


Abstract
The class Frizzled (FZD) or class F of G protein–coupled receptors consists of 10 FZD paralogues and Smoothened (SMO). FZDs coordinate wingless/Int-1 signaling and SMO mediates Hedgehog signaling. Class F receptor signaling is intrinsically important for embryonic development and its dysregulation leads to diseases, including diverse forms of tumors. With regard to the importance of class F signaling in human disease, these receptors provide an attractive target for therapeutics, exemplified by the use of SMO antagonists for the treatment of basal cell carcinoma. Here, we review recent structural insights in combination with a more detailed functional understanding of class F receptor activation, G protein coupling, conformation-based functional selectivity, and mechanistic details of activating cancer mutations, which will lay the basis for further development of class F–targeting small molecules for human therapy.
SIGNIFICANCE STATEMENT Stimulated by recent insights into the activation mechanisms of class F receptors from structural and functional analysis of Frizzled and Smoothened, we aim to summarize what we know about the molecular details of ligand binding, agonist-driven conformational changes, and class F receptor activation. A better understanding of receptor activation mechanisms will allow us to engage in structure- and mechanism-driven drug discovery with the potential to develop more isoform-selective and potentially pathway-selective drugs for human therapy.
Introduction
Wingless/Int-1 (WNT) and Hedgehog (HH) signaling is essential for proper coordination of embryonal development by regulating cellular fate, orientation, positioning, polarity, proliferation, and differentiation. In addition to their crucial involvement early in life, these pathways maintain importance in the adult in a plethora of processes such as in the maintenance of functional stem cell niches to govern tissue renewal, for example, in epithelial tissues, the hematopoietic system, bone, the nervous system, or liver (Ingham and McMahon, 2001; Chien et al., 2009; Wend et al., 2010; Holland et al., 2013; Ingham, 2018; Steinhart and Angers, 2018). While tightly regulated, WNT signaling and HH signaling are both required during embryogenesis and in the adult, and deregulation of the pathways manifests in devastating diseases including developmental disorders, bone and cardiovascular diseases, neurologic disorders, fibrosis, and various forms of cancers (Polakis, 2000; Gould and Manji, 2002; Nusse, 2005; Sen, 2005; De Ferrari and Moon, 2006; Chien et al., 2009; Hoeppner et al., 2009; Freese et al., 2010; Königshoff and Eickelberg, 2010; Wend et al., 2010; Clevers and Nusse, 2012; Holland et al., 2013; Foulquier et al., 2018). Thus, targeting these pathways brings along a large—yet mostly untapped—therapeutic potential, which needs to be balanced with an obvious risk for unwanted side effects.
Class F Receptors and Their Ligands
While WNT effects are mediated by 10 mammalian paralogues of Frizzleds (FZDs) 1–10, HH effects are indirectly mediated by Smoothened (SMO), all of which comprise the class F of G protein–coupled receptors (GPCRs) (Foord et al., 2005; Schulte, 2010). WNT lipoglycoproteins are ligands of several WNT receptors, of which the seven transmembrane (TM)-spanning FZDs are seen as the main component acting in concert with a series of coreceptors such as low-density lipoprotein receptor-related protein 5/6 (LRP5/6), receptor tyrosine kinase-like orphan receptor, receptor tyrosine kinase, reversion-inducing cysteine-rich protein with kazal motifs, and others (Vanhollebeke et al., 2015; Cho et al., 2017; Driehuis and Clevers, 2017; Vallon et al., 2018). In the case of HH/SMO signaling, the cholesterol transporter Patched binds the HH protein and alters cholesterol levels in the inner leaflet of the cell membrane, and cholesterol/sterol interaction with SMO are thought to regulate SMO activity (Qi et al., 2018; Kong et al., 2019; Kowatsch et al., 2019).
A Short Overview of Molecular Mechanisms of WNT and HH Signaling
The complexity of WNT and HH signaling was recently reviewed elsewhere. Here, we will only present basic concepts of signaling diversity, referring the reader to excellent recent reviews in the field (Nusse and Clevers, 2017; Wu et al., 2017; Grainger and Willert, 2018; Steinhart and Angers, 2018; Kong et al., 2019).
