


Abstract
Vascular smooth muscle cells (VSMC) in situ function to control contraction and are said to express a contractile phenotype. However, during development or in response to vascular damage, VSMC proliferate and express a more synthetic phenotype. A survey of literature values for contractile and synthetic VSMC phosphodiesterase (PDE) 3 and PDE4 activities identified a marked difference in the PDE3 and PDE4 activities of these cells. In this study, a comparison of PDE3 and PDE4 activities in contractile and synthetic VSMC demonstrates that a reduced PDE3/PDE4 activity ratio in synthetic VSMC correlates with a reduced PDE3 activity and is associated with marked reductions in PDE3A mRNA and protein levels. Because we show that similar reductions in PDE3 activity and PDE3A levels occur upon culture of human aortic VSMC and that this phenomenon associates with the phenotypic switch that occurs to VSMC in response to vascular damage, our findings are presented in the context that PDE3 inhibition might be expected to selectively alter functions of contractile VSMC.
In healthy blood vessels, vascular smooth muscle cells (VSMC) express a specific set of contractile proteins, exhibit low proliferative or migratory capacities, and control blood vessel contractions (Owens, 1998; Thyberg, 1998). In contrast, during development, in response to vascular damage, or during angiogenesis, VSMC express significant extracellular matrix and are highly proliferative and migratory (Owens, 1998; Thyberg, 1998). The similar phenotypic switch that occurs when VSMC are cultured make cultured VSMC a commonly used model of synthetic VSMC (Li et al., 1999; Worth et al., 2001).
Increases in cAMP or cGMP influence relaxation-contraction coupling, proliferation, migration, and cellular metabolism of VSMC (Rybalkin and Bornfeldt, 1999; Lucas et al., 2000; Koyama et al., 2001). Amounts of VSMC cAMP and cGMP are coordinately regulated by their synthesis by adenylyl cyclases and guanylyl cyclases, respectively, and their hydrolysis by cyclic nucleotide phosphodiesterases (PDE) (Dousa, 1999;Soderling and Beavo, 2000). Eleven distinct mammalian PDE families have been described, and activities of five of these families have been detected in homogenates of blood vessels or of cultured arterial VSMC (Polson and Strada, 1996; Rybalkin and Bornfeldt, 1999). Although members of the PDE1 and PDE5 families hydrolyze VSMC cGMP, VSMC cAMP is inactivated by members of the PDE1, PDE3, and PDE4 families (Polson and Strada, 1996; Degerman et al., 1997; Rybalkin et al., 1997; Houslay et al., 1998; Corbin and Francis, 1999; Kakkar et al., 1999; Palmer and Maurice, 2000). Although PDE3 and PDE4 activities account for greater than 80% of the cAMP PDE activity in VSMC isolated from intact blood vessels (contractile VSMC) or from cultured VSMC (synthetic VSMC), potentially important differences in the relative abundance of these activities have been noted in the two VSMC phenotypes (Polson and Strada, 1996). Thus, whereas the ratio of the percentage PDE3 and PDE4 activities (PDE3/PDE4 ratio) in contractile VSMC is about 1.5, the PDE3/PDE4 ratio is usually about 0.8 when measured in cultured VSMC.
In this report, we demonstrate that a marked reduction in PDE3 activity and in the levels of PDE3A mRNA and protein accompanies the phenotypic switch associated with culturing of rat and human aortic VSMC. In addition, we demonstrate that the synthetic VSMC that accumulate in the intimal layer of rat aorta after balloon-induced damage also have reduced PDE3 activity and PDE3A protein compared with medial layer VSMC. Our data is presented in the context of continuing efforts to use PDE3 inhibitors to limit cardiovascular diseases (Haslam et al., 1999;Movsesian, 1999; Osinski and Schror, 2000; Park et al., 2000; El Beyrouty and Spinler, 2001; Tanabe et al., 2001).
Materials and Methods
Cell Culture.
