


Abstract
The potential role of dimerization in controlling the expression and pharmacological properties of α1-adrenoceptor subtypes was examined using coimmunoprecipitation of epitope-tagged receptors. Human α1-adrenoceptor subtypes (α1A, α1B, α1D) were tagged at their amino-termini with Flag or hemagglutinin epitopes and transfected into human embryonic kidney 293 cells. Homodimerization of all three subtypes was observed by coimmunoprecipitation of receptors with different tags and was not altered by norepinephrine treatment. Heterodimer formation between hemagglutinin-tagged α1B-adrenoceptors and Flag-tagged α1A- or α1D-adrenoceptors was also observed. However, no α1A/α1D-adrenoceptor heterodimers were observed, suggesting that dimerization is subtype-specific. The extent of heterodimerization was also unaltered by norepinephrine treatment. α1-Adrenoceptor truncation mutants lacking carboxyl or amino-terminal sequences formed homo- and heterodimers similarly to full-length receptors, suggesting that these domains play little or no role in dimerization. Biotinylation with a membrane-impermeable agent showed that monomers and homo- and hetero-oligomers of all three subtypes are expressed on the cell surface. Radioligand binding studies showed that heterodimerization did not alter the affinity of α1-adrenoceptors for norepinephrine, prazosin, or subtype-selective antagonists, suggesting that dimerization does not result in pharmacologically distinct subtypes. However, coexpression of α1B-adrenoceptors significantly increased both binding site density and protein expression of α1A- and α1D-adrenoceptors, and increased cell surface expression of α1D-adrenoceptors, suggesting a functional role for heterodimerization. Conversely, coexpression of α1A-with α1D-adrenoceptors, which did not heterodimerize, had no effect on receptor density or protein. These studies demonstrate subtype-selective heterodimerization of α1-adrenoceptors, which does not change their pharmacological properties but seems to have functional consequences in regulating receptor expression and trafficking.
Dimerization of growth factor and cytokine receptors is essential for their signaling (Heldin, 1995). However, G-protein coupled receptors (GPCRs) have traditionally been thought to function as monomers, with a single receptor binding ligand and activating G-proteins. Early studies, using muscarinic/adrenergic receptor chimeras, suggested that the monomeric GPCR model may not completely explain the mechanisms of GPCR function. These studies demonstrated that two binding-deficient muscarinic/adrenergic chimeras could form a receptor capable of specific ligand binding and signaling upon coexpression. Thus, it was suggested that a direct interaction between the two chimeras could reconstitute a ligand binding pocket (Maggio et al., 1993).
Strong evidence now suggests that some GPCRs, such as GABAB and taste receptors, exist as obligate heterodimers and that dimerization may alter the functional, pharmacological or regulatory properties of many other GPCRs (Bouvier, 2001; Rios et al., 2001). These observations have revolutionized contemporary views of GPCR function, raising previously unconsidered possibilities of multiple subunits, multiple binding sites, and additional subtypes that cannot be explained by existing GPCR clones expressed in isolation. GPCR dimerization may be mediated by covalent and/or noncovalent interactions, may involve extracellular, transmembrane, or intracellular domains, and, in some cases, may be altered by agonist occupancy (Bouvier, 2001; Rios et al., 2001). Strikingly, two closely related opioid receptors (κ and δ) heterodimerize to form receptors with distinct ligand binding profiles and functional properties (Jordan and Devi, 1999; Jordan et al., 2000). These studies suggest that dimerization of GPCRs may represent a novel mechanism modulating receptor pharmacology, trafficking and/or function.
α1-Adrenoceptors (α1-ARs) are Gq/11 coupled receptors that mediate responses to the neurotransmitters and hormones norepinephrine (NE) and epinephrine. α1-ARs initiate signals by activating phospholipase C and generating second messengers that release stored intracellular Ca2+ and stimulate protein kinase C. Three human α1-AR subtypes (α1A, α1B, and α1D) have been cloned, each encoded by different genes (Zhong and Minneman, 1999). These subtypes are highly homologous within their transmembrane domains but share little homology at their amino and carboxyl termini (Schwinn et al., 1995). Localization studies have shown that human tissues such as brain, heart, and vascular smooth muscle express mixtures of α1-ARs (Rokosh et al., 1996; Zhong and Minneman, 1999), and homogeneous cell populations, such as rat cardiomyocytes and clonal human SKN-MC neuroepithelioma cells, have also been found to coexpress all three α1-AR subtypes (Rokosh et al., 1996; Zhong and Minneman, 1999). Furthermore, there is pharmacological evidence for additional α1-AR subtypes that cannot be explained by the existing clones (Endoh, 1996; Ford et al., 1996, 1997; Rokosh et al., 1996; Zhong and Minneman, 1999). For example, α1L-AR, a subtype that has a relatively low affinity for prazosin, has been described in functional studies but has not been identified molecularly (Ford et al., 1997; Muramatsu et al., 1998). Other studies have also indicated discrepancies between native α1-ARs and recombinant subtypes (Johnson and Minneman, 1986; Han and Minneman, 1991). The possibility that native α1-AR subtypes may form dimers could partially explain such complexity.
Recent studies from our laboratory suggested that epitope-tagged human α1-AR subtypes can exist as both monomers and SDS-resistant homodimers and oligomers (Vicentic et al., 2002). In this study, we used biochemical and pharmacological approaches to examine the potential role of dimerization of human α1-AR subtypes in controlling their expression and pharmacological properties.
Materials and Methods
Materials. The human α1A-AR cDNA (Hirasawa et al., 1995) was kindly provided by Dr. Gozoh Tsujimoto (National Children's Hospital, Tokyo, Japan), human α1B-AR cDNA (Ramarao et al., 1992) was provided by Dr. Dianne Perez (Cleveland Clinic), and human α1D-AR was cloned in our laboratory (Esbenshade et al., 1995). Other materials were obtained from the following sources: HEK293 cells (American Type Culture Collection, Manassas, VA); fura-2/acetoxymethylester and n-dodecyl-β-d-maltoside (Calbiochem, La Jolla, CA); (-)-norepinephrine bitartrate, BMY7378, S-(+)-niguldipine, Dulbecco's modified Eagle's medium, penicillin, streptomycin, anti-Flag M2 affinity resin, and HRP-conjugated anti-Flag M2 antibody (Sigma Chemical Co., St. Louis, MO); anti-hemagglutinin (HA) affinity matrix (Roche, Indianapolis, IN); anti-HA tag polyclonal antibody and mammalian expression vector pCMV (BD Biosciences Clontech, Palo Alto, CA); sulfo-LC-NHS-biotin, Supersignal enzyme-linked immunosorbent assay Pico chemiluminescent substrate, and Immunopure immobilized streptavidin (Pierce, Rockford, IL); prazosin (Pfizer, Groton, CT); ECL reagent, [125I]arylazidoprazosin and carrier-free Na 125I (Amersham, Chicago, IL). G418 (geneticin), all electrophoresis reagents, and precast 4 to 20% Tris-Glycine polyacrylamide gels were obtained from Invitrogen (Carlsbad, CA), and Superfect Transfection Reagent was from QIAGEN (Valencia, CA).
Epitope-Tagged α1-AR Constructs. Human α1-AR cDNAs were generated by polymerase chain reaction and subcloned into the mammalian expression vector pDT containing in-frame N-terminal hexahistidine and Flag epitope tags as described previously (Robeva et al., 1996; Vicentic et al., 2002). These subtypes were also subcloned into the mammalian expression vector pCMV containing an in-frame N-terminal HA tag (YPYDVPDYA). After sequencing, unique restriction enzyme sites were used to replace the Flag-hexahistidine tag in pDT with the HA-epitope tag to allow generation of stable cell lines under the selection of 400 μg/ml G418.
Cell Culture and Transfection. HEK293 cells were maintained in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum, 10 mg/ml streptomycin, and 100 U/ml penicillin at 37°C in a humidified atmosphere with 5% CO2 (Esbenshade et al., 1993). Cells were transfected with the tagged human α1-AR subtypes by calcium phosphate precipitation (30 μg/15-cm plate). Transfected cells were propagated for several weeks in the presence of 400 μg/ml G418, and subclones were screened by radioligand binding for receptor expression. HA-tagged receptors were transiently transfected into a stable cell line expressing individual Flag-tagged receptors for receptor coexpression studies using 20 to 30 μg of cDNA, and cells were harvested 48 to 72 h after transfection.
