


Abstract
Identification of a G protein-coupled receptor activated by UDP-glucose led us to develop a sensitive and specific assay for UDP-glucose mass and to test whether this sugar nucleotide is released as an extracellular signaling molecule. Mechanical stimulation of 1321N1 human astrocytoma cells by a change of medium resulted in an increase in extracellular levels of both ATP and UDP-glucose. Whereas ATP levels peaked within 10 min and subsequently returned to resting extracellular levels of 3 nM, UDP-glucose levels attained a steady state that exceeded that of resting ATP levels by 3- to 5-fold for at least 3 h. Similar rates of basal release of UDP-glucose and ATP (72 and 81 fmol/min/106 cells) combined with a rate of UDP-glucose metabolism approximately three times lower than ATP hydrolysis account for the elevated extracellular UDP-glucose levels on resting cells. A medium change also resulted in rapid appearance of UDP-glucose on the luminal surface of highly differentiated polarized human airway epithelial cells but at levels 2- to 3-fold lower than ATP. However, nucleotide sugar levels increased 3- to 5-fold over the ensuing 2 h, whereas ATP levels decayed to a resting level; consequently, resting extracellular UDP-glucose levels exceeded those of ATP by 5- to 10-fold. UDP-glucose also was observed at levels that equaled or exceeded those of ATP in the extracellular medium of Calu-3 airway epithelial, COS-7, CHO-K1, and C6 glioma cells. Consistent with the observation of significant extracellular UDP-glucose levels, expression of the UDP-glucose-activated P2Y14 receptor in COS-7 cells resulted in G protein-promoted inositol phosphate accumulation that was partially reversed by enzymatic removal of UDP-glucose from the medium. Taken together, these results indicate constitutive release of UDP-glucose from physiologically relevant tissues and suggest that UDP-glucose acts as an autocrine activator of the P2Y14receptor. Because cellular UDP-glucose is concentrated in the lumen of the endoplasmic reticulum, we speculate that UDP-glucose release may occur as a result of vesicle transport during trafficking of glycoproteins to the plasma membrane.
Adenine and uridine nucleotides are released as extracellular signaling molecules in most tissues effecting a broad range of physiologically important responses (Dubyak and El-Moatassim, 1993; Ralevic and Burnstock, 1998). Two large classes of receptors transduce these effects. The P2X receptors are ligand-gated ion channels primarily mediating fast responses to ATP and potentially other nucleotides in excitatory tissues (Khakh et al., 2000). The P2Y receptors are G protein-coupled receptors that exist both in the central and autonomic nervous systems as well as on most nonexcitatory cells (Harden et al., 1998). A complex array of ectoenzymes regulates the levels of extracellular nucleotides and terminates their action (Zimmermann 2000).
An orphan G protein-coupled receptor exhibiting up to 45% identity to P2Y receptor subtypes recently was shown to exhibit signaling responses to UDP-glucose but not to adenine or uridine nucleotides (Chambers et al., 2000; Freeman et al., 2001). The relatively widespread distribution of this G protein-coupled receptor [now referred to as the P2Y14 receptor (Abbracchio et al., 2003)] and its high selectivity for activation by UDP-glucose are consistent with the idea that UDP-glucose is released in a regulated fashion to act as an extracellular signaling molecule. To test this hypothesis, we developed a radiometric assay for UDP-glucose that allows reliable and specific detection at concentrations of the nucleotide sugar as low as 1 nM. This assay was applied to illustrate that UDP-glucose is released from 1321N1 human astrocytoma and other cells and accumulates on the cell surface at steady-state concentrations often in excess of those of extracellular ATP and UTP. Expression of the recombinant human UDP-glucose receptor in COS-7 cells resulted in enhanced G protein-promoted inositol phosphate accumulation that was partially inhibited by removal of medium UDP-glucose with nucleotide pyrophosphatase. The presence of extracellular UDP-glucose in the medium of several cell types, including highly differentiated airway epithelial cells, establishes the existence of this nucleotide sugar as an extracellular signaling molecule.
Materials and Methods
Cell Culture and Incubations.
1321N1 Human astrocytoma, C6 rat glioma, CHO-K1, NIH-3T3, and COS-7 cells typically were grown to confluence on 24-well plastic (Costar, Cambridge, MA) plates. Primary human bronchial epithelial cells produced from normal and cystic fibrosis specimens, and human Calu-3 serous airway epithelial cells were grown to confluence on 12-mm Transwell-Col supports under air-liquid conditions as described previously (Matsui et al., 2000; Huang et al., 2001). The cells were washed three times and incubated in minimum essential medium at 37°C in a humidified incubator supplemented with 5% CO2. At the times indicated, aliquots (200 μl) were removed, boiled for 2 min, and either used immediately or stored at −20°C for subsequent nucleotide analysis. For intracellular nucleotide measurements, cultures were lysed with 5% trichloroacetic acid, which was subsequently extracted with 12 volumes of ethylic ether.
Luciferin-Luciferase Assay for ATP.
We previously described this assay in detail (Lazarowski et al., 2000). Briefly, 30-μl samples were added to a test tube containing 270 μl of H2O, placed in the light-protected chamber of an Auto-Lumat LB953 luminometer, and 100 μl of 4× luciferin-luciferase reaction buffer was added using a built-in injector. Luminescence was subsequently recorded for 10 s and compared against an ATP standard curve generated in parallel. Luminescence was linear with a slope of unity between 0.1 and 1000 nM ATP.
Enzymatic Synthesis of Radiolabeled Pyrophosphate.
[32P]Pyrophosphate (PPi) was obtained by incubating 100 μCi of [γ-32P]ATP (100 nM) with 0.5 units/ml nucleotide pyrophosphatase from Crotalus adamanteus (Sigma, St. Louis, MO) at 30°C for 15 min, as described previously (Lazarowski et al., 2000). Freshly prepared [32P]PPi was frozen at −20°C and used within 5 days.
UDP-Glucose Measurements.