Establishment of primary cultures of rat aortic VSMC (P0), and their subculture were essentially as described previously (Rose et al., 1997). Briefly, for each study, four male Wistar rats (250–300 g) were sacrificed and their thoracic aortae were removed. After mechanical removal of the endothelial and adventitial layers, medial layers of these aortae were pooled, finely minced, and incubated for 2 to 4 h in 2 ml of Hanks' balanced salt solution (HBSS) supplemented with 100 units/ml of penicillin, 100 μg/ml streptomycin and collagenase (Sigma, 2 mg/ml) at 37°C in a 95% air-5% CO2 humidified atmosphere. The enzymatically dissociated VSMC were collected by centrifugation (1000g, 5 min), resuspended in 3 ml of Dulbecco's modified Eagle's medium supplemented with calf serum (20%, v/v), 100 units/ml of penicillin, 100 μg/ml streptomycin, seeded in a 25-cm2 flasks and kept at 37°C in a humidified atmosphere saturated with 95% air/5% CO2. At day 3, attached VSMC (usually about 50% of those plated) were given fresh medium and cultured to confluence (3–6 days). At confluence, (3–4 × 106 cells/25 cm2 flask), the primary VSMC culture (P0) was rinsed with HBSS and the VSMC were detached from the flask by incubation with 0.05% trypsin and 0.53 mM EDTA. Although one third (1 × 106 VSMC) of the detached P0 VSMC were seeded in three 25-cm2 flasks (about 330,000 VSMC/flask) and allowed to grow to confluence (4–7 days, P1), the remaining 2 × 106 VSMC were divided such that equal amounts were lysed in a cAMP PDE lysis buffer (Rose et al., 1997), or TRIzol reagent (see below). The VSMC lysed in the cAMP PDE lysis buffer were used for cAMP PDE assays and immunoblotting, while VSMC lysed in TRIzol reagent were used to isolate RNA. When the cultures in the three 25-cm2 flasks' (i.e., derived from the P0 VSMC) were confluent (referred to as P1), one flask was lysed in cAMP PDE lysis buffer, one flask was lysed in TRIzol reagent, and one flask was used to establish a further three VSMC cultures in 25-cm2 flasks. These procedures were repeated until VSMC P5 cultures were obtained for analysis. To ensure that VSMC cAMP PDE activities at different passage numbers could be compared directly, VSMC were always lysed when the cultures had reached confluence, a point at which rat aortic VSMC are contact inhibited (Liu et al., 2000).
Cyclic Nucleotide Phosphodiesterase Activity Assay.
cAMP PDE specific activities were determined by a modification of Davis and Daly (1979) as described previously (Rose et al., 1997), using 1 to 5 μg of VSMC protein and at 1 μM cAMP as substrate. Data were expressed as picomoles per minute per milligram of protein with protein concentrations determined using the bicinchoninic acid protein assay (Pierce, Rockford, IL) according to the manufacturer's protocol. Cilostamide (1 μM), Ro 20-1724 (10 μM), or IBMX (100 μM) were used to determine PDE3, PDE4, or “total” cAMP PDE activities, respectively.
Immunoblotting.
VSMC lysates were boiled in sample buffer and aliquots (5–30 μg) were subjected to SDS-polyacrylamide gel electrophoresis and transferred to nitrocellulose membranes (Bio-Rad, Hercules, CA). Immunoreactivity was determined as described previously (Liu and Maurice, 1998) using two antibodies: a monoclonal anti–β-actin antibody (Sigma) and a pan-reactive PDE3 antiserum raised against murine PDE3 (a generous gift of Dr. J. A. Beavo, Department of Pharmacology, University of Washington, Seattle, WA; Zhao et al., 1997). Although this antiserum recognizes both PDE3A and PDE3B, these proteins are easily distinguished on a gel based on their distinct sizes. Immunoreactive proteins were visualized using horseradish peroxidase-conjugated secondary antisera, and an enhanced chemiluminescence kit (Amersham, Baie d'Urfé, QC, Canada). Amounts of β-actin, or PDE3A and PDE3B, were determined by scanning densitometry using Corel Photo-Paint 8.0 software (Corel, Ottawa, ON, Canada) per manufacturer's recommendations as described previously (Liu and Maurice, 1998, 1999).
Reverse Transcriptase Polymerase Chain Reaction.
RNA isolation was carried out using TRIzol reagent as per manufacturers recommendations, and reverse transcriptase-PCR was carried out as described previously (Liu and Maurice, 1998). Briefly, reverse transcription was catalyzed by SuperScript Moloney murine leukemia virus reverse transcriptase using 10 μg of total RNA and oligo(dT)18 primers. Amplification was catalyzed by Taq DNA polymerase using 0.01 of the reverse transcription product and 20 pmol each of sense and antisense primers. Oligonucleotide primers used in these experiments included those for rat PDE3A (5′-CCGAATTCCCTTATCATAACAGAATCCACGCCACT-3′; 5′-GGGAATTCGTGTTTCTTCAGGTCAGTAGCC-3′), for rat PDE3B (5′-CCGAATTCTATCACAATCGTGTGCATGCCACAGA-3′; 5′-CCGAATTCTTTGAG ATCTGTAGCAAGGATTGC-3′), and for rat β-actin (5′-TCAGAAGGACTCCTACGTGGGCGAC-3′; 5′-GCATGAGGGAGCGCGTAACC-3′). These primer pairs allowed amplification of selective products encoding either PDE3A (508 bp), PDE3B (499 bp) or β-actin (281 bp). Conditions used for PCR were optimized to allow linear amplification of the β-actin or PDE3A or PDE3B gene products and these were determined as described previously (Liu and Maurice, 1998). PCR products were separated on 1.5% agarose, visualized by ethidium bromide under UV light, and quantified after digital photography and analysis of digital images, as described previously (Liu and Maurice, 1999).