Radioligand Binding and Ca2+Measurements. For radioligand binding, confluent 15-cm plates were washed with phosphate-buffered saline (PBS; 20 mM NaPO4, 154 mM NaCl, pH 7.6) and harvested by scraping. Cells were collected by centrifugation and homogenized with a Polytron homogenizer (Kinematica, Basel, Switzerland). Cell membranes were collected by centrifugation at 30,000g for 20 min and resuspended by homogenization in 1× buffer (25 mM HEPES, 150 mM NaCl, pH 7.4, 5 mM EDTA) with a protease inhibitor cocktail (1 mM benzamidine, 3 μM pepstatin, 3 μM phenylmethylsulfonyl fluoride, 3 μM aprotinin, and 3 μM leupeptin). Radioligand binding sites were measured by saturation analysis of specific binding of the α1-AR antagonist radioligand 125IBE 2254 (125IBE; 20-800 pM). Nonspecific binding was defined as binding in the presence of 10 μM phentolamine. The pharmacological specificity of radioligand binding sites was determined by displacement of 125IBE (50-70 pM) by selected agonists and antagonists, and data were analyzed by nonlinear regression (Theroux et al., 1996). Intracellular Ca2+ mobilization was measured using fura-2 as described previously (Theroux et al., 1996).
Photoaffinity Labeling. Photoaffinity labeling was performed on membranes from HEK293 cells expressing Flag- or HA-tagged α1-adrenoceptors. Membranes were prepared as described above for radioligand binding (1 μg protein/μl), and treated in the dark for 1 h at room temperature with 6 nM [125I]arylazidoprazosin (Vicentic et al., 2002). Nonspecific labeling was determined in the presence of 1 μM unlabeled prazosin. While still in the dark, open tubes were exposed to 6000 μJ/cm2 ultraviolet light for 3 min using a Stratalinker. Membranes were then washed once with 1 ml of 1× buffer A with protease inhibitors, centrifuged at 16,000g for 5 min and the pellet homogenized in 0.5 ml of 2× buffer A (50 mM HEPES, 300 mM NaCl, pH 7.4) containing protease inhibitors. Radiolabeled membranes were solubilized with 2% n-dodecyl-β-d-maltoside (DβM) in 1× buffer supplemented with protease inhibitor cocktail for 2 h at 4°C with gentle agitation. After solubilization, the soluble fractions (16,000g) and remaining insoluble material were counted on an automatic gamma counter (1470; PerkinElmer Wallac, Gaithersburg, MD),
Solubilization and Immunoprecipitation. Membrane preparations (2-3 mg protein) were prepared as described above and solubilized with 2% DβM in 1× buffer supplemented with protease inhibitor cocktail for 2 h at 4°C with gentle agitation. After solubilization, samples were centrifuged at 16,000g or 100,000g and the supernatant was diluted to 0.2% DβM in 1× buffer with protease inhibitors. Soluble receptors were incubated with M2 anti-Flag or anti-HA affinity matrix overnight at 4°C with gentle agitation. Resin was collected by centrifugation, washed with 1× buffer, deglycosylated on the resin with N-glycosidase F (PNGase F) with gentle agitation for 2 h at room temperature, and then eluted with 4× sample buffer (62.5 mM Tris-HCl, pH 6.8, 20% glycerol, 2% SDS, and 5% β-mercaptoethanol).
Western Blots. Immunoprecipitated samples were run on 4 to 20% Tris-glycine SDS-PAGE, transferred to nitrocellulose, and membranes blocked with 5% nonfat dried milk in Tris-buffered saline containing 0.1% Tween-20 (TBST) for 1 h at room temperature or overnight at 4°C with gentle agitation. Membranes were then incubated with the appropriate concentration of HRP-conjugated M2-anti-Flag antibody or anti-HA-polyclonal antibody for 1 h at room temperature. Membranes were washed with TBST and detected with ECL or incubated with the appropriate concentration of secondary IgG antibody and then detected with ECL.
Biotinylation of Cell Surface α1-ARs. Before membrane preparation, intact HEK293 cells expressing single Flag-α1-AR subtypes or coexpressing different Flag- and HA-tagged α1-ARs were rinsed with PBS and treated with 1.7 mg/ml Sulfo-LC-NHS-biotin, a membrane impermeant biotinylation agent, for 30 min at room temperature. The reaction was quenched by incubating cells with 500 mM Tris-HCl, pH 7.5, for 5 min at room temperature. Membranes were prepared and solubilized as described above and precipitated with immobilized streptavidin or anti-Flag affinity resin overnight at 4°C with gentle agitation. Resin was collected by centrifugation, washed with 1× buffer, receptors were deglycosylated with PNGase F on the resin with gentle agitation for 2 h at room temperature, and eluted with 4× sample buffer. Fractions (30 μl) were separated by SDS-PAGE, transferred to nitrocellulose, and blocked as described above. Membranes were incubated with HRP-conjugated M2-anti-Flag antibody or HRP-conjugated streptavidin, washed with TBST, and detected with ECL.
Surface Expression Assay. Stably transfected HEK293 cells were transiently transfected with tagged human α1-AR subtypes by Superfect transfection reagent (10 μg/10-cm plate). One day after transfection, cells were split into poly-d-lysine-coated 35-mm dishes (Biocoat) and grown overnight at 37°C. The cells were rinsed in PBS, fixed with 4% paraformaldehyde in PBS for 30 min, and then rinsed three times in PBS. The cells were blocked with blocking buffer (2% nonfat milk in PBS, pH 7.4) for 30 min and then incubated with the appropriate concentration of HRP-conjugated M2-anti-Flag antibody in blocking buffer for 1 h at room temperature. The cells were washed three times with blocking buffer, then once with PBS, and then incubated with enzyme-linked immunosorbent assay ECL reagent for 15 s. The luminescence, which corresponds to the amount of receptor on the cell surface (Xu et al., 2003), was determined by placing the plate inside a TD20/20 luminometer (Turner Designs, Sunnyvale, CA).
Results
Characterization of HA-Tagged α1-AR Subtypes. We made N-terminal HA-tagged human α1-AR subtypes for use with our previously characterized Flag-tagged subtypes (Vicentic et al., 2002) to directly examine homo- and heterodimerization. Constructs were expressed in HEK293 cells, which do not endogenously express detectable α1-ARs, and the density of recombinant receptor expression was determined by radioligand binding to be 0.3 to 16 pmol/mg of protein (Table 1). The three HA-α1-AR subtypes exhibited similar KD values for the radioligand 125IBE (Table 1), which were similar to KD values reported for 125IBE at both wild-type and Flag-tagged human α1-ARs (Esbenshade et al., 1993; Han and Minneman, 1991; Vicentic et al., 2002). The pharmacological specificities of the binding sites were also unaffected by the N-terminal tag, because KI values for the agonist NE, the nonsubtype selective antagonist prazosin, the α1A-selective antagonist (+)-niguldipine, and the α1D-selective antagonist BMY7378 in competing for specific 125IBE binding to HA-tagged subtypes showed no significant differences from those observed at wild-type and Flag-tagged α1-AR subtypes (Table 1) (Han and Minneman, 1991; Esbenshade et al., 1993; Lei et al., 2002). HA-tagged subtypes also showed NE-stimulated increases in intracellular Ca2+ (data not shown). Epitope-tagged receptor proteins were examined by Western blots after solubilization, immunoprecipitation, and deglycosylation (Fig. 1). An anti-HA antibody detected what seemed to be both monomers (α1A, ∼50 kDa; α1B, ∼65 kDa; α1D, ∼80 kDa) and dimers (∼100,130,160 kDa) and oligomers (∼150-240 kDa) of each receptor as observed previously with Flag-α1-AR subtypes (Vicentic et al., 2002).
Binding properties of HA-tagged α1-AR subtypes expressed in HEK293 cells
α1-AR subtypes with N-terminal HA tags were stably expressed in HEK293 cells, membranes were harvested, and saturation of specific 125IBE binding measured as described under Materials and Methods. Bmax and KD values were calculated by nonlinear regression of saturation curves, and pKI values for inhibition of binding by NE, prazosin, (+)-niguldipine, and BMY7378 were determined. Values are presented as mean ± S.E.M. of three experiments performed in duplicate.