UDP-glucose was assayed under the conditions delineated experimentally as described underResults. Incubations were in a final volume of 150 μl containing known or unknown amounts of UDP-glucose, 25 mM HEPES, pH 7.4, and 0.5 U/ml UDP-glucose pyrophosphorylase from baker's yeast (Sigma), and 100 nM [32P]PPi (200,000 cpm) or the indicated concentrations. Incubations were terminated by addition of 0.3 mM PPi and immediate heating of samples for 2 min at 95°C.
HPLC Analysis.
Nucleotides and nucleotide-sugars were separated by HPLC (Shimadzu) via a 3.9 × 150 mm (60 Å, 0.5 μm) Nova-Pack C18 column (Waters) with an ion pairing mobile phase (1 ml/min) consisting of 8 mM tetrabutylammonium hydrogen sulfate, 60 mM KH2PO4, pH 5.3, and 15% methanol. Absorbance at λ = 260 nm and radioactivity were monitored on-line as described previously (Lazarowski et al., 2000).
Expression of the Recombinant UDP-Glucose P2Y14Receptor in COS-7 Cells.
cDNA comprising the coding sequence of the human P2Y14 receptor was inserted into the pcDNA3 vector with an hemagglutinin epitope tag at the amino terminus. The Gαqi5 chimera (Coward et al., 1999), generously provided by Dr Bruce Conklin (University of California at San Francisco), was in pcDNA1 vector. COS-7 cells (grown in 12-well plates) were transfected with the expression vectors for the P2Y14receptor or M2 muscarinic receptor and/or Gαqi5 using Fugene 6 transfection reagent (Roche Applied Science, Indianapolis, IN) at a ratio of 5 μl of Fugene to 1 μg of DNA.
Measurement of [3H]Inositol Phosphates.
Approximately 24 h after transfection, the medium was replaced with inositol-free DMEM containing [myo-3H]inositol (1 μCi/well). Sixteen hours after labeling, the cells were incubated with 10 mM LiCl in the absence or presence of test drugs. The reaction was stopped after 45 min by aspiration of the medium and the addition of 50 mM ice-cold formic acid. The accumulation of [3H]inositol phosphates was quantified by Dowex chromatography as described previously (Lazarowski et al., 1997).
Reagents.
[γ-32P]ATP (3000 Ci/mmol), UDP-d-[6-3H]glucose (27 Ci/mmol), [3H]glucose (22 Ci/mmol), and [14C]glucose-1-P (287 mCi/mmol) were obtained from Amersham Biosciences (Piscataway, NJ). [myo-3H]inositol (20 Ci/mmol) was from American Radiolabeled Chemicals (St. Louis, MO). Firefly luciferase and luciferin were from BD Biosciences PharMingen (San Diego, CA). Crotalus adamanteus nucleotide pyrophosphatase, UDP-galactose epimerase, grade III apyrase, and nucleoside diphosphokinase were from Sigma. Other chemicals were of the highest purity available and from sources previously reported (Lazarowski et al., 2000).
Results