Balloon-Catheter Induced Aortic Damage.
Male Wistar rats (300 g) were anesthetized using a ketamine/water/xylazine cocktail (1:2:1). An abdominal incision was made, and a 2 French Fogarty balloon catheter (Baxter, Deerfield, IL) was inserted into the left iliac artery. Once in position, the balloon was maximally inflated using air (200 μl) and passed three times through the abdominal portion of the aorta. After removal of the balloon catheter, the left iliac artery was sutured closed and the rats were allowed to recover in individual cages. After 14 days, rats were sacrificed and abdominal aortae were removed. The isolated aortae were rinsed free of blood, mechanically cleared of fat and connective tissue, and cut longitudinally into three equal portions using a sharp blade. The three aortic segments were separated and used either for cAMP PDE activity determinations, immunoblot analysis, or immunohistochemical studies. The cAMP PDE activity assays and immunoblotting were carried out as described above and the procedures used for immunohistochemistry are described below.
Immunohistochemistry.
Blood vessel segments were fixed with 4% paraformaldehyde and embedded in paraffin, and slides were prepared from successive aortic slices (Core Laboratory, Department of Pathology, Queen's University, Kingston, ON, Canada). Paraffin was removed by successive incubations in toluol, absolute ethanol, distilled water, and 0.1 M Tris. To maximally expose antigens from fixed tissue segments, slides were rinsed in water, heated to boiling twice for 5 min in 0.1 mM EDTA, pH 8.0. After a cooling period (30 min), aortic segments were rinsed with 0.1 M Tris, incubated with bovine serum albumin [10% (w/v), 1 h], and incubated with the primary antibody of interest (30 μg/ml, 48 h, ambient temp), or 4′,6′-diamidino-2-phenylindole dihydrochloride, hydrate (DAPI). Antisera used included a polyclonal anti-PDE3 antiserum raised in rabbit using a murine PDE3B fragment that reacts equally with both PDE3A and PDE3B (Zhao et al., 1997) and a monoclonal anti–β-actin antiserum (Sigma, St. Louis, MO). After incubation with the antiserum, segments were rinsed with 0.1 M Tris, incubated with a biotin-conjugated goat anti-rabbit antiserum (1:200 dilution), rinsed with 0.1 M Tris, incubated with 5.6 μl/ml streptavidin-conjugated dichlorotriazinyl amino fluorescein (Jackson Immunoresearch, West Grove, PA) for 30 min, rinsed in 0.1 M Tris and with anti-fade reagent (Molecular Probes, Eugene, OR), and covered with cover slips. Stained sections were viewed on an Axiovert S100 fluorescent microscope (Zeiss, Welwyn Garden City, UK) and digital images of the sections were captured using a SensiCam high performance CCD camera (Optikon, Kitchener, ON, Canada). To create presentation images, image files were transferred to Adobe Photoshop 6.0 software (Adobe Systems, Mountain View, CA). Intrinsic elastin fluorescence was removed by subtracting a mask of this fluorescence, generated from an aortic slice processed without antiserum using the mask-editing function of the Slidebook digital microscopy software (Intelligent Imaging Innovations, Denver, CO).
Isolation of Intimal VSMC after Balloon Injury.
Healthy rat aortae contain an intima composed of a single layer of endothelial cells, with no VSMC. VSMC that had accumulated in the intimal layer after balloon-induced damage were isolated by a controlled trypsinization of isolated intact aortae. Thus, the lumen of intact excised blood vessels were exposed to 0.05% trypsin and 0.53 mM EDTA in a Ca2+/Mg2+ free HBSS solution (4 × 50 μl). After these incubations, the intimal layer derived tissues from four individual aortae were pooled; intimal VSMC were collected by centrifugation and lysed in cAMP PDE lysis buffer. For these experiments, these samples are defined as intimal VSMC. Amounts of intimal tissue recovered by this protocol were usually in the range of 40 to 60%, based on comparisons of intimal thickness before and after trypsin-EDTA treatment. Residual aortic tissues, which were composed primarily of medial tissue and of some residual intimal tissue, were also lysed in cAMP PDE lysis buffer. These samples are defined as medial VSMC samples.
Results and Discussion
cAMP PDE Activity in Contractile and Synthetic Rat Aortic VSMC
VSMC in situ, which normally express a contractile phenotype, can adopt a synthetic phenotype in response to vascular damage or when cultured (Owens, 1998; Thyberg, 1998; Li et al., 1999; Worth et al., 2001). Interestingly, a literature review of PDE3 and PDE4 activities in arterial VSMC (contractile phenotype) and cultured arterial VSMC (synthetic phenotype) reported differences in the relative percentages that PDE3 and PDE4 (PDE3/PDE4 ratio) represented in these two types of VSMC (Polson and Strada, 1996). Indeed, although contractile VSMC had a higher percentage of PDE3 activity than PDE4 activity (60 versus 30%), the opposite was true of synthetic VSMC (20 versus 70%) (Polson and Strada, 1996). We surmised, based on these data, that PDE3 and/or PDE4 activities might be regulated in a VSMC phenotype-dependent fashion and that this effect might alter the impact of PDE3 or PDE4 inhibition in contractile and synthetic VSMC phenotypes. Many of our data are consistent with this proposal.