Western blot analysis of HA-tagged-α1-ARs expressed in HEK293 cells. Membranes from cells stably expressing each HA-tagged subtype were solubilized and immunoprecipitated with anti-HA affinity resin. Samples were deglycosylated with PNGase F, subjected to SDS-PAGE, transferred to nitrocellulose, blotted with anti-HA-polyclonal antibody, and detected with ECL. Arrows indicate apparent monomers and dimers of each subtype, and the figure is representative of three or four experiments.
Homodimerization of α1-AR Subtypes. To obtain direct evidence for α1-AR homodimerization, each HA-tagged α1-AR subtype was transiently transfected into HEK293 cells (100-500 fmol/mg) stably expressing the same subtype containing a Flag tag (either Flag-α1A-, Flag-α1B-, or Flag-α1D-; 150-600 fmol/mg). Membranes from cotransfected cells were solubilized and immunoprecipitated with anti-Flag or anti-HA antibody. After elution and separation on gels, proteins were visualized by Western blots with anti-Flag antibody or anti-HA antibody. Figure 2 shows that the anti-Flag antibody apparently detected monomers (α1A,∼50 kDa; α1B, ∼65 kDa; α1D,∼80 kDa), dimers and oligomers (∼110-240 kDa) of all three subtypes in immunoprecipitates from either anti-Flag or anti-HA antibody (Fig. 2). Because no cross-reactivity between the two antibodies and tags was observed (data not shown), Flag-α1-ARs must have coprecipitated with HA-α1-ARs, supporting the existence of receptor homodimers and oligomers. Similar results were obtained when coexpressed receptors were detected with anti-HA antibody (data not shown).
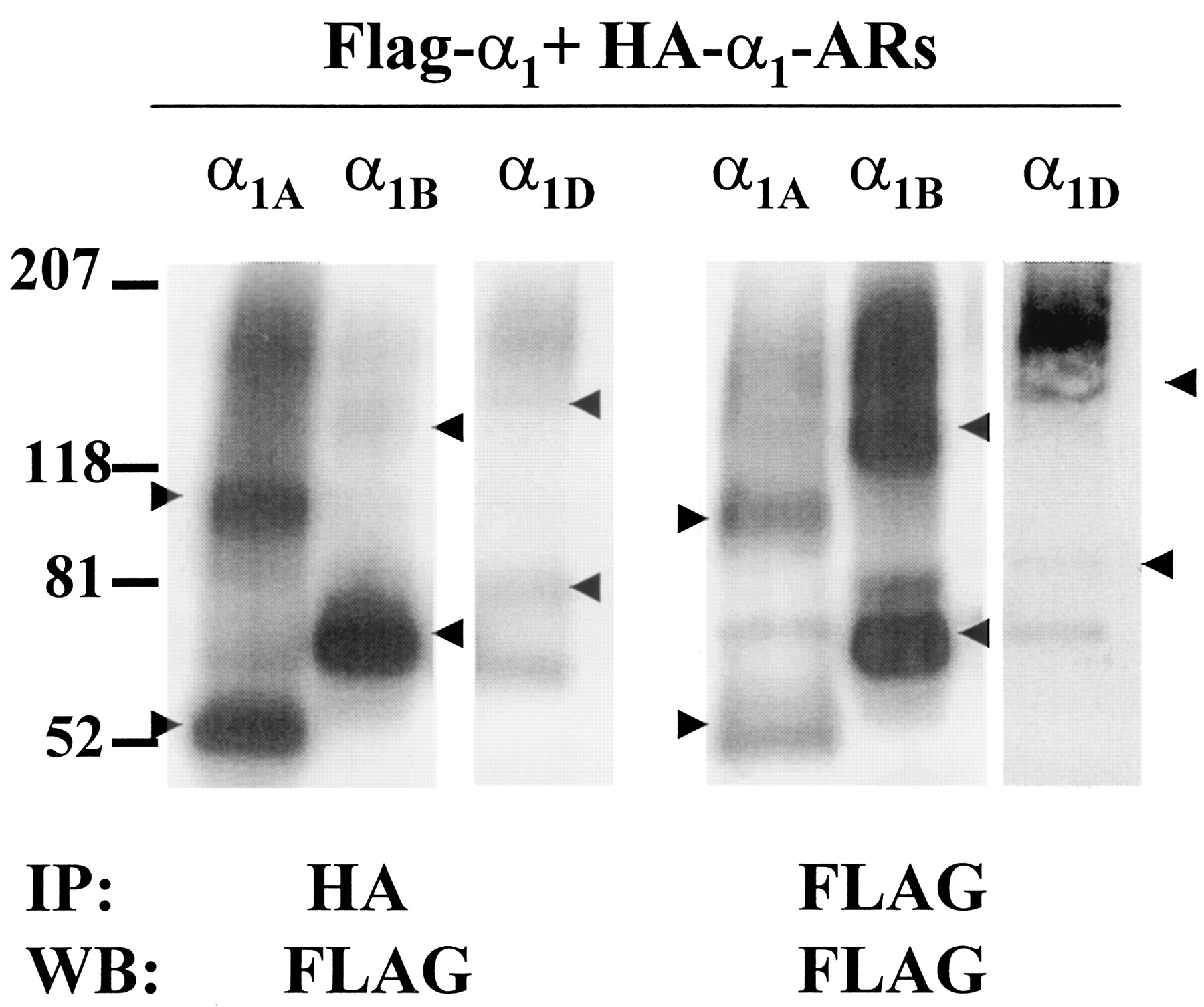
Coimmunoprecipitation of HA-α1-AR subtypes transiently coexpressed in HEK293 cells stably expressing the same Flag-α1-AR subtype. Cells coexpressing HA- and Flag-tagged α1A-; HA- and Flag-tagged α1B-; or HA- and Flag-α1D-ARs were harvested and membrane preparations solubilized and immunoprecipitated (IP) with anti-HA (left) or anti-Flag (right) affinity resin as indicated. Samples were deglycosylated, and Western blots (WB) performed with HRP-anti-Flag antibody. Arrows indicate apparent monomers and dimers of each subtype, and each blot is representative of three or four experiments.
Subtype-Specific Heterodimerization of α1-AR Subtypes. Differentially tagged combinations of α1-AR subtypes were coexpressed at similar levels in HEK293 cells, and coimmunoprecipitation was used to determine whether these subtypes form heterodimers. In Fig. 3, left, HA-α1B-ARs were transiently transfected into HEK293 cells (∼300-450 fmol/mg) stably expressing Flag-α1A- or Flag-α1D-ARs (150-600 fmol/mg). Expression levels of each subtype were within physiological levels (Schwinn, 1994; Graham et al., 1996; Michelotti et al., 2000). Membranes from these cotransfected cells were solubilized, immunoprecipitated with anti-Flag or anti-HA antibodies, and Western blots performed with anti-Flag or anti-HA antibodies. Figure 3 demonstrates an interaction between coexpressed Flag-α1A and HA-α1B-ARs. As expected after anti-Flag immunoprecipitation, the anti-Flag antibody detected bands migrating as monomers (52 kDa) and dimers (110 kDa) of Flag-α1A. After anti-HA immunoprecipitation, anti-Flag antibody also detected monomers and dimers of Flag-α1A, presumably caused by association with HA-α1B (Fig. 3, Left). After anti-HA immunoprecipitation, the HA-α1B seemed as both monomers and dimers of a higher molecular mass, as expected.

Immunoprecipitation of HA-tagged α1-AR subtypes transiently coexpressed with stable Flag-tagged α1-AR subtypes. Left, Flag-α1A-and HA-α1B-ARs coexpressed in HEK293 cells. Center, coexpressed Flag-α1D-ARs and HA-α1B-ARs. Right, coexpressed Flag-α1D-ARs and HA-α1A-ARs. Membrane preparations from cells expressing different combinations of subtypes were solubilized and immunoprecipitated (IP) with anti-Flag or anti-HA affinity resin as indicated. Samples were deglycosylated, electrophoresed and transferred, and Western blots (WB) were performed with anti-Flag or anti-HA antibodies. Arrows indicate apparent monomers and dimers of each receptor. This blot is representative of three or four experiments.
