


Abstract
Metformin, an oral antihyperglycemic, is increasingly being prescribed to pregnant women with gestational diabetes. Metformin is a hydrophilic cation and relies on organic cation transporters to move across cell membranes. We previously demonstrated that human and mouse placentas predominantly express organic cation transporter 3 (OCT3), but the impact of this transporter on maternal and fetal disposition of metformin is unknown. Using immunofluorescence colocalization studies in term human placenta, we showed that OCT3 is localized to the basal (fetal-facing) membrane of syncytiotrophoblast cells with no expression on the apical (maternal-facing) membrane. OCT3 positive staining was also observed in fetal capillaries. To determine the in vivo role of OCT3 in maternal and fetal disposition of metformin, we determined metformin maternal pharmacokinetics and overall fetal exposure in wild-type and Oct3-null pregnant mice. After oral dosing of [14C]metformin at gestational day 19, the systemic drug exposure (AUC0–∞) in maternal plasma was slightly reduced by ∼16% in the Oct3−/− pregnant mice. In contrast, overall fetal AUC0–∞ was reduced by 47% in the Oct3−/− pregnant mice. Consistent with our previous findings in nonpregnant mice, metformin tissue distribution was respectively reduced by 70% and 52% in the salivary glands and heart in Oct3−/− pregnant mice. Our in vivo data in mice clearly demonstrated a significant role of Oct3 in facilitating metformin fetal distribution and exposure during pregnancy. Modulation of placental OCT3 expression or activity by gestational age, genetic polymorphism, or pharmacological inhibitors may alter fetal exposure to metformin or other drugs transported by OCT3.
Introduction
Gestational diabetes mellitus (GDM) is a complication characterized by insulin resistance and glucose intolerance during pregnancy. GDM affects about 5%–13% of pregnancies in the United States; uncontrolled GDM can lead to serious adverse outcomes for the mother and fetus, such as difficult labor, hypertensive disorders, and preeclampsia, as well as neonatal hypoglycemia, respiratory distress syndrome, and macrosomia (Farrell, 2003; Simmons, 2011; Ryu et al., 2014). Historically, insulin has been the mainstay drug for the treatment of GDM. In recent years, oral hypoglycemic agents are increasingly being used in the management of GDM due to the ease of administration, lower cost, and better patient adherence than insulin (Ryu et al., 2014).
Metformin, a frontline drug for type 2 diabetes outside pregnancy, is increasingly being used in the treatment of GDM. Metformin acts as an insulin sensitizer and lowers blood glucose levels by suppressing hepatic gluconeogenesis, reducing intestinal glucose absorption, and stimulating glucose uptake in skeletal muscle and adipose tissues (Hundal and Inzucchi, 2003; Gong et al., 2012). Metformin is predominantly eliminated by the kidney with negligible hepatic elimination (Scheen, 1996; Kimura et al., 2005a, b; Tanihara et al., 2007). Membrane transporters play a critical role in metformin pharmacokinetics and pharmacodynamics because it is hydrophilic and positively charged at physiologic pH. Metformin is a substrate of multiple transporters, including the organic cation transporters 1–3 (OCT1–3), multidrug and toxin extrusion protein 1 and 2-K (MATE1, MATE2-K), and the plasma membrane monoamine transporter (Zhou et al., 2007; Gong et al., 2012). In humans, the liver-specific OCT1 (SLC22A1) mediates metformin uptake into the hepatocyte, a major site of metformin action (Hundal and Inzucchi, 2003; Gong et al., 2012; Wagner et al., 2016). The kidney-specific OCT2 (SLC22A2) works in concert with MATE1/2-K to mediate active secretion of metformin in renal tubular cells (Kimura et al., 2005; Masuda et al., 2006; Tanihara et al., 2007; Morrissey et al., 2013). The third member of the human OCT family, OCT3 (SLC22A3), is broadly distributed in many tissues, including the placenta, salivary glands, heart, brain, and intestine (Wu et al., 2000; Lee et al., 2014). We previously showed that OCT3/Oct3 is the primary organic cation transporter expressed in salivary gland epithelial cells and is responsible for metformin accumulation and secretion in salivary glands (Lee et al., 2014). Recently, we and others observed a reduced metformin oral bioavailability in nonpregnant Oct3−/− mice, suggesting that this transporter may also play a role in intestinal absorption of metformin (Chen et al., 2015; Shirasaka et al., 2016).
The use of metformin in pregnant women with GDM has brought attention to its passage across the placenta barrier (Charles et al., 2006; Wensel, 2009; Eyal et al., 2010). Despite very low passive membrane permeability, metformin readily crosses the placenta barrier (Vanky et al., 2005; Charles et al., 2006; Eyal et al., 2010). Clinical studies performed by us and others have reported that umbilical cord serum concentrations of metformin at the time of delivery were comparable to or even exceeded maternal concentrations, suggesting that metformin is actively transported from maternal circulation to fetal blood (Vanky et al., 2005; Charles et al., 2006; Eyal et al., 2010). Several in vitro and ex vivo studies also indicated the involvement of organic cation transporters in transplacental transport of metformin (Zwart et al., 2001; Kovo et al., 2008a,b). Among the known organic cation transporters, OCT3/Oct3 is the major isoform expressed in the placenta (Wu et al., 2000; Verhaagh et al., 2001; Zwart et al., 2001; Lee et al., 2013). Using uptake studies and Western blot analysis in membrane vesicles prepared from human term placenta, Sata et al. (2005) first demonstrated that OCT3 is expressed in basal but not microvillus membrane vesicles. Using quantitative PCR and targeted proteomics, we previously demonstrated that human and mouse placentas predominantly express OCT3/Oct3 mRNA and protein with minimal expression of other metformin transporters, including OCT1/2, MATE1/2 and plasma membrane monoamine transporter (Lee et al., 2013). Expression of OCT3/Oct3 in human and mouse placentas is influenced by gestational age with higher expression in late pregnancy (Lee et al., 2013). Based on these observations, we hypothesized that OCT3/Oct3 is involved in placental metformin transport and influences fetal exposure to metformin. The goal of this study is to determine the subcellular localization of OCT3 in human placenta by immunofluorescence colocalization and evaluate the in vivo significance of Oct3 in fetal metformin exposure using a pregnant mouse model with deletion of the Oct3 (Slc22a3) gene.