WNT signaling can be divided into dishevelled (DVL)-dependent signaling, including both the β-catenin-dependent WNT/β-catenin pathway as well as the planar cell polarity (PCP) pathway, and DVL-independent signaling. Even though the underlying mechanisms of pathway selection are not fully understood, it is established that the WNT coreceptors LRP5/6 are essential to drive WNT signals—orchestrated by FZDs—toward the transcriptional regulator β-catenin (MacDonald et al., 2007; Nusse and Clevers, 2017). It should be mentioned that FZDs play an important role in pathway selectivity since some FZDs, such as FZD3 and FZD6, most likely do not mediate WNT/β-catenin signaling (Golan et al., 2004; Wang et al., 2016; Corda and Sala, 2017). Even in the presence of LRP5/6, the WNT/PCP pathway does not involve β-catenin but also involves the phosphoprotein DVL in addition to a multitude of other so-called PCP-core proteins (Humphries and Mlodzik, 2018). While WNT/β-catenin signaling relays transcriptional and proliferative input, PCP signaling orchestrates cytoskeletal changes translating into cellular asymmetry, migration, tissue elongation, polarity, and tissue structuring information most obvious in two-dimensional, planar epithelial tissues (Semenov et al., 2007; Humphries and Mlodzik, 2018). The pathways that we define here as DVL-independent signaling branches are mediated by heterotrimeric G proteins not excluding pathways that could be independent of both DVL and G proteins. The subdivision of WNT signaling into DVL-dependent and -independent pathways emerges as mechanistically likely, as we will elaborate later on in this review when discussing conformational and functional selection of signaling pathways by different FZD conformations. Albeit the literature contains contradictory data (Liu et al., 2001; Kilander et al., 2014b; Koval et al., 2016), the concept emerges that simultaneous interaction of DVL and heterotrimeric G proteins with a single FZD is mutually exclusive (Kilander et al., 2014b; Gammons et al., 2016; Hot et al., 2017; Strakova et al., 2017; Schulte and Wright, 2018; Bowin et al., 2019; Wright et al., 2019). The network of heterotrimeric G protein signaling downstream from FZDs is not completely mapped, although substantial progress has been made in recent years to define the FZD-G protein coupling selectivity, mechanisms, and the physiologic relevance of FZD-G protein signaling (Katanaev et al., 2005; Katanaev and Buestorf, 2009; Kilander et al., 2011, 2014a,b; Koval and Katanaev, 2011, 2018; von Maltzahn et al., 2011; Halleskog et al., 2012; Halleskog and Schulte, 2013; Dijksterhuis et al., 2014; Lüchtenborg et al., 2014; Park et al., 2015; Arthofer et al., 2016; Ramírez et al., 2016; Weivoda et al., 2016; Hot et al., 2017; Petersen et al., 2017; Strakova et al., 2017; Wright et al., 2018, 2019).
The release of constitutive inhibition of SMO upon HH-binding to Patched results in the translocation of active SMO to the cilia and activation of an oncogene glioma-associated homolog (GLI)–dependent transcriptional program (Wu et al., 2017; Kong et al., 2019). This pathway was—similar to the WNT/β-catenin pathway—described as being independent of heterotrimeric G proteins. Similar to the discussion of whether FZDs are GPCRs, the question of whether SMO is a GPCR was a matter of intense debate (Ayers and Thérond, 2010; Arensdorf et al., 2016). Interestingly, the overall picture between WNT and HH signaling diverges here substantially since there is now overwhelming evidence that HH/GLI signaling involves direct coupling of SMO to heterotrimeric G proteins (Riobo et al., 2006; Riobo and Manning, 2007; Shen et al., 2013; Manning et al., 2015; Guo et al., 2018; Qi et al., 2019), whereas WNT/β-catenin signaling most likely does not (Bowin et al., 2019). Furthermore, classic GPCR-regulating entities, such as GPCR kinases and β-arrestins were identified as intrinsic elements of SMO signaling (Chen et al., 2004b; Kovacs et al., 2008).
