


Abstract
Regulators of G protein signaling (RGS) and RGS-like proteins are a family (>30 members) of highly diverse, multifunctional signaling proteins that bind directly to activated Gα subunits. Family members are defined by a shared RGS domain, which is responsible for Gα binding and markedly stimulates the GTPase activity of Gα subunits leading to their deactivation and termination of downstream signals. Although much has been learned in recent years about the biochemistry of RGS/Gα interactions, considerably less is known about the broader cellular roles and regulation of RGS proteins. Recent findings indicate that cellular mechanisms such as covalent modification, alternative gene splicing, and protein processing can dictate the activity and subcellular localization of RGS proteins. Many family members also directly link G proteins to a growing list of signaling proteins with diverse cellular roles. New findings indicate that RGS proteins act not as dedicated inhibitors but, rather, as tightly regulated modulators and integrators of G protein signaling. In some cases, RGS proteins modulate the lifetime and kinetics of both slow-acting (e.g., Ca2+ oscillations) and fast-acting (e.g., ion conductances, phototransduction) signaling responses. In other cases, RGS proteins integrate G proteins with signaling pathways linked to such diverse cellular responses as cell growth and differentiation, cell motility, and intracellular trafficking. These and other recent studies with animal model systems indicate that RGS proteins play important roles in both physiology and disease. Recognition of the central functions these proteins play in vital cellular processes has focused our attention on RGS proteins as exciting new candidates for therapeutic intervention and drug development.
I. Introduction
Our understanding of G protein signaling has undergone fundamental changes in recent years. Established models based on information gathered over the last quarter century suggest that most hormones, neurotransmitters, and sensory input rely upon a G protein-coupled receptor (GPCR1), a heterotrimeric guanine nucleotide-binding regulatory protein (G protein), and a limited number of well described downstream effector proteins (e.g., adenylyl cyclases, phospholipases) and chemical second messengers to transmit their signals across the plasma membrane (Bourne et al., 1990; Simon et al., 1991; Hepler and Gilman, 1992; Hamm, 1998). However, recent studies indicate that GPCRs and G proteins engage a growing list of newly appreciated proteins and linked signaling pathways to carry out their cellular functions (Bockaert and Pin, 1999;Hall et al., 1999). Prominent among these new binding partners are theregulators of G protein signaling (RGS proteins). RGS proteins are a large family of highly diverse, multifunctional signaling proteins, which share a conserved signature domain (RGS domain) that binds directly to activated Gα subunits to modulate G protein signaling. RGS proteins differ widely in their overall size and amino acid identity, and many family members possess a remarkable variety of structural domains and motifs that regulate their actions and/or enable them to interact with protein binding partners with diverse cellular roles (Hepler, 1999; Siderovski et al., 1999). Several comprehensive reviews have appeared recently, which examine RGS biochemistry and cellular functions from different perspectives (Burchett, 2000; De Vries et al., 2000; Ross and Wilkie, 2000; Zhong and Neubig, 2001).
Although considerable information is now available describing the biochemical and cellular properties of RGS proteins as blockers of G protein signaling, less is known about cellular mechanisms that regulate RGS functions per se. In addition, although early evidence suggested that RGS proteins acted primarily as negative regulators of G protein signaling, recent findings indicate that these proteins act as tightly regulated modulators and/or as multifunctional integrators of G protein signaling. This review will highlight emerging concepts regarding RGS proteins as modulators and integrators of multiple signaling pathways and focus on new information regarding cellular mechanisms that regulate RGS functions. A brief discussion will also center on roles of RGS proteins in physiology and their potential as therapeutic targets.
II. RGS Proteins Directly Regulate G Protein Activity
A. G Protein Activation and Deactivation and Early Evidence for RGS Proteins
G proteins consist of Gα, Gβ, and Gγ subunits, and Gα subunits bind and hydrolyze GTP to act as molecular switches. Agonist occupancy of GPCRs stimulates the exchange of GTP for GDP on Gα subunits and subunit dissociation, and the amplitude and lifetime of G protein-directed signaling events are dictated by the lifetime of GTP on Gα. Purified Gα subunits in solution are inefficient GTPases with intrinsic rates of GTP hydrolysis in the range of 0.1 to 0.3 Pi/mol Gα/min for most Gα (Gilman, 1987). In the absence of receptor, GTP hydrolysis is limited by the rate of GDP release from Gα following hydrolysis. Agonist-occupied GPCRs stimulate the release of GDP from Gα, opening the guanine nucleotide binding pocket for rapid binding of abundant intracellular GTP. Thus, in the presence of receptors and agonist using reconstituted systems, the observed rates of Gα-directed GTP hydrolysis are enhanced 10-fold or more and are reflective of intrinsic rates of GTP hydrolysis (Gilman, 1987; Ross and Wilkie, 2000).
However, for many G protein-regulated signaling responses, their rates of deactivation in a cellular context are much faster (100- to 300-fold) than is predicted from observed rates of Gα-GTP hydrolysis using purified components. G protein signaling events in the retina, brain, and heart proceed on a much faster time scale than is the case with other non-electrically excitable tissues. For example, G protein-directed (i.e., Gt or transducin) phototransduction in intact rod outer segments begins and ends within milliseconds, with recovery times of less than 200 ms. These rates are much faster than is expected from known rates of rhodopsin and Gαt-mediated GTP hydrolysis in light-stimulated rod outer segment membranes (for review, see Arshavsky and Pugh, 1998). Similar discrepancies between in vitro and in vivo data were also observed for rates of deactivation of G protein-regulated potassium and calcium channels (for review, seeZerangue and Jan, 1998). These observations predicted the existence of unidentified factors or proteins that regulate the rates of Gα-GTP hydrolysis to fine-tune G protein activation state and signaling responses.
B. Discovery of RGS Proteins
Initial evidence for cellular regulators of G protein signaling came from genetic studies of lower eukaryotes (Dohlman and Thorner, 1997; for review, see Koelle, 1997). Studies in yeast nearly two decades ago recognized a gene product (Sst2p) that, when mutated, presented a phenotype that was supersensitive to G protein-directed pheromone responses (Chan and Otte, 1982; Weiner et al., 1993; Dohlman et al., 1995). A similar gene (flbA) was identified as a negative regulator of G protein signaling responses in the fungal organism Aspergillus nidulans (Lee and Adams, 1994). Other investigators studying mammalian systems independently identified a novel gene (GOS8) that was rapidly up-regulated in stimulated monocytes (Siderovski et al., 1994), and a new protein that bound activated Gαi3 in yeast two-hybrid screens, which was termed G alphainteracting protein (GAIP, or later RGS-GAIP) (De Vries et al., 1995). Although cellular roles for GOS8 and RGS-GAIP remained obscure at that time, GOS8 was recognized to share a novel conserved domain with other mammalian proteins (Siderovski et al., 1996). Full appreciation that each of these proteins belonged to a larger superfamily of signaling proteins came from subsequent genetic studies of Caenorhabditis elegans describing a gene (egl-10) that negatively regulated Gαo-directed locomotion and egg-laying behavior (Koelle and Horvitz, 1996). Partial nucleotide sequences for 15 mammalian genes were identified from a brain cDNA library that shared a conserved 130-amino acid core domain with the egl-10,sst2p, flbA, and GOS8 genes. These proteins were termed regulators of G proteinsignaling and numbered consecutively (RGS1–RGS15), and the conserved domain was termed the RGS domain, henceforth recognized as the protein family hallmark. The previously discovered GOS8 was identical with one sequence and was renamed RGS2, and RGS7 was recognized as the mammalian homolog of Egl-10. Separate studies showed that mammalian RGS4 could substitute for Sst2p as an inhibitor of pheromone responses in yeast, demonstrating a conservation of RGS function across species (Druey et al., 1996). Since that time, full-length cDNA for these and other mammalian RGS proteins have been reported to reveal a large family of highly divergent, multifunctional proteins (Fig. 1; Table1).
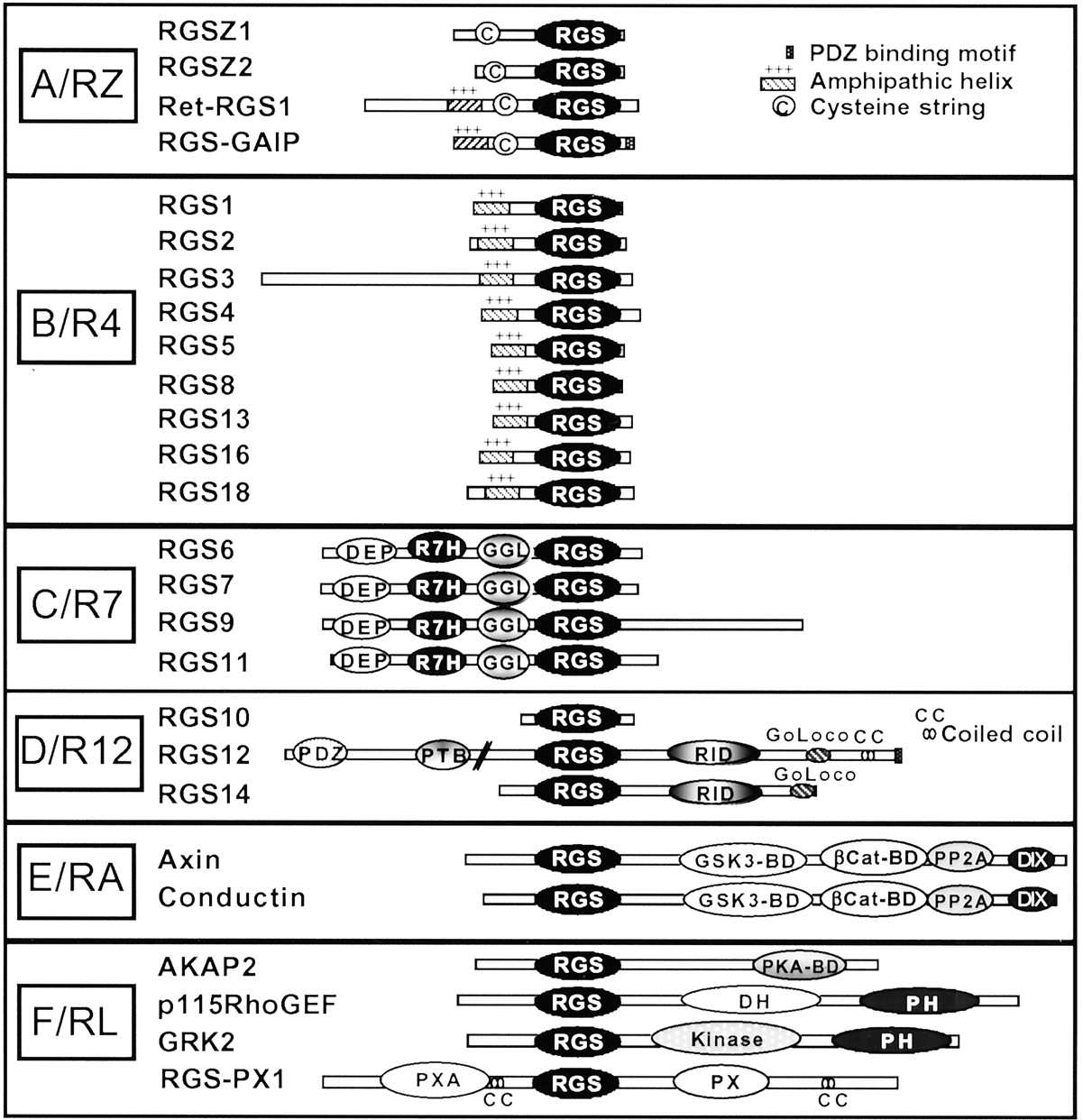
Structures and classification of mammalian RGS and RGS-like proteins. RGS and RGS-like proteins are classified into subfamilies based on alignment of RGS domain amino acid sequences (Zheng et al., 1999; Ross and Wilkie, 2000). Proteins are oriented with their N termini on the left and their C termini on the right. See the text for a description of the domains and motifs.
Mammalian RGS proteins: general information and binding partners
C. RGS Proteins Are GTPase-Activating Proteins for Gα
Although RGS proteins were first identified as negative regulators of G protein signaling, the biochemical mechanisms whereby these proteins regulated Gα signaling were unknown. G proteins act as molecular switches, and RGS proteins could block Gα signaling by preventing GTP binding to Gα or by limiting the lifetime of GTP bound to Gα. Following the discovery of RGS proteins, a series of studies demonstrated that various RGS proteins act as GTPase-activating proteins (GAPs) to greatly accelerate (up to 1000-fold) the rate of Gα-GTP hydrolysis and limit the lifetime of the active Gα-GTP species (Berman et al., 1996b; Hunt et al., 1996; Watson et al., 1996;Hepler et al., 1997; Kozasa et al., 1998). These and other studies demonstrated that RGS proteins bind directly and preferentially to the active GTP bound forms of Gαi/o, Gαq, Gα12/13, or Gαs and that RGS domains exhibit highest affinity for the GDP-Mg2+-AlF4− bound Gα, which mimics the transition state during GTP hydrolysis (for review, see Berman and Gilman, 1998 or Ross and Wilkie, 2000). Additional studies demonstrated that RGS proteins can also bind tightly to active Gα to block effector activation independent of GAP activity by acting as effector antagonists (Hepler et al., 1997; Carman et al., 1999). For a comprehensive discussion of the biochemical properties of RGS proteins as GTPase-activating proteins, see Ross and Wilkie (2000).
D. Structure and Classification of RGS Proteins
Completion of the human genome project has confirmed the existence of more than 30 distinct proteins that contain an RGS or an RGS-like (RL) domain (see below; Fig. 1 and Table 1). Based on amino acid identities within the conserved RGS domain, two independent research groups have classified RGS proteins into six distinct subfamilies (Zheng et al., 1999; Ross and Wilkie, 2000). These groupings also correlate well with overall structure and identified functions within subfamilies. In one classification, subfamily names are arbitrarily designated A–F (Zheng et al., 1999) whereas in the other classification (Ross and Wilkie, 2000), subfamily names are derived from a prototypical RGS protein member (e.g., the RZ subfamily is typified by RGSZ). Both classifications are consistent except for the assignment of RGS10, which in one case is not assigned to a subfamily (Zheng et al., 1999) and in the other is grouped in the R12 subfamily based on RGS domain similarities (Ross and Wilkie, 2000). The six groupings (Fig. 1; Table 1) include the A or RZ subfamily (prototype RGSZ); the B or R4 subfamily (prototype RGS4); the C or R7 (prototype RGS7); the D or R12 subfamily (prototype RGS12); the E or RA subfamily (prototype Axin); and the F or RL subfamily (containing proteins with RL domains). The RL domains of this subfamily are only distantly related to each other and to other RGS domains both in amino acid sequence identities and in Gα recognition. Proteins containing RL domains include D-AKAPs (dual specificity A kinase anchoring proteins), p115RhoGEFs, RGS-PX1, and G protein receptorkinases (GRKs). Unlike the other RGS subfamilies, proteins within the RL category are a collection of miscellaneous proteins, classified together only because they each contain a weakly homologous RGS domain. Members of the RZ and R4 subfamilies, with two exceptions (RGS3 and RET-RGS1), are small 20- to 30-kDa proteins that contain short N- and C-terminal regions flanking the RGS domain. In contrast, members of the R7, R12, RA, and RL subfamilies, with one exception (RGS10), are much larger proteins (up to 160 kDa) that possess longer N and C termini encoding various binding domains and motifs for other proteins.
Aside from a shared RGS domain, RGS proteins differ widely in their overall size and amino acid identity, and possess a remarkable variety of structural domains and motifs (Fig. 1). Unlike members of the A/RZ and B/R4 subfamily, which are simple proteins with little more than an RGS domain, members of the C/R7, D/R12, E/RA, and F/RL subfamilies each have additional subfamily-specific domains. All members of the C/R7 subfamily (RGS6, RGS7, RGS9, RGS11) contain a DEP (disheveled, Egl-10, pleckstrin) domain, a previously unknown conserved region (Sondek and Siderovski, 2001), which we call the R7 homology or R7H domain, and a GGL (G protein gamma subunit-like) domain. Members of the D/R12 family (RGS12, RGS14) share a RBD (Rap1/2-binding domain) and a GoLoco motif. Members of the E/RA family (Axin, Conductin) each contain a glycogen synthase kinase3β-binding domain (GSK3β), a β-catenin binding site (Cat), a protein phosphatase 2A(PP2A) homology region, and a dimerization domain (DIX). Within the RL subfamily, the RhoGEF proteins each contain DH (dblhomology) and PH (pleckstrinhomology) domains, the GRK proteins each contain a Ser/Thr kinase catalytic domain, whereas RGS-PX1 contains Phox homology (PX) and Phox-associated (PXA) domains. Individual RGS proteins within subfamilies also contain additional protein-specific domains (Fig. 1). These structural features impart to RGS proteins the capacity to interact with a growing list of protein binding partners that mediate RGS signaling, RGS subcellular targeting, and regulation of RGS functions. Reported non-G protein binding partners for RGS proteins are illustrated in Fig. 4, and their roles as regulators or mediators of RGS signaling functions are discussed elsewhere within the text.

RGS protein binding partners. Reported RGS binding partners are illustrated along with their subcellular localization. For protein identities and a discussion of potential cellular roles, see the text. For reported RGS/Gα interactions, see Table 1.