Materials and Methods
Animals.
The Oct3 (Slc22a3) null mice of the FVB inbred strain were originally developed by Dr. Denise Barlow (Zwart et al., 2001) and maintained by Dr. Alfred Schinkel (Netherlands Cancer Institute). After rederivation at Charles River Laboratories (Wilmington, MA) (Zhu et al., 2010), breeding pairs of the wild-type (Oct3+/+) and knockout (Oct3−/−) FVB mice were generously provided to us by Dr. John Markowitz at the University of Florida with approval from Dr. Schinkel. These mice were housed in the specific pathogen-free facility at the University of Washington. Mice were maintained on a 12/12-hour automatically timed light/dark cycle under a standard diet. The animal studies were approved by the Institutional Animal Care and Use Committee of the University of Washington.
Uptake Experiments.
Human OCT3 (hOCT3)-expressing HEK cells (Flp-In-293; Invitrogen, Waltham, MA) were previously generated (Duan and Wang, 2010). Cells stably transfected with pcDNA5/FRT vector were used as a control. Cells were maintained in Dulbecco’s modified Eagle’s medium containing 10% fetal bovine serum, 1% l-glutamine, 1% penicillin/streptomycin, and hygromycin B (150 μg/ml) at 37°C in 5% CO2 and 95% humidity. Metformin uptake and kinetic parameters were determined by radiotracer uptake assay as described previously (Lee et al., 2014; Wagner et al., 2017). Uptake experiments were performed in triplicate and repeated three times. Data are presented as mean ± S.D.
Localization of OCT3 in Human Placenta.
Human term placentas from healthy uncomplicated pregnancies were obtained at the Labor and Delivery Unit at the University of Washington (Lee et al., 2013). Upon collection, placentas were immediately snap-frozen and stored at −80°C until use. Frozen sections were prepared by Pathology Research Services at the University of Washington, and immunofluorescence staining was carried out using a procedure developed in our laboratory (Lee et al., 2014). Briefly, frozen human term placenta sections were fixed in ice-cold acetone for 5 minutes, immersed in PBS for 5 minutes, and then blocked in goat serum in PBS for 45 minutes at room temperature. The sections were then incubated overnight at 4°C with a polyclonal rabbit anti-OCT3 (1:125 dilution; Genway, San Diego, CA), colabeled with either anti-human CD31-PECAM1 (1:50 dilution; R&D Systems, Minneapolis, MN), anti-human laminin (1:1000 dilution; Sigma-Aldrich, St. Louis, MO), or anti-human CD71 (transferrin receptor) (1:200 dilution; Abbiotec, San Diego, CA). After primary antibody incubation, the sections were washed with PBS, and fluorescent tag-labeled secondary antibodies (Alexa 488 and Alexa 568, 1:500 dilution) were applied for 1 hour at room temperature. For nonspecific control, placenta tissue sections were incubated with blank serum, followed by secondary antibody incubation under the same conditions. After washing, ProLong Gold antifade medium with DAPI was mounted on the slides and placed under a coverslip. Fluorescent images were obtained with a Zeiss Axiovert 200 fluorescence microscope.
Transporter Expression Analysis in Oct3+/+ and Oct3−/− Placentas.
To determine if there were compensatory changes in the expression of other organic cation and related transporters in placentas of Oct3−/− mice, pregnant Oct3+/+ and Oct3−/− mice at gestational day (gd) 19 were euthanized using CO2. The placentas were immediately collected, flash-frozen in liquid N2, and stored at −80°C until use. Total RNA was extracted using Qiagen Mini RNeasy Kit (Valencia, CA) and reverse transcribed to first-strand cDNA using Invitrogen Superscript III reverse transcriptase according to the manufacturers’ instruction. Taqman real-time PCR assays were carried out according to the manufacturer’s protocols on an Applied Biosystems 7900HT fast real-time PCR system (Foster City, CA) as described previously (Duan and Wang, 2010; Lee et al., 2013).
In Vivo Pharmacokinetics Studies.
