


Abstract
Neuromedin U (NmU) is a structurally highly conserved neuropeptide. It is ubiquitously distributed, with highest levels found in the gastrointestinal tract and pituitary. Originally isolated from porcine spinal cord, it has since been isolated and sequenced from several species. Amino acid alignment of NmU from different species reveals a high level of conservation, and particular features within its structure are important for bioactivity. Specifically, the C terminus, including a terminal asparagine-linked amidation, is essential for activity. The conservation of NmU across a wide range of species indicates a strong evolutionary pressure to conserve this peptide and points to its physiological significance. Despite this, the precise physiological and indeed pathophysiological roles of NmU have remained elusive. NmU was first isolated based on its ability to contract rat uterine smooth-muscle (hence the suffix “U”) and has since been implicated inthe regulation of smooth-muscle contraction, blood pressure and local blood flow, ion transport in the gut, stress responses, cancer, gastric acid secretion, pronociception, and feeding behavior. Two G-protein-coupled receptors for NmU have recently been cloned. These receptors are widespread throughout the body but have differential distributions suggesting diverse but specific roles for the receptor subtypes. Here we detail the isolation and characterization of NmU, describe the discovery, cloning, distribution, and structure of its two receptors, and outline its possible roles in both physiology and pathophysiology. Ultimately the development of receptor-specific ligands and the generation of animals in which the receptors have been selectively knocked out will hopefully reveal the true extent of the biological roles of NmU and suggest novel therapeutic indications for selective activation or blockade of either of its receptors.
I. Discovery, Isolation, Structure, and Synthesis of Neuromedin U
In the 1980s, a novel family of peptides termed “neuromedins” were isolated from porcine spinal cord based on their ability to contract smooth-muscle (Kangawa et al., 1983; Minamino et al., 1984a-c; 1985a,b). This family of peptides comprises the neuromedins B and C (bombesin-like), K (neurokinin B) and L (neurokinin A, substance K, or neurokinin α), N (neurotensin-like), and U. These neuromedins are found in many tissues including the central nervous system and have been implicated in a wide variety of physiological functions.
Purification and characterization of porcine neuromedin U (NmU1) identified two peptides with similar biological activity (Minamino et al., 1985a). Both peptides contracted strips of rat uterus in vitro (hence the suffix U), and intravenous injections caused sustained increases in systemic blood pressure in rats. The two peptides were an icosapentapeptide (NmU-25) and an octapeptide (NmU-8) that was identical to the C terminus of NmU-25 (Fig. 1). Indeed, porcine NmU-8 results from the cleavage of NmU-25 on the C-terminal side of the two arginine residues immediately preceding the last eight C-terminal amino acids of NmU-25 (Arg16 and Arg17), which form a typical peptide cleavage signal (Fisher and Scheller, 1988).

The structure of NmU isolated from different species. Sequences were obtained from original isolated peptides (see Section I.) with the exception of human NmU-25 (from NmU precursor protein, see Section I.) and mouse NmU-23 (NCBI peptide accession number Q9QXH8). Sequences were aligned with Genedoc sequence alignment editor (for review, see Nicholas et al., 1997). The highly conserved C terminus is shown in bold and the di-basic cleavage recognition site, Arg-Arg, in positions 16 and 17, is shown underlined where present. To show more clearly the structural similarities, rat NmU-23, mouse NmU-23, and tree frog skin secretion NmU-23 are elongated with gaps represented by an asterisk (*). Residues are numbered above the amino acid sequence. All versions of NmU have an amidated Asn at the C terminus resulting in -CONH2, which has been abbreviated to -NH2. Numbering of the smaller versions of NmU starts at their N termini.
Following the initial identification of porcine NmU, this peptide was purified and sequenced from rat (Conlon et al., 1988; Minamino et al., 1988), frog (Domin et al., 1989), rabbit (Kage et al., 1991), dog (O'Harte et al., 1991a), and chicken (O'Harte et al., 1991b; Domin et al., 1992). The amino acid sequences of these are shown in Fig. 1. Most of the NmU analogs that have been isolated are icosapentapeptides with the exceptions of a 23-amino acid NmU in rat (NmU-23) and nonapeptides (NmU-9) from guinea pig (Murphy et al., 1990) and chicken (O'Harte et al., 1991b). Chicken NmU-9 was the predominant molecular form isolated from the chicken intestines and was indeed the only form isolated from extracts of whole brain (O'Harte et al., 1991b). In addition, as with porcine NmU, an NmU-8 analog has also been identified in dog (O'Harte et al., 1991a) and is identical to the C terminus of dog NmU-25. Again, the C-terminal NmU-8 is preceded by an Arg-Arg motif (Fig. 1), the cleavage of which gives rise to the smaller NmU analog. The source of the nonapeptides from guinea pig and chicken is less clear. Larger NmU analogs have not been reported in guinea pig, suggesting that NmU-9 may arise independently. Similarly, avian NmU-25 lacks the di-basic Arg-Arg cleavage site, suggesting that avian NmU-9 may be produced independent of avian NmU-25 although the possibility of cleavage at a mono-basic site cannot be excluded.
The human cDNA encoding the precursor protein of NmU has been sequenced (Austin et al., 1995) revealing that human NmU is also an icosapentapeptide and has no internal di-basic cleavage site for the generation of shorter versions. Comparison with the rat cDNA encoding the precursor protein (Lo et al., 1992) demonstrates approximately 70% homology between the two and indicates that in both instances NmU is synthesized as a 174-amino acid precursor containing the NmU peptide within the C terminus. In addition to the signal peptide, these NmU precursors have several characteristics associated with the precursors of most other small regulatory peptides. In particular there are a number of paired di-basic amino acids forming putative proteolytic cleavage sites (five in rat, four in human) that, in addition to releasing NmU, may release a series of other peptides. Two of these sites surround a 33-amino acid sequence that differs by only two amino acids between rat and human. This high level of conservation suggests a biological role for this peptide, although this has not been confirmed.
There is remarkable amino acid homology between the forms of NmU isolated from different species (Fig. 1). For example, 15 of 25 amino acids are conserved between porcine NmU-25 and amphibian NmU-25. Human, rat, rabbit, avian, and amphibian NmU lack the di-basic Arg-Arg cleavage sequence in positions 16 and 17, accounting for the absence of shorter analogs in these species (see above). In particular, the C terminus of NmU displays remarkable homology between species. Thus, human NmU-25, porcine NmU-8 and NmU-25, rabbit NmU-25, and guinea pig NmU-9 have identical C-terminal octapeptides, whereas dog NmU-8 and NmU-25, rat NmU-23, frog NmU-25, and avian NmU-25 and NmU-9 have a single amino acid difference. Furthermore, across all species, the C-terminal pentapeptide (Phe-Arg-Pro-Arg-Asn-NH2) is identical, although in mammalian species, the C-terminal heptapeptide (Phe-Leu-Phe-Arg-Pro-Arg-Asn-NH2) is entirely conserved. This high degree of amino acid conservation is not only indicative of the importance of the C terminus (see Section II.), but considering that the evolutionary paths of these animals diverged millions of years ago, it also shows strong evolutionary pressure to preserve NmU. The evolution of neuropeptides has been reviewed elsewhere (Holmgren and Jensen, 2001).
Intriguingly, a novel NmU analog has been isolated and characterized from the defensive skin secretions of the Australian tree frog, Litoria caerulea (Salmon et al., 2000). This 23-amino acid peptide shows smooth-muscle contractile properties comparable with that of porcine NmU-25 and binds to uterine membrane preparations with an affinity equivalent to that of rat NmU-23. The function of NmU in such skin secretions is unclear but could provide a defensive mechanism by triggering peripheral or neurocentral effects in predators. Alternatively, it could act on the frog itself to elicit hypertensive effects following injury, thus reducing blood loss (Salmon et al., 2000). Such skin secretions have long been known as rich sources of biologically active peptides, but our understanding of their biological roles is in its infancy.
NmU appears to be unique in both structure and function, and it is difficult to relate these aspects to other peptides with any confidence. However, the C-terminal amidation is a structural feature associated with many peptides that exhibit hormonal or physiological activities and is found in both gastrointestinal hormones and hypothalamic releasing factors (Tatemoto, 1978). Conversely, an amidation linked to asparagine, as observed with NmU, has to date only been identified in vasoactive intestinal polypeptide (VIP). The amino acid sequence of VIP, 28 residues in length (Bodanszky et al., 1973; Mutt and Said, 1974), shows weak but evident structural homology with NmU. Structurally NmU-25 also shares limited homology with the 36-amino acid peptide, human pancreatic polypeptide (PP) (Boel et al., 1984), with the C-terminal sequence of -Arg-Pro-Arg-X-CONH2, existing in both. The amino acid sequences of human NmU-25, VIP, and PP have been compared with Aplysia small cardioactive peptide B (SCPB), human ghrelin, and human neurotensin (NT) (Fujii et al., 2000) and are also detailed in Fig. 2. All show low but evident homology. Despite these structural similarities, with the exception of Aplysia SCPB, which shows weak but evident agonism (Fujii et al., 2000), only human NmU-25 and related NmU analogs show any agonist effect at the NmU receptors (for details, see Section IV.).