Because changes in PDE3, PDE4, or both PDE3 and PDE4 activities could have contributed to differences in the PDE3/PDE4 ratio observed between contractile and synthetic VSMC, we first measured PDE3 and PDE4 activities in rat aortic VSMC before culture (contractile phenotype) and at each of a series of passages in culture (synthetic phenotype). In these experiments, synthetic rat aortic VSMC were shown to have a significantly lower cAMP PDE specific activity than was detected in contractile rat aortic VSMC (Fig. 1A). Based on the effect of cilostamide (a PDE3-selective inhibitor), or Ro 20-1724 (a PDE4-selective inhibitor), synthetic VSMC had a significantly lower level of PDE3 activity than was measured in contractile VSMC. When measured at different passages, PDE3 activity was shown to be maximally decreased between P1 and P2. In contrast to these results, no differences in PDE4 activity or in the residual nonPDE3/PDE4 activity, as assessed using the broad-selectivity inhibitor (IBMX), were seen. In fact, when IBMX-mediated inhibition was taken as a measure of total cAMP PDE activity, the decrease in PDE3 activity observed in these experiments accounted for the entire difference between contractile and synthetic rat aortic VSMC (Fig. 1A).

Different levels of PDE3 activity and PDE3A expression in rat aortic medial layer-derived VSMC and cultured rat aortic VSMC. Wistar rat aortic medial layer-derived tissue containing contractile VSMC was either lysed in cAMP PDE assay buffer (10% of total tissue, referred to as aorta), or incubated with collagenase to allow dissociation of medial VSMC (Materials and Methods). Enzymatically dissociated medial VSMC were used to establish a primary culture of rat aortic VSMC (P0). At confluence, P0 VSMC were isolated with trypsin-EDTA, and used to establish P1-P5 VSMC cultures. A, cAMP PDE specific activity (no inhibitor), PDE3 activity (1 μM cilostamide), or PDE4 activity (10 μM Ro 20-1724) were determined in lysates of VSMC as described in Materials and Methods. Data are presented as means ± S.E. of three individual experiments, each of which was carried out in triplicate. B, reverse transcription-PCR–based determination of levels of PDE3A or PDE3B encoding mRNA in Wistar rat aortic medial layer-derived tissue (referred to as aorta) and in consecutive passages of VSMC (P0–P5) using selective PDE3A or PDE3B oligonucleotide primers was carried out as described under Materials and Methods and in Liu and Maurice (1998). Shown are means ± S.E. of three experiments in which amplified amounts of PDE3A or PDE3B were corrected for amounts of a control mRNA (β-actin). C, representative immunoblot of three experiments in which levels of PDE3A and PDE3B were determined. Lysates of rat aortic medial layer-derived tissue (rat aorta, lane 1) or of P5 cultures of rat aortic medial layer-derived VSMC (lane 3) were determined by immunoblot analysis using a PDE3 antibody. Lysates of rat epididymal fat-derived tissue (lane 2) and subcellular fractions of P5 cultures of rat aortic medial layer-derived VSMC (lanes 4 and 5) were used as controls.
PDE3A mRNA and Protein Are Decreased in Synthetic Rat Aortic VSMC
In a series of earlier papers, we showed that rat and human aortic VSMC, and VSMC cultured from each of these aortae, expressed both PDE3A and PDE3B (Maurice et al., 1995; Liu and Maurice, 1998; Palmer and Maurice, 2000). To establish whether synthetic VSMC had a generalized reduction in PDE3, or if the reduced PDE3 activity was reflective of a PDE3 isoform-specific effect, we measured mRNA and protein levels for both PDE3A and PDE3B in contractile and synthetic rat aortic VSMC. Our data was consistent with a marked reduction in the levels of PDE3A mRNA and protein in synthetic VSMC compared with that detected in contractile rat aortic VSMC (Fig. 1, B and C). Thus, although synthetic rat aortic VSMC showed a marked reduction in PDE3A mRNA from their preculture status to passage 2 (P2), no change in PDE3B mRNA was observed over this transition (Fig. 1B). Consistent with these results, immunoblots of VSMC lysates showed that synthetic VSMC expressed markedly less PDE3A protein than contractile VSMC, although the low level of PDE3B was not different between synthetic and contractile VSMC (Fig. 1C). The lack of observed difference in PDE4 activity between contractile and synthetic VSMC, was supported by the fact that no difference in the level of PDE4D mRNA or PDE4D3 protein, the major PDE4D expressed in these cells (Liu and Maurice, 1999), was detected in our experiments (data not shown).