All in vivo studies were carried out in pregnant mice at gd 19 (term in mice is approximately 20 to 21 days), as we previously showed that the expression of Oct3 in the mouse placenta peaks around gd 19 (Lee et al., 2013). To obtain pregnant Oct3+/+ or Oct3−/− mice at gd 19, male and female mice (8–10 weeks old) of each genotype were mated. Gestational age was determined based on the estimated time of insemination, and the day that a vaginal plug was observed was assigned as gestational day 1. Progression of pregnancy was regularly monitored by visual inspection and by measuring the increase in body weight. At gd 19, mice were administered 15 mg/kg metformin containing 0.2 mCi/kg [14C]metformin (Moravek Biochemicals, Inc., Carlsbad, CA) by oral gavage. At various time points (15, 30, 45, 60, 120, 180, 240, and 480 minutes), mice (n = 3–5 mice at each time point) were euthanized by CO2 inhalation followed by cardiac puncture. Blood was collected using a heparin-coated syringe. Plasma was separated by centrifugation at 5000 g and stored at −20°C until analysis. Fetus and various maternal tissues were collected at each time point, snap-frozen in liquid N2, and stored at −80°C until use. Metformin concentrations in maternal plasma, tissue, and fetus homogenates were determined by scintillation counting using the method described by us previously (Lee et al., 2014). Metformin concentration was expressed as nanograms per gram for tissues and nanograms per milliliter for plasma.
Pharmacokinetic Data Analysis.
A population-based bootstrap method (Mager and Göller, 1998; Lee et al., 2014) was used to calculate the mean and confidence intervals of area under the concentration-time curves (AUCs) because metformin concentrations in plasma and tissue were sampled in different animals at each time point (one-point sampling). The following equations were used to obtain maternal plasma AUC, oral clearance (CL/F), terminal half-life (t1/2,β), and volume of distribution at terminal phase (Vβ)



 The terminal slope (β) was calculated by performing a linear regression of log concentrations at the last three to five time points that provided the best correlation (R2). The 95% confidence intervals for pharmacokinetic parameters were generated using a bootstrap method as described previously (Lee et al., 2014). Briefly, plasma or tissue concentrations were resampled with random replacement of individual animals using the R program to create pseudo concentration-time profiles. For each profile, the pharmacokinetic parameters were calculated using the equations defined above. The 95% confidence intervals for each parameter were calculated by taking the 2.5% and 97.5% quartiles. Peak plasma concentration (Cmax) was estimated by visual inspection of the plasma concentration-time curve averaged from all animals.
The terminal slope (β) was calculated by performing a linear regression of log concentrations at the last three to five time points that provided the best correlation (R2). The 95% confidence intervals for pharmacokinetic parameters were generated using a bootstrap method as described previously (Lee et al., 2014). Briefly, plasma or tissue concentrations were resampled with random replacement of individual animals using the R program to create pseudo concentration-time profiles. For each profile, the pharmacokinetic parameters were calculated using the equations defined above. The 95% confidence intervals for each parameter were calculated by taking the 2.5% and 97.5% quartiles. Peak plasma concentration (Cmax) was estimated by visual inspection of the plasma concentration-time curve averaged from all animals.
Statistical Analysis.
Data from in vitro studies were analyzed for statistical significance using unpaired Student’s t test. For pharmacokinetic data, 95% confidence intervals around the estimate for each pharmacokinetic parameter were determined using the nonparametric bootstrap method (Efron and Tibshirani, 1994). Two-sided P values were calculated using permutation tests as described previously (Westfall and Young, 1993; Lee et al., 2014). A P value < 0.05 was considered statistically significant.
Results
Transport of Metformin by hOCT3 Stably Expressed in HEK 293 Cells.
Using HEK293 cells stably expressing hOCT3, we determined the time- and concentration-dependent metformin uptake by hOCT3 (Supplemental Fig. 1). hOCT3-mediated metformin uptake was saturable, with a Km value of 1.16 ± 0.19 mM and a Vmax value of 3.12 ± 0.17 (nmol/min)/mg total protein. The Km value is similar to that previously obtained by Chen et al. (2010) for hOCT3 and is also comparable to the Km values reported for mouse Oct3 (Lee et al., 2014; Chen et al., 2015).
Expression and Membrane Localization of OCT3 in Human Placenta.
To determine the membrane localization and cell type-specific expression of OCT3 in human placenta, we performed immunofluorescence localization studies in healthy human term placentas. The specificity of the OCT3 polyclonal antibody was validated in our previous studies (Lee et al., 2014). As shown in Fig. 1, positive staining of OCT3 was observed in syncytiotrophoblasts as well as in fetal capillaries. Double immunostaining revealed significant costaining with laminin, a strong marker of the basement membrane, as well as CD31-PECAM1, a marker of the fetal vascular endothelium. In contrast, transferrin receptor, a marker of the microvillus (i.e., apical) membrane, showed no overlap with OCT3. No marked fluorescence was observed in slides treated with secondary antibodies (data not shown). These data demonstrated that OCT3 is localized to the basal (fetal-facing) membrane of the placental trophoblast cells and is also present in fetal endothelial cells.

Localization of OCT3 in human term placenta. Human term placenta sections were colabeled with antibodies for OCT3 (B, F, and J; green) and various membrane markers (C, CD31-PECAM1; G, laminin; K, the transferrin receptor; red). The nuclei were shown in blue (A, E, and I). Merged images are shown in (D, H, and L). Arrow indicates representative staining of fetal capillaries (D) and basal membrane of trophoblasts (H).
Metformin Pharmacokinetic Studies in Pregnant Oct3+/+ and Oct3−/− Mice.