Comparative amino acid alignment of NT, VIP, Aplysia SCPB, ghrelin, and PP, with human NmU-25. Amino acid sequences were obtained through the NCBI protein database with the following accession numbers: human NmU-25; NP_006672, VIP; NP_003372, PP; NP_002713; Aplysia SCPB; P09892, NT; NP_006174; and ghrelin; BAA89371. Residues identical to human NmU-25 are shown in bold and * denotes spaces to maximize alignment. Similar comparisons have been made previously (Fujii et al., 2000).
II. Structure-Activity Relationships of Neuromedin U
The consistency of NmU structure across species demonstrates a tight structural requirement for function. Furthermore, the very highly conserved C-terminal region of the NmU variants implies much of the activity of the peptide is located here and the absolute conservation of the last five amino acids suggests that this is essential for function. Within porcine NmU-8, replacement of either Phe2, Phe4, Arg5, Pro6, Arg7, or Asn8 with Gly or replacement of either Phe2, Phe4, Arg5, Pro6, or Asn8 with d-amino acids significantly reduces the ability of NmU to contract chicken crop smooth-muscle (Hashimoto et al., 1991).
Studies examining the ability of modified versions of dog NmU-8 to cause contraction of isolated chicken crop smooth-muscle have also probed the role of this highly conserved region of the NmU peptides. Thus, the Phe residues at positions 2 and 4 (which are conserved across the NmU peptides) are important for the ability of dog NmU-8 to contract chicken crop smooth-muscle (Kurosawa et al., 1996). In particular, the aromatic side chain of Phe at position 2 contributes significantly to activity (Kurosawa et al., 1996), whereas the Arg residue at position 7 (which is also conserved across the NmU peptides) is indispensable for receptor binding and activation (Sakura et al., 2000). These studies also demonstrate that the carboxylic acid group situated at the NH2 terminus of dog NmU-8 is a major contributory factor to bioactivity (Hashimoto et al., 1995). Interestingly, dog NmU-8 is considerably more potent than porcine NmU-8 (Sakura et al., 1995), leading to the suggestion that the pyroglutamic acid (pGlu) residue at position 1 of dog NmU-8 provides resistance to degradation by aminopeptidases. Indeed, modification of the N terminus of dog NmU-8 to give aminopeptidase resistance is associated with increased the contractile activity on chicken crop smooth-muscle (Sakura et al., 1995).
Despite the importance of the C terminus, the NH2 terminus of the longer NmU peptides greatly influences function, particularly in respect to potency. Indeed, porcine NmU-25 is approximately three to five times more potent than porcine NmU-8 in mediating hypertensive effects in the rat and causing contraction of either rat uterine smooth-muscle or chicken crop smooth-muscle in vitro (Minamino et al., 1985a; Okimura et al., 1992). Similarly in the rat, intravenous porcine NmU-25 is a more potent mediator of mesenteric vasoconstriction (see Section V.B.) than NmU-8 (Gardiner et al., 1990). Furthermore, NmU-25 causes repeat twitching of isolated uterus and increases the sensitivity to NmU, whereas NmU-8 does neither, leading to the suggestion that the NH2 termini of the longer NmU forms may be involved in strengthening and prolonging stimulant activity (Minamino et al., 1985a).
Using rat NmU-23 and a series of peptides derived from its fragmentation, amino acids 16 to 23 (the C-terminal octapeptide) were shown to be vital in mediating the contraction of chicken crop smooth-muscle and rat uterus, whereas increasing the length of the N terminus was associated with an increase in activity. In particular, the sequences Tyr-Gln-Gly-Pro corresponding to positions 6 to 9 and Ser-Gly-Gly corresponding to positions 13 to 15 of rat NmU-23 were shown to be important for high potency (Sakura et al., 1991). These sequences may act to fold the peptide, holding it in a conformation that is more favorable for activity. Similarly, fragmentation of the NH2-terminal regions of NmU-25 from rabbit, pig, and frog demonstrates that the C-terminal 11-peptide amides retain contractile activity on chicken crop smooth-muscle that is at least three times more potent than their C-terminal 8-peptide amide equivalents (Okimura et al., 1992). This suggests a functional role for the three amino acids directly extending from the C-terminal octapeptide, despite considerable heterogeneity between species. However, all are hydrophilic and immediately precede a 4-amino acid sequence rich in hydrophobic residues, namely Tyr(or Phe)-Phe-Leu(or Val)-Phe. This, in turn, precedes a hydrophilic and basic C-terminal Arg-Pro-Arg-Asn-NH2 sequence in all species, suggesting that an amphiphilic structure could be involved in the enhancement of bioactivity.
Post-translational amide modification of the C-terminal amino acid is common among neuropeptides and is essential to their bioactivity (Bradbury and Smyth, 1987; Escher et al., 1982; Eipper and Mains, 1988). Such enzymatic amidation may prevent the carboxy terminus of peptides from becoming ionized, therefore making them more hydrophobic and better able to bind to their receptors. Although peptides terminating in amidated versions of all 20 amino acids have been identified, amidated amino acids are generally neutral. Amidation of the neutral amino acid, Asn, in NmU is crucial for activity as synthetic des-amido-NmU-8, which lacks the asparagine-amide structure at the C terminus, is unable to cause uterine contraction or hypertension (Minamino et al., 1985a). Furthermore, non-amidated NmU-8 does not cause intracellular Ca2+ signaling by the recombinant human NmU receptor, GPR66/FM-3 (NmU receptor 1; NmU-R1) (for details, see Section IV.) (Hedrick et al., 2000) or recombinant murine versions of the two NmU receptors, NmU-R1 and NmU-R2 (Funes et al., 2002).
Intracerebroventricular administration of NmU reduces food intake in rat (see Section V.D.), and recent studies have demonstrated that the aromatic rings of Phe residues at positions 16, 17, and 19 of rat NmU-23 reduce the effect of NmU on food intake, whereas those on Tyr residues at positions 1 and 5 are essential for activity (Abiko and Takamura, 2002 and 2003).
In a structure-activity relationship study based on receptor activation, the individual substitution of each of the eight amino acids of porcine NmU-8 with alanine resulted in marked decreases in agonist potency on Ca2+ signaling by either recombinant murine NmU-R1 or NmU-R2 expressed in HEK293 cells, thereby demonstrating the contribution of the individual amino acids to bioactivity (Funes et al., 2002). Indeed, substitution of Arg7 resulted in a complete loss of activity. Moreover, although all the remaining Ala-substituted peptides retained full agonist properties at murine NmU-R2, substitution at Phe2, Phe4, and Arg5 resulted in the generation of partial agonists at the murine NmU-R1 (Funes et al., 2002). The selective effects of substitutions on agonist potencies at the two different receptor subtypes were most apparent with replacement of Arg5, which greatly reduced both potency and efficacy at murine NmU-R1 compared with murine NmU-R2, leading the authors to suggest that this may provide a basis for the development of receptor-specific agonists.
Potency differences between naturally occurring versions of NmU have not, however, been a consistent finding in functional assays with isolated tissues (Benito-Orfila et al., 1991; Westfall et al., 2001) or in studies using the two recently cloned G-protein-coupled receptors (GPCR) for NmU (see Section IV.). For example, NmU-8 and NmU-25 are approximately equipotent at stimulating recombinant versions of these receptors to either elevate intracellular [Ca2+] ([Ca2+]i) (Hedrick et al., 2000; Shan et al., 2000; Szekeres et al., 2000; Funes et al., 2002) or mediate the release of arachidonic acid (Fujii et al., 2000; Hosoya et al., 2000). Alternatively, porcine NmU-8 has been reported to be 2.5- to 5-fold more potent than NmU-25 in Ca2+-signaling assays using recombinant receptors (Raddatz et al., 2000). Although using rat uterine membranes, rat NmU was shown to have a 60-fold higher affinity than porcine NmU-8 (Nandha et al., 1993); direct comparison of the affinities of porcine NmU-8 and NmU-25 using the cloned receptors have not always revealed such differences. Thus, based on their ability to compete with radiolabeled NmU, the binding affinities of porcine NmU-8 and NmU-25 are either remarkably similar (Hosoya et al., 2000; Raddatz et al., 2000) or at best NmU-25 has 3-fold higher affinity than NmU-8 (Fujii et al., 2000) at the cloned human receptors. In contrast, porcine NmU-25 has been reported to have an 85-fold higher affinity than porcine NmU-8 at the cloned canine NmU-R2 expressed in HEK293 cells (Westfall et al., 2001). Thus, the contribution of any differences in affinity to potency differences in functional assays is unclear. The possibility that altered peptide stability may contribute, particularly in tissues or intact organisms, should at least be considered. For example, the longer versions of NmU could be more resistant to proteolytic cleavage of the conserved biologically active C-terminal region. Rat uterine membranes are able to degrade 125I-labeled rat NmU in a temperature-dependent manner (Nandha et al., 1993), demonstrating the susceptibility of such peptides to proteolytic cleavage. Although the issue of ligand degradation has been considered during in vitro and in vivo studies (Westfall et al., 2001; Cao et al., 2003; Yu et al., 2003) the impact of degradation on agonist potency and potential differences between the various forms of NmU has not been fully addressed.
III. Regional, Cellular, and Subcellular Distribution of Neuromedin U
Our understanding of the distribution of NmU owes much to the application of chromatographic and immunological techniques, particularly radioimmunoassay (RIA) and immunocytochemistry (ICC) using antibodies, most commonly raised against synthetic porcine NmU-8, to detect NmU-like immunoreactivity (NmULIR). More recently, detection of mRNA for NmU or its precursor molecule have also been employed. Such studies have enabled localization of NmU to areas, cell types, and subcellular structures, thereby providing detailed evidence of expression patterns. This knowledge has provided considerable support to the current ideas about the physiological roles of NmU.