Because PDE3B mRNA and protein levels were not different between contractile and synthetic VSMC, and no change in PDE4 activity or PDE4D3 level was noted in our experiments, we conclude that the decreased PDE3/PDE4% activity ratio observed in synthetic VSMC is most probably caused by a decrease in PDE3A level in synthetic VSMC. A similar decrease in cardiac PDE3A has been noted to be associated with development of decompensated dilated cardiomyopathy (Smith et al., 1997). The authors of this earlier report suggested that the reduced PDE3A in cardiomyocytes could account, in part, for the ineffectiveness of PDE3 inhibitors, such as milrinone, in the treatment of severe congestive heart failure. Whether similar mechanisms are involved in reducing PDE3A expression in synthetic VSMC and diseased cardiomyocytes awaits further study.
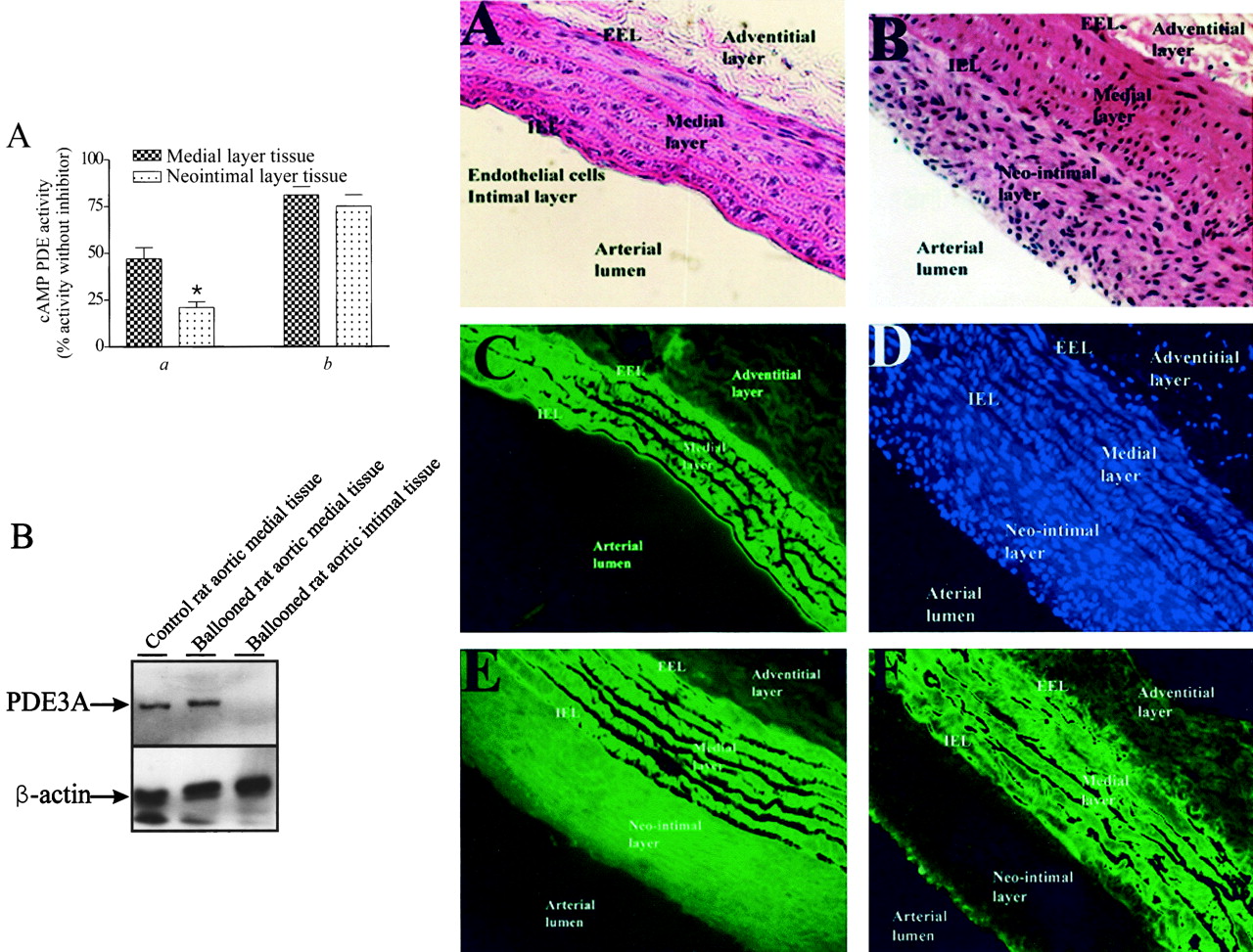
Rat Aortic Intimal VSMC Have Reduced PDE3 Activity and PDE3A Protein Levels
As stated above, the synthetic phenotype expressed by cultured VSMC has often been used as a model of the synthetic phenotype expressed by VSMC that accumulate in blood vessels after damage (Owens, 1998). In fact, several pharmacological approaches designed to reduce the accumulation of intimal VSMC and reduce the incidence of intimal lesions and cardiovascular disease are based on results obtained with cultured synthetic VSMC (Rybalkin and Bornfeldt, 1999). To assess whether the reduced PDE3 activity and PDE3A protein level that accompanied the development of a synthetic phenotype in subcultured VSMC also accompanied the VSMC phenotypic switch in vivo, PDE3 activity and PDE3A protein levels were determined in neointimal tissues isolated from balloon-damaged aortae. It has been established previously that neointimal tissue is composed primarily of dedifferentiated medial VSMC (Owens, 1998). Our results suggest that a reduction in the level of PDE3A accompanies the vascular-damage–induced in vivo phenotypic switch of rat aortic VSMC (Fig. 2A-C). Thus, intimal VSMC PDE3 activity was significantly lower (about 60%) than that detected in medial VSMC isolated from damaged rat aortae (Fig. 2A) or from undamaged aortae (compare Figs. 1A and 2A). Immunoblot analysis of medial and intimal VSMC isolated from ballooned aortae showed that intimal VSMC expressed markedly less PDE3A than was detected in medial VSMC isolated from these aortae (Fig. 2B). In fact, in our experiments, no PDE3A immunoreactivity was detected even though similar amounts of β-actin were present (Fig. 2B). Regrettably, amounts of intimal and medial VSMC obtained from ballooned aortae were too low for the level of PDE3B to be determined. Thus, although it is tempting to speculate the residual PDE3 activity present in intimal VSMC (Fig. 2A) represents activity derived from PDE3B expressed in these cells, further experiments will be needed to confirm this supposition. Consistent with the activity measurements (Fig. 2A), and with results obtained using synthetic rat aortic VSMC generated by our subculturing protocol, levels of PDE4 activity (Fig. 2A) and of PDE4D3 protein expressed in intimal and medial derived VSMC were similar (data not shown).