E. Simple versus Complex RGS Proteins
Given the extraordinary diversity of RGS proteins and their functions, emerging ideas suggest that the smaller simpleRGS proteins (primarily those of the A/RZ and B/R4 subfamilies) serve almost exclusively as negative regulators of G protein signaling. However, the functions of these proteins are tightly regulated such that they act as modulators rather than dedicated inhibitors of G protein signaling. In contrast, the larger RGS family members (C/R7, D/R12, E/RA, and F/RL subfamilies) are multifunctional proteins that have the capacity to bind both G proteins and other signaling proteins. As such, these complex RGS proteins act asintegrators of G protein signaling, possibly as novel G protein effectors or as scaffolding proteins. In this regard, the complex RGS proteins are similar to the growing list of signaling proteins that share a single modular domain but are functionally dissimilar, e.g., proteins that contain SH2 or SH3 domains (see Zhong and Neubig, 2001). We will discuss examples of simple RGS proteins that serve as modulators of G protein signaling, examples of complex RGS proteins as integrators of G protein signaling and then examine mechanisms that regulate their cellular functions.
III. RGS Proteins Modulate G Protein Signaling
A. RGS4 Modulation of Gq/11-Directed Ca2+Signaling
RGS4 was among the first RGS proteins discovered (Druey et al., 1996; Koelle and Horvitz, 1996) and its biochemical and cellular properties have been studied more extensively than those of any other family member. Thus, we have chosen to focus our discussion on RGS4 as the best developed (although still incomplete) case study for understanding mechanisms that underlie RGS modulation of G protein signaling. RGS4 is the prototypical member of the B/R4 family, of which nearly all members are relatively simple proteins composed of an RGS domain flanked by minimal N and C termini lacking prominent modular domains (Fig. 1). Because of their relative simplicity, emerging ideas suggest that the principle cellular role for these proteins is to modulate G protein signaling through their RGS domains, while their N and C termini dictate RGS subcellular localization and signaling capacity. Consistent with this idea, a large body of literature demonstrates that heterologous expression of recombinant forms of RGS4 and other simple B/R4 family members blocks receptor and G protein signaling (for review, see Burchett, 2000; De Vries et al., 2000; Zhong and Neubig, 2001). However, growing evidence now suggests that endogenous RGS proteins act not as simple inhibitors of G protein signaling but, instead, as tightly regulated modulators that fine tune G protein signaling events in a cell- and context-dependent manner. To illustrate this point, we will focus on emerging models of cellular roles for RGS4 as a modulator of Gq/11 and Ca2+ signaling.
Recombinant RGS4 is an effective GAP for both Gαi/o family members and Gαq, and its heterologous high-level expression blocks both Gαi/o-mediated signaling events and Gq/11-directed inositol lipid/Ca2+ signaling in mammalian cells. However, in the case of RGS4 and other simple RGS proteins, where examined, native protein levels are typically low in host cells even when their mRNA levels are high. When RGS4 and other simple B/R4 family members are introduced into permeabilized cells at low levels reflective of their physiological concentrations, these proteins do not block receptor and Gq/11-directed Ca2+ signaling. Instead, these RGS proteins quite unexpectedly elicit rhythmic Ca2+ oscillations (Xu et al., 1999; Luo et al., 2001), suggesting that the observed complete blockade of G protein responses by RGS in other circumstances may be a consequence of overexpressing the protein.
Sufficient information is now available to propose a working model that describes cellular roles and regulation of RGS4 as a modulator of the rhythmic Ca2+ oscillations elicited by many hormones and neurotransmitters (Thomas et al., 1996). This model (illustrated in Fig. 2) is derived in large part from a recently proposed hypothesis (Sierra et al., 2000;Luo et al., 2001), and as is true of all models, the supporting findings are open to other interpretations. The chief limitation of this hypothesis is that many of the supporting observations have not yet been independently confirmed by other laboratories. In addition, some of the cellular components of the model have been extrapolated from in vitro studies, and other supporting studies ignore the fact that B/R4 RGS family members also modulate Gαi/o signaling in parallel. Thus, this model should be considered not as fact but, instead, as a provocative and testable scenario for describing cellular mechanisms whereby simple RGS proteins may act as highly regulated modulators of GPCR signaling. We will discuss each step of the model in some depth to better understand possible mechanisms that regulate the contribution of RGS4 to this process.

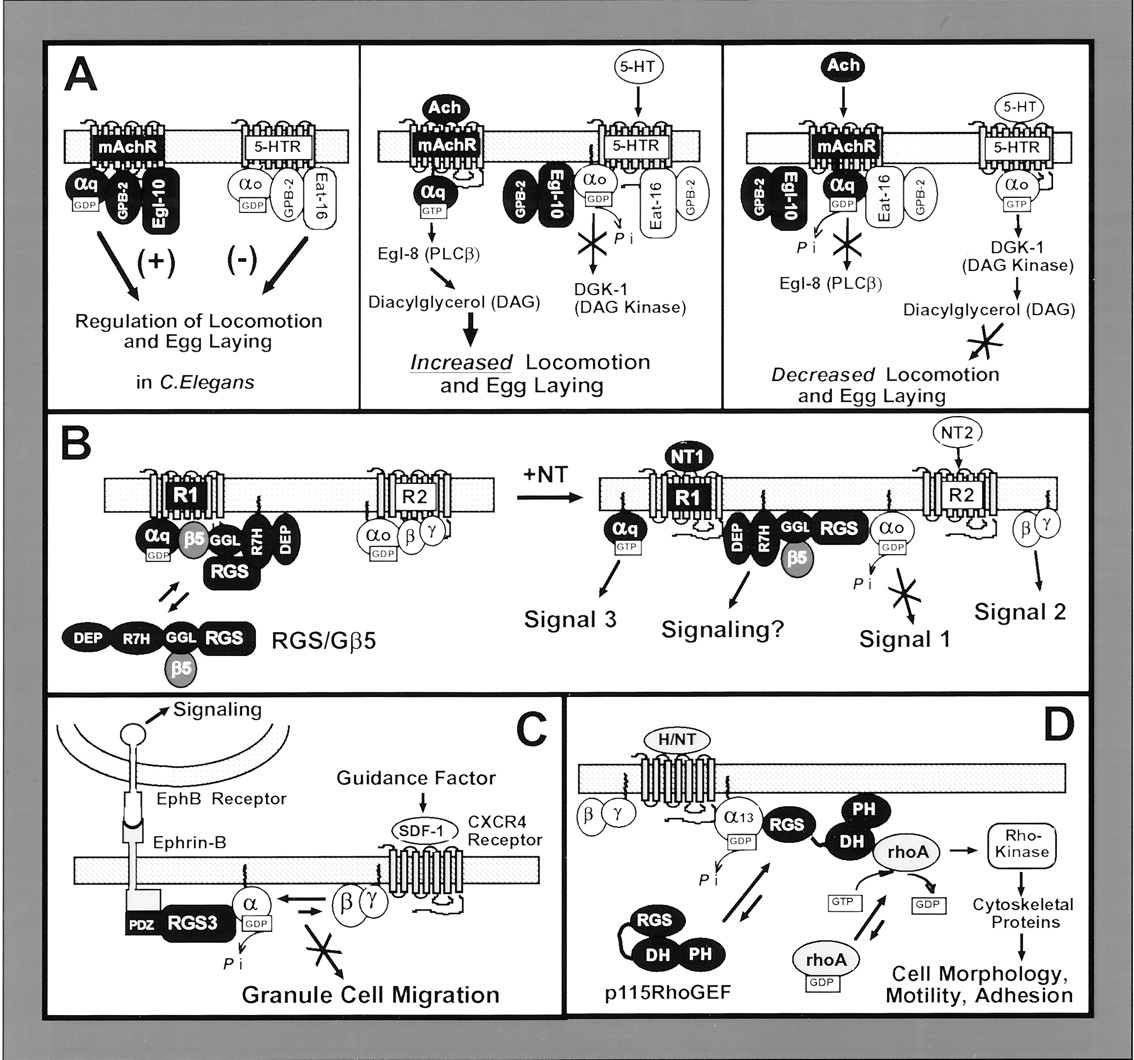
Proposed model depicting RGS4 modulation of Ca2+ oscillations in mammalian cells. The following model is based on recently proposed ideas (Popov et al., 2000; Luo et al., 2001) and supporting data (see text and references therein). Top panel, schematic diagram illustrating peak amplitudes (1 and 3) and refractive periods (2 and 4) of rhythmic, oscillatory Ca2+ spikes in cells. Corresponding diagrams illustrating the proposed role of RGS4 in modulating each stage of the Ca2+ oscillations (1–4) are presented in panels 1–4. Panel 1, hormone (H) activation of GPCR stimulates the Gq/IP3/Ca2+ pathway, resulting in RGS4 membrane recruitment and receptor association. Depicted are associations of the amphipathic helix on the N terminus of RGS4 (+++) with anionic lipids in the plasma membrane (- - -). Panel 2, RGS4 forms a complex with GPCR, Gαq-GTP, and PLCβ and exerts its GAP effects on Gαq to shut off Ca2+ mobilization. Gβγ activates PI3K, which synthesizes PIP3. Panel 3, PIP3 binding to RGS4 inhibits its GAP activity toward Gαq, allowing resumption of IP3 production and Ca2+ mobilization. Ca2+ (●) activated CaM competes with PIP3 for binding to RGS4. Panel 4, Ca2+/CaM binding uncouples RGS4 from PIP3 but does not inhibit RGS4 GAP activity. RGS4 reassociates with the Gαq/GPCR complex to shut off IP3/Ca2+ production. As Ca2+ levels fall, CaM is deactivated and dissociates from RGS4, allowing rebinding of PIP3. See text for further discussion.
1. Cellular Mechanisms That Influence RGS4 Membrane Recruitment and Attachment.
RGS4 (as well as other RGS proteins) is predicted to exist as a soluble hydrophilic protein, but it is found both in the cytosol and tightly bound to membranes (Srinivasa et al., 1998;Bernstein et al., 2000). To modulate Gαq/11 and Gαi signaling events, RGS4 needs to be present at the cytoplasmic face of the plasma membrane. Indeed, recombinant RGS4 and other B/R4 family members are recruited from cytosol to membranes by activated forms of these Gα subunits (Srinivasa et al., 1998; Druey et al., 1999; Heximer et al., 2001). However, in reconstituted systems using purified proteins, the isolated N terminus of RGS4 can associate rapidly and irreversibly with anionic lipid vesicles independent of whether receptor and/or Gα and Gβγ subunits were present (Tu et al., 2001). Apparently, Gα and Gβγ subunits can enhance but are not necessary for constitutive RGS4 membrane association (Dowal et al., 2001). Structural features on RGS4 responsible for its membrane attachment have been identified. The N terminus of RGS4 contains a 33-amino acid cationic amphipathic α helix that drives RGS4 membrane attachment (Bernstein et al., 2000; Tu et al., 2001). RGS4 is also reversibly palmitoylated near its N terminus at Cys2 and, to a lesser extent, Cys12 (Bernstein et al., 2000). However, acylation does not seem to be essential for RGS4 binding to anionic lipid vesicles but does appear to accelerate the process, likely due to its hydrophobic contributions (Tu et al., 2001). N-terminal palmitoylation also targets RGS4 to specialized cholesterol and glycosphingolipid-rich vesicles in vitro, and it has been suggested that reversible acylation may target RGS4 and other RGS proteins to specialized lipid rafts within the plasma membrane (Moffett et al., 2000).
These findings suggest that RGS4 can associate with membranes spontaneously, and that any G protein contributions to this process likely depend on the activation state of the receptor/G protein complex in cells. Germane to this idea, RGS4 association with anionic membranes also greatly increases its GAP activity and that of other RGS proteins toward target Gα (Tu et al., 2001), suggesting that other factors or binding partners at the lipid membrane influence RGS selectivity and potency toward target Gα in cells.
2. Once Bound to Membranes, What Factors Influence RGS4 Specificity for Target Gα in Cells?
In solution-based reconstituted systems using purified proteins, RGS4 is quite promiscuous (as are most other simple RGS proteins) and can block Gαi and Gαq signaling functions in vitro (Berman et al., 1996a;Hepler et al., 1997) and when introduced into intact cells (Druey et al., 1996; Huang et al., 1997a; Yan et al., 1997; Heximer et al., 1999). However, under experimental conditions where protein can be introduced directly to cells at defined concentrations, RGS4 appears to regulate Gα function based on recognition of receptors rather than association with Gα. In pancreatic acinar cells, carbachol, bombesin, and CCK each stimulate Ca2+ signaling to similar extents by activating Gq/11-linked to their respective receptors. Wilkie, Muallem, and coworkers demonstrated that introduction of purified RGS4 directly into these cells selectively inhibited inositol lipid/Ca2+ signaling by carbachol at concentrations that were 4- and 33-fold more potent than required to block bombesin- and CCK-directed Ca2+ signaling, respectively (Xu et al., 1999). The B/R4 family members RGS1 and RGS16 also displayed receptor selectivity, and RGS1 was nearly 1000-fold more potent at blocking carbachol- than CCK-directed Ca2+ signaling (Xu et al., 1999). In stark contrast, RGS2 displayed no preference between the three receptors. This receptor selectivity of RGS4 is conferred by its N terminus since truncated RGS4 lacking this domain exhibited reduced potency and no receptor selectivity. Furthermore, RGS4 potency and selectivity for affecting muscarinic receptor-directed Ca2+signals was restored by combined addition of the N terminus and the RGS core domain (Zeng et al., 1998). Considering all of these findings together, a plausible explanation forwarded by the authors is that a feature associated with the N terminus of RGS4 selectively recognizes certain receptors but not necessarily the linked Gα. Although there is no direct evidence that RGS4 physically contacts receptors, this possibility cannot be ruled out. Although receptor and G protein subunits do not appear to be required for RGS binding to anionic vesicles, they may conspire to help localize and orient RGS4 (and other RGS) to optimize their GAP activities toward Gα (Tu et al., 2001) (Fig. 2, panel 2).
3. Once RGS4 Is Bound to Membranes and Functionally Linked to Receptor and G Protein, What Factors Regulate Its Effects on Ca2+ Signaling?
RGS4 binds very selectively to the anionic lipid PIP3 (Popov et al., 2000). PIP3is formed transiently from PIP2 by the actions of phosphatidylinositol 3-kinase (PI3K), certain isoforms of which are directly stimulated by Gβγ subunits (Fig. 2, panel 3) (Stephens et al., 1997). PIP3 binds RGS4 at a site within the RGS domain (distinct from and opposite to the RGS/Gα contact face), and PIP3 binding inhibits RGS4 GAP activity toward Gα (Popov et al., 2000). When complexed with Ca2+, activated calmodulin (Ca2+/CaM) apparently binds to this same site. Although Ca2+/CaM competes with PIP3 for binding at this site, it has no effect on Gαq GAP activity (Popov et al., 2000) (Fig. 2, panel 4).
Based on this information, a model has been proposed whereby RGS4 is recruited to specific Gq/11-linked receptors to modulate the frequency of Ca2+ oscillations elicited by those receptors (Luo et al., 2001). Following the initial burst of Ca2+ signaling, RGS4 is recruited to membranes to form a stable complex with specific Gq/11-linked GPCRs and blocks IP3/Ca2+ signaling. RGS4 could be precomplexed with the receptor and G protein prior to the signaling event, or it could be recruited after the fact, although the currently available evidence is too limited to differentiate between these scenarios. PIP3, formed in parallel by Gβγ-mediated activation of PI3K, subsequently sequesters RGS4 and blocks its GAP activity toward Gαq/11. This could provide a feedback loop to relieve RGS inhibition of Ca2+ signaling. A rise in intracellular Ca2+ due to resumed IP3production then activates CaM, which competes with PIP3 binding to RGS4 at the membrane (Fig. 2, panel 4), and the newly formed RGS4/Ca2+/CaM complex is capable of serving as a GAP for Gαq-GTP to block IP3/Ca2+ signaling (Fig. 2, panel 4). As cellular levels of Ca2+ fall, CaM becomes deactivated and dissociates from RGS4 thus allowing a new round of RGS4/PIP3 interactions. By alternatively binding PIP3 and Ca2+/CaM, RGS4 is capable of fine-tuning the frequency of Ca2+ oscillations (Fig. 2-5) (Luo et al., 2001). Evidence to support this model comes from two recent studies of Ca2+ signaling in intact cells. One study demonstrates that the frequency of Ca2+oscillations and fluctuations in cellular IP3levels in cells are superimposable (Nash et al., 2001), whereas the other study demonstrates that formation of Ca2+/CaM complexes in cardiac myocytes is required for RGS4 actions on G protein activation of muscarinic K+ channels (Ishii 2001).
The outlined scenario is plausible if receptor, Gαq, and PLCβ remain as a stable active complex in the presence of continued receptor agonist as has been proposed (Biddlecome et al., 1996). This model also can explain the observation that low physiological concentrations of agonist elicit Ca2+ oscillations whereas saturating concentrations of agonist elicit a sustained Ca2+signal (Thomas et al., 1996; Luo et al., 2001). Although direct roles for PI3K, calmodulin, and PIP3 in this scenario are extrapolated from in vitro studies (Popov et al., 2000) and remain untested in a cellular context, this model provides a plausible mechanism for RGS modulation of hormone- and receptor-directed Ca2+ oscillations, which can be readily tested in future studies. Limited information also suggests that other B/R4 family members may act similarly to modulate Ca2+signaling (Xu et al., 1999; Popov et al., 2000).