NmU-LIR is widely distributed throughout the body. It must be noted that there may well be differences between species and developmental stages but these remain to be thoroughly and systematically explored. In the rat, highest levels are in the anterior pituitary and the gastrointestinal tract (Domin et al., 1987; Steel et al., 1988), whereas significant levels are also found in the brain, spinal cord, and both the male and female genito-urinary tract (Domin et al., 1986). These expression patterns of NmU-LIR have been supported by dot blot analyses or quantitative RT-PCR of mRNA for NmU or its precursor in both rat and human tissues (Fujii et al., 2000; Szekeres et al., 2000).
Circulating NmU-LIR has not been detected, (Domin et al., 1987) and this, along with its absence from endocrine cells, for example in the stomach (Augood et al., 1988) and small intestine (Domin et al., 1987), suggests that NmU acts more as a neuropeptide or neuromodulator than as a circulating hormone. However, it should be noted that a small number of NmU-LIR-positive endocrine cells in crypts of guinea pig small intestine have been reported (Furness et al., 1989).
A. Central Nervous System
RIA of tissue extracts indicates high levels of NmULIR in both the spinal cord and brain of rats with considerable regional differences (Domin et al., 1987). In the spinal cord, levels are greater in the dorsal horn than the ventral horn. This and the high concentrations in the dorsal root ganglia are consistent with a sensory role (Domin et al., 1987).
In the brain, the highest levels of NmU-LIR are within the nucleus accumbens, hypothalamus, septum, amygdala, medulla oblongata, and globus pallidus (Domin et al., 1987). Although cell bodies positive for NmU-LIR were reported only in the rostrocaudal portion of the arcuate nucleus with a much more widespread distribution of NmU-LIR-positive fibers (Ballesta et al., 1988), another detailed ICC study demonstrated that within rat brain, distribution of NmU-LIR-positive cell bodies is uneven and largely restricted to a variety of regions associated with distinct functional systems including somatosensory, motor, and auditory (Honzawa et al., 1987). Thus, immunoreactivity was present in cranial motor nuclei, reticular nuclei, nucleus vestibularis lateralis, trigeminal nuclei, colliculus superior, and inferior, lemniscus lateralis, nucleus pontis, nucleus ruber, zona incerta, substantia innominata, horizontal limb of the diagonal band, and the cerebral cortex (Honzawa et al., 1987). Furthermore, nerve fibers positive for NmU-LIR were observed in other regions including the nucleus reticularis thalami, nucleus ventralis posteromedialis, nucleus ventralis posterolateralis, nucleus tegmentalis dorsalis and ventralis, vertical limb of the diagonal band, nucleus olivaris superior, and nucleus pontis. NmU-LIR has also been demonstrated in rat, mouse, and human pituitary, particularly the anterior pituitary where it is colocalized in corticotrophs with adrenocorticotrophic hormone (ACTH) (see Section V.C.) (Ballesta et al., 1988; Domin et al., 1988; Steel et al., 1988). The selective distribution of NmU throughout the human CNS has been supported by dot blot analysis of cDNA libraries (Hedrick et al., 2000), in situ hybridization (Howard et al., 2000) and quantitative RT-PCR of the NmU precursor (Szekeres et al., 2000). This latter study indicates that high levels of NmU precursor message occur in the hypothalamus, medulla oblongata, nucleus accumbens, parahippocampal gyrus, substantia nigra, superior frontal gyrus, and thalamus with greatest levels in the cingulate gyrus, locus ceruleus, medial frontal gyrus, and pituitary (Szekeres et al., 2000). Quantitative RT-PCR has also shown regional expression of NmU mRNA in rat CNS, with by far the highest levels seen in the pituitary gland (Fujii et al., 2000). Similar methodology examining the distribution of NmU precursor mRNA in the mouse CNS demonstrated highest levels in the medulla and spinal cord with lower levels in the hypothalamus and pons (Funes et al., 2002). Using in situ hybridization, the mRNA for NmU has also been shown to be most abundant in the ventromedial hypothalamic regions (lateral arcuate nucleus and median eminence) and in the caudal brainstem (nucleus of solitary tract, area postrema, dorsal motor nucleus of the vagus, and inferior olive) of the rat (Howard et al., 2000). The presence of NmU mRNA in the arcuate nucleus and median eminence of the rat has been questioned with the authors arguing that expression is localized in the pars tuberalis (Ivanov et al., 2002). Indeed, it has been shown that NmU mRNA is abundantly expressed in rat pars tuberalis (Graham et al., 2003), a region that surrounds the hypophyseal stalk, extending along the ventral surface of the median eminence, and which may have a photoperiodic function modulating seasonal changes in pituitary activity (prolactin secretion) in some species, although its function in nonseasonal animals such as the rat is unknown (Morgan and Williams, 1996). A recent study examining the detailed anatomical localization of NmU mRNA in both the rat and mouse hypothalamus has revealed species differences (Graham et al., 2003). Thus, NmU is abundantly expressed in the dorsomedial hypothalamus (DMH) and ventromedial hypothalamus, arcuate nucleus, and suprachiasmatic nucleus (SCN) of the mouse, but expression is weaker in the rat. In contrast, expression within the pars tuberalis is more pronounced in the rat. These data suggest that NmU has both common and distinct functions in the two species. Given the lack of detailed information about NmU expression in humans, it is currently difficult to identify the most suitable animal model of NmU function in humans (Graham et al., 2003).
B. Peripheral Distribution
RIA has shown NmU-LIR to be present throughout the gastrointestinal tract of the rat, being particularly high in the small intestine, cecum, colon, and rectum with considerably lower levels in the esophagus and stomach (Domin et al., 1987). NmU-LIR is not detectable in the liver or pancreas (Domin et al., 1987). Northern blot analysis of mRNA for the NmU precursor has also demonstrated that in the gastrointestinal tract, highest levels are in the duodenum and jejunum (Austin et al., 1994). Detailed ICC studies have demonstrated that in the gut, NmU-LIR is confined to nerve cell bodies in the submucosal and myenteric plexuses and in fibers in these regions and the mucosa of all areas except the stomach (Augood et al., 1988; Ballesta et al., 1988; Bishop et al., 1988; Honzawa et al., 1990). Ultrastructural studies using immunogold labeling demonstrated that NmU-LIR within the cell bodies is localized to large (60-70 nm), electron-dense granules characteristic of those found in nerves containing peptides (Ballesta et al., 1988). Many NmU-LIR-positive fibers are present close to the mucosal epithelium, whereas the muscularis mucosa has a small number in the small intestine but not the large intestine (Ballesta et al., 1988). The intrinsic nature of the nerves containing NmU-LIR is supported by the observation that extrinsic denervation does not affect the number or distribution of NmU-LIR-positive fibers (Ballesta et al., 1988; Bishop et al., 1988). Similarly in the gastrointestinal tract of the guinea pig, NmU-LIR is within nerve cells of the myenteric and submucous plexuses (Furness et al., 1989). It is also present in nerve fibers of these plexuses (Augood et al., 1988; Furness et al., 1989), around submucous arterioles and in the mucosa (Furness et al., 1989), but there are few or no fibers supplying the muscle layers (Augood et al., 1988; Furness et al., 1989). A study of the distribution of nerve cell bodies containing NmU-LIR in the submucosal nerve plexus of the pig demonstrated considerably higher levels in the internal submucosal plexus (Meissner) compared with the external submucosal plexus (Schabadasch), consistent with the projection of the nerve fibers from the submucosal plexus predominantly to the muscularis mucosa and lamina propria of the outer mucosal layer (Timmermans et al., 1989, 1990). These studies in the pig also demonstrated that NmU-LIR can coexist with neurons positive for calcitonin gene-related peptide or substance P immunoreactivity. Similarly, in rat small intestine, NmULIR has been shown to coexist with calcitonin gene-related peptide (Ballesta et al., 1988; Bishop et al., 1988), whereas in the submucosal ganglia of guinea pig small intestine, all cells that were immunoreactive for either vasoactive intestinal peptide, neuropeptide Y, or substance P also contained NmU-LIR (Furness et al., 1989). These peptides define specific populations of neurons within the submucosal plexus, and this therefore places NmU-LIR in noncholinergic secretomotor neurons, cholinergic secretomotor neurons, and sensory neurons (Furness et al., 1989). This suggests multiple roles for NmU in the gut.
Although levels are not as high as within the gastrointestinal tract, RIA reveals significant amounts of NmU-LIR in most of the genitourinary tract of the rat, with highest levels in the ureter, vas deferens, prostate, fallopian tubes, and urethra, although it is not present in seminal vesicles (Domin et al., 1987). Quantitative RT-PCR of NmU mRNA also supports expression in the testis and ovary of rat (Fujii et al., 2000). NmU mRNA (Fujii et al., 2000) and NmU-LIR (Domin et al., 1990) also exists in the rat thyroid gland where immunoreactivity is localized to a minor population of parafollicular C-cells.