Different levels of PDE3 activity and PDE3A expression in rat aortic medial layer VSMC and in intimal VSMC formed in response to balloon-induced vascular damage. Abdominal aortae of rats were damaged by inflation of 2 French Fogarty balloon catheters that had been introduced into the aortae via the left iliac artery as described under Materials and Methods. After 14 days, aortae were removed and processed for analysis. A, a, comparison of percentage of total specific cAMP PDE activity inhibited by the PDE3-specific inhibitor cilostamide (1 μM) in intimal layer and medial layer-derived VSMC (*, p < 0.05;n = 3) after tissue isolation by trypsin treatment as described under Materials and Methods. b, percentage of the total specific cAMP PDE activity inhibited by the dual PDE3 and PDE4 inhibitor zardaverine (10 μM) in intimal and medial layer-derived VSMC (note PDE3 and PDE4 compose >80% of the total cAMP PDE activity in VSMC). B, intimal layer (lane 3) and medial layer tissues (lane 2) from ballooned aorta, and tissue isolated from the medial layer of undamaged rat aorta (lane 1) were subjected to immunoblot analysis using anti-PDE3 or anti–β-actin antisera as described under Materials and Methods. A representative immunoblot of PDE3A (top) and β-actin (bottom) is shown and analysis of similar samples isolated from four rats subjected to this procedure yielded similar results. C, immunohistochemical analysis of rat aortic sections. Fixed sections of control undamaged rat aortae (A and C) or aortae subjected to balloon-induced damage (B, D, E, F) were stained with hematoxylin and eosin (A, B), incubated with a nuclear stain (DAPI; D), incubated with an anti–β-actin antiserum (E), or incubated with an anti-PDE3 antiserum (C and F). Images were viewed on Zeiss Axiovert S100 fluorescent microscope and captured by photography on colored film (A, B) or digitally with a SensiCam high-performance charge-coupled device camera at 20× magnification. Similar data were obtained with sections derived from three control and four ballooned aortae.
To more directly test our conclusion that PDE3A protein level is specifically reduced in intimal VSMC, an immunohistochemical approach was used (Fig. 2C). Overall, our data in these studies are consistent with an intimal VSMC-selective reduction in PDE3A protein level. Thus, although hematoxylin and eosin (Fig. 2C, panel B) or DAPI (Fig. 2C, panel D) staining of sections of ballooned aortae showed that similar numbers of VSMC were present in intimal and medial layers, a marked difference in anti-PDE3–dependent immunostaining was noted between these layers (Fig. 2C, panel F). Similar amounts of anti-PDE3 staining were seen when control undamaged and balloon-damaged aortic medial layers were compared (compare Fig. 2C, panels C and F). No selective PDE3B antisera were available for these experiments; therefore, levels of PDE3B in these sections could not be determined directly. Because β-actin staining was consistent with the idea that similar amounts of protein were expressed by VSMC in both the intimal and medial layers (Fig. 2C, panel E), we conclude that intimal VSMC express very low levels of PDE3A (a result consistent with data obtained by immunoblotting Fig. 2B).