Figure 3, center, also shows interactions between coexpressed Flag-α1D and HA-α1B-ARs. After coexpression, solubilization, and immunoprecipitation with anti-Flag antibody, monomers (80 kDa), dimers (160 kDa), and trimers (240 kDa) of Flag-α1D-ARs were apparent as observed previously. After anti-HA immunoprecipitation, anti-Flag antibody also detected similar Flag-α1D-ARs, indicating interactions between Flag-α1D and HA-α1B-ARs (Fig. 3, center). Dimers and oligomers were also observed after anti-HA precipitation. As expected, after immunoprecipitation with anti-HA antibody the HA-α1B seemed as both monomers and dimers on an anti-HA blot, which were smaller than Flag-α1D-ARs.
Because α1B-ARs formed heterodimers with both α1A- and α1D-ARs, we wanted to determine whether α1A/α1D-AR heterodimers could also be detected. HA-α1A-ARs were transiently expressed (∼200-300 fmol/mg) into stably expressing Flag-α1D-AR HEK293 cells (∼150-300 fmol/mg), solubilized, and immunoprecipitated with anti-HA or anti-Flag antibodies. Figure 3 (right) shows that after immunoprecipitation with anti-HA antibody, Flag-α1D-ARs were not detected by anti-Flag antibody, even after long exposure times. However, parallel blots of anti-Flag immunoprecipitated Flag-α1D-ARs and anti-HA immunoprecipitated HA-α1A-ARs detected bands consistent with monomers and oligomers of each receptor, respectively (Fig. 3, right), confirming that these receptors were efficiently expressed at similar levels. Taken together, these results suggest that interactions between α1-ARs are restricted to particular combinations of subtypes.
To determine whether these observations were caused by interactions of these highly hydrophobic proteins occurring after solubilization, different HEK293 cell lines, each expressing only one (Flag-α1A or HA-α1B) tagged subtype, were solubilized, mixed together, and sonicated again to allow the two receptor populations to come together in the same micelle. When these mixtures were immunoprecipitated with anti-HA antibody, anti-Flag antibody did not detect Flag-α1A-AR on Western blots (Fig. 4A, left); and when the mixture was immunoprecipitated with anti-Flag antibody, anti-HA antibody did not detect HA-α1B-ARs (Fig. 4A, right). Similar results were seen with Flag-α1D- and HA-α1B-AR mixtures (not shown), demonstrating that these interactions are not solubilization artifacts, and that receptors must be coexpressed in cells to interact.

A, lack of interaction between presolubilized α1-AR subtypes. Membrane preparations from HEK293 cells expressing only Flag-α1A or only HA-α1B-ARs were solubilized, mixed together, re-sonicated, and immunoprecipitated (IP) with anti-Flag or anti-HA affinity resin as indicated. Samples were deglycosylated with PNGase F, and Western blots (WB) were performed with anti-Flag antibody (left) or anti-HA antibody (right). B, membrane preparations from HEK293 cells coexpressing Flag-α1A and HA-α1B-ARs were solubilized, centrifuged at 16,000g (16) or 100,000g (100) and immunoprecipitated (IP) with anti-Flag or anti-HA affinity resin as indicated. Samples were deglycosylated, and Western blots (WB) were performed with anti-Flag or anti-HA antibody as indicated. Arrows indicate apparent monomers and dimers of each receptor. Results are representative of three or four experiments.
Extent of Solubilization of Epitope-Tagged α1-ARs. Membranes from HEK293 cells stably expressing epitope-tagged α1-ARs were solubilized with 2% DβM, which has previously been used to effectively solubilize G-protein coupled receptors (Gether et al., 1995). We used photoaffinity labeling to determine the efficiency of DβM solubilization of α1-ARs. Receptors were covalently labeled with [125I]arylazidoprazosin before solubilization, and the proportion of labeled receptor solubilization was determined. Membranes expressing α1A-ARs, α1B-ARs, or a combination of α1A- and α1B-ARs were labeled with [125I]arylazidoprazosin, and total radioactivity incorporated was determined before and after solubilization and centrifugation. We found that 83, 86, and 88% of α1A-, α1B-, and α1A+α1B-ARs, respectively, were found in the soluble fraction, showing that a large majority of photoaffinity-labeled receptors are solubilized with this protocol. Because of the low degree of photoaffinity labeling of α1D-ARs (Vicentic et al., 2002), the degree of solubilization of this subtype was not determined.
Ultracentrifugation was also used to determine whether these receptors might be only partially solubilized. Solubilized samples were ultracentrifuged at 100,000g and compared by Western blot analysis to soluble fractions spun at our standard 16,000g. Figure 4B shows that there are no differences between solubilized Flag-α1A- and HA-α1B-ARs after 100,000g or 16,000g spins when immunoprecipitated with anti-Flag or anti-HA resin and blotted with anti-Flag or anti-HA antibodies. Similar results were seen with the other α1-AR combinations (not shown), demonstrating that these interactions are not caused by partial receptor solubilization.
Effect of Agonist Activation on Dimerization. To determine whether agonist occupancy altered homodimer formation, HEK293 cells expressing single Flag-α1-AR subtypes were pretreated with 100 μM NE for 15 min at 37°C. Western blots of immunoprecipitated samples from both stimulated and nonstimulated cells were quantitated by densitometry and relative band intensities determined. These experiments showed that NE did not significantly alter either the density of the bands on Western blots (not shown) or the ratios of monomers to higher order oligomers (Fig. 5A).

Lack of effect of NE on dimerization of α1-AR subtypes. Intact cells expressing one or more tagged subtypes were serum starved for 2 h and treated with or without 100 μM NE for 15 min. Membranes were harvested, solubilized, immunoprecipitated, and blotted with anti-Flag antibody. Western blots (not shown) were analyzed by densitometry for monomers and dimers as determined by their apparent molecular masses. A, percentage relative band intensity comparing distribution of stimulated and unstimulated Flag-α1A-, Flag-α1B-, or Flag-α1D-ARs expressed in isolation. B and C, Flag-α1A-ARs or Flag-α1D-ARs were cotransfected with HA-α1B-ARs, immunoprecipitated with anti-HA affinity resin, and blotted with anti-Flag antibody. The percentage of relative band intensities of the Flag-tagged subtypes in the presence or absence of NE were quantitated. The density of each band is expressed as a percentage of the density of the unstimulated monomer band (set as 100%). Data represents mean ± S.E.M. of three independent experiments.
Similar experiments were performed to determine whether agonist occupancy altered heterodimer formation. HEK293 cells coexpressing HA-α1B- (∼300-450 fmol/mg) and Flag-α1A- or Flag-α1D-ARs (150-600 fmol/mg) were pretreated with 100 μM NE for 15 min at 37°C before lysis. Solubilized fractions were immunoprecipitated with anti-HA affinity resin, and Western blots were performed with anti-Flag antibody. Blots were quantitated by densitometry, showing that NE did not alter either expression (not shown), or the extent of heterodimer formation for the Flag-tagged subtype in either combination (Fig. 5, B and C). NE also did not induce heterodimer formation between α1A- and α1D-ARs (data not shown). These results indicate that dimerization of α1-AR subtypes is independent of agonist activation.