The unambiguous identification of a G protein-coupled receptor that selectively recognizes UDP-glucose as an agonist (Chambers et al., 2000; Freeman et al., 2001) supports the notion that UDP-glucose acts as an extracellular signaling molecule. To test this hypothesis, we devised a sensitive radiometric assay for detection of UDP-glucose mass. We previously established an assay for UTP based on the strict substrate selectivity of UDP glucose pyrophosphorylase for this nucleotide (Lazarowski et al., 1997; Lazarowski and Harden, 1999). Given that this enzyme catalyzes the reversible interconversion of UDP-glucose and pyrophosphate to UTP and glucose-1P (i.e., PPi + UDP-glucose ↔ UTP + glucose-1P),1 we reasoned that an assay for UDP-glucose mass could be developed using [32P]PPi.
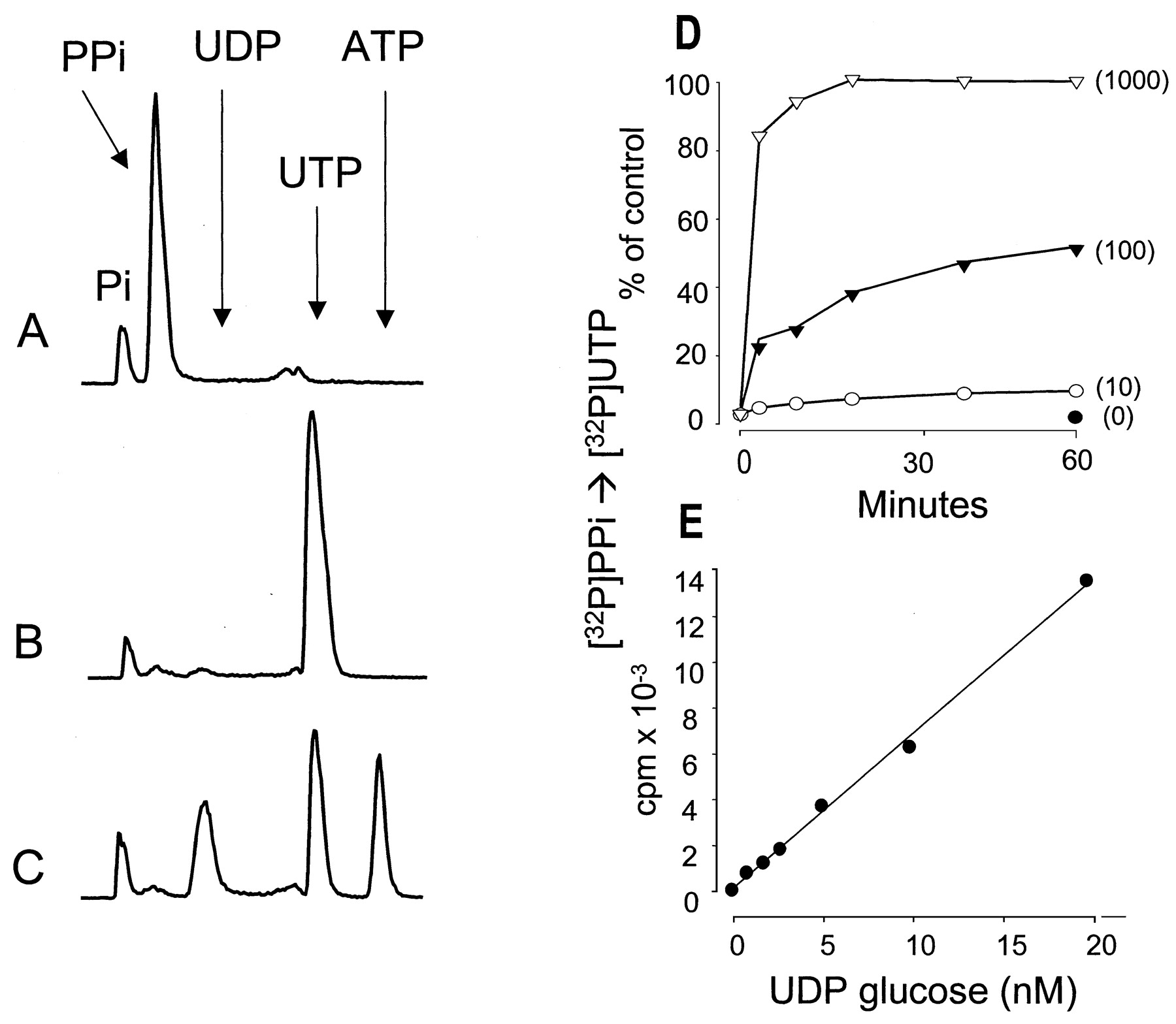
As illustrated in Fig. 1, A and B, incubation of 100 nM [32P]PPi with a molar excess of UDP-glucose (1 μM UDP-glucose) in the presence of UDPG-pyrophosphorylase resulted in formation of a radioactive species that comigrated with authentic UTP during HPLC analysis. The identity of the radioactive species as UTP was confirmed by illustrating that, in the presence of ADP, nucleoside diphosphokinase catalyzed the transfer of the terminal phosphate from this32P-containing molecule to ATP (Fig. 1C). Because only one phosphate in [32P]PPi is labeled and assuming that the radioactive species originally formed from UDP-glucose and [32P]PPi was [32P]UTP, 50% of the enzymatically transferred radioactive phosphate would be expected to exist in the β-phosphate position and 50% should exist in the γ-phosphate position. This relationship was observed as illustrated in Fig. 1C. Therefore, UDP-glucose-pyrophosphorylase effectively converts UDP-glucose to UTP under the conditions tested, and use of [32P]PPi as a cosubstrate provides a potential basis for establishing a radiometric assay for this nucleotide sugar.
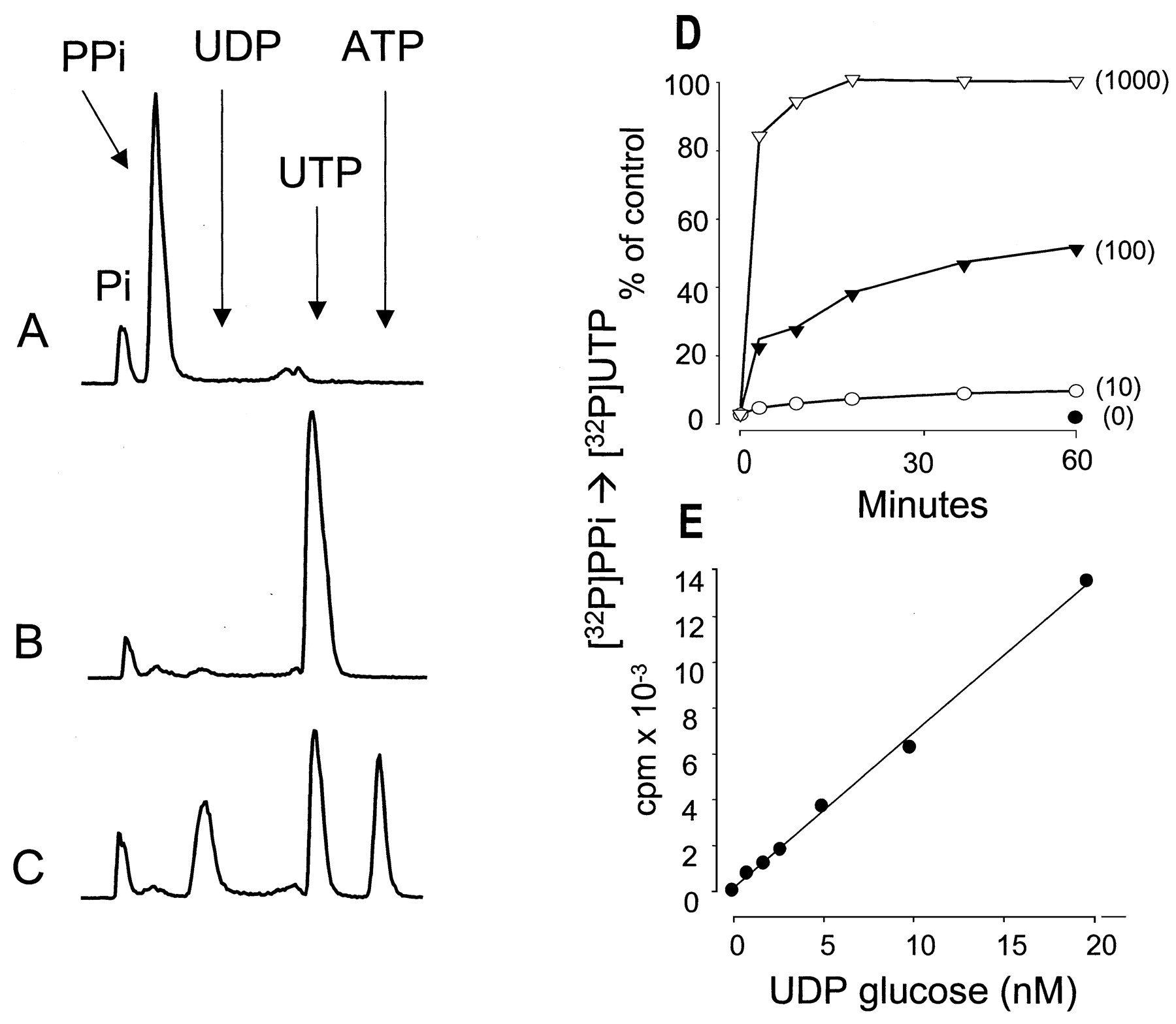
UDP-glucose dependent conversion of [32P]PPi to [32P]UTP. UDP-glucose pyrophosphorylase (0.5 U/ml) and 100 nM [32P]PPi were incubated for 60 min in the absence (A) or presence (B) of 1 μM UDP-glucose. UDP-glucose pyrophosphorylase was inactivated by heat, and the sample was subsequently incubated for 5 min in the presence of 0.2 U/ml nucleoside diphosphokinase and 1 μM ATP (C). D, time course for the UDP-glucose pyrophosphorylase-catalyzed conversion of [32P]PPi to [32P]UTP in the presence of 100 nM [32P]PPi. UDP-glucose concentrations are indicated in parentheses. E, a UDP-glucose calibration curve was obtained in the presence of 100 nM [32P]PPi. Other details as indicated under Materials and Methods.