Human Aortic Contractile and Synthetic VSMC Express Different Levels of PDE3A
To determine whether the decrease in the level of PDE3A that accompanied the phenotypic switch of rat aortic VSMC when cultured (in vitro) and in vivo after aortic ballooning also occurred when human VSMC were cultured, PDE3 and PDE4 activities were measured in lysates of human aorta (contractile phenotype) and in cultured human aortic VSMC (synthetic phenotype). As shown in Table1, synthetic human aortic VSMC express three cAMP PDE activities: PDE1, PDE3, and PDE4. Interestingly, as observed with rat aortic VSMC, synthetic human aortic VSMC expressed significantly less PDE3 activity than contractile human aortic VSMC (Table 1). Corroborating the reduced PDE3 activity observed in synthetic human aortic VSMC, immunoblot analysis of contractile and synthetic human aortic VSMC identified a marked difference in the level of PDE3A expression in these two human aortic VSMC phenotypes when corrected for β-actin content of the samples (Fig.3). Our finding of a cAMP PDE1 activity in synthetic human aortic VSMC, and the absence of this activity in contractile human aortic VSMC, was reported previously (Rybalkin et al., 1997; Palmer and Maurice, 2000). Consistent with our results with rat aortic VSMC (Fig. 1A), no differences in PDE4 activity (Table 1) or PDE4D3 level (data not shown) were observed when the two human aortic VSMC phenotypes were compared. We conclude therefore that a similar reduction in the level of PDE3A accompanies the phenotypic switch that occurs when human aortic VSMC are cultured. Although an immunohistochemical analysis similar to that carried out after balloon-induced vascular damage would directly test this conclusion, no vascular specimens with actively developing intimal layers were available to us. The need to investigate the significance of our finding with vascular specimens with actively developing intimal lesions was shown in earlier work, in which other researchers attempted to investigate expression of PDE1C in human intimal VSMC. In this earlier report, human intimal VSMC were shown to express an overall phenotype more similar to that of contractile VSMC at later stages of lesion progression (Rybalkin et al., 1997).
Different PDE activity in human aorta and cultured human aortic VSMC

Different levels of PDE3 activity and of PDE3A protein expression in human aorta and cultured human aortic VSMC. Segments of human thoracic aorta, or primary cultures of human aortic smooth muscle cells (HASMC) were lysed and processed for immunoblot analysis using anti-PDE3 and anti-β-actin antisera as described previously (Materials and Methods). A representative immunoblot is shown in which amounts of PDE3A and β-actin in an aliquot of human platelet lysate (lane 1, 25 μg), increasing amounts of human aortic lysate (lanes 2–4), or 5 μg of cultured human aortic VSMC (lane 5) are shown. Similar results were obtained with several cultures of human VSMC.
Pharmacological and Therapeutic Implications of our Findings.
cAMP versus cGMP Regulation of VSMC Function.
It is becoming more generally recognized that cultured VSMC often display a significant reduction in their capacity to use cGMP to bring about changes in cell function. Consistent with this, cultured arterial VSMC have been shown to express reduced levels of several cGMP-regulated enzymes compared with these cells before culture. These include the soluble guanylyl cyclase, the cGMP-dependent protein kinase, and PDE1B, a cGMP-hydrolyzing member of the PDE1 family of PDE (Boerth et al., 1997; Rybalkin and Bornfeldt, 1999; Anderson et al., 2000). In one report (Boerth et al., 1997), data consistent with the level of cGMP-dependent protein kinase being determinant in promoting the phenotypic switch of cultured aortic VSMC were presented. We suggest, based on the data presented in this report, that PDE3A, a cGMP-inhibited cAMP hydrolyzing PDE3, should be added to the list of cGMP-regulated enzymes whose expression is phenotypically regulated. Of course, significantly more work will be required to determine whether the reduced expression of PDE3A plays a determinant or secondary role in the phenotypic switch. Interestingly, the level of PDE3B, a cAMP-hydrolyzing PDE3 enzyme that is 10 times less sensitive to cGMP inhibition (Degerman et al., 1997), was unaltered during the phenotypic switch described in our studies. Again, the significance of this finding will require more study.
PDE3 versus PDE4 Regulation of VSMC Function.