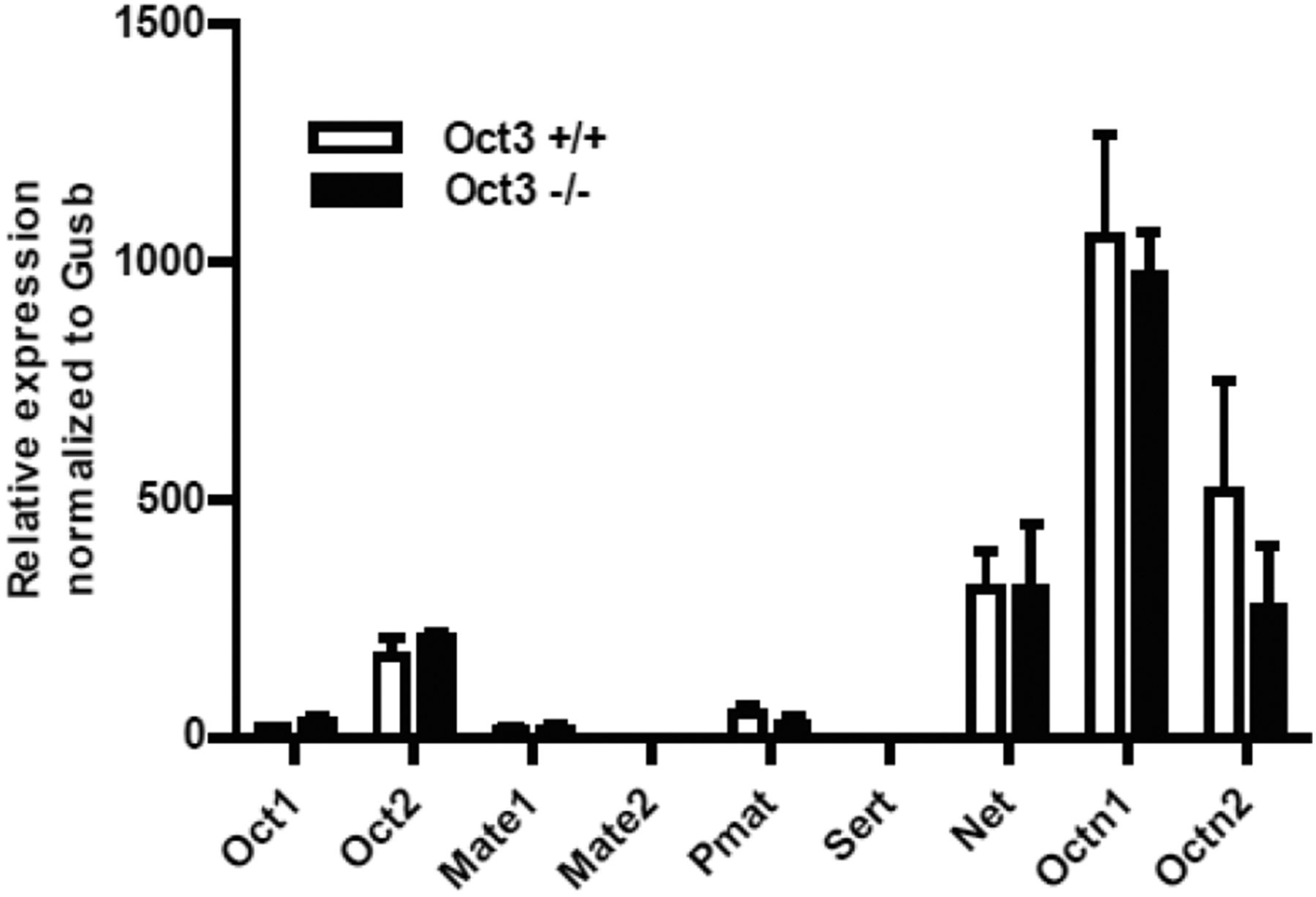
To determine the in vivo significance of OCT3 in maternal and fetal metformin disposition, we performed metformin pharmacokinetic studies in pregnant Oct3+/+ and Oct3−/− mice at gestational day 19. Prior to the pharmacokinetic studies, the mRNA expression of organic cation and related transporters was measured by quantitative RT-PCR in the placentas of Oct3+/+ and Oct3−/− mice. The data revealed no significant compensatory change in mRNA expression of these transporters (Fig. 2). At gd 19, Oct3+/+ or Oct3−/− pregnant mice were given by oral gavage a dose of 15 mg/kg metformin containing 0.2 mCi/kg [14C]metformin. Mice were euthanized at each time point (15–480 minutes), and metformin concentrations in maternal plasma, fetus, and tissue homogenates were measured. The maternal plasma concentration-time profiles and pharmacokinetic parameters in Oct3+/+ and Oct3−/− pregnant mice are shown in Fig. 3 and Table 1. After drug administration, peak metformin concentrations were achieved around 30 minutes, and the drug was mostly eliminated from the maternal circulation at 480 minutes in both Oct3+/+ and Oct3−/− pregnant mice (Fig. 3). Compared with the wild-type pregnant mice, Oct3−/− pregnant mice showed a narrower peak in the plasma concentration-time curve with a slightly lower (∼16%) AUC (Table 1). Other pharmacokinetic parameters were not statistically different between the Oct3+/+ and Oct3−/− pregnant mice (Table 1).

Expression of various organic cation transporters in the placentas of Oct3+/+ and Oct3−/− mice. Transcript of various organic cation and monoamine transporters in Oct3+/+ (n = 3) and Oct3−/− (n = 3) mouse placentas at gd 19 was determined by real-time PCR.