The time and substrate concentration dependence for conversion of UDP-glucose and [32P]PPi to [32P]UTP were delineated to establish optimal conditions for reliable quantification of the mass of UDP-glucose. In reactions carried out with 100 nM PPi and 100 nM UDP-glucose, about 50% of the initial [32P]PPi was converted to [32P]UTP within 60 min (Fig. 1D). Although an increase of the specific activity of [32P]PPi obtained by lowering the PPi concentration potentially would result in a higher sensitivity for detection of UDP-glucose, conversion of [32P]PPi to [32P]UTP was inefficient at these lower substrate concentrations, with only 5% conversion observed with equimolar amounts (either 1 or 10 nM) of PPi and UDP-glucose (data not shown). Therefore, 100 nM PPi was used in subsequent assays. In the presence of 100 nM PPi, UDP-glucose was quantitatively converted to UTP irrespective of its concentration (Fig.1D). Concentrations of UDP-glucose as low as 1 nM were detected reliably under the conditions of this assay (i.e., 100 nM PPi), and the formation of radioactive product was linear up to 100 nM UDP-glucose concentration (Fig. 1E and data not shown).
An assay for mass of UDP-glucose in biological tissues necessarily requires absolute selectivity for detection of this nucleotide sugar over other molecules. As is illustrated in Fig.2, A and B, only UDP-glucose was a substrate under the conditions of this assay. UMP, UDP, UTP, UDP-galactose, UDP-glucuronic acid, and UDP-N-acetylglucosamine (100 nM each) all were inactive substrates for conversion of 32PPi to [32P]UTP in the presence of UDP-glucose pyrophosphorylase. Although UDP-galactose is not a substrate under the conditions of the assay for UDP-glucose, this assay potentially could be adapted to measure UDP-galactose by using UDP-galactose epimerase to enzymatically convert UDP-galactose to UDP-glucose. As is illustrated experimentally in Fig. 2C, addition of epimerase to UDP-galactose–containing samples resulted in formation of UDP-glucose. Because the presence of UTP in cell samples potentially could favor the backward reaction toward formation of PPi and UDP-glucose, we examined the effect of UTP at concentrations present in the extracellular medium of various cell types (Lazarowski and Harden 1999). Figure 2D indicates that 1 to 100 nM UTP did not substantively affect the UDP-glucose–dependent conversion of [32P]PPi to [32P]UTP. Thus, based on these and other data, we conclude that the assay we have developed is specific for quantitation of UDP-glucose mass in biological samples with a sensitivity of 1 nM (150 fmol per 150-μl sample).

Specificity of the UDP-glucose pyrophosphorylase reaction. The UDP-glucose pyrophosphorylase-catalyzed conversion of [32P]PPi to [32P]UTP was measured as detailed under Materials and Methods in the absence or in the presence of 100 nM of the indicated nucleotide or nucleotide sugar (A), in the presence of 100 nM UDP-glucose (B), or after a 15-min incubation of 100 nM UDP-galactose and 1 U/ml of UDP-galactose epimerase at 30°C (C). The HPLC tracings are representative of two experiments in duplicate with similar results. D, the formation of [32P]UTP in the presence of either 10 nM (○) or 100 nM UDP-glucose (●) was quantified in the presence of the indicated concentrations of UTP. The results are expressed as percentage of control formation of [32P]UTP obtained in the absence of UTP addition (3760 ± 230 and 27750 ± 780 cpm for 10 and 100 nM UDP-glucose, respectively). The values represent the mean ± mean difference from two experiments performed with duplicates. UDP-gal, UDP-galactose; UDP-gluAc, UDP-glucuronic acid; UDP-glcNAc, UDP-N-acetyl-glucosamine.
We reported previously that mechanical stimulation of 1321N1 human astrocytoma and other cells results in nonlytic release of ATP and UTP (Lazarowski et al., 1997; Lazarowski and Harden, 1999); we recently also observed constitutive release of ATP from 1321N1 cells and from airway epithelial cells under resting conditions (Lazarowski et al., 2000). The assay for UDP-glucose mass elucidated as described above was applied to determine whether 1321N1 cells also released this nucleotide sugar under resting and mechanically stimulated conditions. Gentle replacement of the 0.3 ml of medium bathing a well (113 mm2) of a 24-well dish of surface cultures of 1321N1 cells resulted in mild mechanical stimulation and therefore a marked release of ATP, which reached an extracellular concentration of approximately 100 nM within 10 min (Fig.3A). Extracellular ATP concentration subsequently decreased with t1/2 = 20.5 min to a resting steady-state level of 3 ± 1 nM in the bulk medium. In contrast, a relatively minor accumulation of extracellular UDP-glucose was observed during the first minutes after a medium change, which represented approximately 10% of ATP accumulation. However, UDP-glucose levels remained stable for 3 h at a higher concentration (10 nM) than the resting levels of ATP (Fig. 3B).