Much of our previous work has shown that a strong positive correlation exists between levels of PDE3 and PDE4, activities in VSMC, and the impact that inhibition of these activities has on VSMC functions. Thus, PDE3 activity dominates in contractile VSMC and PDE3 inhibitors are potent inhibitors of blood vessel contraction and synergize with activators of adenylyl cyclases to relax blood vessels (Maurice and Haslam, 1990). In contrast, PDE4 activity is much less abundant in contractile VSMC, and PDE4 inhibitors are poor relaxants of blood vessels, unless used in combination with PDE3 inhibitors, or in blood vessels with intact endothelium (Komas et al., 1991), a condition under which one would predict that VSMC PDE3A could already be significantly inhibited by cGMP (Maurice et al., 1991). Similarly, consistent with the fact that PDE4 activity dominates in synthetic VSMC, PDE4 inhibitors are more effective inhibitors of VSMC proliferation or migration than PDE3 inhibitors (Souness et al., 1992; Palmer et al., 1998). Although it is tempting to speculate that the dominance of inhibition of PDE3 and PDE4 activities in these VSMC phenotypes simply reflects their relative catalytic abundance in these cells, evidence that these enzymes may regulate distinct pools of cAMP in cells is emerging (Chini et al., 1997). More detailed analysis of the effects that are altered downstream from these enzymes may be necessary to further clarify this important issue. Although altered PDE expression in different endothelial phenotypes has also been reported (Keravis et al., 2000), the impact of these findings on our data remains to be established.
PDE3A as a Therapeutic Target.
PDE3 has been a significant therapeutic target for the treatment of several cardiovascular disorders (Haslam et al., 1999; Movsesian, 1999; Osinski and Schror, 2000; Park et al., 2000; El Beyrouty and Spinler, 2001; Tanabe et al., 2001). Inhibitors of PDE3 activity may result in stimulated myocardial contractility, inhibition of platelet aggregation, and relaxation of vascular and airway smooth muscle. Indeed, milrinone and, more recently, cilostazol have been shown to markedly affect the functioning of cardiovascular tissues (Haslam et al., 1999; Movsesian, 1999;Osinski and Schror, 2000; Park et al., 2000; El Beyrouty and Spinler, 2001; Tanabe et al., 2001). Recently, cilostazol, a structural analog of the PDE3 inhibitor used in our studies (cilostamide), has been considered a promising candidate drug for prevention of restenosis because of its vascular and platelet effects (Ochiai et al., 1999;Tanabe et al., 2001). Indeed, several studies have indicated some degree of success in suppressing neointimal formation in animal models using cilostazol (Inoue et al., 2000; Aoki et al., 2001). Results of trials in which both aspirin and cilostazol were delivered in combination after stenting showed results comparable to those obtained when aspirin and ticlopidine, an inhibitor of platelet aggregation, were used (Park et al., 2000). Based on our present results, we suggest that cilostazol may have acted preferentially to inhibit platelet PDE3A when used in these trials. In fact, our data demonstrating a clear and marked reduction in the level of PDE3A in the proliferative and migratory synthetic VSMC provides strong support for the idea that the effects of this agent are most probably not caused by direct vascular effects.
Based on the results obtained here, a strategy of PDE1 or PDE4 inhibition could perhaps prove to be more effective than targeting PDE3 in reducing accumulation of intimal VSMC after vascular injury. Although no PDE1C-selective agents have yet been described, previous work indicating that Ro 20-1724, a PDE4 inhibitor, was more potent at inhibiting rat aortic VSMC migration than the PDE3 inhibitor cilostamide strengthens the idea of using PDE4 inhibitors (Palmer et al., 1998). In addition, because PDE4 inhibitors are not potent vasodilators in blood vessels without intact endothelial function (Komas et al., 1991), use of these agents might result in fewer undesirable cardiovascular effects such as vascular relaxation. Clearly, further work will be required to test these ideas.
Acknowledgments
Human aorta was generously provided by Dr. J. G. Pickering (John P. Robarts Research Institute, London, ON, Canada) and human aortic VSMC was provided by Dr. C. H. Graham (Queen's University, Kingston, ON, Canada).
Footnotes
- Received November 6, 2001.
- Accepted February 7, 2002.
-
H.A.D. is the recipient of an Ontario Graduate Scholarship, D.G.T. is a Heart and Stroke Foundation of Ontario (HSFO) Research Trainee, D.P. is the recipient of a Canadian Institutes of Health Research (CIHR) Doctoral Research Award, and D.H.M. is an HSFO Career Investigator. This work was made possible through a grant from the Heart and Stroke Foundation of Ontario (T4675).
Abbreviations
- VSMC
- vascular smooth muscle cells
- PDE
- phosphodiesterase
- HBSS
- Hanks' balanced salt solution
- Ro 20-1724
- 4-[(3-butoxy-4-methoxyphenyl)-methyl]-2-imidazolidinone
- IBMX
- 3-isobutyl-1-methylxanthine
- PCR
- polymerase chain reaction
- bp
- base pair(s)
- DAPI
- 4′,6′-diamidino-2-phenylindole dihydrochloride, hydrate
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}