Maternal plasma metformin concentration-time profile in pregnant Oct3+/+ (▢) and Oct3−/− mice (■). Pregnant (gd 19) Oct3+/+ (▢) and Oct3−/− (■) mice were given a dose (15 mg/kg) containing 0.2 mCi/kg of [14C]metformin by oral gavage. At various time points (0–480 minutes) after drug administration, animals were sacrificed and blood, tissues, and fetuses were collected. Metformin concentrations in maternal plasma were measured by liquid scintillation counting. Data represent mean ± 1.96 S.E. (n = 3–5 mice/time point). Inset depicts the same data on a semilogarithmic scale.
Metformin pharmacokinetic parameters from pregnant Oct3+/+ and Oct3−/− mice
Data are presented as mean ± S.E. (Cmax) or with 95% confidence intervals in parentheses.
Impact of Oct3 Deletion on Fetal Disposition of Metformin.
At each time point of the pharmacokinetics study (Fig. 3), fetuses were collected and metformin concentrations in fetal tissue homogenate were determined (Fig. 4). After maternal drug administration, metformin fetal concentration increased progressively and peaked at ∼120 minutes in Oct3+/+ mice and ∼60 minutes in Oct3−/− mice (Fig. 4). Compared with fetuses in wild-type pregnant mice, a trend of lower fetal concentrations of metformin was observed in the Oct3−/− pregnant mice throughout the time course. At 120 and 180 minutes, fetal drug concentrations in the Oct3−/− pregnant mice were only ∼50% of those in the wild-type mice. The calculated fetal AUC0–480 minute and extrapolated AUC0–∞ in the Oct3−/− pregnant mice were reduced by 39% and 47%, respectively (Table 2). When the fetal-to-maternal plasma AUC ratio was calculated, the Oct3−/− pregnant mice showed approximately ∼30%–40% reduction (Table 2). These data clearly demonstrated that fetal metformin exposure is significantly reduced in the Oct3−/− pregnant mice.
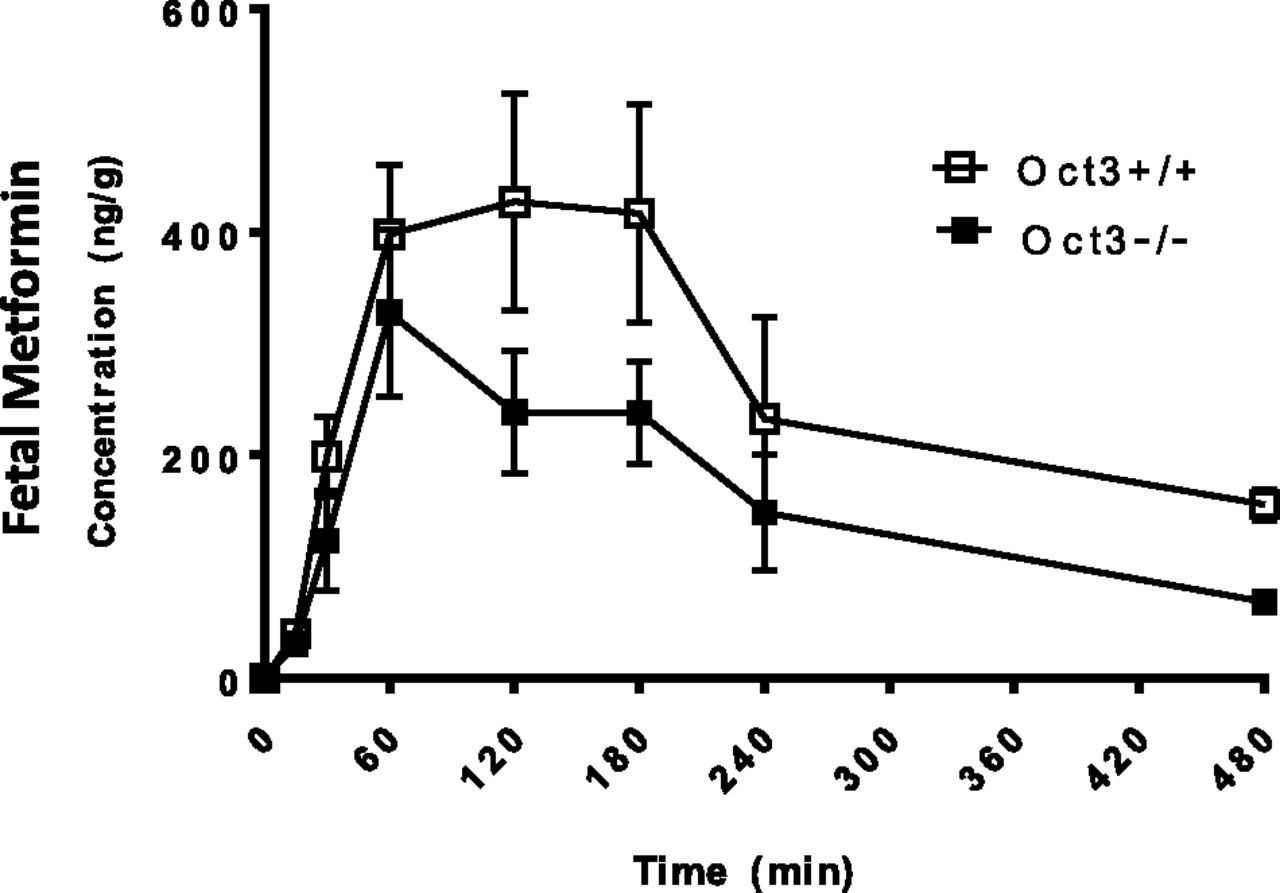
Fetal metformin concentration-time profile in Oct3+/+ and Oct3−/− mice following oral administration. Pregnant (gd 19) Oct3+/+ (▢) and Oct3−/− (■) mice were given a dose (15 mg/kg) containing 0.2 mCi/kg of [14C]metformin by oral gavage. At various time points (0–480 minutes) after drug administration, animals were euthanized, and blood, tissues, and fetuses were collected. Metformin concentrations in fetus homogenates were determined by liquid scintillation counting. Data represent mean ± 1.96 S.E. (n = 3–5 mice/time point).