The identification of the plant alkaloid cyclopamine as a SMO antagonist laid the foundation for a rather rich pharmacological toolbox for SMO including a wide range of small molecule ligands with varying efficacies, with important clinical applications for the treatment of basal cell carcinoma and acute myeloid leukemia (Chen, 2016; Hoy, 2019). In contrast, hardly any small molecules that target FZDs have been identified and thoroughly validated regarding their mode of action (Generoso et al., 2015; Zhang et al., 2017a; Riccio et al., 2018) and information from the recent crystal structure of a ligand-free FZD4 suggests that targeting FZDs with small molecules could be challenging (Yang et al., 2018).
Frizzleds: Ligand-Receptor Interaction
WNT proteins (19 members of the family in humans) interact with the cysteine-rich domain (CRD) of FZDs, but not SMO, involving their lipid modification and a common fold including an index finger and a thumb (Janda et al., 2012; Willert and Nusse, 2012; Hirai et al., 2019). In the published crystal structures, the lipidated thumb domain of WNT proteins binds a lipophilic groove on the CRD and an elongated index finger recognizes the CRD close to its C terminus (Janda et al., 2012; Hirai et al., 2019). This interaction, which shows affinities in the lower nanomolar range explains—at least in part—WNT-FZD selectivity (Dijksterhuis et al., 2015). However, how WNT-CRD interaction physically translates into structural rearrangements in the seven TM cores of FZDs to establish and stabilize an active receptor conformation remains a mystery. Coreceptors of different kinds are implicated to mediate WNT-FZD and pathway selectivity (Hendrickx and Leyns, 2008; Dijksterhuis et al., 2015; Wang et al., 2016; Eubelen et al., 2018). In addition to WNTs, Norrin presents a soluble FZD4-selective ligand, which also interacts with the FZD-CRD forming a complex with LRP5/6 to feed into the WNT/β-catenin pathway (Xu et al., 2004; Ke et al., 2013; Chang et al., 2015).
All WNT proteins, except for Drosophila melanogaster WNTD are lipidated at a conserved serine residue rendering them highly lipophilic (Willert et al., 2003; Willert and Nusse, 2012). Substantial evidence exists that the lipidation of WNTs is essential for their agonist activity; furthermore, the crystal structures of WNTs and FZD-CRDs suggest that the lipid moiety interacts with a conserved lipophilic groove on the CRD. This concept poses an interesting thermodynamic resolvation conundrum (Willert et al., 2003; Schulte et al., 2005; Janda et al., 2012; Willert and Nusse, 2012; Deshpande et al., 2019). WNT transport from the trans-Golgi network depends on the seven TM-spanning protein Wntless (or Evenness Interrupted) (Bänziger et al., 2006; Bartscherer et al., 2006) and it remains thus far unclear how mature WNT is either embedded with the lipid moiety in the plasma membrane or packaged into lipoprotein particles, carrier proteins, or exosomes (Ching and Nusse, 2006; Herr and Basler, 2012). Irrespective of the packaging state of the WNT protein, the lipid moiety is shielded from the aqueous surrounding and lipid exposure to water is energetically unfavorable. Purified WNTs are solubilized with detergents such as 3-[(3-cholamidopropyl)dimethylammonio]-1-propanesulfonic acid and stabilized by bovine serum albumin, indicating that the acyl group is bound to a carrier protein (Willert, 2008; Mihara et al., 2016). The thermodynamic problem arises when the shielded fatty acid group needs to leave the lipophilic surrounding to be transported through an aqueous surrounding in order to engage with the lipophilic groove on the CRD of the receptor. This process is intrinsically unfavorable, and would therefore require energy, catalysis, or as yet unidentified mechanisms to be accomplished. Thus, it remains obscure how cells handle the lipid modification of WNTs during release from the cell, extracellular transport, and recognition by the receptors through CRD binding in agreement with the published crystal structures (Janda et al., 2012; Hirai et al., 2019). This aspect becomes particularly interesting in the context of the ability of FZDs to mediate CRD-independent WNT effects and the ability of nonlipidated WNTs to induce signaling (Chen et al., 2004a; Speer et al., 2019). In conclusion, CRD interactions with WNTs are essential, providing affinity and proximity; however, other mapping additional interactions with the receptor could be important to fully understand the mode of action of WNTs as FZD agonists.