4. What Factors Contribute to Turning Off This Signaling Loop?
The simplest means for shutting off Ca2+oscillations would involve withdrawal of agonist. However, in the continued presence of agonist, several cellular mechanisms may contribute to turning off the RGS signal. At the level of the RGS protein, regulated post-translational modification of sites within the RGS domain may block further RGS/Gα interactions. Consistent with this idea, addition of palmitate to a conserved Cys residue on helix 4 of the RGS domain blocks RGS4 interactions with Gα (Tu et al., 1999). Alternatively, palmitoylation at other sites within the N terminus may target RGS4 to specialized “lipid rafts” within the plasma membrane and thereby limit its availability (Druey et al., 1999; Moffett et al., 2000). Other post-translational modifications may also contribute to feedback and inhibit RGS functions. Phosphorylation of several B/R4 family members modulates their capacity to interact with Gα. For example, phosphorylation of RGS2 by PKC (Cunningham et al., 2001) or RGS16 by undefined kinases (Chen et al., 2001a) blocks their interaction with Gα. Another possibility is that phosphorylation of certain B/R4 family members at sites within the RGS domain promotes their binding with the cytosolic scaffolding protein 14-3-3 to prevent their interactions with Gα (Benzing et al., 2000). Taken together, these findings suggest that mechanisms for uncoupling RGS from Gα could proceed in conjunction with well defined classical mechanisms that desensitize receptor and G protein actions such as receptor phosphorylation and internalization (Ferguson, 2001).
B. RGS4 As a Possible Scaffolding Protein That Links Receptors to Related Signaling Proteins
Experimental evidence supporting the model of RGS4 regulation of Ca2+ oscillations (Fig. 2) also suggests that RGS4 and certain other RGS proteins may bind directly to GPCRs to form a stable ternary complex between the GPCR, Gαq, PLCβ, and CaM (for discussion, see Sierra et al., 2000). Although no direct evidence has been reported demonstrating RGS4 physically binding to receptors, several lines of indirect evidence support the idea that RGS4 assembles related signaling proteins, perhaps as a stable complex with receptors. As discussed above, a synthetic peptide corresponding to the N terminus of RGS4 selectively blocks certain GPCR signals (Zeng et al., 1998; Xu et al., 1999). RGS4 also binds Ca2+/CaM at a regulatory site within the RGS domain (Popov et al., 2000), Gαq at the Gα/RGS interface (Hepler et al., 1997), and PLCβ1 at an undefined site (Dowal et al., 2001). RGS4 also displays relatively weak but significant affinity for binding Gβγ (Wang et al., 1998; Dowal et al., 2001). These data support the idea that RGS4 could potentially act as a multifunctional scaffold to assemble related proteins in a shared pathway in concert, specifically IP3-mediated Ca2+ signaling. In this regard, RGS4 and other RGS proteins may act in a manner similar to the β-arrestins, which form a stable signaling complex with GPCR and serve as a scaffold to assemble related kinases (JNK, Ask1, and MKK) to facilitate MAPK signaling at the plasma membrane (Miller and Lefkowitz, 2001). Further studies will be necessary to determine whether RGS4 and other simple RGS proteins make direct physical contact with GPCR and, if so, whether RGS and GPCRs form a stable complex. The relative role of G proteins in this process is currently unknown. Even so, other studies (discussed elsewhere in the text) provide compelling evidence that certain of the larger complex RGS proteins bind directly to GPCRs by PDZ domain interactions to serve as multifunctional integrators of receptor and G protein signaling.
C. RGS Modulation of the Kinetics of Fast-Acting Signaling Responses
Unlike inositol lipid/Ca2+ signaling and other slow-acting G protein-regulated processes (e.g., MAPK cascades), ion conductances in electrically excitable cells respond to signals on a subsecond time scale. GPCR and linked G protein subunits directly regulate several important fast-acting signaling events in the brain, retina, and heart (Arshavsky and Pugh, 1998; for review, see Zerangue and Jan, 1998). Most notable among these are phototransduction in the retina (mediated by Gαt), G protein-regulated inwardly rectifying potassium channels (GIRK) in brain and heart, and the voltage-dependent N-type Ca2+ channels in the brain (both channels are directly mediated by Gβγ). The onset and deactivation of these signaling events are very rapid, and compelling evidence now demonstrates that RGS proteins modulate the kinetics of these responses. In atrial myocytes, GIRK currents deactivate within 600 ms. However, when GIRKs are exogenously expressed inXenopus oocytes lacking RGS proteins, their deactivation occurs at rates markedly slower than the intrinsic GTPase rates of Gαi/o (Higashijima et al., 1987). Introduction of RGS4 and other simple RGS proteins of the B/R4 family (RGS1, RGS3, and RGS8) markedly accelerates GIRK activation and deactivation rates, and RGS4 restores GIRK kinetics in mammalian cell lines to levels similar to those observed in heart and brain (Doupnik et al., 1997;Saitoh et al., 1997, 1999). The RGS in question had no effect on the amplitude of GIRK currents, indicating that they are not dedicated inhibitors, but rather modulators that fine-tune these signal responses.
Mechanisms underlying RGS modulation of activation and deactivation rates for GIRK are unclear, although several possibilities have been proposed (Zerangue and Jan, 1998). Since GIRK and N-type Ca2+ channels are directly regulated by Gβγ and not Gα-GTP, rates of channel activation and deactivation presumably reflect availability of free Gβγ. RGS may stabilize an active GPCR/G protein/channel complex to limit the diffusion time required for activation and deactivation. To affect deactivation rates, RGS proteins may modulate the lifetime of free Gβγ. RGS and Gβγ compete for the same face of Gα, and binding to Gα is predicted to be mutually exclusive (Tesmer et al., 1997). Unstable and transient RGS/Gα-GDP interactions following GTP hydrolysis would promote Gα-GDP and Gβγ reassociation and increase deactivation rates. In support of this idea, RGS proteins block slow-acting Gβγ-mediated responses such as MAPK signaling in mammalian cells (Yan et al., 1997) and pheromone responses in yeast (Druey et al., 1996). Alternatively, formation of a stable RGS/Gα-GDP complex would prolong Gβγ availability thereby slowing deactivation rates and possibly enhancing the amplitude of response. Consistent with this idea, coexpression of the B/R4 family members RGS4 or RGS3 with GIRK markedly enhances basal current in a Gβγ-dependent manner (Bunemann and Hosey, 1998). Both processes may be at work in a cell-dependent context, depending on the signaling responses and proteins involved. Other studies clarify this point further by demonstrating that low levels of expressed RGS proteins modulate the rate of GIRK channel deactivation whereas higher levels of protein enhance the current amplitude (Keren-Raifman et al., 2001).
RGS proteins also modulate the kinetics of Ca2+currents carried by voltage-dependent N-type Ca2+channels. Stimulation of Gαi/o-linked GPCR by various neurotransmitters inhibits N-type Ca2+channels in neurons, and this inhibition is mediated by free Gβγ. In mammalian cells, overexpression of B/R4 RGS family members RGS3, RGS4, or RGS8 accelerates the rate of recovery of Ca2+ currents from neurotransmitter (Gβγ-mediated) inhibition, as well as decreasing the potency of agonist required (Jeong and Ikeda, 1998; Melliti et al., 1999). Consistent with these findings, expression of a mutant form of GαoA that is insensitive to endogenous RGS (DiBello et al., 1998; Lan et al., 1998) resulted in an increase in agonist potency, a marked reduction in recovery time, and an increase in the time to reach steady state after application of agonist (Jeong and Ikeda, 2000). Increased agonist potency and slow recovery times are consistent with RGS-mediated GAP effects on Gα and hence availability of free Gβγ. Reasons for RGS effects on the delayed rate to steady state are unclear, but could be explained if RGS promoted formation of a stable GPCR/G protein/channel complex that limited the diffusion time required for Gβγ (for further discussion, see Jeong and Ikeda, 2000).
RGS proteins also modulate the time course of phototransduction (Arshavsky and Pugh, 1998; He et al., 1998). In rod outer segments, elevated levels of cytosolic cGMP bind to and open channels to maintain resting membrane potential. In response to light, the photon-activated GPCR rhodopsin stimulates GTP binding to Gαt, which in turn, binds to the inhibitory γ-subunit of cGMP-phosphodiesterase (γ-PDE). Gαt-GTP sequesters γ-PDE and disinhibits the catalytic α-subunit of PDE (α-PDE), which becomes free to rapidly hydrolyze cGMP. In turn, a reduction in cytosolic cGMP levels causes channels to close and initiates electrical signaling pulses to the visual cortex. Remarkably, this entire signaling cascade proceeds within 150 ms with a recovery time of 200 ms while providing single photon reliability and maximal signal amplification. Intrinsic rates of rhodopsin-stimulated Gαt GTPase activity are 100-fold too slow to account for the onset and deactivation of this signaling event, predicting the existence of a regulatory factor. It was recognized that an essential unidentified cellular factor, in concerted action with γ-PDE, served as a GAP for Gαt to limit the lifetime of the signaling response (Angleson and Wensel, 1994;Arshavsky et al., 1994). Wensel and coworkers identified a membrane bound protein that is a GAP for Gαt (Angleson and Wensel, 1993), and later showed that this protein was the RGS protein, RGS9 (He et al., 1998). In these studies, RGS9-1 was found to meet all of the requirements of the essential factor, i.e., it is expressed exclusively in rod outer segment, is tightly membrane-bound, and is observed to act synergistically with γ-PDE as a potent GAP for Gαt. Thus, RGS9-1 is capable of determining the lifetime of active Gαt-GTP/γ-PDE complex. To achieve this, RGS9-1 forms a high-affinity ternary complex with active Gαt and γ-PDE (Slep et al., 2001) thereby eliminating the slow binding constants typical of protein-protein diffusion. Confirmation of the importance of RGS9-1 as the key modulator of the recovery step in phototransduction comes from recent studies of mouse retinas derived from homozygous RGS9(−/−) knockouts (Chen et al., 2000; Lyubarsky et al., 2001). Unlike RGS9, other RGS proteins (RGS4 and RGS-GAIP) fail to act cooperatively with γ-PDE as GAPs for Gαt, even though each is an effective GAP for Gαt in isolation (Nekrasova et al., 1997). In fact, these proteins inhibit rather than enhance γ-PDE-directed GAP activity toward Gαt, suggesting a high degree of specificity for the proteins involved in this signaling response.
IV. RGS Proteins Integrate G Protein Signals
We have discussed examples of RGS proteins as modulators of both slow- and fast-acting G protein signals. Evidence suggests that the simple RGS proteins (members of the B/R4 and perhaps the A/RZ subfamilies) serve as highly regulated modulators of G protein signaling responses rather than as dedicated inhibitors. However, the larger more complex RGS proteins (members of the C/R7, D/R12, E/RA, and F/RL subfamilies) in many cases likely perform additional cellular functions. For example, although RGS9 is clearly an essential modulator of Gαt-directed phototransduction, it differs from the simple RGS proteins in that it is larger and exists as multiple splice variants (RGS9-1 and RGS9-2). Both forms of RGS9 bind the Gβ5 subunit and the longer variant, RGS9-2, may also bind other signaling proteins (Chen et al., 2001b) at its C-terminal extension (Figs. 1 and 4; Table 1). The extended N and C termini confer additional regulatory and/or signaling functions to RGS9 and other complex RGS proteins. Emerging concepts suggest that these complex RGS proteins link active Gα subunits to other signaling pathways to serve as multifunctional integrators of G protein signaling.
A. RGS Proteins Integrate Distinct G Protein Signaling Pathways
Activation of multiple receptors in a single cell initiates complex signaling cascades that must be integrated for proper cellular responses. This integration can occur through the activation of kinases, recruitment of cellular scaffolds and associated signaling proteins, or by direct receptor interactions. Recent evidence indicates that some cells use RGS proteins to link distinct receptor and Gα activation to parallel downstream signaling cascades.
Members of the C/R7 family (RGS6, 7, 9, and 11) could represent important contributors to signal integration from multiple receptors. In addition to their RGS domain, C/R7 subfamily proteins contain DEP, R7H, and GGL domains (Fig. 1). GGL domains bind the Gβ subunit Gβ5 specifically and with high affinity (Snow et al., 1999). Gβ5 is unique among Gβ subunits in that it has reduced sequence homology with other family members (53%), is expressed almost exclusively in the nervous system, and does not bind well to most Gγ subunits (Watson et al., 1994). In studies designed to identify binding partners for Gβ5, RGS7 was the main binding partner to copurify out of retinal extracts (Cabrera et al., 1998). Further studies showed that the other members of the C/R7 family also bind Gβ5 with high affinity, both in vitro and in vivo (Snow et al., 1998b; Posner et al., 1999; Zhang and Simonds, 2000). This interaction, taken together with evidence from genetic studies in lower eukaryotes (described below), has led to working models that propose that RGS/Gβ5 complexes can potentially substitute for Gβγ in Gαβγ heterotrimers, although no direct evidence for this has been reported.
Genetic evidence from C. elegans supports this model, showing that C/R7 subfamily members couple competing G protein-regulated behaviors (Hajdu-Cronin et al., 1999; Chase et al., 2001). In worms, the interplay between Gαq- and Gαo-linked signals controls egg laying and locomotion. Proposed models suggest that serotonin (Gαo-linked), and acetylcholine (Gαq-linked), cross-regulate these behaviors (Hajdu-Cronin et al., 1999). Activation of Gαocauses lethargic movements, delayed egg laying, and reduced mating, whereas activation of Gαq has the opposite effect. Egl-10 and Eat-16, C. elegans C/R7-like proteins which bind the Gβ5 homolog GPB-2, cross-regulate these signals. Loss-of-function mutations in Eat-16 suppress the constitutively active Gαo phenotype, indicating that Eat-16 acts downstream of Gαo. However, reducing the levels of Gαq reverses the Eat-16 loss-of-function phenotype, indicating that Eat-16 acts downstream of Gαo by limiting Gαqactivity (Chase et al., 2001).
The genetic evidence can be interpreted in a number of ways. In the simplest model, the RGS/GPB-2 dimers act only as negative regulators of their respective Gα homologs (van der Linden et al., 2001). However, this interpretation does not fully take into account the interplay between the different Gα signaling pathways. A more comprehensive model is derived from scenarios proposed by several research groups (Guan and Han, 1999; Hajdu-Cronin et al., 1999; Sierra et al., 2000) (depicted in Fig. 3A). In this model, Eat-16, GPB-2, and Gαo-GDP exist as a heterotrimer at rest whereas Egl-10 and GPB-2 complex with Gαq-GDP. In this case, Eat-16 acts as a Gγ subunit for Gαo whereas Egl-10 is a Gγ for Gαq. Neurotransmitter activation of either receptor releases the RGS protein. When the Eat-16/GPB-2 complex disengages from Gαo-GTP, Eat-16 is free to act as a GAP on Gαq via its RGS domain. In parallel, activation of Gαq by competing neurotransmitters releases Egl-10/GPB-2 from Gαq-GTP and allows Egl-10 to limit Gαo signals. While the physiological purpose of this cross-talk is uncertain, reciprocal inhibition among neurotransmitters allows the worms a tighter level of control over reproduction and locomotion (van der Linden et al., 2001). Many aspects of this model remain to be tested. For example, there is no direct evidence that the RGS-Gβ5 dimers bind the inactive Gα homologs. Even given the need for further testing, this scenario provides a testable model to explain the physiological role of RGS-Gβ5 interactions.
RGS proteins as integrators of G protein signaling. Panel A, speculative model for RGS-directed integration of G protein signaling in the worm C. elegans. The worm RGS proteins Egl-10 and Eat-16 each form a heterotrimeric complex with the C. elegans equivalent of Gβ5 (GPB-2) and Gα subunits to couple to GPCR (left panel). Following receptor activation by either acetylcholine (Ach) (middle panel) or serotonin (5-HT) (right panel), the corresponding G protein signaling pathway is stimulated and the linked RGS/Gβ5-like complex is released to inhibit the opposing G protein signaling pathway. Gαq activation of the Egl-8 (PLCβ) and formation of diacylglycerol stimulates locomotion and egg laying, whereas Gαo activation of GDK-1 (diacylglycerol kinase) opposes this behavior. See text for further details. Panel B, a hypothetical model for RGS-directed integration of G protein signaling in the mammalian central nervous system. Left, an R7 subfamily protein (containing DEP, R7H, GGL, and RGS domains) exists as a complex with Gβ5 (RGS/Gβ5) in the cytosol and at the membrane. RGS/Gβ5 forms a heterotrimeric complex with Gαq and couples to a GPCR (R1). Right, activation of receptor (R1) with neurotransmitter (NT1) stimulates Gαq signaling and releases RGS/Gβ5, which is available to block subsequent signaling initiated by a second neurotransmitter (NT2) that activates a GPCR (R2) linked to Go. See text for a more detailed description of the model and appropriate references. Panel C, PDZ-RGS3 mediates reverse signaling by Ephrin-B. In response to EphB receptor binding to EphB on granule cells, the PDZ domain of PDZ-RGS3 binds the C-terminal tail of Ephrin-B at the plasma membrane. This positions PDZ-RGS3 to block G protein-stimulated granule cell migration initiated by SDF-1 (guidance factor) binding to the CXCR4 GPCR. See text for further details about the model and appropriate references. Panel D, the RGS-like protein p115RhoGEF mediates Gα13 stimulation of RhoA. Following activation of a Gα13-linked GPCR by hormone or neurotransmitter (H/NT), p115RhoGEF (containing RGS, DH, and PH domains) is recruited from the cytosol and associates with Gα13 at the plasma membrane. The DH domain of p115RhoGEF recruits and stimulates GTP binding to RhoA, which activates Rho kinase and downstream changes in cell morphology, adhesion, and motility. The GAP activity of the RGS domain feeds back to shut-off Gα13 signaling. See text for a more detailed description of the model and appropriate references.