Effect of C-Terminal Truncation on Dimerization. To examine a possible role for the intracellular C terminus in α1-AR dimerization, we constructed C-terminal truncation mutants of each Flag-α1-AR subtype. Truncations were made 14 to 16 amino acids (aa) after the end of the predicted 7th transmembrane domain, at a conserved glutamine (α1A, Gln344; α1B, Gln366) or adjacent arginine (α1D, Arg418) by mutation into a stop codon, and C-truncated (Ctr) Flag-α1-ARs stably expressed in HEK293 cells. In Fig. 6A, it is apparent that after solubilization and immunoprecipitation with anti-Flag antibody, Western blots with anti-Flag antibody detected bands migrating as monomers and dimers of the Flag-Ctrα1A-, α1B-, and α1D-ARs. As expected, C-truncated mutants were about 30% smaller than full-length receptors, with the corresponding monomers running around 36, 40, and 45 kDa and dimers around 75 to 100 kDa. Full-length HA-α1B-ARs (∼300-450 fmol/mg) were then transfected into HEK293 cells stably expressing Flag-Ctr-α1A, Flag-Ctr-α1B, or Flag-Ctr-α1D-ARs at physiological levels (100-400 fmol/mg). After solubilization and immunoprecipitation with anti-HA affinity resin and deglycosylation, Western blots were performed with anti-Flag antibody. Figure 6B shows that anti-Flag antibody detected monomers and dimers of all three C-truncated mutants after immunoprecipitation with anti-HA antibody, indicating interactions between full-length HA-α1B-ARs and each Flag-Ctr-α1-AR subtype. The size of the anti-HA coprecipitated Ctr-α1-ARs were similar to those observed after anti-Flag immunoprecipitation, although the ratio of monomers to oligomers was higher in the anti-HA versus anti-Flag immunoprecipitated samples. These results suggest that full-length α1B-ARs form dimers with α1A-, α1B, or α1D-AR C-truncated mutants, supporting the hypothesis that the C-terminal domain is not critical for this interaction.

Western blot analysis of Flag-α1-AR C-terminal (Ctr) or N-terminal (Ntr) truncation mutants. Flag-tagged Ctr-α1A, Ctr-α1B, or Ctr-α1D-ARs (CtrA, CtrB, CtrD) were expressed alone (A) or coexpressed (B) with full-length HA-α1B-ARs (HA-B) in HEK293 cells. C, Flag-tagged N-truncated α1D-ARs (NtrD) were coexpressed with full-length HA-α1B-ARs (HA-B) in HEK293 cells. Membrane preparations were solubilized and immunoprecipitated (IP) with anti-Flag or anti-HA affinity resin as indicated. Samples were deglycosylated and Western blots (WB) performed with HRP-anti-Flag or anti-HA antibody. Arrows in B and C indicate apparent monomers and dimers of each subtype. Results are representative of three or four experiments.
Effect of N-Terminal Truncation on α1D-AR Dimerization. α1D-ARs have an unusually long extracellular N terminus (95 aas) compared with α1A- and α1B-ARs (25 and 45 aa). We examined the possibility that the N terminus of α1D-ARs might be involved in heterodimerization by generating mutants truncated at Gly79 (Pupo et al., 2003). N-Truncated-α1D coding sequence was subcloned into the expression vector pDT containing an N-terminal Flag tag, and Flag-Ntr-α1D-ARs were transiently coexpressed with full-length HA-α1B-ARs in HEK293 cells. After immunoprecipitation with anti-Flag antibody, Western blots were performed with anti-Flag or anti-HA antibodies. Figure 6C shows that anti-Flag antibody detected monomers and dimers of Flag-Ntr-α1D-ARs with approximate molecular masses of 55 and 110 kDa, as previously reported (Pupo et al., 2003). Figure 6C also shows that anti-HA antibody detected monomers and dimers of full-length HA-α1B-ARs immunoprecipitated by the anti-Flag antibody, indicating an interaction between the full-length HA-α1B and Flag-Ntr-α1D-ARs, suggesting that the N terminus is not required for heterodimerization.
Biotinylation of Monomers and Higher Order Oligomers. To determine whether α1-AR homodimers were present on the cell surface, intact HEK293 cells expressing single Flag-α1-AR subtypes were treated with the membrane-impermeant biotinylating reagent sulfo-LC-NHS-biotin before membrane preparation. After solubilization, biotinylated proteins were precipitated with streptavidin, deglycosylated, and eluted with 4× sample buffer. Western blots of streptavidin-purified samples were performed with anti-Flag antibody to determine which receptor species were subject to cell-surface biotinylation. Monomers of all three receptors corresponding to their predicted molecular masses (α1A,∼50 kDa; α1B, ∼65 kDa; α1D,∼80 kDa) could be detected in the surface biotinylated samples (Fig. 7, top left), although the α1A signal was weak and required a long exposure time for detection. In addition, homodimers and trimers of all three subtypes, corresponding to their predicted molecular masses, were detected in the streptavidin-purified samples. To compare with total receptor expression, parallel samples were immunoprecipitated with anti-Flag affinity resin, and blotted with anti-Flag antibody. All three anti-Flag immunoprecipitated receptors were also visible as monomers, dimers and trimers (Fig. 7, top right). Sequential coimmunoprecipitations were also done to further determine which receptor complexes were present on the cell surface. In these experiments, biotinylated α1-AR samples were immunoprecipitated with anti-Flag and then anti-HA resin before detection with streptavidin on Western blots. However, no bands were detected on these blots (data not shown) even after long exposure times, probably because of poor recovery of samples after this complex procedure.

Surface expression of Flag-α1-AR subtypes detected with a membrane-impermeant biotinylating reagent. Intact HEK293 cells expressing individual Flag-α1-AR subtypes (top) or a combination of subtypes as indicated (bottom) were treated with sulfo-LC-NHS-biotin and harvested, and membranes were prepared. Membranes were solubilized and precipitated (IP) with either immobilized streptavidin or anti-Flag affinity resin, as indicated. Samples were deglycosylated and eluted with 4× sample buffer. Equal amounts of protein (in 30 μl) were loaded onto Western blots (WB) detected with HRP-anti-Flag antibody (top) or HRP-streptavidin (bottom). Closed arrows indicate apparent monomers and dimers of each receptor (top) or apparent heterooligomers of Flag-α1A- + HA-α1B-ARs or Flag-α1D- + HA-α1B-ARs (bottom); hollow arrow indicates apparent HA-α1B-AR dimers (bottom) that were observed in both coexpression samples. These blots are representative of two to six experiments.
To determine whether heterodimers were present on the cell surface, cells cotransfected with Flag-α1A- and HA-α1B-ARs, or cotransfected with Flag-α1D- and HA-α1B-ARs, were also treated with sulfo-LC-NHS-biotin. After solubilization, biotinylated proteins were immunoprecipitated with anti-Flag affinity resin, deglycosylated, eluted with sample buffer, and blotted with HRP-streptavidin. In Fig. 7, bottom, high molecular mass species apparently corresponding to trimeric complexes of α1A- and α1B-ARs (∼150-160 kDa) were detected by streptavidin on the cell surface, along with apparent HA-α1B-AR dimers (∼125 kDa). After a long exposure, dimers of Flag-α1A-ARs were also detected on the cell surface (data not shown). Figure 7, bottom, also shows large molecular mass species consistent with heterodimers and -trimers of α1B- and α1D-ARs (∼145-200 kDa) detected by streptavidin, along with apparent HA-α1B-AR homodimers. These data indicate that oligomeric complexes of α1B/α1A-ARs and α1B/α1D-ARs are expressed on the cell surface, although it is difficult to determine, in these higher order complexes, which receptors are actually present and in what proportion.
Pharmacological Characterization of α1-AR Heterodimers. The binding properties of heterodimers were determined using the antagonist radioligand 125IBE. The density of binding sites (Bmax) and affinity (KD) for 125IBE were determined by saturation analysis of specific 125IBE binding in membranes from cells transfected with different combinations of tagged subtypes expressed alone or together (Tables 2 and 3), with each experiment representing three to four different transfections. No significant differences in affinity for 125IBE were found with any combinations of subtypes examined (Tables 2 and 3).
125IBE binding to Flag-α 1A and HA-α 1B-ARs expressed alone and together
Differentially tagged α 1-AR subtypes were expressed alone or HA-α 1B-ARs were transiently expressed in HEK293 cells stably expressing Flag-α 1A-ARs, membranes were harvested, and Bmax and KD values were calculated by nonlinear regression of saturation curves of specific 125IBE binding. Estimated Bmax values for each coexpressed subtype were calculated from the fraction of high- and low-affinity binding sites for (+)-niguldipine (see Fig. 8). Values are presented as mean ± S.E.M. of three independent transfection experiments performed in duplicate.