Quantitative RT-PCR of the NmU precursor has also shown levels to be high in human stomach, intestine, and bone marrow with some expression in a variety of other tissues including spleen, lymphocytes, adipose tissue, prostate, and placenta (Szekeres et al., 2000). Similar studies have also determined distinct patterns of NmU expression among different lymphoid cells (Hedrick et al., 2000). There appears to be some developmental regulation of NmU expression given that adult liver and lung express little or no NmU (or precursor) mRNA, whereas levels in fetal tissues are high (Hedrick et al., 2000; Szekeres et al., 2000).
IV. Receptors for Neuromedin U
A. Discovery
Binding sites for NmU were characterized well before the molecular nature of the receptor had been identified. Early studies demonstrated that the binding of 125I-labeled rat NmU to membranes prepared from rat uterus was saturable, specific, reversible, and dependent on time, temperature, and pH (Nandha et al., 1993). Scatchard analysis suggested a single class of binding site with a Kd of 0.35 nM. Furthermore, guanosine 5′-[γ-thio]triphosphate (GTPγS) reduced the binding implying a GPCR. Despite such studies, it was not until molecular biology had established itself alongside classical pharmacology that NmU receptors were cloned and their molecular nature determined.
In 1998, a GPCR (FM-3) was cloned from human and murine cDNA libraries due to its homology with both the growth hormone secretagogue receptor [subsequently identified as the receptor for ghrelin (Kojima et al., 1999)] and the neurotensin receptor (33 and 29% DNA sequence homology, respectively), which were used as tags to screen a genetic database (Tan et al., 1998). This cloning enabled a subsequent “reverse-pharmacological” approach to the identification of the ligand for the receptor, in which recombinantly expressed FM-3 was screened with putative ligands. This approach led directly to the identification of NmU as a ligand for the orphan human GPCR, FM-3 (or GPR66) (Fujii et al., 2000; Hedrick et al., 2000; Hosoya et al., 2000; Howard et al., 2000; Raddatz et al., 2000; Shan et al., 2000; Szekeres et al., 2000). NmU binds to and activates human FM-3 (hFM-3) with subnanomolar affinity and potency, respectively. Extracts of rat tissue were also found to contain natural ligands for FM-3 based on the ability to elevate [Ca2+]i in CHO cells expressing the recombinant receptor (Kojima et al., 2000). Extracts from brain and small intestine were the most active, and the endogenous ligand was purified chromatographically to a 23-amino acid peptide that was identical to rat NmU-23.
Using the ability of activated hFM-3 to elevate [Ca2+]i and increase inositol phosphate accumulation, over 1500 proposed GPCR ligands, including many peptides, have been screened as potential ligands. The various forms of NmU are the only peptides that potently activate the receptor (Fujii et al., 2000; Hedrick et al., 2000; Hosoya et al., 2000; Howard et al., 2000; Kojima et al., 2000; Raddatz et al., 2000; Shan et al., 2000; Szekeres et al., 2000). Peptides that show some similarity to NmU, such as the neuromedins B, C, K, and N, as well as neurotensin, ghrelin, motilin, vasoactive intestinal peptide, and pancreatic polypeptide are inactive. However, Aplysia SCPB shows a very low but significant ability to elevate [Ca2+]i in CHO cells expressing recombinant hFM-3 (Fujii et al., 2000). This peptide shares a conserved motif of Leu-X-X-Pro-Arg-X-amide with NmU (Fig. 2), suggesting the importance of this sequence in the interaction of NmU with its receptor. However, this motif is also present in pancreatic polypeptide (see Fig. 2), but this is not an agonist of hFM-3, suggesting its longer NH2 portion may interfere with receptor binding.
At the time of identification of hFM-3 as the NmU receptor, a second NmU receptor was identified from human and rat (Hosoya et al., 2000; Howard et al., 2000; Raddatz et al., 2000; Shan et al., 2000). The murine orthologues of the receptors have also been identified and cloned (Howard et al., 2000; Funes et al., 2002). Given the identification of two receptors for NmU, the first (FM-3/GPR66) has been designated NmU-R1 and the second NmU-R2.
Although there are differences in the structure of the NmU peptide between species, there is a very high conservation of the C terminus that is largely responsible for its biological activity (see Section III.). Furthermore, neither human (Fujii et al., 2000; Hedrick et al., 2000; Hosoya et al., 2000; Howard et al., 2000; Raddatz et al., 2000; Shan et al., 2000; Szekeres et al., 2000) nor murine (Funes et al., 2002) NmU receptors have significant abilities to distinguish between different forms of NmU. Despite this, there are apparent species and tissue selectivity in the biological actions of NmU. For example, both forms of porcine NmU stimulate contraction of uterine muscle from the rat but not the guinea pig (Minamino et al., 1985a). This suggests that differences in the distribution and/or function of NmU receptors between species contributes to functional diversity.
B. Characterization of Neuromedin U-R1 and Neuromedin U-R2
The genes for human NmU-R1 (hNmU-R1) and hNmU-R2 have been mapped respectively to SHGC-33253 that is localized to chromosome 2q34-q37 and SHGC-8848 that is localized to chromosome 5q31.1-q31.3 (Raddatz et al., 2000). Their genomic structures differ in that the predicted open reading frame of hNmU-R1 is encoded on two exons, whereas that of hNmU-R2 exists on four (Shan et al., 2000). There is, however, a level of homology between the two genes and conservation of certain exon-intron boundaries suggesting that the two receptors may be the products of a duplication event (Shan et al., 2000). Interestingly, several expressed sequence tags from GenBank have been identified that have identity with NmU-R2, and three of these may represent an alternative transcript (Shan et al., 2000). Although this may represent a sterile transcript, its potential presence should urge caution in the interpretation of receptor expression profiles based on mRNA expression, particularly if it is unclear whether the probes used are able to hybridize to the alternate transcript (Shan et al., 2000).
When a new gene is cloned it is not always a simple task to identify conclusively the methionine residue used to initiate translation. Indeed, for both human NmU receptors, more than one N-terminal variant of the protein sequence has been published (e.g., Raddatz et al., 2000; Shan et al., 2000; Tan et al., 1998). Although all variants of the receptors published to date are functionally active, it is informative to consider which form of the receptor is most likely to exist physiologically.
For hNmU-R1, the first group to identify this receptor reported the cloning of a 403-amino acid protein (Tan et al., 1998), whereas a subsequent publication extended the putative N terminus of the receptor by 23 amino acids, suggesting translation initiation from an inframe, upstream AUG (Raddatz et al., 2000). Analysis of the nucleotide sequences at the two putative translation initiation sites reveals that the shorter 403-amino acid form seems to have a stronger Kozak sequence (Kozak, 1986). In addition, comparison with the mouse orthologue shows that the murine receptor agrees with the shorter form of the human receptor. Thus, for hNmU-R1 the evidence for the shorter form of the receptor is stronger, although it is not inconceivable that initiation at both human AUG codons may occur, producing two proteins from one mRNA, as reported previously for other proteins (for review, see Kozak, 1991a,b).
Similarly, two forms of hNmU-R2 have been reported that differ in their initiating methionine, a 415-amino acid form (Raddatz et al., 2000; Shan et al., 2000) and a 412-amino acid form (Hosoya et al., 2000; Howard et al., 2000). Again, analysis of the nucleotide sequences surrounding the two putative translation initiation sites and comparison with rodent orthologue sequences provides stronger evidence for the slightly shorter form.
The two receptors are typical GPCRs possessing seven putative trans-membrane domains, as represented in Figs. 3 and 4. A comparative amino acid sequence alignment of both receptors is also shown in Fig. 4. The hNmU-R1 consists of 403 amino acids and has a predicted molecular mass of 44,979 Da; hNmU-R2 is slightly longer at 412 amino acids and has a predicted molecular mass of 47,450 Da. The two receptors show about 45 to 50% amino acid homology with each other, and both show some homology to the human growth hormone secretagogue receptor, the human motilin receptor, and human neurotensin receptor, with identities of about 30%. Single nucleotide polymorphisms (SNPs) have been identified, particularly in the noncoding regions of both hNmU-R1 and hNmU-R2 [National Center for Biotechnology Information (NCBI) Database]. Additionally, five SNPs have been reported that result in a change in the amino acid sequence of hNmU-R2. Based on the sequence presented in Fig. 4, these are S295T, F312L, P380L, M385V, and T392A. The frequency and functional significance of these SNPs are, however, unknown.