SMO: Ligand-Receptor Interaction—a Mystery
While WNTs directly interact with FZDs to induce WNT signaling, HH proteins (desert HH, Indian HH, and sonic HH) interact with a 12 TM-spanning protein called Patched (Alcedo et al., 1996; Stone et al., 1996). Patched acts as a cholesterol transporter and negative regulator of SMO activity by constitutively reducing cholesterol levels in the inner leaflet of the membrane (Zhang et al., 2018; Kong et al., 2019; Kowatsch et al., 2019). There is recent evidence that cholesterol and oxysterols could act as primary endogenous ligands at SMO through interaction with the SMO-CRD or the TM-spanning core (Nachtergaele et al., 2012; Huang et al., 2016, 2018; Luchetti et al., 2016; Raleigh et al., 2018; Deshpande et al., 2019; Hedger et al., 2019). However, similar to the lack of knowledge about how lipidated WNTs reach the CRD of FZDs, it remains unclear how cholesterol reaches the extracellular CRD binding pocket on SMO in living cells. In this regard, the role of a postulated intramolecular tunnel in SMO for cholesterol trafficking has neither been functionally nor experimentally confirmed thus far (Huang et al., 2018; Deshpande et al., 2019; Hedger et al., 2019; Qi et al., 2019).
Similarly to the WNT/β-catenin pathway, it remains a matter of debate under which circumstances the SMO/GLI pathway is mediated through heterotrimeric G proteins. The most recent data strongly support the concept that SMO acts as a Gi/o and G12/13-coupling GPCR feeding into the SMO/GLI pathway (Riobo et al., 2006; Ogden et al., 2008; Shen et al., 2013; Manning et al., 2015; Qi et al., 2019; Wright et al., 2019). However, the role of G12/13 proteins downstream from SMO remains less well defined (Guo et al., 2018; Wright et al., 2019).
Signal Initiation: Different Concepts
Frizzleds.
Currently, two mechanisms emerge that explain WNT/FZD-dependent signal initiation. In the WNT/β-catenin pathway, WNT binding to the CRD of FZD results in signalosome formation (Gammons et al., 2016; Bilic et al., 2017; DeBruine et al., 2017a). The WNT/FZD complex appears to serve as a platform—in cooperation with the coreceptors LRP5/6—for the orchestration of dynamic DVL recruitment, which results in the inhibition of the β-catenin destruction complex and transcriptional regulation through T-cell/lymphoid enhancer-binding transcription factors (Nusse and Clevers, 2017; Gammons and Bienz, 2018; Steinhart and Angers, 2018). Interestingly, in this model it remains completely unclear what protein dynamic changes are evoked by ligand binding in FZDs to promote and regulate productive FZD-DVL interaction and signaling. While some data point to a constitutive, ligand-independent interaction between FZD and DVL, particularly in overexpressing cell systems (Valnohova et al., 2018; Wright et al., 2019), other studies argue for a WNT-induced dissociation of a preformed FZD-DVL complex (Gammons et al., 2016) or enhanced recruitment of DVL to FZD in response to agonist (the FZD4-selective Norrin) (Bang et al., 2018). These discrepancies clearly underline that the dynamics in the FZD-DVL interaction are not sufficiently understood. The modus of interaction between FZD and DVL proteins is complex and the functional role of FZD-DVL to mediate signaling is rather unclear and is best described as a scaffolding function orchestrating interaction of signaling components, such as axin, casein kinase 1, β-arrestin, adapter protein 2, and others (Bryja et al., 2007; Schwarz-Romond et al., 2007; Yu et al., 2007; Strakova et al., 2018). Initially, a highly conserved, unconventional, internal PDZ ligand domain in the C terminus of FZDs, a KTxxxW sequence, was identified to mediate FZD-DVL contact to the PDZ domain of DVL (Umbhauer et al., 2000). Furthermore, other regions on the cytoplasmic parts of FZDs were pointed out to be important for a FZD-DVL interface. The flanking regions of intracellular loop (IL) 3 of FZDs provided a discontinuous docking site for DVL (Tauriello et al., 2012; Wright et al., 2018). Importantly, the DEP domain of DVL appears to have a key role for FZD interaction as well as for membrane recruitment involving electrostatic interaction between positive charges on the DEP domain and the negative head groups of the phospholipids (Simons et al., 2009; Gammons et al., 2016). Additional regions in FZDs, such as IL1, IL2, and C terminal residues distal of the KTxxxW sequence on the helix 8 of the receptor were identified by mutagenesis to be involved in FZD-DVL interaction (Pau et al., 2015; Bertalovitz et al., 2016; Strakova et al., 2017). However, it remains unclear if IL1 or IL2 are, in fact, direct interaction sites or if mutation of those residues affect the overall conformation of the receptor reducing its ability to recruit DVL. Particularly in the case of the conserved FZD4 Y2502.39 (superscript numbers refer to Ballesteros-Weinstein nomenclature for GPCRs) (Ballesteros and Weinstein, 1995) in the C terminal end of IL1, we could show that an overall structural change of mutagenesis rather than direct interaction of the amino acid with DVL was the reason for reduced FZD-mediated DVL recruitment (Strakova et al., 2017).