Similar pathways may exist in mammalian cells, although in vitro studies using purified mammalian RGS proteins have failed to show association of Gα-GDP/RGS/Gβ5 heterotrimers, and RGS/Gβ5 dimers do not mimic conventional Gβγ signals such as modulation of adenylyl cyclase or activation of PLCβ (Posner et al., 1999). However, based on recent unpublished studies involving RGS9/Gβ5 dimers and their involvement in M2AchR signaling, Sondek and Siderovski (2001) have proposed a model in which members of the C/R7 subfamily, complexed with Gβ5, act as a Gβγ. A variation of this model is diagramed in Fig. 3B. If RGS/Gβ5 dimers can substitute for Gβγ, several questions remain to be addressed. These include how Gα subunits discriminate between conventional Gβγ subunits and RGS/Gβ5 complexes in cells, how RGS/Gβ5 target specific Gα subunits, and what role RGS GAP activity plays in their signaling functions. Unlike other Gβγ dimers, RGS/Gβ5 complexes, particularly RGS7/Gβ5, are both membrane bound and cytosolic (Cabrera et al., 1998; Rose et al., 2000). Signaling roles for the cytosolic subpopulation are unclear but may relate to interactions with cytosolic-binding proteins such as 14-3-3 (discussed below).
In addition to the RGS and GGL domains, all C/R7 family members also contain several other conserved regions that may influence their function, including the DEP domain. DEP domains exist in many unrelated signaling proteins and may influence membrane interactions or possibly influence RGS/Gα interactions. C/R7 family members also share a conserved region of residues identified on the PfamB data base (Fig. 1) (Sondek and Siderovski, 2001), which we term the R7H domain (Fig. 1). Although roles for this domain are unknown, one possibility is that it may be involved in regulated membrane attachment. This attachment could occur through post-translational lipid modifications, which, in at least one C/R7 family member, RGS7, localize to this region (Rose et al., 2000). This conserved region may also have independent signaling functions through interactions with as yet unidentified binding partners.
Models proposing that C/R7 family members act as both a Gβγ and a GAP can help account for signal integration at multiple levels. Such a mechanism could help focus signals from multiple receptors activated by a single ligand, integrate signals from competing ligands, or auto-inhibit a Gα for which the RGS acts as both a Gγ and a GAP. Because all C/R7 family members, as well as Gβ5, are found exclusively in brain, fine-tuning of neuronal transmission is a hypothetical function of these complexes. For example in the striatum, where the C/R7 family member RGS9-2 is highly enriched, this protein could help fine-tune glutamate signals and present a therapeutic target to help regulate striatal activity in diseases such as Parkinson's. In the striatum, glutamate is released onto cells expressing multiple subtypes of metabotropic receptors including Gαq-linked group I mGluR (mGluRI), as well as Gαi/o-linked group II mGluR (mGluRII). In a hypothetical model (Fig. 3B), the RGS9-2/Gβ5 complex associates with the mGluRI subpopulation of postsynaptic receptors, acting as a Gβγ subunit. When glutamate is released, both mGluRI and mGluRII receptors would be activated and mGluRI-linked Gαq would release RGS9-2/Gβ5. The RGS domain of RGS9-2 would then be free to act as a GAP for the Gαi/o subunits activated by mGluRII receptors. Through this interaction, RGS9-2 expression could limit ion channel modulation through Gi/o-linked receptors while enhancing Ca2+ and PKC signals from Gαq.
Additional binding partners may modulate RGS/Gβ5 function. For example, RGS9-2 interaction with the protein evectin could affect its localization or interactions (Chen et al., 2001b). Although little is known about the signaling properties of evectins, they are membrane-anchored proteins with an N-terminal PH domain and may therefore secure RGS9-2 to the membrane. Other C/R7 family members also recruit a variety of additional binding partners including γ-PDE, polycystin, and 14-3-3, as discussed elsewhere in the text. Interactions between C/R7 family members and these proteins could recruit the RGS to membrane compartments, direct interactions with Gα or Gβ5, or alter RGS/Gβ5 activity, and thereby regulate the novel signaling functions of this RGS subfamily.
B. RGS Proteins Integrate G Protein and Non-G Protein-Linked Signals
A compelling example of RGS proteins directly linking different receptor systems is found in mouse cerebellar granule cells (Lu et al., 2001). Ephrin B (EphB), a single transmembrane-spanning cell surface ligand for a tyrosine kinase receptor, is implicated in a variety of developmental processes. Although membrane-spanning ligands were traditionally thought to signal exclusively through their receptors, evidence now indicates that many also relay signals through their own C termini. In the case of EphB, this “reverse signaling” mediates several developmental processes including axon guidance and vascularization and allows proper migration of granule cells in the developing cerebellum. The EphB tail binds directly to the PDZ domain of PDZ-RGS3, a potential new splice variant of RGS3 identified by yeast two-hybrid screening using the cytoplasmic tail of EphB as bait. In migration assays, EphB inhibits chemoattraction by the chemokine receptor CXCR4, a G protein-coupled receptor. PDZ-RGS3 mediates this inhibition, which requires both the PDZ and RGS domains of the protein. The proposed model for EphB reverse signaling is depicted in Fig. 3C (Lu et al., 2001). At birth, granule cells are retained at the pia by chemoattraction through activation of CXCR4. At approximately postnatal day 3, EphB is up-regulated and interacts with its receptor, which promotes binding of PDZ-RGS3 to the cytoplasmic tail of EphB. With the PDZ domain recruited to EphB, the RGS domain is free to inhibit CXCR4 signals and allow cells to begin migrating through the cerebellum. A caveat to this model is that EphB and CXCR4 must be in close proximity, since they directly link through PDZ-RGS3. In this model, PDZ-RGS3 is an EphB effector, directly integrating the tyrosine kinase ligand with a GPCR signaling cascade.
C. RGS Proteins Link Gα to Monomeric GTPases
One of the most exciting areas of research in the RGS field is in the newfound appreciation that RGS proteins can directly link Gα to nontraditional signaling cascades, particularly to regulation of monomeric GTPases. The first example of this was the RhoA exchange factor p115RhoGEF (Hart et al., 1998; Kozasa et al., 1998). The structure of RhoGEFs consists of an N-terminal RGS domain, a more C-terminal Rho guanine nucleotide exchange factor (DH) domain, and a PH domain (Fig. 1). In reconstituted systems using purified proteins, the RGS domain specifically interacts with and acts as a GAP for Gα12/13 family members whereas the DH domain exchanges GTP for GDP on RhoA. A model illustrating Gα12/13 regulation of Rho signaling is pictured in Fig. 3D. At rest, the RGS and DH domains of the cytosolic RhoGEF inhibit one another. After receptor activation, Gα13-GTP recruits RhoGEF through interaction with the RGS domain, releasing the DH domain that then binds RhoA, draws it to the membrane and initiates nucleotide exchange. These proteins therefore directly link GPCR activation to the cytoskeletal changes initiated by activated RhoA. The discovery of the p115RhoGEF family and the elucidation of their mechanism of action accounts for some of the morphological and proliferative changes induced by hormones such as thrombin or endothelin and provides new models to explain Gα12/13-induced oncogenesis (Fukuhara et al., 2001; Kozasa, 2001).
The resting state of p115RhoGEF in which the two catalytic domains inhibit each other points to a potentially widespread mechanism of RGS regulation. The simplest models presume that RGS domains are always active and that recruitment to the membrane by activated Gα subunits is the only necessary step to promote RGS GAP activity. Clearly, this is not the case for p115RhoGEF, since its in vitro GAP activity is limited when the DH and RGS domains are coexpressed (Kozasa et al., 1998). In the case of PDZ-RGS3, membrane targeting alone is also not sufficient to elicit signals since adding a membrane anchor does not mimic EphB reverse signaling in oocytes (Lu et al., 2001). In this model, EphB not only localizes PDZ-RGS3 but also frees the RGS domain, allowing it to act as a GAP at the CXCR4-linked Gα. Stimulation of GAP activity through interaction with binding partners may be a common mechanism of RGS regulation that is not yet fully appreciated.
RGS12 and RGS14 may also directly link heterotrimeric and monomeric GTPases. The cellular roles of this RGS subfamily are not as well understood as those of the RhoGEFs, but investigations promise to lead to exciting new insights into nontraditional GPCR signals. This is especially true in light of recent evidence that RGS12 can regulate multiple receptor signals and that both RGS12 and RGS14 are themselves highly regulated (Mao et al., 1998; Snow et al., 1998a; Chatterjee and Fisher, 2000b; Schiff et al., 2000; Kimple et al., 2001). Similar to RhoGEFs, RGS12 and RGS14 share an N-terminal RGS domain and a binding domain for the monomeric GTPases Rap1 and Rap2 toward the C terminus. In addition, these proteins share a C-terminal G protein regulatory or “GoLoco” motif (Fig. 1). The RGS domains of RGS12 and RGS14 are specific GAPs for Gαi/o family members in vitro (Snow et al., 1998a; Cho et al., 2000; Traver et al., 2000; Hollinger et al., 2001) although both proteins can inhibit Gα12/13-mediated signals in cell systems (Mao et al., 1998; Cho et al., 2000). The RBD or Rap interacting domain interacts specifically with the GTP bound form of Rap1 and Rap2 (Traver et al., 2000), members of the Ras family of GTPases. Apparently, this binding does not increase GTP binding or hydrolysis on Rap1 (Traver et al., 2000; Hollinger et al., 2001). Although the functional consequences of RGS/Rap interaction remain elusive, possibilities include that the binding of RGS12 or RGS14 to Rap may interfere with Rap-effector binding, or may recruit activated Rap to the membrane to initiate MAPK cascades. Knowing whether RGS12 or RGS14 coexpression enhances or limits Rap-mediated signals will help distinguish between these possibilities but has not yet been tested.
In addition to the RGS and RBD domains, RGS12 and RGS14 also contain a GoLoco motif toward their C terminus (Fig. 1). Recently, several groups showed that GoLoco domains bind inactive Gαi-GDP, but not Gαo-GDP, subunits. They potently inhibit guanine nucleotide exchange, permitting a protein to act as aguanine nucleotide dissociationinhibitor (GDI) (Hollinger et al., 2001; Kimple et al., 2001; Natochin et al., 2001). Because RGS12 and RGS14 are GAPs for Gαi/o family members, the presence of a GoLoco motif may allow specific inhibition of Gαisubunits. However, the consequences of interplay between Gα and other binding partners of RGS12 and RGS14 are not yet clear. Taken together, Gαi targeting by the GoLoco motif, the interaction with Rap1/2, and the interplay of G protein regulatory functions position RGS12 and RGS14 as meaningful players in investigating novel mechanisms of G protein regulation and signal transduction.
D. RGS Proteins As Scaffolds to Assemble Related Signaling Components
In some cases, RGS proteins act as platforms in signaling cascades, bringing together both Gα and non-G protein partners. As discussed previously in the case of RGS4, RGS binding partners can include components of traditional G protein signals such as the receptor Gα and Gβγ. They also can include proteins that may link Gα to novel downstream signaling cascades.
Perhaps the best example of an RGS protein as a scaffold is Axin. Axin is part of the Wnt signaling cascade, which directs vertebrate axis formation and neural development in embryos (Vasicek et al., 1997). Some models of learning and memory formation also implicate this cascade, and forms of Alzheimer's disease may involve disorders in this pathway. Axin regulates the function of the transcription factor β-catenin by coordinating its phosphorylation. It does this by bringing together adenomatous polyposis coli (APC), GSK3β, and β-catenin. When complexed with these proteins, β-catenin is highly phosphorylated by GSK3β, which initiates its degradation and reduces β-catenin-mediated gene transcription (Nakamura et al., 1998;Hedgepeth et al., 1999). APC binds at the RGS domain near the N terminus of Axin. However, based on the crystal structure of the RGS domain of Axin, APC binds at an extended groove distinct from the putative Gα interacting surface and, therefore, potentially independent of Gα binding (Spink et al., 2000). Therefore, although Gα interactions are not yet demonstrated, Axin could bind Gα while complexed with its other binding partners, creating an additional level of regulation of Wnt-mediated signals through Axin recruitment or activation.
The GRKs are a group of RGS proteins in the F/RL subfamily with multiple interacting partners. The primary identified role of GRKs is GPCR phosphorylation, which enhances arrestin binding and receptor desensitization (Ferguson, 2001). However, GRK family members also contain an RGS domain at their N terminus, within the putative receptor recognition site, and certain isoforms bind Gβγ subunits at a PH domain toward their C terminus (Siderovski et al., 1996). Expression of the isolated PH domain of GRK2 limits Gβγ signals, a feature often adapted as a tool to identify Gα- versus Gβγ-mediated signals in cells. Although the RGS domain of GRK2 is only a weak GAP for Gαq, it binds to both native and recombinant Gαq in an activation-dependent manner. GRK2 expression also effectively limits Gαq signals. This outcome most likely occurs by GRK2 sequestering Gαq or interfering with effector activation rather than through increased GTP hydrolysis (Carman et al., 1999;Sallese et al., 2000; Usui et al., 2000). Because GRK2 binds directly to receptors, Gαq and Gβγ, GRK-mediated scaffolding could target inhibition to specific receptor signals as well as sequester signaling components.
Interaction of RGS proteins, particularly RGS16, with MIR16 could influence the scaffolding properties of the cell membrane (Zheng et al., 2000). MIR16 is the first cloned glycerophosphodiester phosphodiesterase. Although the physiological role of this family of proteins in mammalian systems has not yet been investigated, in lower eukaryotes they regulate the phospholipid composition of cell membranes. This controls the existence of microdomains within the membrane known as lipid rafts in which protein complexes assemble for proper signaling. While interaction between MIR16 and RGS proteins may regulate RGS localization or function, particularly since MIR16 binds at the RGS domain, this interaction may actually regulate the enzymatic activity of MIR16 (Zheng et al., 2000). Regulating the enzymatic activity of MIR16 could impact cells by affecting signaling cascades that depend on components being properly oriented within lipid rafts.
RGS-GAIP may represent one component of a scaffolding complex linking nerve growth factor-mediated signals to GPCRs. At its C terminus, RGS-GAIP interacts with the PDZ domain of GIPC (RGS-GAIPinteracting protein C terminus) (De Vries et al., 1998b). A recent report indicated that GIPC links TrkA nerve growth factor receptors to RGS-GAIP, and consequently to Gαi/o-linked GPCRs (Lou et al., 2001). In PC12 cells, GIPC interacts with the TrkA receptor through its PDZ domain. Although RGS-GAIP binds in the same region of TrkA, the binding sites are on opposing faces of the molecule, and binding is therefore not mutually exclusive. All three molecules coprecipitate out of cell extracts, showing that they can indeed form a trimeric complex. The authors speculate that this interaction may indicate TrkA cross-talk with a GPCR. Supporting this model, nerve growth factor has been shown to elevate cAMP levels in neurons, which could result from Gαi/o inhibition through recruitment of RGS-GAIP to a TrkA/GPCR complex.
E. RGS Proteins Regulate Intracellular Trafficking
Regulated transport of proteins and intracellular membranes is one of the most vital mechanisms supporting signal transduction. These processes include regulated internalization, recycling of plasma membrane proteins, and the transport of proteins within the cytoplasm. Factors that affect intracellular transport can fundamentally alter the signaling properties of a cell.
RGS-PX1, a newly identified RGS protein, boasts several characteristics that position it as a potentially important link between G proteins and intracellular trafficking (Zheng et al., 2001). RGS-PX1 contains a central RGS domain, several domains likely involved in membrane recruitment, and a Phox domain, which is commonly found in sorting nexins that regulate trafficking between intracellular compartments. One of the most intriguing features of RGS-PX1 is that, unlike any other RGS protein identified to date, it specifically regulates Gαs and Gαs-linked receptor signals in vitro and in intact cells. However, in addition to its capacity to inhibit Gαs-linked signals, RGS-PX1 may also affect transport of internalized receptors. Consistent with a role as a type of sorting nexin, RGS-PX1 selectively binds to phospholipids that are enriched in endosomes, and it is found primarily in early endosomes when expressed in HEK293 cells. In these cells, RGS-PX1 inhibits transport of EGF receptor (EGFR) from endosomes to lysosomes, reduces EGFR degradation through lysosomes, and enhances EGFR signals. Taken together, these findings demonstrate that RGS-PX1 is both a Gαs-specific GAP and a functional sorting nexin within these cells. With its dual capacity, RGS-PX1 provides new insights into the link between signaling and trafficking inside the cell (von Zastrow and Mostov, 2001).