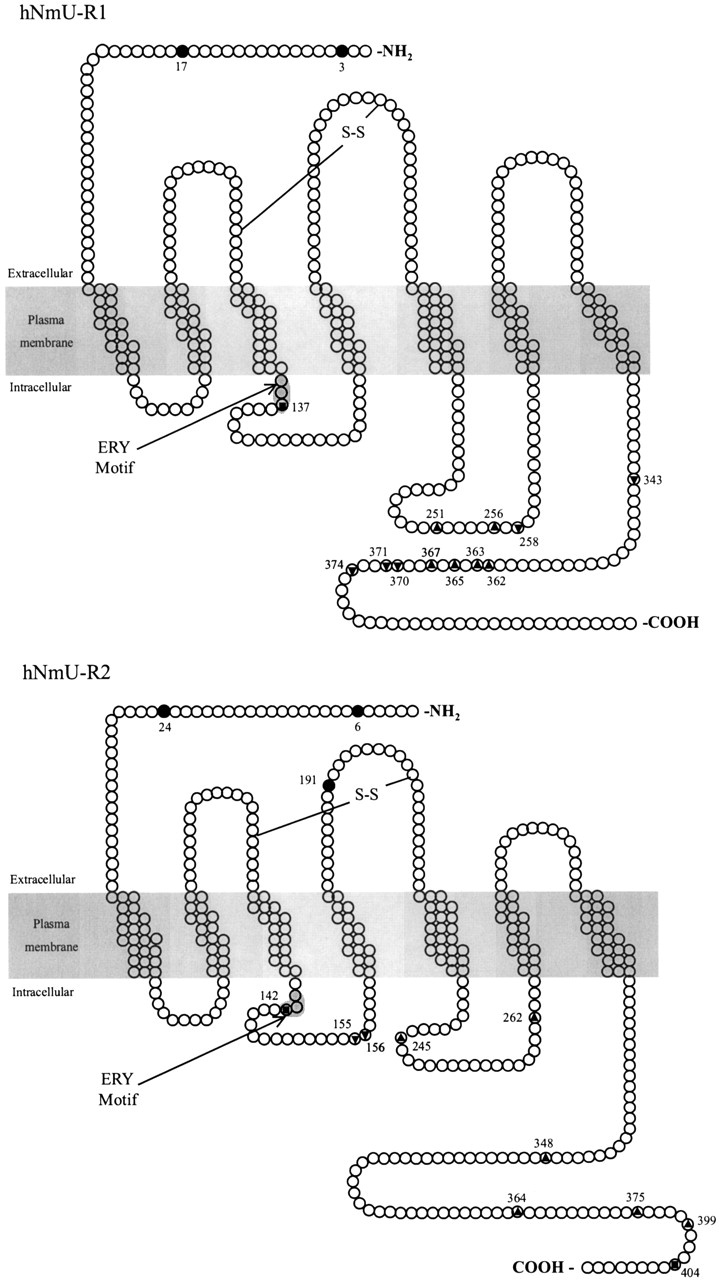
Schematic representation of hNmU-R1 and hNmU-R2. Sequences were acquired with NCBI accession numbers AF272362 (hNmU-R1) and AF272363 (hNmU-R2). Prediction of transmembrane spanning helices was carried out using HMMTOP (v.2.0) (Tusnády and Simon, 2001), which uses the Hidden Markov model (Tusnády and Simon, 1998) for predicting transmembrane helices. Identification of N-linked glycosylation sites (•) within extracellular domains and the identification of residues most likely to be phosphorylated (serine residues (▴), tyrosine residues (▾), and threonine residues (▪)) within internal domains were accomplished using ExPASy on-line proteomics tools (Blom et al., 1999). The positions of these residues within the protein that could be subject to post-translational modification are provided. Conserved cysteine residues that may be involved in the formation of disulphide bridges and the position of the ERY motif at the boundary of TM 3 and the second intracellular loop are also illustrated. Further details are given in Fig. 4.

Sequence alignment of amino acids for hNmU-R1 (top) and hNmU-R2 (bottom). Full sequences were obtained with accession numbers AF272362 (NmU-R1) and AF272363 (NmU-R2) and aligned with Genedoc sequence alignment editor (for review, see Nicholas et al., 1997). Spaces, as represented by dashes (-), indicate gaps to maximize alignment, with shaded areas highlighting conserved residues. The putative transmembrane spanning regions were determined on-line using HMMTOP (v.2.0) (Tusnády and Simon, 2001), which uses the Hidden Markov model for predicting transmembrane helices (Tusnády and Simon, 1998). These are indicated by lines above and below the sequence for NmU-R1 and NmU-R2, respectively. Symbols above the sequence are representative of residues with potential for post-translational modification for NmU-R1, and those below, for NmU-R2. These were determined using ExPASy online proteomics tools using NeTNGlyc (Gupta and Brunak, 2002) to identify N-linked glycosylation (▪) and NetPhos (Blom et al., 1999) to identify potential serine, threonine, and tyrosine-linked phosphorylation sites. Only those residues with consensus sequences for phosphorylation by specific kinases (Pearson and Kemp, 1991; Kennelly and Krebs, 1991) are detailed here and include protein kinase C (▴), protein kinase A (•), casein kinase I (⋄), casein kinase II (○). Some residues highlighted in Fig. 3 as potential phosphorylation sites do not have consensus sequences for specific kinase activity. These have not been detailed here.
In comparison with hNmU-R1, the third intracellular loop is shortened in hNmU-R2, as is the N terminus. Furthermore, the C terminus of hNmU-R1 is 67 residues in length and is considerably shorter than the 92 residues of hNmU-R2. Conservation is highest within the proposed transmembrane domains, whereas the N and C termini show little homology. Both receptor subtypes possess many of the characteristics linking them to GPCR family 1, such as the ERY variant of the DRY motif at the boundary of transmembrane domain 3 and the second intracellular loop and the conservation of cysteine residues in extracellular loops 2 and 3 (see Figs. 3 and 4). The D/ERY motif is a highly conserved triplet amino acid sequence involved in the modulation of ligand binding (e.g., Rhee et al., 2000), the preservation of inactive receptors (e.g., Barak et al., 2001; Wilbanks et al., 2002), and G-protein-coupling (Strader et al., 1994; Scheer et al., 2000). The conserved cysteine residues exist in similar positions to those within other Family 1 GPCRs where they form a disulfide bridge (Strader et al., 1994; Perlman et al., 1995) that may be critical for the stability of receptor conformation, protein folding, and ligand binding (e.g., Savarese et al., 1992). Furthermore, consensus sites for serine-, threonine-, and tyrosine-linked phosphorylation and asparagine (N)-linked glycosylation, as depicted in Figs. 3 and 4, exist throughout the receptors. Within intracellular domains, consensus recognition sites (Kennelly and Krebs, 1991; Pearson and Kemp, 1991) exist for several kinases including protein kinase A, protein kinase C, and casein kinases I and II. These predominate within the C-terminal tail and the third intracellular loop of both receptors, as well as within the second intracellular loop of hNmU-R2. Both receptor subtypes have two potential asparagine-linked glycosylation sites within the NH2 terminus, but in addition, hNmU-R2 possesses a third within the second extracellular loop. Potential sites for phosphorylation by G-protein receptor kinases (Pitcher et al., 1998) are not shown due to lack of information concerning consensus sequences.
C. Tissue Distribution of Neuromedin U-R1 and Neuromedin U-R2
Despite many studies on the distribution of human NmU receptors, an absolutely clear picture has yet to emerge. Different techniques have been applied to determine the pattern of receptor expression, and this could account for some discrepancies. Dot blots, Northern blots, quantitative RT-PCR, and in situ hybridization have been popular, but these address the mRNA signal and may not reflect protein expression. To date, distribution of NmU-Rs has not been investigated at the level of the protein due to the lack of suitable antibodies. Species differences could also contribute to the somewhat ambiguous picture concerning the sites of receptor expression. Despite these difficulties, a pattern has emerged that suggests that hNmU-R1 is expressed predominantly in the periphery, especially the gastrointestinal tract, whereas hNmU-R2 expression is predominantly in the CNS.
The mRNA for hNmU-R1 is expressed in a wide variety of tissues but levels are greatest in peripheral tissues, particularly the small intestines and stomach (Hedrick et al., 2000; Howard et al., 2000; Raddatz et al., 2000; Szekeres et al., 2000; Westfall et al., 2001). It is also present in the pancreas, adrenal cortex, heart, lung, trachea, mammary gland, bone marrow, peripheral blood leukocytes (particularly T cells and NK cells), genitourinary system, placenta, mammary gland, spleen, and adipose tissue (Hedrick et al., 2000; Howard et al., 2000; Raddatz et al., 2000; Szekeres et al., 2000; Westfall et al., 2001). A similar distribution pattern of NmU-R1 mRNA exists in rat, with highest levels in the small intestine, lung, and femur (Fujii et al., 2000). NmU-R1 mRNA is also expressed in approximately 25% of the small/medium diameter neurons within the dorsal root ganglia (Yu et al., 2003).
Although several studies examining mRNA expression have failed to demonstrate hNmU-R1 in human (Hedrick et al., 2000; Howard et al., 2000) and mouse (Funes et al., 2002) brain, others have shown low levels, but with a widespread distribution within the CNS (Raddatz et al., 2000; Szekeres et al., 2000). Highest levels of hNmU-R1 mRNA expression are found in the cerebellum, dorsal root ganglion, hippocampus, and spinal cord, albeit at levels 5- to 25-fold less than the peripheral organs.
In humans, NmU-R2 mRNA is confined predominantly to specific regions within the brain, with greatest expression observed in the substantia nigra (Howard et al., 2000), medulla oblongata, pontine reticular formation (Raddatz et al., 2000), spinal cord (Shan et al., 2000), and thalamus (Howard et al., 2000; Raddatz et al., 2000). Moderate to high levels are also observed in the hippocampus, hypothalamus, and cerebral cortex (Raddatz et al., 2000). The peripheral distribution of hNmU-R2 mRNA is a little unclear. High levels of NmU-R2 mRNA are present in testes (Hosoya et al., 2000; Raddatz et al., 2000; Westfall et al., 2001), whereas lower levels are present in a variety of peripheral tissues including the gastrointestinal tract, genitourinary tract, liver, pancreas, adrenal gland, thyroid gland, lung, trachea, spleen, thymus, and thyroid (Shan et al., 2000; Raddatz et al., 2000; Westfall et al., 2001). Others have failed to detect NmU-R2 mRNA in liver, heart, skeletal muscle, intestines, pancreas, placenta, and kidneys (Howard et al., 2000) or bladder (Westfall et al., 2001).