In addition to the formation of a heterogenous signaling platform consisting of FZDs, LRP5/6, additional coreceptors, and DVL, recent reports point at important dynamic changes in FZD-FZD interactions (DeBruine et al., 2017b). It has been known for some time that FZDs can form dimers or oligomers, even though their function and relevance for signal initiation were more unclear (Kaykas et al., 2004). The FZD4-selective agonist Norrin acts as a homodimeric peptide and is capable of binding two CRDs of FZD4 (Ke et al., 2013). Furthermore, it was shown that unsaturated fatty acids can dimerize CRDs of FZDs, implicating that the lipid modification of WNTs could bridge two CRDs resulting in ligand-induced receptor dimerization or oligomerization (DeBruine et al., 2017a; Nile et al., 2017). In contrast to the model that ligand binding brings receptors together, we reported that FZD6 dimers dissociate upon WNT-5A stimulation to promote signaling toward extracellular signal–regulated kinases 1/2 (Petersen et al., 2017). Thus, the dimeric form of FZD6 constitutes the ligand-interpreting receptor species, whereas the monomeric form presents the active species initiating signaling. Reassociation of FZD6 dimers in the range of minutes after WNT addition correlated with signal termination, further supporting the concept of dynamic dimerization. The argument that a monomeric GPCR presents the minimal signaling unit is established for many receptors that show ligand-dependent G protein coupling when solubilized in detergent micelles at monodispersion or embedded in high-density lipoprotein particles or nanodiscs (Whorton et al., 2007; Liang et al., 2017; Zhang et al., 2017c; Draper-Joyce et al., 2018; García-Nafría et al., 2018a,b; Krishna Kumar et al., 2019). Furthermore, inactive GPCRs frequently crystallize as dimers (see also SMO) (Wang et al., 2013), whereas not a single active G protein-bound GPCR structure was reported as a dimer.