At least one well studied RGS protein, RGS-GAIP, is also clearly involved in traffic of both intracellular and internalized vesicles (De Vries et al., 1998a,b; Wylie et al., 1999). Both native and recombinant RGS-GAIP is found on clathrin-coated vesicles. These vesicles usually are not directly associated with the plasma membrane, although some are partially derived from plasma membrane-associated clathrin-coated pits. RGS-GAIP is also found on trans-Golgi derived vesicles both in native fractions (De Vries et al., 1998a) and when expressed as recombinant protein (Wylie et al., 1999). Interestingly, RGS-GAIP and its preferred Gα substrate, Gαi3, are found primarily on separate intracellular membranes. Overexpression of either RGS-GAIP or Gαi3 inhibits vesicle transport at the trans-Golgi network. One model predicts that RGS-GAIP and Gαi3 interact in a time-dependent manner such that vesicles expressing RGS-GAIP interact with the Golgi stacks expressing Gαi3, but only when Gαi3 is activated. Hydrolysis of GTP, enhanced by RGS-GAIP, releases the RGS-GAIP-expressing vesicles, and this alternating cycle moves cargo through the trans-Golgi. Confounding this interpretation, RGS-GAIP overexpressionmimics overexpression of Gαi3 rather than inhibiting it. This data supports a model in which the primary function of RGS-GAIP in the cell may be independent of its GAP activity.
The interaction between RGS-GAIP and the PDZ domain of GIPC may also play a role in regulation of intracellular traffic. GIPC (also known as NIP and SemCAP1) is implicated at multiple levels in axon guidance, although this may be through GIPC interaction with semaphorins and neuropilins rather than with RGS-GAIP (Cai and Reed, 1999; Wang et al., 1999). Interactions of RGS-GAIP with GIPC may account for the inhibition of vesicular traffic seen with overexpressed RGS-GAIP, which mirrors overexpression of Gαi3rather than reversing it (see above). Because GIPC overexpression causes a redistribution of a semaphorin to lipid rafts, GIPC may also help target RGS-GAIP intracellularly.
B/R4 family members may also be involved in vesicular transport. A recent study (Sullivan et al., 2000) noted that in neuronal cell lines, the membrane-associated fraction of RGS4 is found primarily in the cytoplasm rather than at the plasma membrane. This distribution occurs when a protein is localized to intracellular membranes such as Golgi-derived vesicles. In a yeast two-hybrid screen, β-COP, an integral part of the COP-1 complex found on Golgi-derived vesicles was identified as an RGS4 interacting partner (Sullivan et al., 2000). In cell lines, a large percentage of native RGS4 (and RGS2) associates with the large COP-1 complex. β-COP and the COP-1 complex are implicated in anterograde transport of vesicles from the endoplasmic reticulum to the Golgi. Although the interaction of RGS4 with β-COP occurs at the RGS domain, it does not reduce RGS4 enhancement of Gαi GTPase activity. In contrast, RGS4 binding to β-COP does limit COP-1 association with Golgi membranes and RGS4 expression inhibits intracellular transport from the endoplasmic reticulum. Taken together, these findings suggest that RGS4 (and RGS2) can control availability of plasma membrane and secretory proteins through interaction with β-COP.
F. RGS Protein Interactions with Non-G Protein Binding Partners
As the list of non-G protein binding partners for RGS proteins expands, research into the functional significance of these interactions intensifies (Fig. 4). Many occur independently of G protein binding, at domains distinct from the RGS box. For the interactions that do occur at the RGS domain, it is often unclear how they impact Gα binding. In the following section, we will describe several of these newfound binding partners and discuss their potential signaling functions.
Variant forms of RGS12 interact with a number of binding partners and therefore have the capacity to differentially regulate a subset of signals. The longest splice variant of RGS12 contains an N-terminal PDZ domain and a C-terminal PDZ binding motif. PDZ domains, found in a variety of signaling proteins, are believed to act as scaffolds by binding recognition motifs of 4 amino acids at C termini of numerous proteins, including a number of GPCRs (Sheng and Sala, 2001). An extensive survey of binding to receptor tails showed that the PDZ domain of RGS12 interacts specifically with the interleukin-8 receptor (CXCR2), as well as with its own C-terminal PDZ binding motif (Snow et al., 1998a). A proposed model exists in which RGS12 GAP activity is auto-inhibited at rest through the interaction between the N-terminal PDZ domain and the C-terminal binding motif. Recruitment of the PDZ domain to the IL-8 receptor eliminates this inhibition and allows RGS12 to act as a GAP for Gαi/o subunits. However, since some splice variants of RGS12 contain only the PDZ domain (see below), RGS12 could also bring together the receptor with downstream signaling partners such as Gαi/o or Rap.
RGS12 may also bind together other partners in signaling complexes. In cultured chick dorsal root ganglia, Gαi/o-linked GABABreceptors inhibit N-type Ca2+ channels, which causes reduced neurotransmitter release. One report (Schiff et al., 2000) showed that recombinant RGS12 reduces the half-life of the Ca2+ current inhibition. They also show that eliminating native RGS12 dramatically extends the GABAB-mediated signal. These results demonstrate that native RGS12 is responsible for the rapid termination of GABAB-mediated Ca2+ channel inhibition. Because RGS12 is a GAP for Gαi/oand should therefore limit GABAB-activated Gαi/o signals, these findings were not altogether unexpected. However, further study showed that the termination of this signal is not mediated through the RGS domain, but that the phosphotyrosine-binding domain of RGS12 specifically binds the phosphorylated α1Bsubunit of the Ca2+ channel. Why this interaction limits inhibition of the channel is unclear. One hypothesis is the Ca2+ channel subunit recruits RGS12, tethering the channel to Gαo, which is subsequently inhibited by RGS12 (Schiff et al., 2000). Because RGS12 contains a number of other domains, including a GoLoco motif, models of these connections are complex and still incomplete.
Interaction with an important cytosolic scaffolding protein, 14-3-3, could help regulate RGS functions. 14-3-3 binds at phosphorylated residues in a semiconserved consensus sequence reportedly present in a number of RGS proteins (RGS1, 2, 3, 7, 8, 11, 13, 16). Both RGS3 and RGS7 interact with 14-3-3 in a phosphorylation-dependent manner (Benzing et al., 2000). In the case of RGS7, 14-3-3 binding actually interferes with Gαi/o interactions and may therefore limit signals. However, proteins such as 14-3-3 are scaffolds for known signaling cascades such as the ras-raf-MAPK cascade (Fu et al., 2000), without which activation of downstream binding partners does not occur. 14-3-3 interactions with RGS proteins may therefore not only regulate Gα interaction, but also represent part of an intracellular signal cascade that is not yet fully elucidated. In addition to the cytosolic 14-3-3, RGS7 also binds polycystin, a transmembrane protein involved in cystic kidney disease (Kim et al., 1999). As discussed elsewhere in the text, this interaction may regulate RGS7 degradation.
Simple RGS proteins also exhibit surprising interactions with non-Gα binding partners. For example, the B/R4 subfamily member RGS2 blocks cAMP production by adenylyl cyclase in olfactory cells (Sinnarajah et al., 2001). Since RGS2 is a selective GAP for Gαq and, at that time, no RGS protein was known to inhibit Gαs, these findings were unexpected. Inhibition of cAMP production is mediated by a direct interaction of RGS2 with certain subtypes of adenylyl cyclase shown using purified recombinant RGS2 and the purified cytosolic domain of type V adenylyl cyclase. The inhibitory effect of RGS2 on adenylyl cyclase is found both in recombinant expression systems and in olfactory cells, suggesting that native levels of RGS2 determine the response properties of odorant receptors (which signal through Gαs). D-AKAP2 may also be an RGS protein involved in the cAMP signaling cascade. A yeast two-hybrid screen identified D-AKAP2 as a binding partner for the regulatory subunits of protein kinase A (PKA) (Huang et al., 1997b). The PKA binding region is located in the C-terminal “R”-binding domain but D-AKAP also contains a putative RGS-like domain at the N terminus. Although the RGS domain of D-AKAP has never been shown to interact with or act as a GAP on any Gα subunit, it is tempting to speculate that D-AKAP2 may link modulation of cAMP (through Gαi or possibly Gαs) with activation of PKA.
In the preceding sections, we have discussed the roles that both simple and complex RGS proteins play in cell physiology. It is now apparent that most RGS proteins have more signaling functions than suggested by their GAP activity. Simple RGS proteins modulate traditional GPCR signals, influencing the response elicited by hormones and neurotransmitters through targeted receptor interactions. Both simple and complex RGS proteins play a key role as scaffolds for receptors and components of the G protein signal cascade. Additionally, many RGS proteins integrate divergent signals, sometimes interacting with their binding partners in unexpected ways. RGS proteins govern ion channel regulation, intracellular traffic, and cell morphology by regulating both G protein and non-G protein-linked signals. Because of their extensive involvement in so many aspects of signal transduction, cells have evolved multiple mechanisms to tightly regulate RGS functions.
V. Cellular Mechanisms Regulating RGS Protein Functions
As our understanding of the biochemistry and cellular roles of RGS proteins has come into focus, attention has turned to studying the cellular mechanisms regulating them. Many studies in living cells demonstrate RGS functions not predicted by in vitro assays. For example, RGS proteins often do not exhibit the same Gα selectivity in culture as in recombinant assays, their localization is influenced by receptor activation, and their expression levels are under tight control in the cellular environment. The activity of kinases, protein binding partners, and proteases can affect RGS action. At present, our limited understanding of how cells regulate RGS proteins makes for a somewhat confusing field of study. However, understanding how the complex regulation of RGS proteins impacts cell signals will ultimately contribute to the development of an array of highly targeted therapeutics.
A. Regulation of RGS Signaling Capacity by Feedback Phosphorylation
Although regulation can occur at multiple levels, post-translational modifications provide the most direct mechanism to alter protein interactions or localization. One of the most prevalent and important modifications involves the addition or removal of phosphate. Through phosphorylation, signaling cascades can be fine-tuned in either a feedback or feed-forward manner.
Since signals that RGS proteins manipulate activate several kinases, models in which feedback phosphorylation is a primary mechanism regulating RGS function are attractive. Of the growing number of examples in the literature (Table 2), the primary effect of phosphate addition is to alter protein-protein contact either by promoting interactions through binding motifs specific for phosphorylated amino acids, or by sterically interfering with interactions. Phosphorylation can either enhance or inhibit GAP activity, based on the RGS and kinase involved. Protein kinase C (PKC), for instance, phosphorylates RGS2 and reduces its Gα GAP activity, the net effect of which is augmentation of Gq/11 signals (Cunningham et al., 2001). A very recent report reveals that RGS16 is phosphorylated at Ser53 and Ser194 in cells after stimulation of Gαi/olinked α2A-adrenergic receptors (Chen et al., 2001a). Phosphorylation of Ser53, at the N terminus of the RGS domain, reduces RGS16 GAP activity and enhances α2A-adrenergic signals including ERK activation. Although the exact kinase involved is unknown, this phosphorylation represents a positive feedback loop to enhance receptor signals. On the other hand, the GAP activity of RGS-GAIP is actually increased by ERK-mediated phosphorylation of serine 151 in the RGS domain (Ogier-Denis et al., 2000), resulting in reduced Gα-GTP available to activate ERK in the cell. A recent report showed that, in addition to second messenger-activated kinases, transmembrane tyrosine kinases can also regulate RGS phosphorylation and thereby their interaction with Gα subunits (Derrien 2001). At rest, RGS16 exists in a complex with inactive EGFR in several cell systems. Upon EGFR activation, RGS16 is phosphorylated at tyrosine residues within the RGS domain enhancing its GAP activity for Gαi. This translates to an increased inhibition of Gαi-linked signals. A particularly interesting implication of this work hinges on the fact that GPCRs, such as the muscarinic cholinergic receptors, can themselves activate EGFRs, which could then feedback to inhibit Gα signals directly through RGS16.
Reported RGS post-translational modifications
Rather than modifying GAP activity directly, phosphorylation can also affect RGS interactions with other proteins that compete with Gα subunits for binding. In the case of RGS3 and RGS7, phosphorylation is necessary for interaction with the intracellular scaffold protein 14-3-3 (Benzing et al., 2000). PKC phosphorylation allows RGS7 to associate with cytosolic 14-3-3 in lieu of Gαi/o, thereby reducing RGS7 GAP activity. Unfortunately, this study did not explore the distribution of Gβ5, which would have provided insight into the effect that 14-3-3 interactions may have on RGS7-mediated cellular signals.
B. Membrane Targeting and Lipid Modification
As previously discussed, most RGS proteins are predicted to be inherently hydrophilic, yet are found both in the cytosol and tightly bound to the membrane in vivo. Additionally, because receptor-Gαβγ complexes are membrane-bound, cellular mechanisms must be in place to direct RGS proteins to target Gα complexes. Several cytosolic RGS proteins translocate to the plasma membrane when exposed to GTPase-deficient Gα mutants (Druey et al., 1998; Saitoh et al., 2001), suggesting either direct recruitment of RGS by Gα-GTP or membrane recruitment through mechanisms initiated by G protein activation. Such mechanisms could include regulated post-translational modifications such as phosphorylation or lipid modifications, as well as enhanced interaction with protein binding partners.
Recruitment of RGS proteins to membrane-bound signaling complexes can be affected by phosphorylation, either directly or through binding partner interactions. For example, in studies of the mechanism by which atrial natriuretic peptide (ANP) inhibits astrocyte proliferation, it was found that ANP causes RGS3 and RGS4 to migrate from the cytosol to the membrane. These results were surprising since the ANP receptor isnot a GPCR (Pedram et al., 2000). RGS3 and RGS4 binding to the plasma membrane limits activation of Gαqand Gαi/o linked to endothelin receptors, which cause astrocyte proliferation. Their inhibition therefore reduces proliferation. Translocation of RGS3 and RGS4 is mediated by cGMP-dependent protein kinase (PKG)-induced phosphorylation. While it is not yet clear what mechanism is initiated by this phosphorylation, regulated trafficking of RGS proteins may underlie interactions between different signaling pathways as a common means of intracellular communication.
Many factors in addition to phosphorylation affect membrane attachment, most notably addition of fatty acid moieties to mature proteins. The peptide sequence of most RGS proteins does not contain consensus motifs for the covalent addition of either myristate or isoprenoids, important membrane anchors for many signaling proteins. However, RGS proteins do contain exposed cysteine residues, which can rapidly and reversibly incorporate palmitate at the cell membrane in the absence of recognized motifs. The addition of thioester-linked palmitate can contribute to membrane and subcellular localization as well as affect protein interactions (Mumby 1997). The exact mechanisms governing the addition and cleavage of palmitate are not fully understood; however, palmitoylation is a reversible process that can be modulated by GPCR activity (Mumby, 1997). Several members of the RGS family are palmitoylated in vivo and in vitro, with varying effects on their functions (Table 2). As discussed earlier with reference to RGS4 signaling, RGS4 and RGS10 are both palmitoylated at a conserved cysteine residue in their RGS box, which inhibits interaction of both proteins with Gα subunits (Tu et al., 1999; Fig.5). However, in assays with reconstituted receptors, which present a setting that is somewhat more physiological, palmitoylated RGS10 is actually a more effective GAP than the unpalmitoylated form. The hypothesis given for this is that the hydrophilic RGS10, which does not have an amphipathic helix (discussed above), is not recruited to the membrane to associate with Gα in the absence of palmitate. In the case of RGS10, therefore, membrane targeting overcomes the inhibition due to the conformational change induced by the RGS-box palmitate (Tu et al., 2001). The RGS-box cysteines are conserved in a variety of RGS families and represent a mechanism by which membrane proximity and conformational changes can directly affect GAP activity.

Potential molecular targets on RGS proteins for drug action. Schematic model of candidate regions on RGS proteins that could serve as targets for drugs to alter RGS protein functions. Target sites include amino acids and surrounding regions that are essential for direct RGS/Gα binding (1, 2, and 3); those that mediate RGS binding with other proteins/molecules to allosterically regulate RGS/Gα binding (4, 5, and 6); amino acids required for RGS protein membrane attachment (7, 8); and those residues that mediate RGS binding to GPCR (9) and to other regulatory and/or signaling proteins (10).
Palmitoylation of N-terminal cysteines occurs in a variety of signaling proteins and affects membrane attachment and subcellular targeting. RGS16, for example, requires functional cysteines at amino acids 2 and 12 for proper regulation of both Gαi- and Gαq-linked receptors in cells (Druey et al., 1999). The interpretation for this is that palmitoylation of Cys2 and Cys12 is necessary to target RGS16 to membrane bound Gα subunits. Surprisingly however, mutation of these shared residues in RGS4 doesnot inhibit either membrane attachment or RGS4 function in yeast (Srinivasa et al., 1998; Tu et al., 1999). As discussed above, palmitoylation of Cys2 and Cys12 can enhance RGS4 membrane binding and activity but also promotes palmitoylation of Cys95 in the RGS-box thereby reducing interactions between RGS4 and Gα. The different consequences of palmitoylation for RGS16 and RGS4 may therefore depend on the lack of a target cysteine within the RGS-box of RGS16.
Unlike the B/R4 family members, the amino termini of RGS-GAIP and other A/RZ proteins contain a N-terminal cysteine string, which is palmitoylated and acts as a membrane and subcellular targeting motif. Although the B/R4 family is recruited to the membrane through its amphipathic helix, multiple palmitoylation at the cysteine string of A/RZ proteins allows their membrane attachment (De Vries et al., 1996). Palmitoylation and phosphorylation may interact to control the unique subcellular localization of RGS-GAIP on clathrin-coated pits and its potential control of vesicle fusion, because only the membrane bound pool of RGS-GAIP is phosphorylated in cells (Fischer et al., 2000). These findings suggest that multiple types of post-translational modification may conspire to dictate proper localization and function of this RGS family.
