 (3.5 μg; gray bars) was also transfected. Cell cultures were treated or not with BP (2.5 μM) and with catalase (200 U/ml) or ellipticine (2.5 μM) as mentioned. Cells were harvested 40 h later. The reporter gene activities were assayed as described under Material and Methods. The results (mean ± S.E.; n > 6) are expressed as percent of the activity of untreated pCMV/MCS-transfected cells. In each group of results (bars 1 to 3, 4 to 6, and 7 to 9), statistical differences with the pCMV/MCS-transfected control values (i.e., bars 1, 4, and 7, respectively) are indicated by ***P < .0001.
In both cases, the reporter gene expression was restored by catalase addition thus suggesting that the repressive mechanism involves intracellular H2O2generation (compare bars 5 and 6 with bar 4). Moreover, the addition of ellipticine, an inhibitor of CYP1A1 (Lesca et al., 1980), also limited the repressive effect (see bars 7 to 9). In these experiments, catalase and ellipticine alone unexpectedly displayed an inducing effect on the activity of the CYP2E1 gene promoter. However, they clearly prevented the repression of this activity resulting from CYP1A1 expression. These observations suggest that the mechanism involves the production of H2O2 within the cell by CYP1A1. We have previously shown that BP treatment or transfection of a CYP1A1 expression vector both produced a similar amount of ROS within the cell (Morel et al., 1999).
Discussion
In this study, we have shown that the activities of two cytochrome P450 gene promoters (CYP1A1 and CYP2E1) were inhibited by oxidative stress. Exogenous H2O2 addition or intracellular H2O2production resulting from the overexpression of the CYP1A1 and CYP2E1 enzymes themselves seem to trigger the repressive effect. Data from this and from a previous study (Morel et al., 1999) showed that both procedures can indeed lead to similar intracellular ROS production and gene promoter repression. The protective effect observed when catalase was added strongly suggests that H2O2 is involved as a mediator of the repression. It is unclear whether catalase enters the cells; however, in the assays used here, it prevents intracellular ROS production. In addition, the protective effect observed with inhibitors of CYP1A1 and CYP2E1 suggest that these enzymes activity account for the increase in ROS and its consequences on gene expression.
We have used the HepG2 cell line as a model to study the cross-regulation of cytochromes P450 involving H2O2. In these hepatoma cells, the CYP1A1 gene is functional and, as in hepatocytes, is highly inducible. However, HepG2 cells do not express CYP2E1 (this expression was achieved using transfection experiments). Concerning redox regulations, this cell line contains antioxidant systems such as glutathione, which is depleted when γ-glutamylcysteine synthase is inhibited or on treatment with exogenous oxidants. Furthermore, cytokines and other stimuli elicit an ROS generation in this cell line (our unpublished observations). In this article, we have described the involvement of ROS as an endogenous signal modulating gene expression in this particular model. It is consistent with several observations, suggesting that such a mechanism could be relevant in a more physiological context. Indeed, the repression of transfectedCYP promoters by ROS, which is consistent with the decreased levels of the corresponding endogenous mRNA (Morel and Barouki, 1998, and our unpublished observations), could be involved in several physiological regulations. In human hepatocytes primary cultures, inflammatory cytokines such as tumor necrosis factor-α and interleukin-1β were shown to repress several cytochrome P450 monooxygenase mRNAs, including CYP1A1 and CYP2E1 (Abdel-Razzak et al., 1993). ROS are known to be second messengers of such cytokines (Krieger-Brauer and Kather, 1995) and may account for the observed repression. Furthermore, this repressive effect could be relevant for the pattern of CYP2E1 expression along the liver acinus. CYP2E1 is less expressed in the periportal regions, where the oxygen tension is high compared with the perivenous regions (Oinonen and Lindros, 1998). Because H2O2 concentration is correlated with oxygen partial pressure in this organ (Jungermann and Kietzmann, 1997), the oxidative repression of the CYP2E1gene promoter could constitute one mechanism accounting for the distribution of this enzyme. Furthermore, a decrease in microsomal and peroxisomal CYP2E1 has been observed after ischemia-reperfusion, a process that generates oxidative stress (Pahan et al., 1997).
The H2O2-mediated repression of the CYP1A1 and CYP2E1 gene after the expression of CYP1A1 and CYP2E1 could also be part of a biological control mechanism allowing a limitation of cytochrome P450-mediated toxicity. Indeed, these drug-metabolizing enzymes are necessary to eliminate xenobiotics, but their activity may also be deleterious. High CYP2E1 activity may cause ROS generation and subsequent damages to biological macromolecules such as DNA and protein oxidation. In addition, CYP2E1 overexpression was shown to be associated with increased lipid peroxidation, mitochondrial dysfunction, and a high apoptosis rate (Wu and Cederbaum, 1999). CYP2E1-mediated oxidative stress was also associated with collagen production of hepatic stellate cells, and high CYP2E1 activity was associated with pathologies such as steatohepatitis (Weltman et al., 1996) or hepatocellular carcinoma (Tsutsumi et al., 1993). In addition, CYP2E1 can activate some of its substrates into reactive metabolites that can form adducts to macromolecules. For example, it can metabolize small halogenated molecules such as carbon tetrachloride or chloroform into reactive radicals or into phosgene (Aragno et al., 1994). Such metabolites can react directly with the CYP2E1 protein itself (Yang and Cederbaum, 1997). The resulting neoantigens may cause immunotoxicity through the generation of autoantibodies and subsequent hepatic damages such as hepatitis (Eliasson and Kenna, 1996). Furthermore, several studies show that CYP2E1 can activate some molecules into mutagenic compounds (Guengerich and Shimada, 1998).