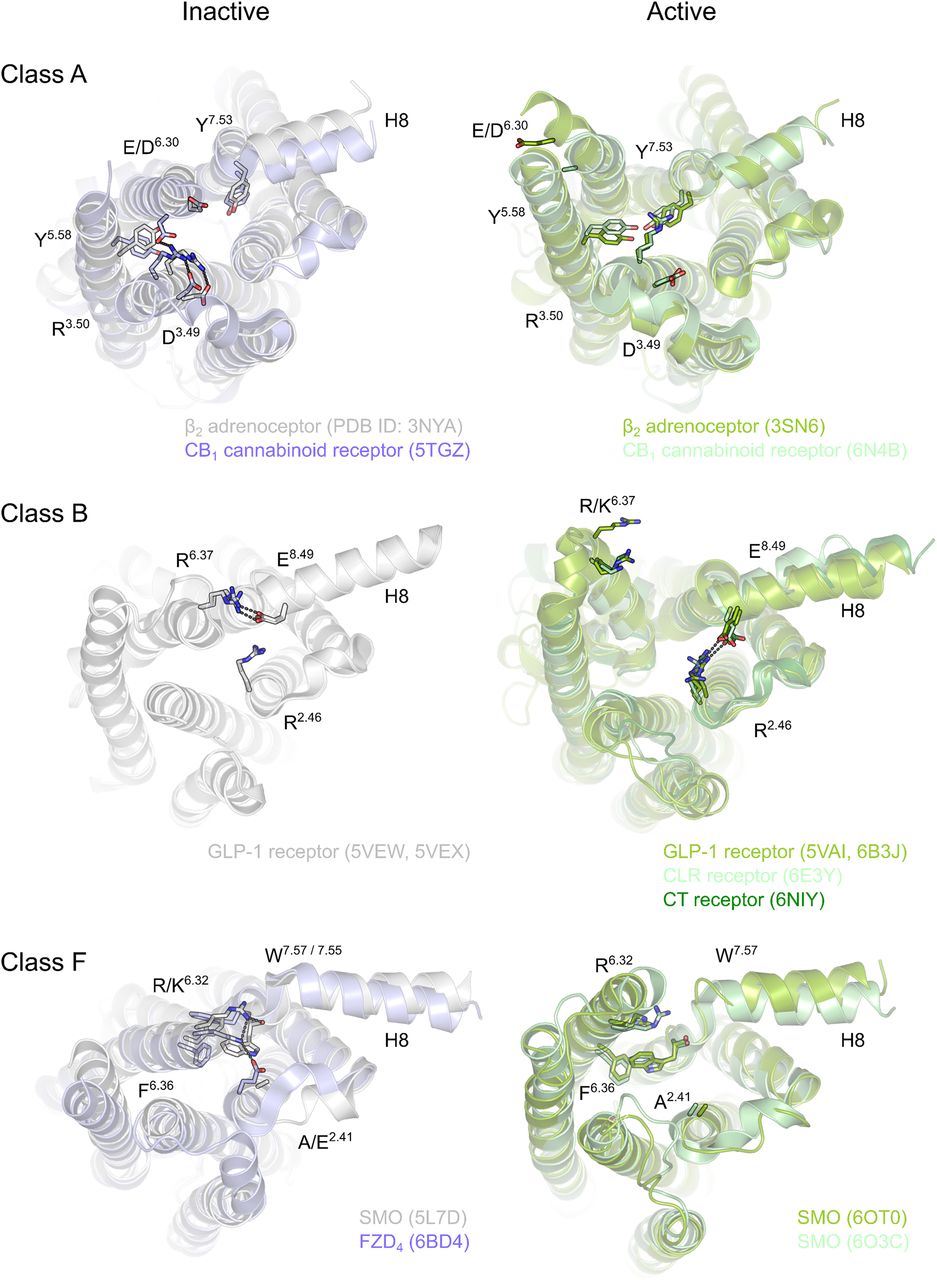
The idea of dimer dissociation upon receptor activation also merges well with the second concept of class F receptor activation, which is reminiscent of what is known for the activation of class A and B GPCRs: the agonist, the receptor, and the heterotrimeric G proteins act in a ternary complex to induce GDP release and subsequent GTP binding of the Gα subunit as the primary output of the ligand-receptor interaction (De Lean et al., 1980; Schulte and Wright, 2018). Decades of structural analysis have identified a common theme in GPCR activation generally involving a swing out of helix 6 of the receptor firmly accommodating the G protein in the active state of the receptor (Fig. 1) (Gether and Kobilka, 1998; Latorraca et al., 2017; Weis and Kobilka, 2018). FZD Förster resonance energy transfer probes designed to monitor agonist-induced dynamics, especially in TM6 of the receptor relative to the C terminus, support the idea that agonist stimulation of FZDs induces an active state similar to those observed in class A and B receptors (Schulte and Wright, 2018; Wright et al., 2018). In fact, the recently identified molecular switch between the lower ends of TM6 and TM7, which is highly conserved among class F receptors, is very similar to a polar network essential for class B receptor activation and G protein coupling (Liang et al., 2018b). Its functional relevance is further underlined by the oncogenic potential of mutations in the residues participating in the molecular switch, such as the cancer-associated SMOM2 mutant in W7.55 (Wright et al., 2019).