Fetal metformin AUC and AUC ratios in Oct3+/+ and Oct3−/− pregnant mice
Data are presented as mean with 95% confidence intervals in parentheses.
Impact of Oct3 Deletion on Metformin Tissue Distribution in Pregnant Mice.
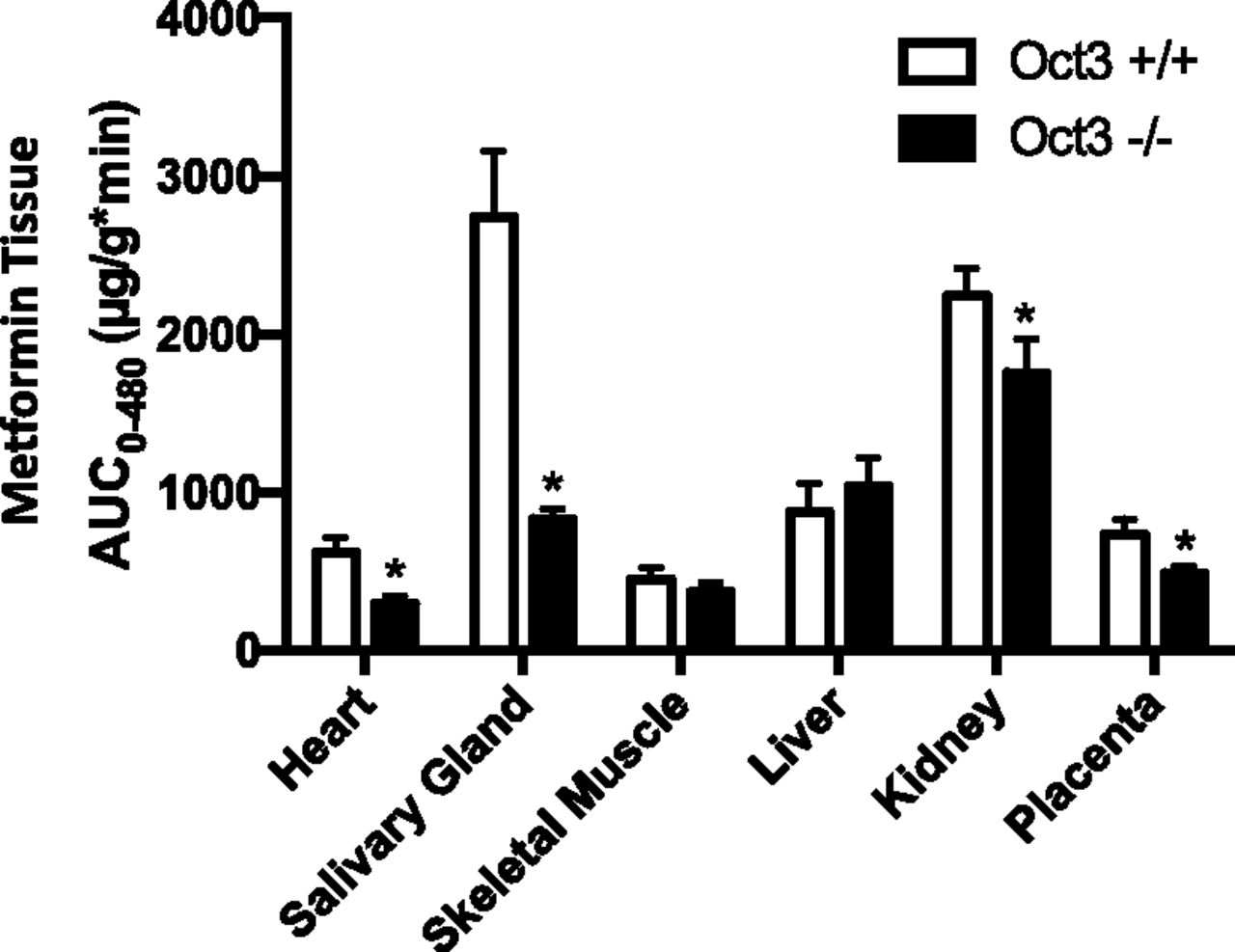
In nonpregnant male and female mice, we previously observed a substantial reduction in metformin tissue exposure in salivary glands and heart in the Oct3−/− mice, consistent with a high and predominant expression of Oct3 in these tissues in wild-type mice (Lee et al., 2014). When tissue metformin concentrations were measured in Oct3+/+ and Oct3−/− pregnant mice, the knockout mice showed a 70% and 52% reduction in metformin AUC0–480 minutes in salivary glands and heart, respectively (Fig. 5). A small reduction was also observed in the kidney and placenta, whereas no significant difference was observed in skeletal muscle and liver (Fig. 5).