125IBE binding to HA-α 1A, HA-α 1B-, and Flag-α 1D-ARs expressed alone and together
Differentially tagged α 1-AR subtypes were expressed alone or HA-α 1B- or HA-α 1A-ARs were transiently expressed in HEK293 cells stably expressing Flag-α 1D-ARs, membranes were harvested, and Bmax and KD values calculated by nonlinear regression of saturation curves of specific 125IBE binding. Estimated Bmax values for each coexpressed subtype were calculated from the fraction of high- and low-affinity binding sites for BMY7378 (see Fig. 8). Values are presented as mean ± S.E.M. of three independent transfection experiments performed in duplicate.
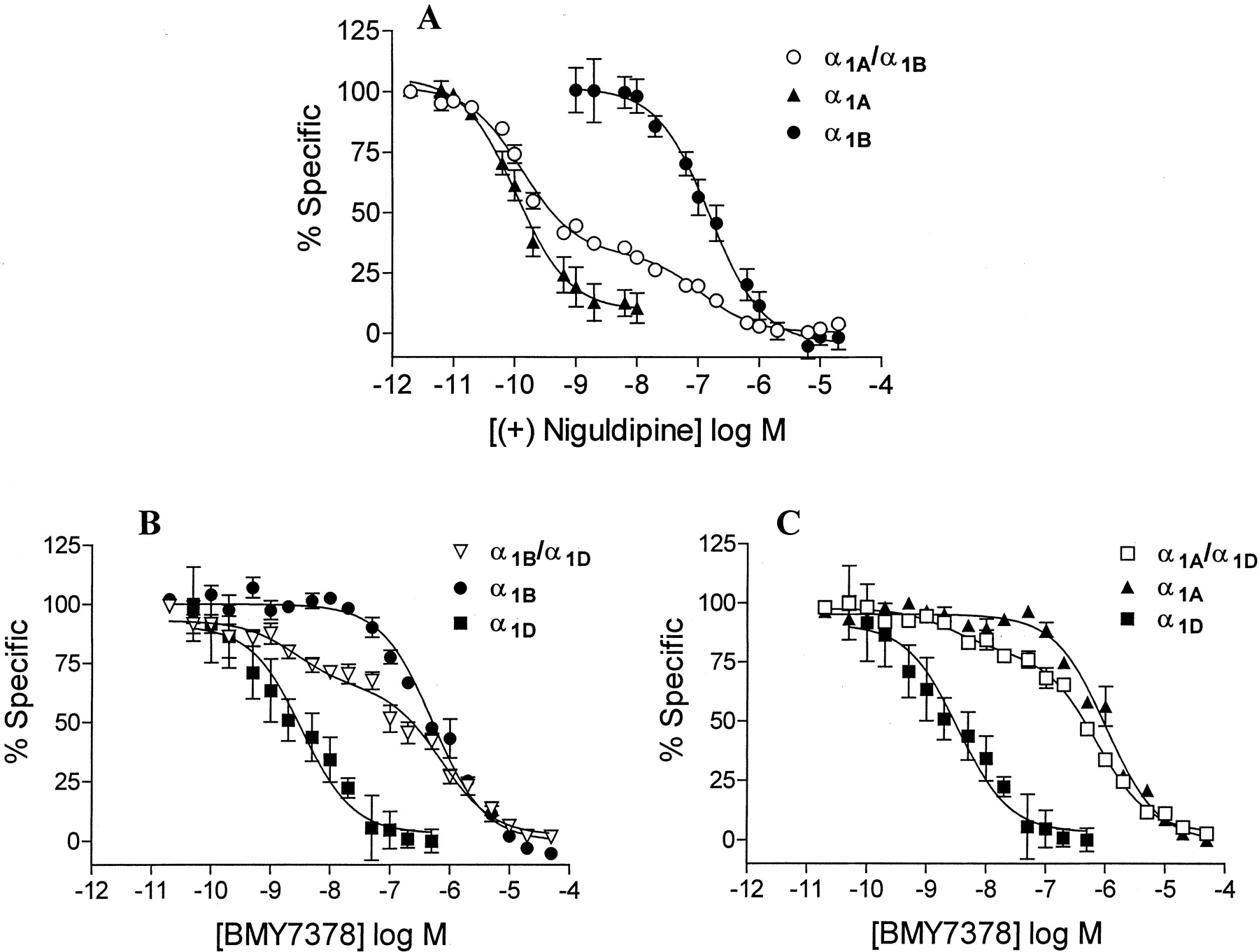
Inhibition of specific 125IBE binding by the agonist NE, the non-subtype-selective antagonist prazosin, the α1A-selective antagonist (+)-niguldipine, or the α1D-selective antagonist BMY7378 was examined in each preparation, and KI values were determined by nonlinear regression analysis (Fig. 8, Table 4). We found no changes in apparent KD values for prazosin, which binds to all cloned subtypes with similar affinities, when combinations of α1-AR subtypes were coexpressed. However, the subtype-selective antagonists (+)-niguldipine and BMY7378 showed different affinities for individual subtypes, as previously reported, and competition curves for these drugs in cells expressing combinations of subtypes were clearly biphasic (Fig. 8). Biphasic curves were separated into two components by nonlinear regression analysis to determine the fraction of high- and low-affinity binding sites and the affinity of the competing drug for each site.

Inhibition of specific 125IBE binding by subtype-selective antagonists. A, inhibition by the α1A-selective antagonist (+)-niguldipine in membranes from HEK293 cells expressing Flag-α1A, HA-α1B, or both. B, inhibition by the α1D-selective antagonist BMY7378 in membranes from HEK293 cells expressing Flag-α1D, HA-α1B, or both. C, inhibition by the α1D-AR selective antagonist BMY7378 in membranes from HEK293 cells expressing Flag-α1D, HA-α1A, or both. Data are plotted as percentage of specific binding remaining at the indicated drug concentrations. Each point is the mean ± S.E.M. from three to four different transfection experiments, each performed in duplicate.
Affinities of drugs in competing for specific 125IBE binding to α 1-AR subtypes expressed alone and together in HEK293 cells
Differentially tagged α 1-AR subtypes were expressed alone or together in HEK293 cells, membranes were harvested, and inhibition of specific 125IBE binding was determined for the drugs indicated. pKi values were calculated by nonlinear regression of one- or two-site fits as described under Materials and Methods. Values are presented as mean ± S.E.M. of three experiments performed in duplicate.
When α1A- and α1B-ARs were coexpressed, two-site analysis of inhibition of specific 125IBE binding by (+)-niguldipine (Fig. 8A) showed that 68% of binding sites had a high affinity characteristic of α1A-ARs. Receptor density in HEK293 cells expressing only α1A-ARs, only α1B-ARs, or a combination of α1A+α1B-ARs is shown in Table 2, and the density of each subtype was calculated by multiplying total Bmax by the fraction of sites showing high or low affinity for (+)-niguldipine. α1A- or α1B-ARs expressed alone showed similar Bmax values (Table 2), however the density of α1A-ARs was significantly increased (by approximately 66%) when coexpressed with α1B-ARs, whereas α1B-AR density was not significantly altered (Table 2). Densitometric analysis of Western blots of samples immunoprecipitated with anti-Flag antibody also showed an 83 ± 1.9% increase in signal strength for Flag-α1A-AR protein upon coexpression of α1A- and α1B-ARs, compared with Flag-α1A-ARs expressed alone (Fig. 9A).

A, Western blot analysis of α1-AR protein expression in membranes expressing single subtypes or mixtures of subtypes. Left, Flag-α1A-ARs (Flag-A) expressed alone or in combination with HA-α1B-ARs (HA-B). Center, Flag-α1D-ARs (Flag-D) expressed alone or in combination with HA-α1B-ARs. Right, Flag-α1D-ARs expressed alone or in combination with HA-α1A-ARs (HA-A). Membranes were solubilized, immunoprecipitated (IP) with anti-Flag affinity resin, and deglycosylated. Western blots (WB) were performed with HRP-conjugated anti-Flag antibody and are representative of three to four different transfection experiments. Arrows indicate apparent monomers and dimers of each receptor. B, cell surface expression of Flag-α1D-ARs expressed alone or in combination with HA-α1B-ARs was quantified via a luminometer-based assay as described under Materials and Methods. The values for each experiment are represented as arbitrary relative absorbance units. The bars and error bars represent the means ± S.E.M. for three to six independent experiments. *, significantly different (p < 0.05), using an unpaired t test analysis, compared with the same subtype expressed alone.