C. Factors Regulating RGS Protein Half-Life
Modulation of protein stability is another mechanism by which cells regulate signaling events. Phosphorylation can change protein stability directly or by regulating interaction with binding partners (Benzing et al., 1999; Garrison et al., 1999). For example, RGS7, which is multiply ubiquitinated and rapidly degraded by proteosomes, has a short half-life. RGS7 protein levels increase after exposure to endotoxins, partially through tumor necrosis factor-mediated phosphorylation of a p38-kinase recognition motif on RGS7, which decreases its proteolysis (Benzing et al., 1999). Although the mechanism by which phosphorylation governs proteolysis is unknown, it could be through interaction of RGS7 with a binding partner. Interactions of RGS7 with its various binding partners (Gβ5, 14-3-3, polycystin) have not been fully explored; however, investigations indicate that binding partners do stabilize RGS7 (Kim et al., 1999;Witherow et al., 2000; Keren-Raifman et al., 2001). Along these lines, RGS7 and other R7 family members are highly degraded when ectopically expressed without Gβ5, indicating that complex formation is necessary for protein stability. Therefore, modifications that regulate these interactions can affect degradation. In yeast, regulation of the proteolysis of Sst2p results in an elegant feedback loop governing pheromone signals. Phosphorylation by G protein-activated MAPK reduces Sst2p degradation, which limits free Gβγ, thereby decreasing active MAPK available to phosphorylate Sst2p (Garrison et al., 1999).
Targeting of proteins for degradation is governed in part by a number of regulatory sequences including poly-proline regions or N-terminal polar residues. The N-end rule of degradation relates the half-life of a protein to the identity of the N-terminal residue, insofar as presence of an aspartate, glutamate, or cysteine at position 2 enhances ubiqutination and subsequent degradation (Davydov and Varshavsky, 2000). RGS4 quickly succumbs to this pathway when expressed in cell lines, with a half-life of less than 1 h (Davydov and Varshavsky, 2000). Cleavage of the RGS4 start methionine reveals Cys2, which is then arginylated, thereby destabilizing RGS4. Inhibition of arginylation blocks ubiquitination and subsequent degradation. The percentage of RGS4 degraded through this pathway varies depending on cell type. A speculation is that, since cysteine 2 is one of the principle sites of palmitoylation (see above), addition of fatty acid could interfere with arginylation and subsequent degradation. Palmitoylation could therefore not only assist with targeting RGS4 to signaling compartments but regulate protein levels within the cell as well.
An unusual mechanism of RGS regulation through proteolysis has been described for the yeast RGS protein Sst2p (Hoffman et al., 2000). While full-length Sst2p is ∼70 kDa and primarily bound to the plasma membrane, yeast also express two proteolytic products, a 36-kDa band corresponding to the C terminus and a matching 45-kDa band corresponding to the N terminus, both products of regulated cleavage of the full-length protein. Quite unexpectedly, this is not a step in the degradation of Sst2p as the N and C termini can re-associate to inhibit growth arrest. However, unlike the uncleaved protein, the reassembled Sst2p does not reduce transcriptional activation, so cells both grow and continue to produce high levels of pheromone-induced mRNA. The proteolytic processing of Sst2p depends on the components of the pheromone signaling pathway that are concentrated at the tip of the mating projection. Theoretically, therefore, proteolysis of Sst2p is enhanced at the mating tip, making it more responsive to transcriptional responses from pheromone signals than the rest of the cell. Clearly Sst2p, which can reduce both growth arrest and transcriptional activation, is under multiple levels of control, the interplay of which are not yet fully understood.
D. Targeted Subcellular Localization of RGS Proteins
Compartmentalization is a major mechanism by which cells regulate protein signaling capacity. Proteins can be compartmentalized individually or with a subset of signaling components to either facilitate or impede signals. Identified cellular mechanisms to regulate RGS localization include masking or exposing targeting sequences, altering post-translational modifications, and forming complexes with protein binding partners.
Some RGS proteins are targeted to specific signaling complexes through interaction with binding partners. These can include G protein-coupled receptors and signaling components, as previously discussed in the case of RGS4 and RGS12 (Snow et al., 1998a; Xu et al., 1999). Other RGS proteins may be localized by interaction with non-GPCR binding partners such as GIPC, β-COP, 14-3-3, polycystin, or MIR16 (discussed above). Since studies are beginning to reveal that the membrane-bound populations may have different signaling properties than those in solution, constitutive interaction with membrane-bound partners may represent a universal mechanism to regulate RGS availability and signals (Tu et al., 2001).
A growing list of RGS proteins are localized to the nucleus, both as native protein and when recombinantly expressed (Table2). Indeed, cytosolic localization of some RGS proteins may reflect competition between nuclear import and export signals located in the N terminus and in the RGS domain (Chatterjee and Fisher, 2000a; Heximer et al., 2001). Cells may use the nucleus as a storage compartment for RGS proteins to regulate their Gα GAP activity. In the case of RGS10, PKA phosphorylation at the C terminus causes translocation to the nucleus and makes RGS10 unavailable to limit GPCR signals at the plasma membrane. However, nuclear RGS proteins may also serve important cellular roles independent of their actions at the plasma membrane. For example, nuclear localization of RGS3T is associated with its capacity to cause apoptosis, although the underlying mechanisms of this are unclear (Dulin et al., 2000). Variants of RGS12 with a short C-terminal region all localize to the nucleus and one variant, RGS12TS-S, is expressed in punctate nuclear foci. Because many tumor suppressor proteins also exhibit this expression pattern, this localization may be functionally relevant and may indicate a role for RGS12 in cell cycle events. Lending credence to this hypothesis, overexpression of this splice variant in COS-7 cells causes abnormal nuclear morphology and division (Chatterjee and Fisher, 2000b). RGS2 and RGS7 have also been reported in the nucleus; however, very little is known about the functional consequences of this localization (Chatterjee and Fisher, 2000a,b; Song et al., 2001; Zhang et al., 2001).
E. Factors Regulating Cellular Expression of RGS Proteins
Regulation of RGS mRNA expression and protein translation represent additional cellular mechanism important for determining the levels of protein available to direct signals in vivo. Differential expression of RGS proteins can lead to vast changes in cellular response properties. The highest proportion of RGS genes are expressed in brain tissue, which is in line with the extensive diversity of neuronal and glial GPCRs and the signal modulation necessary for proper brain function. Because the brain is by far the most differentiated organ, understanding the regulation of RGS expression in neurons and glia is a leading focus of research.
Although a clear understanding of the regulation of RGSprotein levels is important, surprisingly little is known on this subject. Efforts to date to detect native RGS proteins are hampered by the lack of effective antisera as experimental tools. In a few cases where specific antisera are available that readily recognize recombinant protein, these sera fail to detect native protein in cells that express high levels of mRNA. One example of this is RGS14, which, by in situ hybridization, is highly expressed in the cerebellum (Grafstein-Dunn et al., 2001), however, is undetectable in cerebellar lysates using specific antisera (Hollinger et al., 2001). Reasons for this discrepancy are unclear. At rest, the expression of many native RGS proteins may be so low that they fall below the limit of detection of many of the available antibodies. Translational control could therefore present a primary mechanism of regulating RGS protein levels. Some specific antisera are now available that recognize native RGS proteins; however, few descriptions of regulation of RGS protein levels have been reported.
Considerably more information is available describing mRNA levels. Several studies have examined differential steady-state distribution of one or more RGS proteins in the hopes of using this information to speculate on functional implications (see De Vries et al., 2000). In the first of these studies, Gold and colleagues (1997) investigated the regional distribution of nine RGS subtypes (RGS3–11) in brain. Most of these proteins were expressed broadly, although RGS9 was seen almost exclusively in the striatum. Later studies showed that striatal RGS9 is actually the longer splice variant of RGS9, RGS9-2. Some other RGS proteins also show limited expression patterns. For example, RGS16 mRNA is primarily in thalamic and hypothalamic nuclei in brain, regions involved in sensory processing and circadian regulation (Grafstein-Dunn et al., 2001). Studies have also investigated distribution in other tissues, particularly in the heart and in lymphocytes. Unfortunately, whereas some RGS proteins exhibit specific expression profiles, most are found to varying degrees in multiple tissues or brain regions, and therefore these expression profiles furnish limited information about function.
More informative than the cataloging steady-state expression levels is exploring regulation of mRNA expression by extracellular stimuli such as GPCR activation, seizure induction, and long-term potentiation (Burchett et al., 1998; Ingi et al., 1998; Ni et al., 1999). Several studies investigating regulation of mRNA after stimulation have focused on RGS2 and RGS4. Initial research examined reciprocal regulation of simple RGS proteins by signals that they help control (e.g., Gαq-directed increases in RGS2 which could reduce Gαq activation) (Song et al., 1999). However, the regulation of RGS2 and RGS4 mRNA has proven to be more complex than anticipated. RGS2 mRNA is up-regulated in cells by a variety of tested stimuli including cAMP, Ca2+, and diacylglycerol, as well as drug treatments, seizures and LTP induction (see De Vries et al., 2000). In most cases, RGS2 acts as an immediate early gene similar to c-fos (Robinet et al., 2001). RGS4 mRNA regulation in vivo has been very difficult to interpret, with a recent report postulating that RGS4 expression is differentially altered by stress. Data from experimental animals could therefore be confounded by stress-related changes (Ni et al., 1999;Gold et al., 2000). Although the fact that RGS4 is regulated by stress could lead to insights as to its physiological importance, the early studies on RGS4 regulation need to be revisited with an eye toward these new developments.
F. Expression of Alternatively Spliced Gene Products
As we have discussed, RGS regulation can occur post-translationally through modifications of protein interactions and through regulation of transcription or translation. Another common mechanism by which cells can fine-tune signaling responses using a limited template of genes is through alternate splicing of the gene itself, which fundamentally alters the structure and function of a protein by adding or deleting particular motifs. Several RGS proteins are produced as multiple gene products (Table3) (De Vries et al., 2000). Although the functional significance of most of these splice variants is currently unknown, research into several RGS proteins (RGS3, RGS9, RGS12) is lending insight into how structural variations can fundamentally alter protein regulation and function.
Reported RGS splice variants
As previously mentioned, a variant of RGS3, RGS3T, directs apoptotic programs in cells (Dulin et al., 2000). This activity requires the unobstructed N terminus of the truncated product. Although this sequence is present in the full-length protein, the extended N terminus seems to shroud the relevant residues since full-length RGS3 exhibits neither nuclear localization nor pro-apoptotic activity. Several splice variants of RGS3 have been identified (RGS3, RGS3T, RGS3S) that may vary in expression and cellular responses. RGS3S for example is expressed preferentially in heart. Although both are truncated forms of RGS3, the relationship between RGS3S and RGS3T is currently unclear (Mittmann et al., 2001). Another potential splice variant with an extended N terminus, PDZ-RGS3, regulates reverse signaling of the neurotrophin Ephrin B, as described above (Lu et al., 2001).
The two known splice variants of RGS9 are particularly interesting because each shows very specific patterns of expression, suggesting distinct physiological roles. The shorter form, RGS9-1, is expressed exclusively in the retina where it acts as a GAP for transducin (Gαt), as described above. In contrast, RGS9-2, a C-terminal extended variant, is found almost exclusively in the striatum (Granneman et al., 1998; Rahman et al., 1999; Zhang et al., 1999). One demonstrated function of RGS9-2 is to desensitize μ-opioid receptors found specifically in the striatum. RGS9-2 but not RGS9-1 reduces opioid responses in vitro, and when RGS9 protein is reduced in mice, morphine responses are enhanced and no short-term tolerance to morphine is observed (Rahman et al., 1999; Garzon et al., 2001). Although RGS9-1 is not expressed in brain regions responsible for morphine responses, the in vitro studies also demonstrate that RGS9-1 and RGS9-2 are not interchangeable. RGS9 thus represents one gene alternately transcribed to regulate two entirely separate systems.
RGS12, with twelve known splice variants, is the most highly spliced RGS protein described to date. Initially only four variants were identified, containing either a PDZ domain, a PDZ binding motif, both, or neither. As described above, the PDZ domain present at the N terminus of RGS12 binds specifically to the PDZ binding motif in the IL-8 receptor tail as well as to the binding motif at its own C terminus (Snow et al., 1998a). The proposed model suggests that in the longest splice variant, the N and C termini bind to each other, occluding the RGS domain. Receptor recruitment of the PDZ domain opens the protein and allows it to act as a GAP for the receptor-linked Gα subunit. Although this model is plausible, it does not consider the other motifs found in RGS12 (Fig. 1) or the additional RGS12 splice variants (Chatterjee and Fisher, 2000b). All known variants of RGS12 contain identical core motifs (including the RGS domain) but exhibit combinations of three different N and C termini. The N and C termini are expressed in a tissue-specific manner. Forms of RGS12 with a truncated C terminus are particularly intriguing because of their altered cellular localization and their possible relationship to cell cycle control. One variant, RGS12TS-S changes distribution throughout the cell cycle from nuclear foci, in which it is found during most phases of the cell cycle, to the chromosome during metaphase. When overexpressed, this form of RGS12 causes abnormal nuclear shapes (including nuclear blebbing) and altered nuclear division. Taken together these findings suggest that this splice variant of RGS12 is involved in some aspect of nuclear reorganization during cell division.
VI. RGS Proteins As Therapeutic Targets
As cellular roles for RGS proteins come into focus, it is clear that these proteins could represent attractive therapeutic targets. In the following section, we will illustrate some of the potential roles of RGS proteins in physiology and disease based on limited available information. Unfortunately, very little is currently known about RGS proteins expressed at physiological levels in their native environment. Due to technical considerations, most available studies use overexpressed RGS proteins, often in non-native environments, with the hope that this phenotype will provide clues to their biological function. Potentially more informative genetic knock-out approaches are hampered in some cases by embryonic lethality, which limits conclusions that can be drawn. Where possible, we have attempted to infer potential roles of certain RGS proteins in organ physiology and disease by placing relevant data into a larger context. In addition to showing an overview of their potential biological importance, we will also discuss candidate molecular targets for the generation of RGS acting drugs.
A. Roles for RGS Proteins in Cell Migration and Development
In lower eukaryotes, RGS proteins regulate several aspects of embryonic development including glial differentiation, embryonic axis formation, and skeletal and muscle development (Granderath et al., 1999; Fukui et al., 2000; Wu et al., 2000). The Drosophilagene product Loco is an example of an RGS protein that acutely affects the viability of an organism. Drosophila embryos lacking Loco fail to hatch although they appear morphologically normal (Granderath et al., 1999). Upon close examination, however, these embryos exhibit profound defects in glial cell-cell interactions such that axons remain partially unsheathed and embryos lack a blood-brain barrier. These data indicate that Loco is particularly important in glial cell adhesion and motility. Mouse knock-outs of the closely related mammalian gene RGS14 are lethal at early embryonic stages due to improper attachment to the uterus (D. Siderovski, personal communication; Zhong and Neubig, 2001), indicating that this gene may also be involved in cell adhesion during development in mammals. Although defects are found in animals deficient in RGS proteins, other studies show that overexpression of some RGS proteins can also cause developmental defects. For example, exogenous RGS2 or RGS4 inXenopus embryos results in severe skeletal and muscular abnormalities (Wu et al., 2000). Because both of these proteins effectively inhibit Gαq, the observed developmental problems may result from over-inhibition of Gαq-linked pathways.
Several RGS proteins can affect cell migration during development (Bowman et al., 1998). Axin for example inhibits axis formation in embryos by scaffolding binding partners together to alter gene transcription (see above). Mammalian RGS3 can directly affect renal tubule cell migration, which underlies the formation of the kidney (Gruning et al., 1999). The RGS-like proteins in the p115RhoGEF family influence cell migration and shape by activating RhoA. Interestingly, studies have shown that some of the RGS proteins that affect cell migration block Gα12/13 signals in addition to being GAPs for Gαi/o and Gαq (Moratz et al., 2000; Reif and Cyster, 2000). Because Gα12/13 promote both cell migration and oncogenesis more effectively than other Gα subunits (Radhika and Dhanasekaran, 2001), the Gα12/13antagonist function of these RGS proteins may underlie their effects on cell motility.
Certain RGS proteins also play a role in cell proliferation and apoptosis. For example, as discussed previously, inhibition of astrocyte proliferation by atrial natriuretic peptide occurs through translocation of RGS3 and RGS4 to the membrane (Pedram et al., 2000). Certain RGS proteins can also induce cell death. The truncated variant of RGS3, RGS3T, which shares many features of full-length RGS3 but is highly enriched in the nucleus, causes apoptosis. While nuclear localization of RGS3T correlates with apoptosis in transfected cells, the function of native protein in host cells is still a mystery. Based on recent reports, at least three variants of RGS3 are clearly involved in cell cycle and cell migration (Dulin et al., 2000; Reif and Cyster, 2000; Lu et al., 2001).