Time course of release of ATP and UDP-glucose by 1321N1 human astrocytoma cells. Cells (500,000/well) were rinsed and incubated in 0.3 ml of DMEM for the times indicated. ATP (A) and UDP-glucose (B) were quantified as indicated under Materials and Methods. The results are expressed as the mean value ± S.D. from three experiments performed with duplicate samples.
Intracellular levels of ATP and UDP-glucose were 10,970 ± 910 and 751 ± 5 pmol per million cells, respectively. The ratio of ATP to UDP-glucose in the extracellular medium during the initial 3 to 10 min after medium replacement also approximated the ratio of intracellular ATP to UDP-glucose, suggesting that a common mechanism of release occurred upon mechanical stimulation of 1321N1 cells. The fact that the steady state UDP-glucose levels on resting 1321N1 cells (i.e., 3 h after a change of medium), were much higher than that observed with ATP suggested that released UDP-glucose was either not metabolized or that its rate of metabolism is much lower than that of ATP. To test these possibilities, trace amounts of UDP-[3H]glucose were added to the medium of 1321N1 cells, and levels of the radioactive nucleotide sugar were followed over the ensuing 4 h. HPLC analysis of UDP-[3H]glucose metabolism indicated that the 3H-labeled product of UDP-[3H]glucose comigrates with authentic [14C]glucose-1-P standard, and no accumulation of [3H]glucose could be observed (Fig.4A). These results indicated the reaction UDP-[3H]glucose → [3H]glucose-1-P + UMP, which is consistent with the activity of an ectophosphodiesterase/nucleotide pyrophosphorylase (E-NPP), as we have recently observed on 1321N1 cells (Lazarowski et al., 2000).
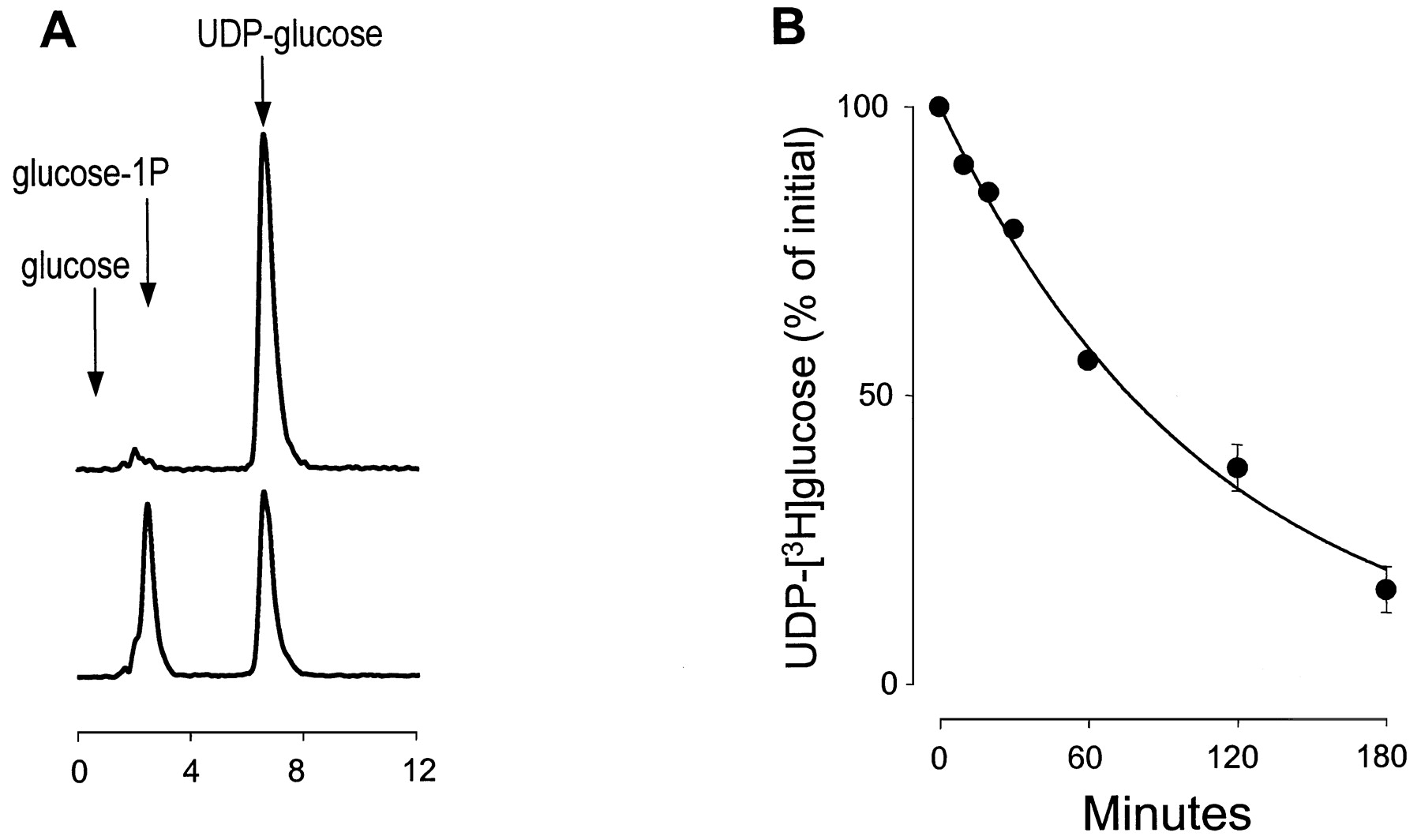
Hydrolysis of UDP-[3H]glucose on 1321N1 human astrocytoma cells. Cells (500,000/well) were incubated in 0.3 ml of DMEM containing 0.5 μCi of UDP-[3H]glucose. The medium was removed at time = 0 (A, top), 60 min (A, bottom), or as indicated (B), and the 3H species was analyzed by HPLC, as described under Materials and Methods. The retention time of commercial standards of [3H]glucose, [14C]glucose-1P, and UDP-[3H]glucose are indicated with arrows.