When α1D-ARs were coexpressed with either α1A- or α1B-ARs, the fraction of binding sites with a high affinity for the α1D-selective antagonist BMY7378 was used to determine the proportion of α1D-ARs. Receptor density was determined in cells expressing α1A-, α1B-, or α1D-ARs alone, α1B-+α1D-ARs, or α1A-+α1D-ARs (Table 3). The proportion of α1D-ARs was 31% for α1B+α1D and 19% for α1A+α1D-AR combinations (Fig. 8, B and C). Bmax values for each subtype were calculated and showed that α1D-AR density increased by 68% and α1B-AR density by 119% when the two subtypes were coexpressed (Table 3). However, when α1A- and α1D-ARs (which do not coimmunoprecipitate) were coexpressed, no significant increases were observed in the density of either subtype (Table 3). Compared with Flag-α1D-ARs expressed alone, densitometric analysis of Western blots also detected an 80 ± 3.0% increase in Flag-α1D-AR total protein expression only when α1B-, and not α1A-ARs, were coexpressed (Fig. 9A). These results show that coexpression of subtypes that form heterodimers (α1B+α1A or α1B+α1D) alters both receptor density and receptor protein levels, whereas coexpression of subtypes that do not form heterodimers (α1A+α1D) does not. This coincidence raises the possibility that the observed alterations in receptor density may be a functional consequence of α1-AR heterodimerization.
Increased Cell Surface Expression. The data from the ligand binding and Western blot analysis suggested that the processing and/or trafficking of α1D-ARs might be regulated by coexpression with α1B-ARs. Therefore, we used a quantitative luminometer-based assay to directly determine whether the increase in α1D-AR expression when coexpressed with α1B-ARs was related to an increase in receptor surface expression. Interestingly, several laboratories have reported that α1D-ARs tend to be localized intracellularly and are poorly expressed on the cell surface (Daly et al., 1998; McCune et al., 2000; Chalothorn et al., 2002). We determined surface expression of Flag-α1D-ARs stably expressed in HEK293 cells (∼150-250 fmol/mg) alone and when coexpressed with HA-α1B-ARs (∼300-450 fmol/mg), which were transiently expressed in these cells. As shown in Fig. 9B, there was a striking 10-fold increase in cell surface expression of α1D-ARs when coexpressed with α1B-ARs compared with α1D-ARs alone. These data reveal that coexpression of α1B-ARs facilitates efficient trafficking of α1D-ARs to the cell surface.
Discussion
Although multiple α1-AR subtypes are known to coexist within tissues and cells, the functional importance of this coexistence remains unclear. All three subtypes are known to respond to the same neurotransmitters and couple to the same major signaling mechanisms, and responses to combinations of subtypes could therefore be additive, redundant, or synergistic (Graham et al., 1996; Zhong and Minneman, 1999; Varma and Deng, 2000). However, with the recent discovery that some GPCRs form dimers that alter their pharmacological and functional properties, the potential implications of the coexistence of these subtypes must be examined more carefully. Therefore, in this study, we have examined the possibility that α1-AR subtypes homo- and heterodimerize into larger multiprotein species that may impact their pharmacological and functional properties.
Our laboratory recently reported that epitope-tagged α1A-, α1B-, and α1D-ARs form SDS-resistant homodimers and higher order oligomers when expressed in HEK293 cells. These SDS-resistant oligomers can be reduced to monomers under very strong denaturing conditions (6 M urea) (Vicentic et al., 2002), suggesting that noncovalent mechanisms are involved. This suggests that there is equilibrium between monomeric and oligomeric forms of the receptor, which can be altered by solubilization conditions. In the studies reported here, we provide direct evidence for homodimerization of all three subtypes in HEK293 cells by coimmunoprecipitation of α1-ARs with different epitope tags. Single subtypes tagged with either Flag- or HA-epitopes were coexpressed in HEK293 cells, solubilized, and immunoprecipitated with antibodies to each tag. Each subtype was found to coimmunoprecipitate with its alternatively tagged version, suggesting formation of homodimers that can be partially separated into monomers on an SDS gel under the denaturing conditions used. Interestingly, larger species with molecular masses characteristic of dimers or trimers were also observed in many of these experiments, although we do not know whether these contain the same or different tags. It seems that some homodimers are dissociated during preparation of samples for Western blots, whereas others are SDS-resistant and run as dimers after partial denaturation and electrophoresis.
Similar approaches were used to study heterodimerization between different α1-AR subtypes. Different subtypes with distinct epitope tags were coexpressed in HEK293 cells, solubilized, and immunoprecipitated with tag-specific antibodies. After electrophoresis and blotting, individual subtypes were detected using anti-tag antibodies. These experiments showed that α1B-ARs coimmunoprecipitate with either α1A-or α1D-ARs when expressed in the same cells. Because no such heterodimerization was observed when solubilized preparations from cells expressing single subtypes were mixed together, such coimmunoprecipitation does not seem to be a result of nonspecific hydrophobic interactions. Furthermore, heterodimerization does not occur via the association of partially solubilized receptors, because similar levels of heterodimers were still present after ultracentrifugation, which should pellet all partially solubilized proteins. Interestingly, coimmunoprecipitation of α1A- and α1D-ARs was not observed after coexpression, suggesting that heterodimer formation occurs only between specific combinations of subtypes.
Similar subtype-specific heterodimerization has been observed with other GPCRs, including opioid, serotonin, and somatostatin receptors (Bouvier, 2001; Rios et al., 2001). Coimmunoprecipitation studies show dimerization between κ and δ but not κ and μ opioid receptors (Jordan and Devi, 1999), whereas another study found δ and μ receptor heterodimers (George et al., 2000). Interestingly, recent work suggests that several different AR combinations, including β1- and β2-ARs (Lavoie et al., 2002) and β1- and α2A-ARs (Xu et al., 2003), which coexist in many tissues, can also form heterodimers that modulate receptor internalization and signaling. Taken together, these studies suggest that subtype-specific GPCR heterodimer formation may be widespread, with multiple functional consequences.
The effect of agonist activation on GPCR dimerization varies depending on the receptors involved. Agonist occupancy has been shown to increase, decrease or have no effect on dimerization of different GPCRs (Bouvier, 2001; Rios et al., 2001). We found that stimulation of intact cells with NE did not alter the levels of homodimers of any α1-AR subtype or the extent of α1A/α1B or α1D/α1B heterodimers and did not induce formation of α1A/α1D heterodimers. These results suggest that both homo- and heterodimerization of α1-ARs is constitutive and agonist-independent, similar to results obtained with κ opioid and M3 muscarinic receptor homodimers, and angiotensin1/bradykinin B2 heterodimers (Jordan and Devi, 1999; Jordan et al., 2000).
Mechanisms that have been implicated in GPCR dimerization include interactions between specific motifs within transmembrane domains, intracellular or extracellular loops, or N- or C-terminal extensions. For example, metabotropic glutamate receptors form dimers via covalent disulfide bonds in their very long extracellular N-termini, GABAB receptor C-termini play a role in GABAB receptor dimerization, and chimeric α2A-AR-m3 muscarinic receptors require the third intracellular loops for dimerization (Bouvier, 2001; Rios et al., 2001). We first examined whether the relatively long C-tails of α1-AR subtypes might be involved in dimerization by the use of C-truncated mutants. We found C-truncated receptors formed both homo- and heterodimers similar to those formed by full-length receptors, suggesting that their C-termini are not required for dimerization. Similar results have been observed with C-truncation mutants of δ opioid and dopamine D3 receptors (Cvejic and Devi, 1997). We also examined the relatively long N terminus of α1D-ARs to determine whether it was required for α1-AR homo- or heterodimerization and found similar results. Mutated α1D-ARs lacking the N-terminal 79 aa formed SDS-resistant homodimers similar to full-length receptors (Pupo et al., 2003), and also formed heterodimers with α1B-, but not α1A-ARs, also similar to full-length α1D-ARs. These results suggest that dimerization of α1-ARs does not require C- and N-terminal domains, suggesting that α1-AR oligomerization may occur by a transmembrane domain-swapping model, as has been proposed previously for β2-ARs (Hebert et al., 1996).