These reports are consistent with NmU-R2 mRNA distribution within mouse CNS (Funes et al., 2002) and rat CNS where expression is detected in the hypothalamus, hippocampus, and spinal cord (Hosoya et al., 2000; Howard et al., 2000). More specific localization in rats suggests that NmU-R2 is expressed within the paraventricular nucleus (PVN), the wall of the third ventricle in the hypothalamus, and in the CA1 region of the hippocampus (Howard et al., 2000). A detailed study using in situ hybridization to determine the localization of NmU-R2 mRNA expression in the hypothalamus of mouse and rat revealed some species differences (Graham et al., 2003). Thus, in rats, expression is predominantly in the ependymal layer of the third ventricle and PVN. Using a more sensitive technique, some expression was also noted in the arcuate nucleus. In the mouse, distribution is less localized, with stronger expression in the arcuate nucleus and also in clusters of neurons in the DMH and surrounding the ventromedial nucleus (Graham et al., 2003). Receptor autoradiographic studies have also localized NmU binding sites in both lamina I and the outer part of lamina II of the spinal cord dorsal horn, and the expression profiling of mRNA indicates that binding is most likely attributable to NmU-R2 (Yu et al., 2003). Based on the tissue distribution of mRNA, the greatest level of NmU-R2 expression in the rat is in the uterus (Hosoya et al., 2000), consistent with the role of NmU in uterine contraction in this species. The lower levels or absence of expression in human (Raddatz et al., 2000; Shan et al., 2000; Westfall et al., 2001) and dog (Westfall et al., 2001) uterus may reflect differences between species and/or physiology.
D. Regulation of Cellular Signaling Pathways by Neuromedin U
Observations regarding the intracellular signal transduction pathways regulated by NmU receptors have, to date, been entirely based around the activity of these receptors when expressed as recombinant proteins. It is clear from the functional screening assays that NmU-R1 and NmU-R2 of human and rodent origin are able to mediate intracellular Ca2+ signaling with potency in the nanomolar range (Fujii et al., 2000; Hedrick et al., 2000; Hosoya et al., 2000; Howard et al., 2000; Kojima et al., 2000; Raddatz et al., 2000; Shan et al., 2000; Szekeres et al., 2000; Funes et al., 2002). For hNmU-R1, this has been shown to be associated with phosphoinositide hydrolysis (Raddatz et al., 2000; Szekeres et al., 2000). Given the lack of effect of pertussis toxin on Ca2+ signaling by either hNmU-R1 or hNmU-R2 (Raddatz et al., 2000; Shan et al., 2000; Szekeres et al., 2000), this indicates a Gαq/11-mediated activation of phospholipase C (PLC) and a lack of PLC activation by βγ-subunits derived from Gαi. Activation of either hNmU-R1 or hNmU-R2 is also able to stimulate the release of arachidonic acid, most likely through a Ca2+-dependent activation of phospholipase A2 (Fujii et al., 2000; Hosoya et al., 2000). We have recently confirmed rapid and robust generation of inositol 1,4,5-trisphosphate by hNmU-R1 and hNmU-R2 and demonstrated that activation of PLC by either receptor is independent of Gαi (Brighton et al., 2003). Additionally, we have shown that Ca2+ responses depend upon a thapsigargin-sensitive intracellular Ca2+ store (Brighton et al., 2003).
Coupling of these receptors to signal transduction pathways other than Ca2+ and phosphoinositide signaling is less clear and could also depend on, for example, the experimental protocol, expression levels of the receptors, and the cellular background. Thus, although activation of hNmU-R2 in CHO cells results in a partial inhibition of forskolin-stimulated cAMP accumulation (Hosoya et al., 2000), activation of transiently expressed hNmU-R1 in HEK293 cells has no affect on either the basal or forskolin-stimulated levels of cAMP. Recently, we have demonstrated in HEK293 cells with stable receptor expression that activation of either hNmU-R1 or hNmU-R2 inhibits forskolin-stimulated cAMP accumulation slightly more potently than they stimulate phosphoinositide hydrolysis (approximately 0.1 nM and 1 nM EC50, respectively) (Brighton et al., 2003). Furthermore, using the immunoprecipitation of specific G-protein α-subunits from cell membranes incubated with [35S]GTPγS (Akam et al., 2001), we have shown coupling of the human NmU receptors to both Gαq/11 and Gαi, but not to Gαs (Brighton et al., 2003). As with many GPCRs (Luttrell et al., 1999), activation of either receptor is also able to activate the mitogen-activated protein kinase signaling pathway, specifically extracellular signal-regulated protein kinase 1 (ERK-1) and ERK-2 (Brighton et al., 2003), although the specific transduction pathways involved have not been established.
Using brain slices from rat, it has recently been shown that NmU depolarizes a subpopulation of hypothalamic paraventricular neurons by enhancing IH channel activity (Qiu et al., 2003). This regulation may be through a Ca2+-dependent activation of soluble guanylate cyclase, an increase in cyclic guanosine monophophate, and a subsequent activation of IH through the binding of cyclic guanosine monophosphate to the C terminus of the channel. NmU-R2 mRNA is expressed within the PVN, and the authors suggest that regulation of IH by NmU may regulate the activation of autonomic output from the brain and spinal cord, thereby influencing cardiovascular function (see Section V.B.).
V. Physiological Roles of Neuromedin U
Despite the variety of activities that have been reported, the precise physiological roles of NmU have yet to be clearly defined. A number of other diverse roles, as well as smooth-muscle contraction, have been documented. All evidence to date points to multiple roles, and indeed the wide distribution of NmU (see Section III.) and the NmU receptors (see Section IV.B.) supports this.
A. Smooth-Muscle Contraction
The smooth-muscle contractile properties of NmU are well documented in a variety of tissues and from a variety of different species. The contractile properties of both porcine NmU-8 and NmU-25 seen on rat uterine preparations (Minamino et al., 1985a) have, however, not been a consistent finding in other species, for example, in the dog (Westfall et al., 2001). Similarly, contractile effects in other tissues are species-specific. For example, a variety of NmU analogs do not mediate contraction of guinea pig or rat small intestine (Minamino et al., 1985a; Bockman et al., 1989); porcine jejunum (Brown and Quito, 1988); circular smooth-muscle of frog stomach; bladder from rat, guinea pig, ferret, rabbit, or mouse (Westfall et al., 2001); or the small and large intestinal longitudinal smooth-muscle of rat and frog (Benito-Orfila et al., 1991). NmU mediates a concentration-dependent contraction of turtle small intestine leading to the suggestion that it may regulate gut motility in reptiles but not mammals where other peptides may be more important (Bockman et al., 1989). However, NmU mediates a concentration-dependent contraction of rat circular smooth-muscle from the fundus in a manner that is direct, involving neither cholinergic activation nor nervous stimulation (Benito-Orfila et al., 1991). These direct motor actions have been reported elsewhere. For example, NmU contracts human smooth-muscle from the ileum, particularly the longitudinal muscle, and this is resistant to both atropine and tetrodotoxin (Maggi et al., 1990). NmU also contracts human urinary bladder in this way (Maggi et al., 1990). NmU-mediated contraction of canine bladder (both in vitro and in vivo), stomach, ileum, and colon has also been shown (Westfall et al., 2001). Despite the relative or absolute lack of NmU in the muscularis externa of the gastrointestinal tract (see Section III.), some of these functional responses suggest a regulatory role for NmU in the gastrointestinal tract.
B. Blood Pressure and Regional Blood Flow
At the time of the identification of NmU, intravenous administration was demonstrated to cause a rapid and sustained increase in arterial blood pressure in rats (Minamino et al., 1985a). A more recent study has supported an effect on blood pressure but found it to be relatively small and transient (Chu et al., 2002). In anesthetized dogs, intravenous administration of NmU also caused a very slight and transient increase in blood pressure (Westfall et al., 2001). In contrast, the intracerebroventricular administration of NmU to conscious, freely moving rats caused rapid, progressive, and concentration-dependent increases in mean arterial pressure that were long-lasting at high concentrations (Chu et al., 2002). Furthermore, NmU increased heart rate, while a high concentration also increased plasma norepinephrine concentration. These data indicate that NmU is able to increase sympathetic activity consistent with its localization in the PVN of the hypothalamus (Howard et al., 2000) and the projection of PVN neurons to sympathetic preganglionic neurons in the spinal cord. Note also that NmU depolarizes parvocellular cells of the PVN in hypothalamic slices, most likely through a direct postsynaptic action (Qiu et al., 2003). However, it is unclear if all of the cardiovascular actions of centrally administered NmU are mediated via this route. Indeed, intravenous NmU mediates potent constrictor effects on the superior mesenteric vascular bed in conscious rats, and this can occur independently of changes in systemic arterial pressure, heart rate, and renal or hindquarters blood flow, suggesting local rather than central effects (Gardiner et al., 1990). Similarly, intravenous NmU reduced blood flow in the superior mesenteric artery and the portal vein of anesthetized dogs with little or no effect on axillary artery and pancreatic tissue blood flow or systemic arterial pressure (Sumi et al., 1987). Thus, NmU may be important within the intestines as a regulator of local blood flow, at least in dog and rat. Whether this phenomenon is true of other species remains to be established.