UDP-[3H]glucose levels in the bulk medium decreased with a t1/2 = 77 min. Thist1/2 value for UDP-glucose is approximately 3-fold higher than thet1/2 value of ATP observed under the same conditions (Fig. 3A) but does not fully explain why extracellular UDP-glucose remained essentially in the same concentration range for at least 3 h after medium replacement. The most likely basis for these results is that UDP-glucose hydrolysis is balanced by constitutive release of this sugar nucleotide. Using the first-order rate constant (k) for UDP-glucose hydrolysis calculated in Fig. 4B (k = 0.396/t1/2 = 0.009 min−1), the rate for constitutive release of the nucleotide sugar was 72 fmol/min/106 cells, and the basal rate of ATP release obtained in the same experiment was 81 fmol/min/106 cells. Taken together, these results indicate that ATP is released at a markedly increased rate relative to that of UDP-glucose for several minutes after mechanical stimulation. However, the rates of release of ATP and UDP-glucose decrease over time and reach comparable values on resting cells. Thereafter, the slower rate of UDP-glucose hydrolysis results in higher resting state levels of the nucleotide sugar in the extracellular medium.
We also determined whether extracellular UDP-glucose could be detected on other cells, including more physiologically relevant tissues such as primary airway epithelia from normal or cystic fibrosis donors. Similar to 1321N1 cells, a gentle change of medium on the luminal surface of polarized normal and CF bronchial epithelial cells resulted in transient accumulation of luminal ATP (5–10 nM ATP), which subsequently decayed to 1 to 2 nM on resting cells (Fig.5A). These results were consistent with previous observations with both mechanically stimulated and resting primary cultures of airway epithelial cells (Grygorczyk and Hanrahan 1997; Watt et al., 1998; Donaldson et al., 2000). UDP-glucose also was detected in the luminal medium at levels somewhat lower than ATP levels 5 min after a medium change. However, extracellular UDP-glucose levels increased by up to 3- to 5-fold over the ensuing 2 h on resting cells (Fig. 5). Moreover, UDP-glucose levels exceeded those of ATP by 5- to 10-fold 2 h after a change of medium on polarized epithelial cells.

Measurement of ATP and UDP-glucose accumulation in the luminal medium of primary bronchial epithelial cells. Polarized primary cultures of normal or CF cells were rinsed and incubated in 0.3 ml of DMEM. Luminal samples were removed at 5 min (░) or 120 min (■) after rinse, and ATP and UDP-glucose measurements were performed as detailed under Materials and Methods. The data represent the mean value (± S.D.) from three experiments performed with duplicate samples.
UDP-glucose levels also were detected in the extracellular medium of resting immortalized cells (Table 1). The highest levels were found in the medium of COS-7 and polarized Calu-3 cells and the lowest level with C6 glioma cells. The extracellular UDP-glucose in the medium of resting cells exceeded ATP levels by 4-fold in Calu-3 human airway epithelial cells, by 3-fold in 1321N1 cells, and by 2-fold in COS-7 cells. Similar extracellular levels of UDP-glucose and ATP were observed with CHO-K1 cells and C6 glioma cells.
Measurement of extracellular UDP-glucose and ATP on resting cells
An inverse correlation between resting levels of UDP-glucose and [3H]UDP-glucose metabolism was observed in experiments measuring the rates of hydrolysis of trace amounts of [3H]UDP-glucose on resting COS-7, Calu-3, and C6 cells (as illustrated with 1321N1 cells in Fig. 4). Whereas thet1/2 for [3H]UDP-glucose in the medium of C6 cells was 41 min, only 38% and 31% of the initial [3H]UDP-glucose was metabolized after 180 min by COS-7 and Calu-3 cells, respectively.
A potential extracellular signaling role for UDP-glucose was first suggested by studies of the P2Y14 receptor expressed as a recombinant protein in human embryonic kidney cells (Chambers et al., 2000; Freeman et al., 2001). UDP-glucose promoted guanine nucleotide exchange in P2Y14receptor-expressing human embryonic kidney cell membranes in a pertussis toxin-sensitive manner. UDP-glucose also promoted Ca2+ responses in cells cotransfected with the P2Y14 receptor and either Gα16, which promiscuously couples most G protein-coupled receptors to Gαq-activated responses, or a Gαqi chimera that specifically converts Gαi-coupled receptor signals into Gαq-dependent second-messenger responses. Using a Gαqi5 chimera in which Gαq contains the five carboxyl-terminal amino acids from Gαi (Coward et al., 1999), we investigated Gαqi5-dependent inositol phosphate production in COS-7 cells transfected with or without recombinant human P2Y14 receptor. Expression of the P2Y14 receptor alone had no effect on basal [3H]inositol phosphate levels relative to levels observed in wild-type, empty vector-transfected, or M2 muscarinic receptor-transfected COS-7 cells (Fig.6 and data not shown). Similarly, in the absence of Gαqi5, no UDP-glucose–stimulated inositol phosphate accumulation was observed in P2Y14receptor-expressing cells (Fig. 6A). In contrast, coexpression of the P2Y14 receptor with Gαqi5 resulted in a large increase in basal [3H]inositol phosphate levels that was further stimulated by the addition of UDP-glucose to the medium. Coexpression of the M2 muscarinic receptor with Gαqi5 did not result in an elevation of basal [3H]inositol phosphate accumulation, but carbachol stimulated [3H]inositol phosphate accumulation to levels similar to those stimulated by UDP-glucose in P2Y14 receptor/Gαqi5-expressing cells (Fig.6A).