Expression of both homo- and heterodimers of α1-ARs on the cell surface was examined by use of a membrane-impermeant biotinylating reagent (Bai et al., 1998) and subsequent interaction with streptavidin These data suggest that the observed α1-AR dimers and oligomers all seem to be expressed on the cell surface. This was particularly noticeable for α1D-ARs, which often have low expression levels in HEK293 cells, and in which the primary species seem to be homodimers and trimers (Vicentic et al., 2002). In fact, the pattern of expression of each subtype was similar for total and cell surface receptor expression, with mixtures of monomers and dimers predominating for α1A-, monomers for α1B-, and dimers and trimers for α1D-ARs. This suggests that homo- and hetero-oligomerization is not specifically confined to a single subcellular compartment.
One possible consequence of α1-AR dimerization might be changes in the pharmacological properties of the receptor binding sites. In fact, there is existing evidence for additional α1-AR subtypes that cannot be explained by the three known cloned subtypes (Endoh, 1996; Ford et al., 1996, 1997). Changes in pharmacological properties have been observed previously with opioid, somatostatin, and dopamine (D2/D3) receptor heterodimers, which have novel ligand binding properties compared with subtypes expressed alone (Jordan and Devi, 1999; Scarselli et al., 2000). However, other GPCRs, such as 5-HT1B/5-HT1D serotonin receptor and opioid/adrenergic receptor (δ-OR/β2-AR) heterodimers, show no apparent differences in binding properties upon heterodimerization (Xie et al., 1999; Jordan et al., 2001). We used radioligand binding to examine the affinities of agonists and antagonists in membranes from cells expressing α1-AR subtypes alone or in different combinations. We found no changes in apparent KD values for the nonsubtype selective antagonist prazosin when combinations of α1-AR subtypes were coexpressed, suggesting that heterodimerization does not explain the “α1L” subtype previously observed in functional studies of smooth muscle (Ford et al., 1996; Muramatsu et al., 1998). However, we cannot rule out the possibility that the α1L subtype is not labeled by 125IBE. We also examined the subtype-selective antagonists (+)-niguldipine (α1A) and BMY7378 (α1D). These drugs showed different KI values for individual subtypes expressed alone as expected, but competition curves for these drugs in cells expressing combinations of subtypes were clearly biphasic. Nonlinear regression analysis showed that inhibition curves from cells coexpressing combinations of subtypes were best fit by a two-site model, with each of the sites characteristic of a single subtype expressed alone. For example, inhibition of specific 125IBE binding in cells coexpressing α1A- and α1B-ARs by (+)-niguldipine resulted in a two-site fit; one site had an affinity characteristic of the α1A-subtype and the other was characteristic of the α1B-subtype expressed alone. Similar results were obtained with the α1D-selective antagonist BMY7378 when α1D-ARs were coexpressed with other subtypes. These results suggest that α1-AR heterodimers do not display any unexpected pharmacological profiles but that their pharmacological properties seem to be caused by combinations of binding sites contributed by individual subtypes. Thus, heterodimerization of α1-ARs is not likely to account for the properties of novel subtypes that have been proposed in native tissues.
In contrast to the lack of effect of subtype coexpression on receptor pharmacological properties, the surface expression of α1D-ARs was strikingly increased upon coexpression with α1B-ARs, suggesting that α1B-ARs may assist in transporting α1D-ARs to the cell surface. Furthermore, coexpression of α1B-ARs with α1A- and α1D-ARs significantly increased the density of α1A- and α1D-AR binding sites and the amount of receptor protein observed on Western blots. The mechanisms by which such effects occur remain to be elucidated. It is interesting that heterodimerization is important in trafficking the GABAB receptor heterodimer to the cell surface, where the R2 subtype acts as a molecular chaperone and is essential for proper transport of the R1 subtype (Margeta-Mitrovic et al., 2001). In addition, dimerization of vasopressin and dopamine receptor C-truncated mutants with wild-type vasopressin V2 and dopamine D3 receptors prevents cell surface expression, supporting a role for dimerization in receptor processing and transport (Karpa et al., 2000; Zhu and Wess, 1998). Our data raise the possibility that heterodimerization may enhance the efficiency of α1-AR processing and/or trafficking, resulting in increased receptor expression.
Interestingly, a number of previous studies have suggested that a large fraction of α1D-ARs are localized intracellularly, with a limited cell surface expression (Daly et al., 1998; McCune et al., 2000; Chalothorn et al., 2002). For example, α1D-ARs fused with green fluorescent protein seem to be mainly cytosolic in HEK293 cells (Chalothorn et al., 2002). These findings support previous observations on low expression levels and poor coupling efficiencies of human α1D-ARs after heterologous expression in different cell lines (Perez et al., 1993; Esbenshade et al., 1995; Theroux et al., 1996; Taguchi et al., 1998), raising the possibility that this subtype cannot be efficiently folded and translocated to the surface membrane on its own. On the other hand, the α1B subtype has been found predominantly on the cell surface in most studies (Hirasawa et al., 1997; Daly et al., 1998; McCune et al., 2000; Chalothorn et al., 2002). Thus, our results suggest that coexpression of the surface-directed α1B-AR promotes surface trafficking of the α1D subtype, possibly by direct heterodimerization, although this remains to be further investigated.
It is known that rat cardiomyocytes express all three α1-AR subtypes (Noguchi et al., 1993, 1995; Zhong and Minneman, 1999) and that rat and human vascular smooth muscle cells express heterogeneous mixtures of subtypes (usually α1B-with α1A-or α1D-ARs) (Price et al., 1994; Graham et al., 1996; Piascik et al., 1997). Interestingly, studies in the peripheral vascular system show that α1B-AR mRNA is widespread, although this subtype does not seem to be involved in contraction (except in mesenteric resistance arteries) (Piascik et al., 1997). All three α1-AR subtypes are also expressed in several human brain regions, including cerebral cortex and cerebellum, where their functions may modulate sleep, arousal, and other important behavioral processes but remain largely unknown (Price et al., 1994; Graham et al., 1996). The widely observed coexistence of these subtypes in many different cells and tissues supports a potential role for heterodimerization in control of expression or trafficking of particular subtypes.
In summary, we used coimmunoprecipitation to show that differentially-tagged human α1-AR subtypes form agonist-independent homodimers that are expressed on the cell surface. We have also presented the first evidence for formation of heterodimers by certain combinations of α1-AR subtypes but not others. These heterodimers are constitutively present, are located on the cell surface, and require neither the extracellular amino terminus nor the intracellular carboxyl tail. Radioligand binding studies suggest that heterodimerization of α1-ARs does not change their affinities for norepinephrine or antagonists, suggesting that dimerization does not result in pharmacologically distinct subtypes. Coexpression with α1B-ARs significantly increases receptor density and total protein expression of both α1A- and α1D-AR subtypes and also significantly increases the proportion of α1D-ARs on the cell surface. These findings suggest that heterodimerization of human α1-AR subtypes are functionally relevant in the regulation of receptor processing and/or trafficking.
Acknowledgments
We thank Drs. Andre Pupo and Rebecca Wright for helpful discussions and experimental support and Mr. George Rogge for excellent technical support.
Footnotes
-
Supported by National Institutes of Health grants to KPM and RAH. RAH thanks the W.M. Keck Foundation for a Distinguished Young Scholar in Medical Research award.
-
ABBREVIATIONS: GPCR, G protein coupled receptor; AR, adrenoceptor; HEK, human embryonic kidney; BMY7378, 8-[2-[4-(2-methoxyphenyl)-1-piperazinyl]ethyl]-8-azaspiro[4.5]decane-7,9-dione dihydrochloride; HRP, horseradish peroxidase; HA, hemagglutinin,; ECL, enhanced chemiluminescence; DβM, n-dodecyl-β-d-maltoside; PBS, phosphate-buffered saline; PNGase F, N-glycosidase F; TBST, Tris-buffered saline containing 0.1% Tween-20; PAGE, polyacrylamide gel electrophoresis; sulfo-LC-NHS-biotin, sulfosuccinimidyl-6-(biotinamido) hexanoate; BE 2254, 2-[β-(4-hydroxyphenyl)-ethylaminomethyl]-tetralone; 125IBE, 125I-BE 2254; NE, norepinephrine.
- Received January 28, 2003.
- Accepted August 29, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}