C. Hypothalamo-Pituitary-Adrenal Axis and the Stress Response
NmU-R2 mRNA is expressed in the hypothalamic PVN (see Section IV.C.), which in addition to its role in feeding behavior (see Section V.D.), is responsible particularly for the release of corticotrophin releasing hormone (CRH). CRH is released following stimuli such as stress and subsequently stimulates the release of ACTH from the anterior pituitary. In turn, ACTH stimulates cortisol production by the adrenal cortex and thus the PVN is key in the control of the hypothalamo-pituitary-adrenal axis. Subcutaneous injection or intracerebroventricular administration of NmU to rats increases plasma concentrations of ACTH and corticosterone (Malendowicz et al., 1993; Ozaki et al., 2002). More specifically, administration of rat NmU-23 into the PVN increases the blood concentrations of both ACTH and corticosterone (Wren et al., 2002). These observations are consistent with a role of NmU in the hypothalamic-pituitary axis, although it has been noted that altered organ blood flow (see Section V.B.) also has the potential to influence the metabolic clearance of plasma hormones (Malendowicz et al., 1993). In support of a central action of NmU on hypothalamic PVN function, NmU stimulates the release of CRH from hypothalamic explants in vitro (Wren et al., 2002). Activation of central neurons by NmU is also indicated by the enhanced expression of c-Fos immunoreactivity or its mRNA in hypothalamic areas of the rat including the PVN (magnocellular and parvocellular divisions), supraoptic nucleus, arcuate nucleus, dorsomedial nucleus, and lateral hypothalamic area following intracerebroventricular administration of NmU (Niimi et al., 2001; Ivanov et al., 2002; Ozaki et al., 2002). Taken together, the above observations suggest central effects of NmU on the hypothalamo-pituitary-adrenal axis and hint at other functions for NmU. Indeed, arginine vasopressin and oxytocin are also synthesized in the PVN (and supraoptic nucleus), and the intracerebroventricular administration of NmU to rats enhances c-Fos expression in neurons containing these peptides (Niimi et al., 2001) while plasma levels of both are increased (Ozaki et al., 2002). Furthermore, NmU stimulates the release of arginine vasopressin from hypothalamic explants (Wren et al., 2002). However, the physiological consequences of these effects remain to be established (see also Section V.D.). Based on studies in which NmU was delivered intracerebroventricularly to ovariectomized rats, it has also been argued that NmU inhibits the release of luteinizing hormone through a CRH-mediated inhibition of pulsatile gonadotropin-releasing hormone release from pituitary gonadotropes (Quan et al., 2003).
In addition to the central effect of NmU on the hypothalamo-pituitary-adrenal axis, in vitro studies have demonstrated that NmU can enhance the secretion of steroids from rat adrenal cortex in a manner dependent upon adrenal medullary chromaffin cells (Malendowicz et al., 1994a,b). This is blocked by inhibitors of either ACTH or CRH, leading to the suggestion that a local ACTH/CRH system may be involved (Malendowicz et al., 1994a). Long-term (6-day) treatment of rats with NmU administered subcutaneously also influences the adrenal cortex, with low concentrations directly stimulating its function and growth, whereas high concentrations are inhibitory (Malendowicz et al., 1994a).
The direct effects of NmU on the adrenal cortex and its effects within the hypothalamus suggest a role for NmU in the physiological responses to stress. In addition to the impact of NmU on adrenal function in rats, intracerebroventricular administration in rats or mice induces stress-related behaviors (Hanada et al., 2001). These behavioral changes are blocked by either a CRH antagonist or anti-CRH IgG, and NmU does not cause locomotor activity in CRH knockout mice (Hanada et al., 2001). CRH neurons involved in the stress response are located in both the central nucleus of the amygdala and the PVN (Wren et al., 2002). However, NmU stimulates the secretion of CRH from hypothalamic explants in vitro and the administration of NmU into the PVN has acute effects on stress-related behaviors in rats (Wren et al., 2002). Thus, the actions of NmU on arousal and stress-related behaviors are likely to be via CRH neurons within the PVN (Wren et al., 2002).
D. Feeding and Energy Homeostasis
Since the first report that a single intracerebroventricular injection of NmU decreases food intake and feeding-associated behavior in the rat (Howard et al., 2000), the ability of centrally administered NmU to decrease food intake and body weight has been demonstrated in several other studies (Kojima et al., 2000; Nakazato et al., 2000; Niimi et al., 2001; Ivanov et al., 2002; Hanada et al., 2003). Intracerebroventricular injection of NmU antiserum into rats also increases food intake (Kojima et al., 2000), whereas fasting reduces levels of NmU in the ventromedial hypothalamus (Howard et al., 2000). Thus, central effects of NmU play an important role in feeding behavior and energy homeostasis. More specifically, the effect of NmU has been mapped to particular hypothalamic nuclei. Thus, injection of NmU into either the PVN or arcuate nucleus immediately decreases food intake in rats (Wren et al., 2002). NmU-R2 mRNA is expressed predominantly in the central nervous system (see Section IV.C.) and indeed has been localized to the PVN and arcuate nucleus in rat and mouse (Howard et al., 2000; Graham et al., 2003). This distribution is consistent with direct effects of NmU via NmU-R2 in these regions. Administration of NmU intracerebroventricularly or into the PVN also increases gross-locomotor activity, body temperature, heat production, and oxygen consumption in rats (Hanada et al., 2001, 2003; Howard et al., 2000; Nakazato et al., 2000; Ivanov et al., 2002; Wren et al., 2002). However, NmU does not induce shivering, suggesting that the effect on body temperature is through chemical means. This generally involves sympathetic stimulation of brown adipose tissues and skeletal muscle. NmU can increase core body temperature in satiated rats in the absence of altered feeding behavior thereby indicating that the effects on feeding and energy expenditure are independent (Ivanov et al., 2002).
The precise way in which the central administration of NmU regulates feeding behavior and energy expenditure is unclear, although several mechanisms have been suggested. NmU induces c-Fos expression in a high proportion of oxytocin-immunoreactive neurons within the PVN and supraoptic nucleus (Niimi et al., 2001). Central infusion of oxytocin reduces food intake (Olson et al., 1991a,b) leading the authors to suggest that the activation of brain stem projecting oxytocin-containing neurons is responsible for the inhibitory effects of NmU on feeding. Interestingly, these authors also reported activation of arginine vasopressin-containing neurons in the PVN and supraoptic nucleus, leading to the suggestion that the previously reported inhibitory effects of intracerebroventricular NmU on water intake (Howard et al., 2000) are mediated by arginine vasopressin (Niimi et al., 2001). This is consistent with an ability of NmU to stimulate release of arginine vasopressin from hypothalamic explants (Wren et al., 2002) and increase plasma levels following central administration (Ozaki et al., 2002).
Intracerebroventricular administration of CRH also inhibits feeding in rats (Morley and Levine, 1982). The suppression of food intake, the increase in oxygen consumption, and the body temperature gain observed following centrally administered NmU are all absent in CRH knockout mice (Hananda et al., 2003). Furthermore, CRH containing neurons are found within the PVN, and NmU stimulates the release of CRH from hypothalamic explants (Wren et al., 2002) (see Section V.C.). Thus, similar to the effects of centrally administered NmU on stress responses (Section V.C.), these data suggest that the effects of NmU on feeding and energy balance are via CRH.
Leptin is a protein hormone with important roles in the regulation of feeding, body weight, and metabolism (Considine and Caro, 1997; Friedman and Halaas, 1998). It is secreted from adipocytes and acts in the hypothalamus to mediate its anorectic effects, and it may have significant interactions with NmU in the control of feeding behavior. This is suggested by the observation that the expression of NmU mRNA is reduced both within the hypothalamic SCN of the leptin-deficient ob/ob strain of mice and also in the ventromedial hypothalamus of fasted rats (Howard et al., 2000). Leptin also stimulates the release of NmU from hypothalamic explants (Wren et al., 2002). Furthermore, in obese Zucker rats that lack functional leptin receptors and are hyperphagic, NmU expression is reduced within the nucleus tractus solitarius (but not the inferior olive) of the brain stem. The nucleus tractus solitarius receives information relating to gastric loading and gut nutrients and relays this to the brain stem and forebrain, regions that regulate feeding (Rinaman et al., 1998). Taken together, these data suggest that NmU release is mediated by leptin and that the synthesis of NmU is restricted in the absence of leptin. In contrast to the decrease in NmU mRNA within the SCN of ob/ob mice (Howard et al., 2000), a recent study has reported an increase within the DMH of both ob/ob mice and in fasted normal mice (Graham et al., 2003). Whether this reflects true regional differences within the hypothalamus remains to be established.
Effects of leptin on hypothalamic neuropeptide signaling and the subsequent effects on feeding and body weight have been documented previously for several other neuropeptides (Inui, 1999). It is not uncommon, therefore, to associate leptin with neuropeptides that control feeding, and it is unlikely that the reduction in feeding and body weight by NmU occurs independently of leptin.
In contrast to the immediate effects of NmU on feeding when administered into the PVN or arcuate nucleus, it has a very delayed inhibitory effect following administration into the medial preoptic (MPO) area of the hypothalamus (Wren et al., 2002). Although the MPO is involved in reproductive function, c-Fos expression and neuropeptide Y levels increase after feeding. Furthermore, α-melanocyte-stimulating hormone reduces food intake when administered into the MPO (Kim et al., 2000), leading to the suggestion that NmU may interact with other regulators of feeding behavior to mediate its delayed effect (Wren et al., 2002).