Autocrine activation of the P2Y14receptor. A, COS-7 cells were transfected with either the M2 muscarinic receptor or the P2Y14 receptor, with or without the Gαqi5 chimera. The cells were labeled with [3H]inositol and subsequently incubated for 45 min in the presence of 10 mM LiCl in the absence of agonist or in the presence of 100 μM UDP-glucose or 300 μM carbachol. B, COS-7 cells cotransfected with Gαqi5 and either the empty pcDNA3 vector or the P2Y14 receptor were labeled with [3H]inositol and incubated for 45 min with LiCl in the absence or in the presence of 2 U/ml of the indicated nucleotidase. The resulting total [3H]inositol phosphates were quantified as detailed under Materials and Methods. The data represent the mean ± S.D. from three (A) or five (B) separate experiments, each performed with triplicate samples. *, the [3H]inositol phosphate accumulation in P2Y14receptor-expressing cells was significantly reduced (ttest) by NTPppase (p < 0.001) but not by apyrase (p > 0.7).
To investigate potential autocrine activation of the P2Y14 receptor, the effects of crotalus adamanteus NTPppase, which hydrolyzes nucleotide triphosphates and sugar nucleotides, and apyrase, which hydrolyzes nucleotides but not nucleotide sugars, were examined. NTPppase caused a partial but significant reduction of basal inositol phosphate accumulation in P2Y14 receptor expressing cells (Fig. 6B), whereas no effect of apyrase was observed. These results suggest activation of the recombinant P2Y14 receptor by endogenous UDP-glucose under basal culture conditions.
Discussion
By applying a sensitive new assay for mass of UDP-glucose, we have illustrated release of this nucleotide sugar from resting cells into the extracellular environment. Release of UDP-glucose occurred from all cells studied including highly differentiated polarized airway epithelial cells, and the extracellular concentrations of UDP-glucose were similar to those previously observed for the important extracellular signaling molecules ATP and UTP (Lazarowski and Harden, 1999). These results for the first time directly implicate UDP-glucose as an extracellular signaling molecule, and although not addressed in this study, our results indicate that similar methodologies can be applied to the evaluation of another potentially important extracellular molecule, UDP-galactose.
The physiological significance of the observation of regulated release of cellular UDP-glucose is heightened by the recent realization that an orphan G protein-coupled receptor [KIA0001; now referred to as the P2Y14 receptor (Abbracchio et al., 2003)] is potently activated by UDP-glucose and UDP-galactose (Chambers et al., 2000; Freeman et al., 2001). This receptor is expressed across a broad range of tissues including lung. The concentrations of extracellular UDP-glucose measured in the extracellular medium are sufficiently high to activate the UDP-glucose receptor, and given results from a variety of other studies with nucleotide-activated receptors (Lazarowski et al., 1997; Beigi et al., 1999), these bulk concentrations of nucleotide sugar probably significantly underestimate the concentration of UDP-glucose occurring at the cell surface. Indeed, we illustrate in the present study that expression of the recombinant UDP-glucose–activated P2Y14 receptor in COS-7 cells resulted in G protein activation. This response of the expressed UDP-glucose receptor was partially reversed by medium addition of nucleotide pyrophosphatase, which hydrolyzes UDP-glucose, but not by addition of apyrase, which does not hydrolyze this sugar nucleotide. Therefore, the UDP-glucose–activated P2Y14receptor, which is a structural member of the P2Y receptor class of receptors for extracellular nucleotides, can be considered a functional member of this class of signaling proteins given the regulated release of its cognate agonist into the extracellular space.
UDP-glucose plays a well established central role in intermediary metabolism. Earlier studies in glycobiology also had indirectly suggested that release of cellular nucleotide sugars might occur. For example, extracellular glycosyltransferase activity (Roth et al., 1971;Den et al., 1975) was detected and extracellular glycosylation was speculated to regulate intercellular adhesion. This idea has not been unambiguously supported by experimental results2 (reviewed in Roseman, 2001). The clear observation of release of nucleotide sugar reported here reopens the question of whether extracellular UDP-glucose or other nucleotide sugars may serve roles in addition to actions as activators of G protein-coupled receptors.
Detection of extracellular UDP-glucose was heretofore limited by lack of a sensitive assay. Previous assays were based on spectrometric quantitation of NADPH in a multienzymatic assay that used UDP-glucose pyrophosphorylase and a molar excess of PPi to generate glucose-1P plus UTP from UDP-glucose. Phosphoglucomutase was used to convert glucose-1-P to glucose 6-P, which was subsequently oxidized to gluconolactone-6-P by the enzyme glucose-6-P dehydrogenase in the presence of NADP (Hansen et al., 1966; Johnson et al., 1968). The sensitivity of this assay is in the micromolar range. Based on a similar principle, we reasoned that the enzyme UDP-glucose pyrophosphorylase should quantitatively convert [32P]PPi into [32P]UTP in a UDP-glucose dependent manner. Moreover, by manipulating the specific radioactivity of the substrate [32P]PPi, quantification of physiologically and pharmacologically relevant levels of endogenous UDP-glucose (i.e., low nanomolar concentrations) in extracellular fluids seemed feasible assuming that [32P]UTP can be quantified in the presence of excess of [32P]PPi. Thus, the assay described here combining the high selectivity of UDP-glucose pyrophosphorylase for UDP-glucose over other nucleotide-sugars and nucleotides with HPLC separation of [32P]UTP from [32P]PPi provides a method sufficiently sensitive and selective for accurate determination of extracellular UDP-glucose concentrations.