Metformin AUC0-480 in various tissues in pregnant Oct3+/+ (▢) and Oct3−/− mice (■). Pregnant (gd 19) Oct3+/+ (▢) and Oct3−/− (■) mice were given a 15 mg/kg metformin dose containing 0.2 mCi/kg of [14C]metformin by oral gavage. At various time points (0–480 minutes) after drug administration, animals were euthanized, and blood, tissues, and fetuses were collected. Metformin concentrations in various maternal tissues were determined and AUC0–480 was calculated as described under Materials and Methods. Data represent mean ± 95% confidence interval. Two-sided P values were calculated using the permutation test as described under Materials and Methods (*P < 0.05).
Discussion
Metformin is increasingly being prescribed to pregnant women with GDM. Metformin readily crosses the placental barrier, but the underlying transport mechanism is unclear. In this study, we determined the subcellular localization of OCT3 in human placenta and investigated the in vivo role of Oct3 in fetal and maternal metformin disposition using a pregnant mouse model with Oct3 gene deletion. Our data demonstrated that OCT3 is localized to the basal membrane of the placental barrier and facilitates maternal-to-fetal transport of metformin. Our data revealed a significant impact of Oct3 on fetal and maternal metformin tissue distribution and pharmacokinetics.
The placenta is an important barrier tissue that mediates the exchange of nutrients, metabolites, and drugs between maternal and fetal circulation. This function is achieved by a multitude of transport proteins differentially expressed in the apical (maternal-facing) or basal (fetal-facing) membrane to mediate substrate transfer across the placenta (Ganapathy et al., 2000; Vähäkangas and Myllynen, 2009). Using functional studies and Western blot analysis in membrane vesicles prepared from human placenta, Sata et al. (2005) previously showed that OCT3 is expressed at basal membrane of human placenta trophoblasts. Using immunofluorescence colocalization studies, we confirmed that OCT3 is localized to the basal, but not apical, membrane (Fig. 1). Our study further revealed that OCT3 is also present in fetal endothelial cells (Fig. 1). While the physiologic function of OCT3 in the placenta is still unclear, the transporter is generally believed to serve as a clearing mechanism for bioactive amines (Ganapathy et al., 2000; Ganapathy and Prasad, 2005; Duan and Wang, 2010). In the fetus, circulating catecholamine levels are kept low for the maintenance of cardiovascular homeostasis (Bzoskie et al., 1997). The presence OCT3 in fetal capillaries may help to clear endogenous amines from the fetal circulation.
Because OCT3 is a bidirectional transporter driven by membrane potential, it can transport a substrate into or out of cells, depending on the electrochemical gradient of the substrate. The basal localization of OCT3 suggests that the transporter may couple with an active uptake transporter at the apical membrane to mediate maternal-to-fetal transport. This would facilitate drug transfer into the feus and increase fetal drug exposure. Alternatively, OCT3 may couple with an efflux transporter at the apical membrane to mediate fetal-to-maternal transport. This would promote drug clearance from the fetal compartment and decrease drug exposure in the fetus. To determine the actual role of Oct3 in placental transport of metformin in vivo, we performed pharmacokinetic studies in Oct3+/+ and Oct3−/− pregnant mice at gd 19. We chose to perform studies at gd 19 because we previously showed that OCT3/Oct3 expression in human and mouse placentas is gestational age dependent and reaches highest in late pregnancy (Lee et al., 2014). Metformin is highly water soluble and not bound to plasma proteins (Tucker et al., 1981; Graham et al., 2011). The clinical dosage of metformin can range from 500 to 3000 mg/day After a single oral dose, the peak plasma concentrations range from 1.0 to 1.6 mg/l after a 0.5-g dose, increasing to about 3 mg/l after a 1.5-g dose (Tucker et al., 1981; Graham et al., 2011). At the oral dose (15 mg/kg) used in our study, the observed Cmax in the maternal plasma in wild-type and knockout mice is 3.3 and 4.2 mg/l (Table 1), which is within the range of clinically observed metformin concentrations. The observed metformin Cmax in wild-type and knockout mice (26 and 32 µM, respectively) are far below its Km toward the OCT3/Oct3 transporter (Supplemental Fig. 1) (Chen et al., 2010, 2015; Lee et al., 2014), suggesting that our pharmacokinetic studies were conducted at plasma concentrations not saturating the Oct3 transporter.
Compared with wild-type pregnant mice, Oct3−/− pregnant mice showed a narrower peak in the maternal plasma concentration-time profile (Fig. 3), similar to that observed in nonpregnant Oct3−/− mice (Lee et al., 2014). This is likely due to a reduction in Oct3-mediated metformin distribution into peripheral tissues such as the salivary glands, heart, and the fetal compartment. The maternal plasma AUC of metformin in the Oct3−/− pregnant mice is only slightly reduced, which may be due to a small reduction in oral absorption as previously observed in nonpregnant Oct3−/− mice (Chen et al., 2015; Shirasaka et al., 2016). In contrast, fetal AUC was substantially reduced in the Oct3−/− pregnant mice (Fig. 4; Table 2). This reduction cannot be accounted for by the small reduction in the maternal AUC in knockout mice as there was still a significant reduction in fetal drug exposure after normalizing to maternal plasma AUC (Table 2). Thus, our in vivo data in mice clearly demonstrated a significant role of Oct3 in facilitating metformin fetal distribution and exposure during pregnancy.