B. Roles of RGS Proteins in Organ Physiology
A number of RGS proteins modulate cardiac development and output (Sierra et al., 2000). One mechanism to control cardiac ouput is by directly controlling the heart rate through activation of the parasympathetic nervous system. This reduces heart rate by modulating ion currents through GIRKs. Several studies have shown that the native kinetics of these K+ channels can only be recreated in recombinant systems when RGS4 is coexpressed, suggesting that native cardiac excitability depends on the expression of this or similar RGS proteins (Fujita et al., 2000; Mark and Herlitze, 2000). Evidence is also mounting that RGS proteins contribute to cardiac growth, in addition to excitability. NFAT, the transcription factor involved in initiating cardiac growth, requires Ca2+ oscillations to translocate to the nucleus and cause gene transcription. As described above (Fig. 2), the proper frequency of these oscillations may be due to the regulation of RGS activity through PIP3 and Ca2+/CaM. Over-expressing RGS4 may shift the ratio of the proteins in this cascade, varying Ca2+ oscillations and thereby affecting cardiac growth. This scenario was tested in studies in which RGS4 was ectopically expressed in adult mouse hearts. After pressure overload in the ventricles of the animals, RGS4 overexpressing mice showed reduced Gq-mediated hypertrophy, indicating that the mechanisms governing cardiac growth were disrupted (Rogers et al., 1999). Further evidence that the relative ratio of RGS proteins is important for regulating signals critical for cardiac function comes from studies of RGS modulation of GIRK1/2 channels inXenopus oocytes (Keren-Raifman et al., 2001). In these studies, expressing RGS4 (or RGS7/β5) could accelerate, inhibit, or have no effect on GIRK currents, depending on the levels of protein in the cell. These results could help to explain some inconsistent reports regarding the physiological effects of particular RGS proteins.
Several studies indicate that RGS proteins can fine-tune immune responses. Both T- and B-cells rapidly regulate several RGS proteins in response to activation by a growing list of agents (Beadling et al., 1999; Cho et al., 2000; Reif and Cyster, 2000). The precise regulation of these genes and proteins depends on both the cell type and identity of the activating agent (Cho et al., 2000; Reif and Cyster, 2000), suggesting that specific immune responses may require modulation by a subset of RGS proteins. Both simple and complex RGS proteins are regulated by immune activation, therefore a combination of these may define the response of the cells. RGS1, a B/R4 family member, is either constitutively expressed or inducible in a number of B-cell lines (Moratz et al., 2000). Increased RGS1 inhibits chemokine-induced B-cell migration, GPCR-linked Ca2+ signals, Gα12-induced stress-activated protein kinase, and serum response-element activation (potential effector antagonist functions). B-cell maturation in particular may be regulated by levels of RGS1. Germinal cells constitutively express high levels of RGS1 and are unresponsive to chemokine signals, whereas mature B-cells express low levels of RGS1 and respond to chemokines unless RGS1 is induced. Therefore, a cell cycle-dependent decrease in RGS1 levels could control the migration of B-cells out of the germinal center in lymphoid tissue (Moratz et al., 2000).
A recent RGS2 genetic knock-out demonstrated that this simple RGS protein also plays an important role in the immune response (Oliveira-Dos-Santos et al., 2000). RGS2(−/−) immune tissue exhibits a reduced response to activating agents both in vitro and in vivo. RGS2(−/−) T cells do not proliferate as well as controls after activation by a variety of stimuli and produce low levels of the T-cell growth factor IL-2. Because these deficits are most pronounced within the first 24 h after stimulation, they are consistent with a model of RGS2 as an immediate early gene involved in T-cell activation. The impaired response seen in these cells extends to the whole animal. When injected with virus, RGS2-deficient mice show reduced footpad swelling, indicative of impaired T-cell activation and recruitment to the site of infection.
C. RGS Proteins in Neuronal Function and Behavior
Numerous studies have investigated RGS modulation of neuronal signals. Because neuronal signals are under such complex control, research into this area is particularly challenging. Some studies have clearly indicated that ion channels underlying signals such as neurosecretion and dendritic Ca2+ fluxes can be tightly regulated by exogenous RGS proteins (Jeong and Ikeda, 2000;Melliti et al., 1999, 2000; Schiff et al., 2000; Zhou et al., 2000). However, the receptor and Gα selectivity of RGS proteins, their multiple interacting partners, and cellular regulation all influence their signaling capacity. Investigating the contribution of native RGS proteins to different neurological processes is technically challenging, particularly because a number of RGS subtypes are often expressed.
Can RGS proteins directly affect behavior? Studies have used RGS-insensitive Gα subunits as well as antisense gene knock-down to identify roles of endogenous RGS proteins in behavior. Whereas several mammalian studies have examined behavioral effects of RGS proteins, genetic manipulation in lower eukaryotes provides the opportunity to examine global effects much more readily. In C. elegans, RGS proteins integrate reproductions with environmental considerations such as food availability (Hajdu-Cronin et al., 1999; Dong et al., 2000;Wilkie, 2000). Egl-10, one of the first RGS proteins discovered, governs egg-laying behavior by negatively regulating the C. elegans Gαo homolog. Yet, C. elegans RGS1 and RGS2, which are also negative regulators of Gαo, cannot substitute for Egl-10. RGS1 and RGS2 do not affect egg-laying unless worms are food-deprived. In normal worms, food deprivation inhibits this behavior, which surges once food is restored. However, if RGS1 and RGS2 are genetically knocked out, worms do not recover from food deprivation. Thus, the interplay of the RGS subtypes produces proper behavioral responses of worms to their surrounding environment.
In mammalian systems, little is known about behavioral outcomes of altering RGS levels. A recent genetic knock-out lends insight into the role that RGS2 plays in the control of both physical and psychological responses to stress (Oliveira-Dos-Santos et al., 2000). In addition to displaying a decreased immune response (described above), engineered animals seemed unusually prone to injuries from fights with nontransgenic littermates. After testing them in a number of paradigms, the investigators unexpectedly found that RGS2 knock-out animals exhibit increased anxiety behaviors. Anatomical studies show that synapses and dendritic spines in the CA1 region of RGS2(−/−) hippocampus are decreased, suggesting that RGS2 plays a role in proper development of synaptic connections in the hippocampus. These studies provide a compelling example of how different RGS proteins can fundamentally affect the behavior of an organism.
To demonstrate the effects of native RGS proteins in behavior, investigators have employed a complimentary approach. While only published in abstract form to date, a group of studies using a potentially fruitful approach to identify functions of native proteins merit description. The investigative team expressed an RGS-insensitive mutant of Gαq(G188S) in rat brain, to isolate the behavioral contribution of native RGS proteins acting on Gαq (Comery et al., 2000; Grauer et al., 2000;Howland et al., 2000). Gαq(G188S) transgenic animals exhibit excessive vibrations in the absence of stimulation, possibly a global motor deficit. The animals also display impaired inhibition of startle by a prepulse of stimulus, indicating a higher level of stress response. Several transgenic rat lines also exhibit increased startle, although consistent increases in global anxiety or fear conditioning are not observed. Differences in expression levels of Gαq(G188S), as well as possible compensatory mechanisms such as increases in Gα11, may limit the conclusions that can presently be gained from these studies since behavioral effects differ somewhat across lines. However, these transgenic rats provide a useful model system for assessing the global effects of blocking native RGS effects on Gαqfunctions in the central nervous system.
Studies of RGS9(−/−) knock-out mice provide another example of RGS roles in the sensory system. RGS9-1, expressed exclusively in the retina, is one of the few RGS proteins for which a discrete biological function has been identified. RGS9-1 is a potent GAP for transducin (Gαt), which underlies the proper regulation of ion currents in photoreceptors. As discussed above, mice lacking RGS9-1 respond poorly to light. In particular, cells do not recover from pulses of light, giving these animals a form of night blindness (Chen et al., 2000; Lyubarsky et al., 2001). Functional RGS9-1 is therefore a principle element needed for proper vision.
D. RGS Proteins in Disease States
Several recent reports suggest that RGS proteins may be involved in a number of diseases. For example RGS16, also known as RGS-r, is a retinally abundant RGS protein that may be modified in autosomal recessive retinitis pigmentosa, a form of blindness (Bressant et al., 2000). Genetic linkage studies designed to identify naturally occurring mutations in affected individuals revealed at least one intron mutation in RGS16. Although the mutation is in the noncoding region of the gene, it could account for some forms of this disease perhaps by altering levels of RGS16 expression. Genetic studies also identified RGS8 as a possible determinant of hereditary prostate cancer (Sood et al., 2001). The chromosomal region associated with this form of prostate cancer carries a module of 13 genes, including the gene for RGS8. Although this investigation did not demonstrate a direct role for RGS8 in the etiology of the disease, these findings raise the intriguing possibility that mutations in the RGS8 coding sequence or in its promoter regions may play a role in the development of prostate cancer.
Recent studies suggest a role for RGS proteins in psychiatric disorders. By examining changes in gene expression to provide insight into the underlying causes of schizophrenia, studies found that RGS4 expression levels are consistently and significantly altered more than any other gene tested (Mirnics et al., 2001). In analyses of differential expression of almost 8000 genes on cDNA microarrays, every patient tested demonstrated decreased RGS4 mRNA in their prefrontal, motor, and visual cortices. In contrast, patients with major depressive disorder do not show changes in RGS4 mRNA levels, indicating that this reduction is specific to schizophrenia. Although the physiological reason for down-regulation of RGS4 mRNA is unclear, several hypotheses exist. For example, low levels of RGS4 could be a risk factor for developing schizophrenia. This hypothesis is supported by the fact that most antipsychotics limit overactive Gi/o signals activated by D2 dopamine receptors, which would be exacerbated by reduced levels of RGS4 (Mirnics et al., 2001). Alternatively, RGS4 down-regulation could be secondary to other problems found in schizophrenia such as reduced synaptic density or modified stress perception in schizophrenic patients (Mirnics et al., 2001).
Initial evidence suggests that striatal RGS9-2 could play a role in Parkinson's disease. In Parkinson's, dopaminergic neurons in the substantia nigra degenerate, leading to a disregulation of thalamocortical circuits. Current models posit that dopamine regulates the opposing “direct” and “indirect” pathways through the basal ganglia, thereby controlling movement. This requires the activity of both D1 and D2 dopamine receptors in the striatum. A recent study of Parkinson's patients investigated variations in striatal levels of proteins that are potentially involved in dopaminergic signaling (Tekumalla, 2001). Although the study used a limited number of patients, they found significant increases in two proteins: a transcription factor (ΔFosB), which is known to be affected by alterations in dopaminergic signals, and RGS9-2. The latter finding was somewhat unexpected, since decreases in agonist concentration are anticipated to sensitize receptor signals. Because RGS9-2 specifically inhibits dopamine D2 receptor signals, increases in this protein could disrupt the equilibrium between D1 and D2 receptors, changing the balance between the direct and indirect pathways. If RGS9-2 levels change early in the disease, they could exacerbate the disregulation caused by the declining dopamine levels. RGS9-2 increases could also limit the efficacy of drugs designed to enhance dopamine signals in the striatum, a phenomenon known to happen in the latter stages of the disease (Tekumalla, 2001). The discovery of the altered regulation of RGS9-2 in the striatum may lead to new insights into the chronic decline in movement control seen in the disease.
Preliminary evidence suggests certain RGS proteins may also be involved in chronic heart failure. Heart failure is correlated with alterations in both Gαq-linked signaling pathways and Gαi (Takeishi et al., 2000). Two studies have independently shown that both mRNA and protein levels of several RGS proteins are up-regulated in failed human hearts. To date, all of these are members of the B/R4 family including RGS2, RGS3, and RGS4. Both RGS2 and RGS4 are effective GAPs for Gαq in vitro, therefore an increase in their protein levels may represent an adaptive mechanism to reduce Gαq over-activity, which causes increased Ca2+ and PKC activity and leads to cardiac hypertrophy and exacerbated heart failure (Takeishi et al., 2000). Additional evidence sustains this hypothesis. In several model systems, cardiac overload induces RGS4 overexpression. Furthermore, exogenous RGS4 reduces cardiac hypertrophy in response to pressure overload (in this case an adaptive mechanism) and delays the heart failure associated with Gαqoverexpression in transgenic mice (Rogers et al., 2001). Increases in RGS2, RGS3, and RGS4 in failing hearts support the existence of a negative feedback loop for long-term regulation of cardiac hypertrophy.
RGS proteins also offer a great deal of therapeutic potential in altering the effects of other drugs, including drugs of addiction. RGS can potentially modulate signal desensitization, modify which signals are targeted, and even change the potency of a compound. In rats, native RGS proteins modulate acute tolerance to morphine (Garzon et al., 2001). Although reducing the native levels of several RGS proteins has an effect on responses to morphine, these studies focused on RGS2 and RGS9. Findings indicate that these proteins have opposing effects: knocking down RGS2 makes morphine less potent, whereas reducing RGS9 produces analgesia at lower concentrations and discourages tolerance to morphine after acute administration or repeated treatments. Native RGS9 therefore facilitates tolerance and reduces the potency of morphine. Drugs targeted to reduce RGS9 signals may therefore show major benefits in the treatment of pain.
E. Molecular Targets for Drug Development
As outlined above, RGS proteins serve important roles as modulators and integrators of G protein signaling and are central participants in both physiology and disease. As such, RGS proteins have become new candidates for therapeutic intervention (Jones et al., 2000;Zhong and Neubig, 2001). Drugs that alter RGS actions could affect cellular functions in a number of ways including: 1) potentiating hormone and neurotransmitter actions, 2) prolonging or enhancing the effects of other drugs that stimulate G protein signaling pathways, or 3) affecting downstream signaling pathways that are activated by RGS proteins. For a more comprehensive discussion of this topic and possible therapeutic applications of RGS-directed drugs, see Zhong and Neubig, 2001. A number of identified regions, both within and outside of the RGS domain, could serve as targets for drug development. The regions involved fall into four categories (illustrated in Fig. 5): 1) direct RGS/Gα binding, 2) allosteric modulation of RGS/Gα binding, 3) RGS membrane attachment, or 4) RGS interactions with GPCRs, regulatory proteins, and/or downstream effectors.
1. Direct Modulation of RGS/Gα Binding.
The most obvious sites for possible drug actions are amino acids at the RGS/Gα contact interface. Drugs that block this interaction could prevent the inhibitory effects of RGS proteins on G protein signaling whereas drugs that mimic RGS actions could directly limit G protein signaling. Two structures of Gα subunits complexed with an RGS protein have been reported: RGS4/Gαi1-GDP-AlF (Tesmer et al., 1997) and RGS9-1/Gαt-GDP-AlF
(Tesmer et al., 1997) and RGS9-1/Gαt-GDP-AlF -complexed with γ-PDE (Slep et al., 2001). In each case, the surface loops of the guanine nucleotide “switch” regions of activated Gα make contact with three sites on the RGS domain (Fig. 5). Seventeen residues on the surface loops of RGS4 connecting helices α3–α4 (site 1), α5–α6 (site 2), and helices α7–α8 (site 3) of the RGS domain make contact with Gαi1.
-complexed with γ-PDE (Slep et al., 2001). In each case, the surface loops of the guanine nucleotide “switch” regions of activated Gα make contact with three sites on the RGS domain (Fig. 5). Seventeen residues on the surface loops of RGS4 connecting helices α3–α4 (site 1), α5–α6 (site 2), and helices α7–α8 (site 3) of the RGS domain make contact with Gαi1.
Mutational analyses demonstrate the importance of certain amino acids essential for RGS/Gα contact, which serve to highlight broader surrounding interfaces that could be targeted with drugs. Alanine substitution of residues in RGS4 that directly contact Thr182 of Gα (Glu87, Asn88 in site 1, or Asn128 in site 2) completely abolishes RGS4 GAP activity and Gα binding (Srinivasa et al., 1998). Site 2 on both RGS4 and RGS9-1 interacts directly with the switch regions of Gα-GTP (Tesmer et al., 1997; Slep et al., 2001). A number of amino acids within this region, when modified directly or when bound by other proteins, have been reported to enhance RGS effects on Gα. ERK-mediated phosphorylation of a Ser151 within site 2 of RGS-GAIP enhances its GAP activity toward Gαi1(Ogier-Denis et al., 2000). Similarly, two different residues within site 2 of RGS9-1 (Arg360 and Trp362) make direct contact with γ-PDE, and this interaction markedly potentiates RGS9-1 effects on Gαt GTPase activity (Slep et al., 2001). Although these individual residues are unique to RGS9-1 and RGS-GAIP, analogous interfaces in other RGS may serve as a platform for direct contact with specific binding partners, which could be targeted by small molecules as possible drug targets. Within site 3 of RGS4, three amino acids (Asp163, Ser164, and Arg121) interact with Thr182 of Gα following initial RGS/Gα binding. Of these amino acids, Ser164 is highly conserved among RGS proteins and is phosphorylated by PKC in RGS3, RGS7 (Benzing et al., 2000). Phosphorylation of RGS2 by PKC blocks its GAP activity toward Gαq/11(Cunningham et al., 2001). In contrast, phosphorylation of this Ser in RGS3 and RGS7 promotes their interactions with the cytosolic scaffolding protein 14:3:3 and prevents RGS interactions with Gα (Benzing et al., 2000). In both cases, RGS-Gα interactions are blocked, albeit by distinct mechanisms.
Regions on Gα that interface directly with RGS provide a potential binding pocket that also could serve as drug targets. Based on comparisons of the solution NMR structure of RGS4 and RGS-GAIP (de Alba et al., 1999; Moy et al., 2000) with the crystal structure of RGS4/Gαi1-GDP-AlF (Tesmer et al., 1997), helices α1 and α9 of the RGS domain reorient upon initial contact with Gαi1 and form a binding pocket for Thr182 of Gαi1. Mutating a conserved Gly residue immediately adjacent to Thr182 in Gαi1 or the equivalent Gly of Gαq abolishes RGS effects on Gα (DiBello et al., 1998; Lan et al., 1998), thereby demonstrating the importance of this binding pocket as a possible drug target (Comery et al., 2000). Taken together, these findings suggest that RGS/Gα contact interfaces could be targeted by small molecules that modify RGS actions on Gα signaling functions important for physiology and disease.