Release of adrenal corticosteroids is enhanced by NmU (see Section V.C.), and this is associated with the promotion of prosurvival mechanisms while limiting other behaviors, such as feeding (Munck and Naray-Fejes-Toth, 1994; Sapolsky et al., 2000). It is therefore possible that some effects of NmU on feeding, particularly longer-term effects, are mediated by this loop.
E. Gastric Acid Secretion and Gastric Emptying
Many other hypothalamic peptides that influence feeding behavior also contribute to the central regulation of gastric acid secretion and gastric emptying (for review, see Tache et al., 1990; Tache and Yang, 1990). Although intraperitoneal injection of NmU has no effect on gastric acid secretion in conscious rats, intracerebroventricular administration reduces it (Mondal et al., 2003). This effect is independent of vagal innervation of the stomach. Furthermore, the effect is independent of prostaglandin E2, which is known to have both direct and indirect inhibitory effects via an action on parietal cells and the inhibition of gastrin secretion respectively. However, the influence of NmU on gastric acid secretion is blocked by intracerebroventricular administration of anti-CRH IgG and yohimbine, suggesting that NmU mediates its effects via CRH (see Section V.C.) and an increased sympathetic outflow. This effect is consistent with the hypothalamic expression of NmU and its receptors, particularly NmU-R2 (see Section IV.C.). Gastric emptying is also delayed by intracerebroventricular administration of NmU, and although CRH can reduce gastric emptying via the parasympathetic nervous system, this appears not to be the mechanism through which NmU acts (Mondal et al., 2003). A reduced gastric emptying is known to be associated with reduced food intake, and fast gastric emptying is associated with overeating (Duggan and Booth, 1980; Kelly, 1980). Thus, the effects of NmU on gastric acid secretion and gastric emptying are consistent with its effects on food intake and energy balance (see Section V.D.). NmU and its receptors are located within the gastrointestinal tract, including the stomach (see Sections III.B. and IV.C.), suggesting that NmU may regulate gastrointestinal function by local effects. Interestingly, compared with other secretagogues, NmU weakly stimulates histamine secretion from enterochromaffin-like cells in vitro (Lindström et al., 1997) and in vivo (Norlen et al., 2001). The histamine released by enterochromaffin-like cells floods adjacent parietal cells to stimulate acid production, and histamine release should therefore provide an indication of acid secretion.
F. Effects on Ion Transport
The contraluminal application of NmU-8 increases electrogenic ion transport in isolated porcine jejunal mucosa without contracting the longitudinal muscle overlying the mucosa (Brown and Quito, 1988). This action is dependent upon external chloride and is mediated by noncholinergic enteric neurons. However, the physiological and pathophysiological roles of NmU in the gastrointestinal tract remain to be fully defined. Expression of hNmU-R1 mRNA in the goblet cells of the ileum suggests that NmU also regulates secretory function and may therefore affect mucosal function (Howard et al., 2000).
G. Cancer
Laser capture microdissection followed by mRNA amplification and the use of high-density oligonucleotide probe arrays has been used for cell-specific profiling of gene expression in normal oral epithelial cells and those from oral tumors. Changes in the mRNA levels of over 600 genes were associated with oral cancer including that for NmU, which was down-regulated in each of the five cases studied (Alevizos et al., 2001). Furthermore, the promoter region of the NmU gene is hypermethylated in esophageal squamous cell carcinoma (ESCC), and this is accompanied by silencing at the mRNA level (Yamashita et al., 2002). Of approximately 120 genes commonly up-regulated in ESCC cell lines following the unmasking of epigenetic silencing, specific selection criteria identified three genes as worthy of further consideration of their tumor suppressing activity. One of these was NmU, and in colony focus assays using an ESCC cell line, the recombinant expression of NmU inhibited growth, thereby highlighting its potential role as a tumor suppressor gene (Yamashita et al., 2002). The gene for NmU is localized at 4q12 and this has been reported to show changes associated with cancer (Yamashita et al., 2002).
H. Pronociception
Both NmU (see Section III.A.) and receptors for NmU (see Section IV.C.) are expressed in the spinal cord. More specifically, NmU binding sites have been identified in lamina I and in the outer part of lamina II of rat spinal cord, and these are associated with the expression of mRNA for NmU-R2 (Yu et al., 2003) (see Section IV.C.). These areas are essential components in central nociceptive pathways, and recent studies indicate that NmU has pronociceptive effects (Cao et al., 2003; Yu et al., 2003). Thus, intrathecal administration of NmU to rats resulted in a reduced threshold for mechanical stimulation, thermal hyperalgesia, and behavioral responses associated with activation of nociceptive pathways including scratching, licking, and biting of the lower body (Yu et al., 2003). Furthermore, NmU administration enhanced flexor α-motorneuron activity in response to touch and noxious pinch. Similarly in mice, intrathecal injection of NmU produced thermal hyperalgesia and morphine-sensitive behavioral responses, whereas systemic administration in a perfused hindquarter preparation altered the electrophysiology of nociceptive dorsal horn neurons consistent with an increased basal activity and enhanced responsiveness to noxious mechanical stimulation (Cao et al., 2003). Thus, NmU may be involved in spinal nociceptive transmission and processing although the precise mechanisms through which it mediates these effects remain to be determined. It is also possible that some of the behavioral responses (see particularly Section V.C.) are due in part to these pronociceptive effects of NmU.
I. Immune Regulation
There are very specific patterns of expression of the mRNA for both NmU and NmU-R1 in immune cells in humans, strongly suggesting that NmU may play an immunomodulatory role. In particular, NmU is expressed in dendritic cells, monocytes, and B cells, whereas NmU-R1 is expressed in T cells and NK cells leading to the suggestion that there is an interaction between helper/antigen-presenting cells and effector populations (Hedrick et al., 2000). This has yet to be explored from a functional perspective.
VI. Summary
Since the discovery of NmU in the mid 1980s, progress toward elucidating the pathophysiological roles of this peptide has been comparatively slow. However, the conservation of structural elements of NmU across species and the widespread distribution of NmU and its receptors throughout the body point to a fundamental role in key physiological processes. Although clues have emerged from the patterns of distribution of NmU and its receptors, the complete spectrum of NmU-related activities remain to be defined. Current evidence suggests roles for NmU in pain, cancer, in the immune system, in the regulation of smooth-muscle contraction in the gastrointestinal and genitourinary tracts, in the control of blood flow and blood pressure, and in the regulation of feeding and energy homeostasis and stress responses. The recent cloning of two NmU receptors from several species should considerably increase the rate of progress in defining NmU function. In addition to allowing the determination of the cellular signaling pathways and cellular functions mediated by NmU, this may also allow the development of small molecule agonists and antagonists, hopefully with receptor subtype-selectivity, that could further help to describe the roles of NmU and the receptor subtypes through which different effects are mediated. Furthermore, the development of mice in which the genes for the receptors have been selectively deleted may greatly improve our understanding of the pathophysiological roles of NmU and may highlight the receptors as important therapeutic targets.
Acknowledgments
We gratefully acknowledge funding from the Biotechnology and Biological Sciences Research Council (BBSRC) and GlaxoSmithKline. We would like also to thank Dr. Alan Wise (GlaxoSmithKline, Stevenage, UK) for his support and contributions to the review.
Footnotes
-
↵1 Abbreviations: NmU, neuromedin U; VIP, vasoactive intestinal polypeptide; PP, pancreatic polypeptide; SCPB, small cardioactive peptide B; NT, neurotensin; pGlu, pyroglutamic acid; NmU-R1, neuromedin U receptor subtype 1; NmU-R2, neuromedin U receptor subtype 2; [Ca2+]i intracellular calcium concentrations; GPCR, G-protein coupled receptor; RIA, radioimmunoassay; ICC, immunocytochemistry; NmU-LIR, neuromedin U-like immunoreactivity; ACTH, adrenocorticotrophic hormone; CNS, central nervous system; DMH, dorsomedial hypothalamus; SCN, suprachiasmatic nucleus; GTPγS, guanosine 5′-[γ-thio]triphosphate; hFM-3, human FM-3; CHO, Chinese hamster ovary; hNmU-R1, human neuromedin U receptor subtype 1; hNmU-R2, human neuromedin U receptor subtype 2; SNP, single nucleotide polymorphism(s); NCBI, National Centre for Biotechnology Information; PVN, paraventricular nucleus; PLC, phospholipase C; cAMP, cyclic adenosine monophosphate; ERK, extracellular signal-regulated protein kinase; CRH, corticotrophic releasing hormone; MPO, medial preoptic; ESCC, esophageal squamous cell carcinoma.
-
DOI: 10.1124/pr.56.2.3.
- The American Society for Pharmacology and Experimental Therapeutics
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
- Article
- Abstract
- I. Discovery, Isolation, Structure, and Synthesis of Neuromedin U
- II. Structure-Activity Relationships of Neuromedin U
- III. Regional, Cellular, and Subcellular Distribution of Neuromedin U
- IV. Receptors for Neuromedin U
- V. Physiological Roles of Neuromedin U
- VI. Summary
- Acknowledgments
- Footnotes
- References
- Figures & Data
- Info & Metrics
- eLetters