Nonlytic release of relatively large amounts of ATP and UTP occurs transiently during mechanical stimulation of nonexcitatory cells and tissues (Lazarowski et al., 1995, 1997, 2000; Grygorczyk and Hanrahan, 1997; Watt et al., 1998; Lazarowski and Harden, 1999; Ostrom et al., 2000; Schwiebert et al., 2002). Mechanically released ATP and UTP eventually decay to baseline concentrations in the low nanomolar range, which reflects a steady state at which rate of hydrolysis equals the rate of constitutive release (Donaldson et al., 2000; Lazarowski et al., 2000; Ostrom et al., 2000; Dubyak, 2002; Schwiebert et al., 2002). Thus, two apparently different mechanisms are involved in the release of nucleotides from nonexcitatory tissues. One mechanism operates in response to exogenously applied mechanical forces (e.g., shear, cell swelling, hydrostatic pressure) and may provide a rapid adaptation of cells to stress through acute activation of ATP- and/or UTP-sensitive P2Y receptors. The mechanism(s) involved in the release of ATP and UTP by mechanically stimulated cells clearly differs from those operating in excitatory tissues such as platelets and nerve terminals but remains poorly understood.
Like ATP (and UTP), UDP-glucose is released from 1321N1 cells during mechanical maneuvers such as a medium change but at a considerably slower rate than the nucleotides. The ratio of extracellular ATP:UDP-glucose observed during a medium change of 1321N1 cells correlates with the relative total cell content of ATP and UDP-glucose and suggests the occurrence of a common transport mechanism that reflects the relative intracellular concentrations of these species. However, purine and pyrimidine nucleotide and nucleotide sugar pools are heterologous (Moyer and Henderson, 1985) and releasable pools are probably compartmentalized. Furthermore, our data indicating persistent extracellular accumulation of UDP-glucose in the presence of active UDP-[3H]glucose hydrolysis reveal the existence of basal or constitutive release of the sugar nucleotide. The calculated rate of basal UDP-glucose release by 1321N1 cells is in the same magnitude as that of basal ATP release [72 and 81 fmol/min/106 cells, respectively (this study andLazarowski et al., 2000). The nearly identical rates of release of UDP-glucose and ATP coexist with a 10-fold difference in the intracellular levels of these two species suggesting that leakage of UDP-glucose and ATP through a nonselective membrane pore (e.g., during cell lysis) was not involved. This is an important observation, because available assays for cell lysis (e.g., LDH release and51Cr-labeling) have a sensitivity of approximately 1% (Lazarowski et al., 1995; Watt et al., 1998), whereas UDP-glucose accumulation on 1321N1 cells represents <0.1% of the UDP-glucose cell content. Moreover, these data are not consistent with the existence of a transport mechanism that simply exports both species according to their cytosolic abundance and suggests that different release pathways may exist for mechanical versus constitutive release and for ATP versus UDP-glucose.
We can only speculate about the mechanism of release and the subcellular source of extracellular UDP-glucose. Our data with 1321N1 cells indicate an UDP-glucose content of ∼0.35 fmol/cell, which would potentially provide a 1000-fold driving force for UDP-glucose export to the extracellular medium via a putative selective plasma membrane transporter. Alternatively, the UDP-glucose/UMP antiporter that concentrates UDP-glucose up to 20 times in the endoplasmic reticulum (ER) (Perez and Hirschberg, 1986; Abeijon et al., 1997; Castro et al., 1999) may indirectly provide a pathway for UDP-glucose release via vesicle transport. UDP-glucose participates in the quality control of protein synthesis in the lumen of the ER, and probably pre-Golgi structures, by tagging denatured domains of newly synthesized glycoproteins for subsequent chaperone recognition (Parodi, 2000; Zuber et al., 2001). Although our data do not rule out the potential release of cytosolic UDP-glucose via a transporter or an unidentified channel, we speculate that UDP-glucose transported to the ER lumen is released from cells as cargo during the constitutive pathway that targets glycoproteins to the plasma membrane. Similarly, vesicular trafficking of membrane-targeted phosphoproteins may provide a pathway for the release of vesicular ATP in resting cells. Exocytotic nucleotide release by nonexcitatory cells has been indirectly suggested by recent studies showing temperature-dependent release of ATP from endothelial cells (Schwiebert et al., 2002), and decreased extracellular ATP accumulation in oocytes (Maroto and Hamill, 2001) and endothelial cells (Knight et al., 2002) pretreated with pharmacological inhibitors of ER/Golgi vesicle trafficking.
Constitutive release of ATP, UTP, and now UDP-glucose may contribute to the tonic control of discrete cellular functions. For example, basal ATP release constitutes an important source of adenosine, which accumulates in pharmacologically relevant concentrations (> 50 nM) on resting airway surfaces and promotes tonic activation of the CFTR Cl− channel via activation of the A2b-adenosine receptor (Huang et al., 2001). UDP-glucose readily accumulates in the medium of 1321N1 human astrocytoma cells as well as in epithelial cells over several hours in the absence of external stimuli. As discussed above, endogenous UDP-glucose accumulates on cells in amounts that basally activate the UDP-glucose receptor heterologously expressed in COS-7 cells. Our results place UDP-glucose as a novel intercellular signaling molecule and suggest a potential mechanism for the constitutive release of nucleotides via vesicular trafficking. The physiological processes regulated by the widely distributed UDP-glucose receptor and its cognate agonist remain to be elucidated.
Acknowledgments
We are indebted to Catja van Heusden for excellent technical assistance.
Footnotes
-
↵1 An equilibrium constant of 0.67 for the reaction in the direction of UTP synthesis was reported (Hansen et al., 1966).
-
↵2 In contrast to most glycosyltransferases, only UDP-xylose:proteoglycan core protein xylosyltransferase has been confirmed to be secreted both by cultured cells and in vivo (Gotting et al., 1998; Kuhn et al., 2001).
-
This work was supported by United States Public Health Service grants HL34322 and GM38213.
- Abbreviations:
- UDP-glucose
- uridine 5′-diphosphoglucose
- HPLC
- high-performance liquid chromatography
- NTPppase
- nucleoside triphosphate pyrophosphatase
- PPi
- pyrophosphate
- DMEM
- Dulbecco's modified Eagle's medium
- ER
- endoplasmic reticulum
- Received October 28, 2002.
- Accepted February 3, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}