(Tesmer et al., 1997), helices α1 and α9 of the RGS domain reorient upon initial contact with Gαi1 and form a binding pocket for Thr182 of Gαi1. Mutating a conserved Gly residue immediately adjacent to Thr182 in Gαi1 or the equivalent Gly of Gαq abolishes RGS effects on Gα (DiBello et al., 1998; Lan et al., 1998), thereby demonstrating the importance of this binding pocket as a possible drug target (Comery et al., 2000). Taken together, these findings suggest that RGS/Gα contact interfaces could be targeted by small molecules that modify RGS actions on Gα signaling functions important for physiology and disease.
2. Allosteric Modulation of RGS/Gα Binding.
Regions indirectly involved with RGS-Gα interactions also may provide effective targets for drugs that act allosterically to modulate RGS actions. At least two examples of allosteric regulation of RGS/Gα binding have been reported. As discussed, PIP3 and Ca2+/CaM bind the RGS domain at charged residues in helix α5, opposite of RGS/Gα contact face, and PIP3 binding blocks RGS-Gα interactions (Popov et al., 2000). In another example of allosteric modulation of RGS/Gα, covalent addition of the fatty acid palmitate to a conserved Cys within helix α4 of the RGS domain blocks RGS-Gα interactions, although this region is not directly involved with Gα contact (Tu et al., 1999). Other proteins also bind RGS domains at sites distinct from the RGS/Gα interface that could provide potential drug targets. For example, MIR16 apparently binds to undefined residues in helices α1 and α2 within the RGS domain of RGS16 (Zheng et al., 2000). Axin binds APC at a site in the RGS domain that is removed from the Gα binding face (Spink et al., 2000). This binding region (also referred to as the “B site”) (Zhong and Neubig, 2001) forms a groove-spanning helix α3, α4, and α5 (Fig.5) which is conserved across other RGS domains. Although no information is available about the effects of MIR16 or APC binding effects on RGS-Gα interactions, these observations raise the possibility that small molecules could target analogous regions on other RGS proteins as possible therapeutic agents.
3. Selectivity of RGS-Gα Interactions.
Ideally, a clinically useful agent should be able to distinguish between discrete RGS-Gα interactions. Therefore, residues that determine RGS specificity for Gα interactions would be of great interest. RGS-PX1 and p115RhoGEF contain distantly related RGS-like domains that exhibit strict selectivity for Gαs and Gα12/13, respectively, which could be targeted with specific compounds. However, a much more difficult challenge will be targeting the closely related RGS subfamilies that interact with both Gαi and Gαq family members. Although many amino acids that contact Gα are conserved among RGS subfamilies, certain RGS proteins have unique amino acids that dictate Gα recognition. For example, RGS2 and RGS4, although closely related members of the B/R4 subfamily, exhibit surprisingly different Gα selectivity. Whereas RGS4 is an effective GAP for both Gαi family members and Gαq (Hepler et al., 1997), RGS2 is highly selective for Gαq in vitro (Heximer et al., 1997) and is a much more potent inhibitor of Gαq than Gαi signaling in cells (Ingi et al., 1998; Heximer et al., 1999). Three amino acids directly involved in RGS4 contact with Gαidiffer in RGS2 (Cys106, Asn184, and Glu191 in RGS2), and exchanging these residues between RGS2 and RGS4 switches their Gα preferences. These studies suggest that discrete structural features are important determinants of selective RGS-Gα interactions and raises the possibility that therapeutic compounds could be designed that selectively target one RGS-Gα interaction but not others.
4. Modulation of RGS Membrane Localization.
A different strategy for blocking RGS protein cellular actions could be to interfere with RGS membrane attachment. The class of cholesterol-lowering drugs known as statins have shown new promise as therapeutic agents because they prevent isoprenylation of many signaling proteins, thereby blocking their membrane attachment and signaling capacity (Bellosta et al., 2000). Since RGS proteins must associate with the plasma membrane to modulate Gα signaling events, small molecules that prevent RGS membrane attachment could serve as effective drugs to alter RGS functions. As discussed elsewhere in the text, several RGS proteins contain amphipathic helices at their N termini that are required for RGS membrane attachment (Figs. 2 and 5). Drugs that bind to involved regions, or drugs that affect palmitoylation, which contributes to RGS membrane attachment, may prevent RGS membrane recruitment and block RGS actions on G protein signaling.
5. Modulation of RGS Binding to GPCR, Effector and/or Regulatory Proteins.
Last, RGS interactions with non-G protein binding partners could provide many future targets for drug development. As discussed, RGS proteins bind directly to a variety of proteins involved in diverse cellular processes. Therefore, therapeutic agents that block RGS interactions with downstream signaling proteins could affect such specific cellular processes as ion conductances, intracellular trafficking, cell growth, and differentiation, among others. For example, drugs that specifically block p115RhoGEF actions could prevent the stimulatory effects of Gα12 and Gα13 on Rho signaling pathways that lead to cell growth and proliferation (Gutkind, 1998; Zhong and Neubig, 2001). Many RGS proteins that bind non-G protein signaling partners are expressed exclusively in specific brain regions (Gold et al., 1997;Grafstein-Dunn et al., 2001), making these proteins attractive targets for possible therapeutic intervention. However, in most cases, our level of understanding of these protein-protein interactions and their physiological consequences is still severely limited. Furthermore, the lack of structural data has stymied efforts thus far to identify critical contact interfaces as possible drug targets. A better understanding of how RGS proteins interact with and modulate the functions of their non-G protein binding partners remains an important research goal and warrants close attention for future drug development.
VII. Conclusion and Future Directions
We have summarized findings that demonstrate the importance of RGS proteins as key contributors to vital processes in cell biology, physiology, and disease. The realization that RGS proteins act as tightly regulated modulators and integrators of G protein signaling has served to highlight these proteins as novel drug targets for therapeutic intervention. However, many questions remain unanswered regarding the cellular roles and regulation of RGS protein as signaling molecules. Evidence indicates that RGS proteins directly bind to GPCR and other cell surface receptors. What factors contribute to this process, and what effects RGS proteins have on receptor signaling, other than as G protein inhibitors, are largely unknown. In addition, preliminary evidence indicates that some RGS proteins can act as scaffolds to assemble signaling complexes, although the contribution of RGS proteins to this process and involved mechanisms are unclear. At least 20 proteins (besides Gα subunits) have now been identified that are direct binding partners for RGS proteins, and additional binding partners almost certainly remain to be found. In most cases, we know little or nothing about the functional significance of these protein interactions. Understanding the part that RGS proteins play as direct links between G proteins and other signaling pathways, the proteins involved, and their possible contribution to disease processes are important research goals. These questions remain to be answered before we can fully understand what roles RGS proteins serve in physiology and disease processes, and how they can best be targeted for drug development.
Acknowledgments
We thank Dr. Leah Bernstein for help in reviewing the manuscript as well as the many researchers and laboratories that have contributed to our current understanding of the fundamental roles that RGS proteins play in physiology and behavior. We also thank Dr. Kathleen Young and colleagues at the CNS Disorders research group at Wyeth Pharmaceuticals for support. We acknowledge grant support that has made our research possible (National Institutes of Health Grants NINDS/R01NS37112 and NIGMS/R01GM61847).
Footnotes
Address for correspondence: John R. Hepler, Department of Pharmacology, Emory University School of Medicine, G205 Rollins Research Center, 1510 Clifton Road, Atlanta, GA 30322. E-mail:jhepler{at}emory.edu
Abbreviations
- GPCR
- G protein-coupled receptor
- GAP
- GTPase activating protein
- RGS
- regulators of G protein signaling
- RL
- RGS-like
- GRK
- G protein receptor kinase
- DEP
- disheveled Egl-10 pleckstrin
- GGL
- G protein gamma subunit-like
- DH
- dbl homology
- PH
- pleckstrin homology
- PX
- Phox
- PXA
- Phox-associated
- CCK
- cholecystokinin
- CaM
- calmodulin
- PKC
- protein kinase C
- GAIP
- G alpha-interacting protein
- GIRK
- G protein-regulated inwardly rectifying potassium channel
- GDI
- guanine nucleotide dissociation inhibitor
- GTPγS
- guanosine 5′-O-thiotriphosphate
- MAPK
- mitogen-activated protein kinase
- PLC
- phospholipase C
- PIP3
- phosphatidylinositol 3,4,5-trisphosphate
- PI3K
- phosphatidylinositol 3-kinase
- PIP2
- phosphatidylinositol 4,5-bisphosphate
- PDE
- phosphodiesterase
- EphB
- Ephrin B
- Ach
- acetylcholine
- RBD
- Rap-binding domain
- APC
- adenomatous polyposis coli
- GSK3β
- glycogen synthase kinase 3β
- GIPC
- GAIP interacting protein C terminus
- EGFR
- epidermal growth factor receptor
- IL
- interleukin
- D-AKAP
- dual specificity A kinase anchoring protein
- ERK
- extracellular signal-regulated kinase
- ANP
- atrial natriuretic peptide
- PKA
- protein kinase A
- The American Society for Pharmacology and Experimental Therapeutics
References
In this issue





{kind=link}
{kind=link}
{kind=link}
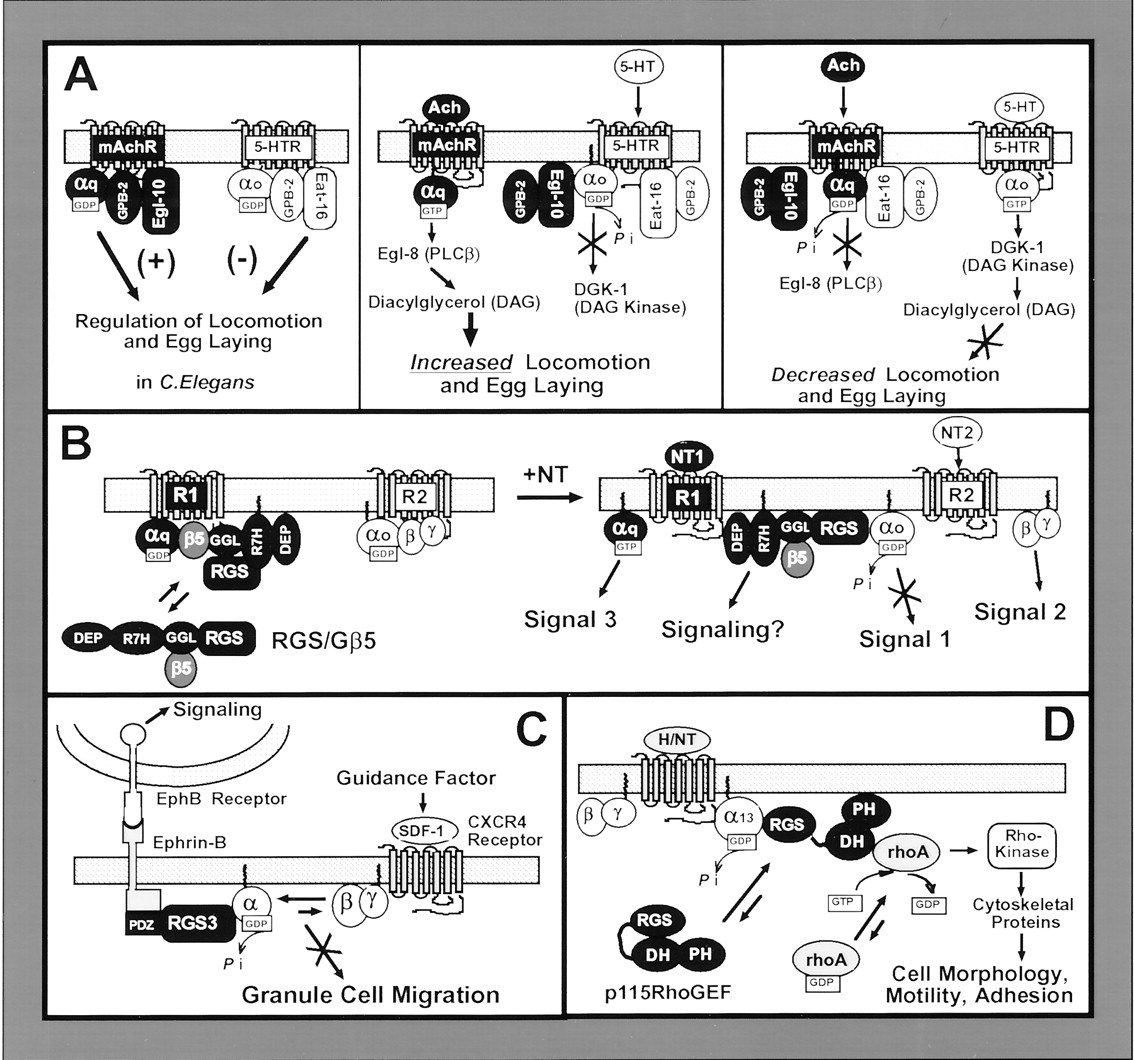{kind=link}
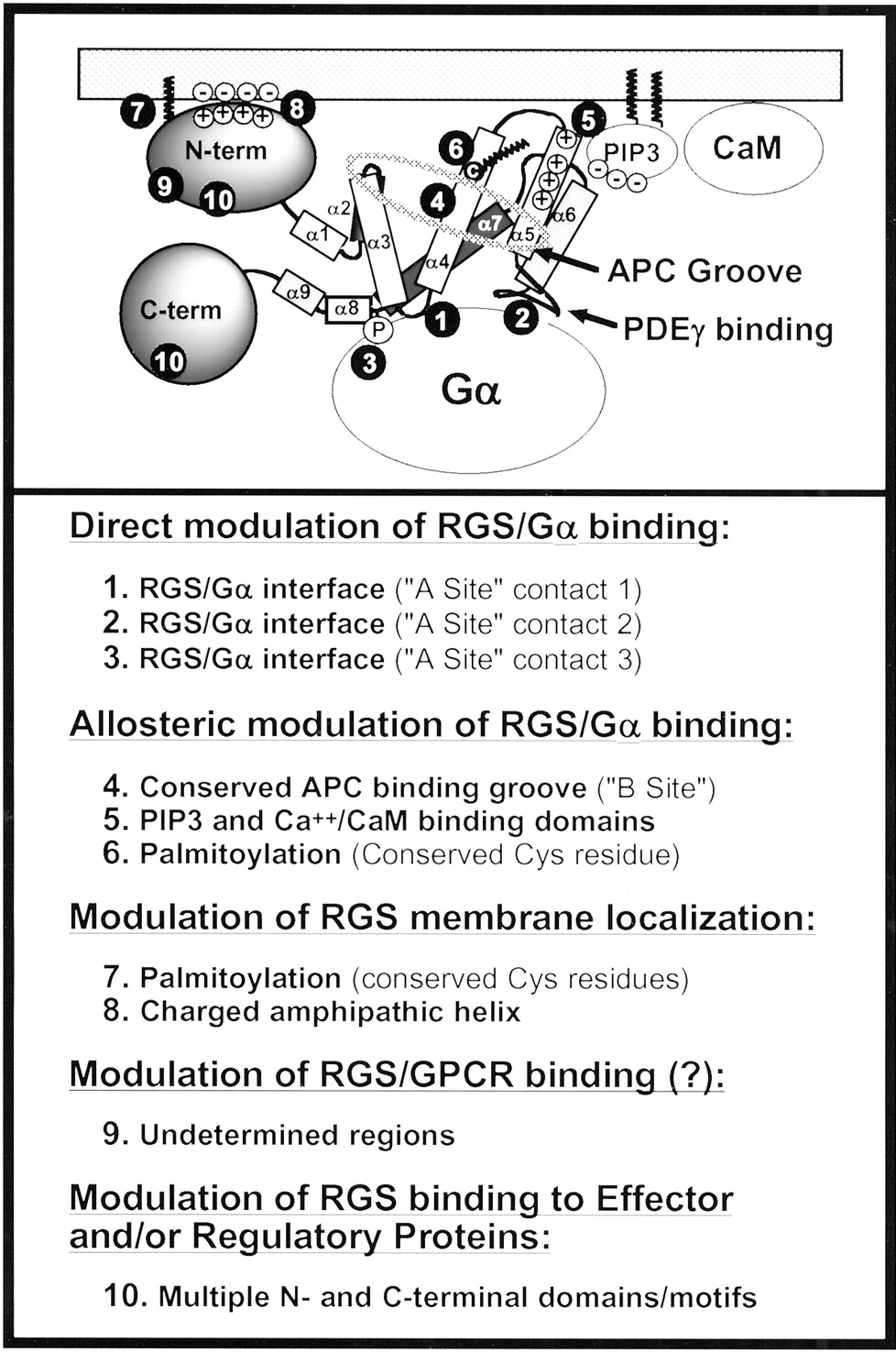{kind=link}
Jump to section
- Article
- Abstract
- I. Introduction
- II. RGS Proteins Directly Regulate G Protein Activity
- III. RGS Proteins Modulate G Protein Signaling
- IV. RGS Proteins Integrate G Protein Signals
- V. Cellular Mechanisms Regulating RGS Protein Functions
- VI. RGS Proteins As Therapeutic Targets
- VII. Conclusion and Future Directions
- Acknowledgments
- Footnotes
- Abbreviations
- References
- Figures & Data
- Info & Metrics
- eLetters