Similarly, CYP1A1 activity may also lead to toxicity. In HepG2 cells, CYP1A1 generates H2O2(Morel et al., 1999) and increases 8-oxo-guanine formation (Park et al., 1996). CYP1A1 inducers have been shown to trigger oxidative stress in vivo (Shertzer et al., 1998). In addition, CYP1A1 is also a potent activator of mutagenic compounds such as BP (for a recent review, seeGuengerich and Shimada, 1998). Consistently high CYP1A1 activity was associated with an increased risk of lung cancer (Kiyohara et al., 1998).
For all the above reasons, high CYP2E1 and CYP1A1 activity could be deleterious, especially if repair and detoxification mechanisms are defective. The autoregulatory mechanism described here for CYP2E1 and in a previous study for CYP1A1 (Morel et al., 1999), together with the cross-regulations between those two systems, tend to fine tune the expressions of these enzymes. These mechanisms could enable cells to limit protein or DNA damage resulting from high cytochrome P450 monooxygenase activity within the cell. In this respect, it is important to note that resistance to ROS toxicity can be achieved both by the induction of detoxifying enzyme (Schiaffonati and Tiberio, 1997) as well as by the repression of ROS-producing ones. The data presented in this study suggest that H2O2 could be an important mediator of the mechanisms involved in these regulations.
The model of cross-regulation between CYP1A1 and CYP2E1 proposed here relies mostly on experiments using transfection assays in hepatoma cells. However, it is consistent with previous studies describing unexpected drug-drug interactions and provides a molecular mechanism for those observations. For example, it has been shown in vivo in rat liver that CYP1A1 inducers (such as 3-methylcholanthrene, β-naphthoflavone, or the polychlorinated biphenyl Aroclor 1254) caused a decrease in rat hepatic CYP2E1 mRNA and protein levels (Borlakoglu et al., 1993; Goasduff et al., 1995) and inhibited the metabolism of dimethylnitrosamine (Kornbrust and Dietz, 1985) and benzene (Schrenk et al., 1996), two typical CYP2E1 substrates. It would be interesting to investigate whether other drug-drug interactions could be mediated by similar mechanisms.
Little is known about the promoter of the CYP2E1 gene. Putative regulatory sequences were described in the rat sequence such as hepatocyte nuclear factor HNF1 binding sites (Liu and Gonzalez, 1995), but the extrahepatic expression of the gene suggests that other factors may be important. Owing to the lack of transcriptional induction by classical CYP2E1 inducers, few studies have addressed the regulation of the promoter activity. The data presented here suggest that this promoter activity is regulated by oxidative stress. In the case of the CYP1A1 gene, we have shown that TNFα and oxidative stress repressed the promoter activity, most likely through the oxidative repression of the Nuclear Factor I (NFI) transcription factor (Morel and Barouki, 1998). This does not preclude a role of redox-sensitive kinases or phosphatases. Concerning theCYP2E1 gene, the transcriptional mechanisms of its repression remain unknown. Interestingly, a conserved NFI site is located in the promoter region of the CYP2E1 gene, just down-stream of the TATA box motif. This sequence binds NFI proteins (our unpublished observations). Additional studies are required to assess the potential role of NFI or other factors in the regulation of the CYP2E1 gene promoter by oxidative stress.
Acknowledgments
We are very thankful to Dr. F. Berthou for the determination of chlorzoxazone hydroxylase activities and to Dr. P. Maurel for the gift of the CYP1A1 gene promoter.
Footnotes
-
Send reprint requests to: Dr. Robert Barouki, Institut National de la Santé et de la Recherche Médicale U490, Université Paris V-René Descartes, Centre Universitaire des Saints-Pères, 45, rue des Saints-Pères, 75006 Paris, France. E-mail: robert.barouki{at}biomedicale.univ-paris5.fr
- Abbreviations:
- BP
- benzo[a]pyrene
- ROS
- reactive oxygen species
- UTR
- untranslated region
- DCF
- 2′,7′-dichlorofluorescein
- BSO
- l-buthionine-S,R-sulfoximine
- 4-MP
- 4-methlpyrrazole
- NFI
- nuclear factor I
- Received October 5, 1999.
- Accepted February 17, 2000.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}