Several groups have investigated the role of OCT3 in drug transport at the placenta barrier (Ganapathy et al., 2000; Zwart et al., 2001; Sata et al., 2005; Vähäkangas and Myllynen, 2009; Ahmadimoghaddam et al., 2012). In their original characterization of the Oct3 knockout mice, Zwart et al. (2001) showed that when MPP+, an experimental substrate of OCT3/Oct3, was administered into maternal circulation during pregnancy, MPP+ concentration measured at a single time point was significantly decreased in the embryos of the Oct3−/− genotype. This study suggested that Oct3 facilitates MPP+ transport from maternal blood into the fetus. However, based on Mate1 expression and ex vivo studies in a perfused rat placenta model, Ahmadimoghaddam et al. proposed a protective role of Oct3 at the placenta barrier. They suggested that analogous to renal organic cation secretion mediated by OCT2 and MATE1, Oct3 at the basal membrane transports metformin from fetal blood to the syncytiotrophoblast, which is then effluxed into the maternal blood by Mate1 at the apical membrane (Ahmadimoghaddam et al., 2012; Ahmadimoghaddam and Staud, 2013). Our data from comprehensive pharmacokinetic analyses in pregnant Oct3−/− mice are consistent with the initial finding by Zwart et al. and clearly support a role of Oct3 in facilitating maternal-to-fetal drug transport in vivo. Moreover, unlike the rat placenta, human, and mouse placentas minimally express MATE1/Mate1 (Otsuka et al., 2005; Lee et al., 2013), indicating that an OCT3/MATE1-mediated fetal-to-maternal elimination pathway may not exist in these species.
Our in vivo studies suggest that metformin in the maternal circulation is transported into the syncytiotrophoblast cells by an apical transporter and is then effluxed into the fetal side by basal Oct3. According to this model, there ought to be an apical transporter to initiate metformin uptake from the maternal blood. The molecular identity of this transporter is currently unclear. Previously, Ganapathy and Prasad had hypothesized that organic cations (e.g., MPP+) may be transported into placenta cells by the serotonin transporter and/or norepinephrine transporter, which are Na+- and Cl−-dependent secondary active transporters known to be expressed at the apical membrane of placenta (Ganapathy and Prasad, 2005). Metformin has been reported to be a transportable substrate of serotonin transporter (Han et al., 2015). In addition, the organic cation/carnitine transporter OCTN2 is expressed at the apical membrane and may mediate organic cation uptake into the syncytiotrophoblast cells (Ganapathy and Prasad, 2005; Grube et al., 2005). While more studies are necessary to validate the transporter(s) responsible for metformin uptake at the apical membrane, we propose a molecular model for organic cation transport at the placenta barrier based on our current study and available information in the literature (Fig. 6).

A proposed model for organic cation transport at the human placenta barrier. Organic cations, such as metformin, in the maternal circulation are first transported into the syncytiotrophoblast cells by NET, SERT, or OCTN2 at the apical membrane. Organic cations are then effluxed down their electrochemical gradient into the fetal side by OCT3 at the basal membrane. Larger and more lipophilic cations can be actively pumped back to the maternal circulation by P-gp and/or BCRP located at the apical membrane. BCRP, breast cancer resistance protein; NET, norepinephrine transporter; OCTN2, organic cation transporter novel type 2; P-gp, P-glycoprotein; SERT, serotonin transporter. This model is based on data in the present study and references (Vähäkangas and Myllynen, 2009; Ganapathy et al., 2000).
In conclusion, our study demonstrated that OCT3 is localized to the basal membrane of human placenta trophoblast cells and fetal exposure to metformin is significantly reduced in the Oct3−/− pregnant mice. Our data support a role of Oct3 in facilitating maternal-to-fetal drug transport at the placenta barrier, leading to enhanced fetal exposure to metformin. To our knowledge, this is the first complete pharmacokinetics study that clearly established Oct3 as an important determinant of fetal exposure to metformin in vivo. It is reasonable to speculate a similar role of OCT3 on fetal exposure to metformin during human pregnancy because human and mouse placentas share a similar expression pattern of the OCT/Oct transporters (Lee et al., 2014). If so, modulation of OCT3 activity by gestational age, genetic polymorphism, or pharmacological inhibitors may have clinical consequence on fetal exposure to metformin or other drugs transported by OCT3.
Acknowledgments
We are thankful to Drs. John Markowitz and Haojie Zhu (University of Florida) for providing breeding pairs of Oct3+/+ and Oct3−/− mice for our study.
Authorship Contributions
Participated in research design: Lee, Hebert, Wang.
Conducted experiments: Lee.
Performed data analysis: Lee, Wagner, Liang, Rice, Wang.
Contributed new reagents or analytical tools: Easterling, Liang, Rice.
Wrote or contributed to writing of the manuscript: Lee, Hebert, Wagner, Wang.
Footnotes
- Received March 12, 2018.
- Accepted June 29, 2018.
This work was supported by National Institutes of Health [Grants U10HD047892, R01GM066233, T32GM007750].
↵
 This article has supplemental material available at molpharm.aspetjournals.org.
This article has supplemental material available at molpharm.aspetjournals.org.

Abbreviations
- AUC
- area under the concentration-time curve
- gd
- gestational day
- GDM
- gestational diabetes mellitus
- HEK
- human embryonic kidney
- MATE
- multidrug and toxin extrusion protein
- MPP+
- 1-methyl-4-phenylpyridinium
- OCT
- organic cation transporter
- PBS
- phosphate-buffered saline
- PMAT
- plasma membrane monoamine transporter
- Copyright © 2018 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}