Comparison of the intracellular rearrangements between the active and inactive states of class A, B, and F GPCRs. In class A, interactions between TM3 and TM6 stabilize the inactive receptor conformations, whereas in classes B and F similar interactions are observed between TM6 and TM7/8 and to a lesser extent in TM2. In each of these receptor classes, outward movement of TM6, manifesting the receptor activation, breaks the observed interaction pattern. Currently, active structures of class C GPCRs remain unsolved, and thus class C is excluded from the comparison. Receptors are shown as a range of gray and green cartoons (inactive and active structures, respectively) and the key residues as corresponding sticks. Black dashes indicate possible hydrogen bonds. View is from the intracellular side of the receptors. PyMOL (The PyMOL Molecular Graphics System, version 2.0; Schrödinger, LLC) was used for visualization.
Smoothened.
Similar to FZDs, different conceptual perceptions of receptor activation mechanisms have emerged. On one hand, SMO translocation to the cilium has served as a measure of receptor and pathway activation (Milenkovic et al., 2009). While this physical relocation of SMO coincides with pathway activation, it could be misleading regarding the biophysical and pharmacological understanding of receptor activation manifesting as a conformational change in the receptor molecule. As an example, the plant alkaloid cyclopamine, which binds SMO without (antagonist) or with negative (inverse agonist) efficacy, affects cilial translocation of SMO, without promoting conformational changes resulting in receptor activation (Incardona et al., 1998; Chen et al., 2002a; Wang et al., 2009; Weierstall et al., 2014; Schulte and Kozielewicz, 2019; Wright et al., 2019). The availability of diverse SMO ligands ranging from the natural compound cyclopamine, smoothened agonist [(SAG) series of compounds], smoothened antagonists (series of compounds), and diverse sterols have been thus far helpful to define small molecule–driven SMO activation and signal initiation (Chen et al., 2002b; Frank-Kamenetsky et al., 2002; Chen, 2016; Kozielewicz et al., 2019).
SMO in Ternary Complex with Heterotrimeric Gi or Nanobody NbSmo8
Over the years, detailed structural and mechanistic insight into the activation of class F receptors was lacking, and the concept of class F receptors coupling to heterotrimeric G proteins was—despite substantial experimental evidence—met with skepticism (Schulte and Bryja, 2007; Schulte, 2010; Malbon, 2011; Schulte and Kozielewicz, 2019). The large amount of SMO structures in inactive states as well as the first crystal structure of a FZD in the apo (ligand-free) state were recently complemented with two mammalian SMO structures in the active state. Despite the different experimental approaches used to resolve SMO activation by: 1) cryo-electron microscopy imaging of a sterol ligand-bound human SMO in complex with heterotrimeric Gi [Protein Data Bank identification (PDB ID): 6OT0] (Qi et al., 2019) and 2) X-ray crystallography of an agonist (SAG21k)-bound human SMO in complex with a stabilizing nanobody (PDB ID: 6O3C) (Deshpande et al., 2019), both structures support a similar concept of receptor activation, the involvement of a common molecular switch mechanism, and overall similar structural rearrangements. In comparison with active, G protein-bound class A and B receptors, the active SMO structures also present an outward movement of TM6 (Fig. 1) (Wacker et al., 2010; Rasmussen et al., 2011; Hua et al., 2016; Song et al., 2017; Zhang et al., 2017c; Liang et al., 2018a,b; Yang et al., 2018; Deshpande et al., 2019; Krishna Kumar et al., 2019; Qi et al., 2019). This outward movement in the active state is a consequence of ligand-induced and G protein–stabilized structural rearrangements breaking interactions in conserved motifs, such as the D(E)RY motif in class A and a polar network in class B receptors. However, the mode of conformational change observed in active SMO is slightly different compared with class A/B receptors. In particular, there is a notable 7 to 8 Å outward movement of TM6 and a 4 to 5 Å inward movement of TM5. The larger changes on the intracellular facing part of the receptor are mirrored by minor rearrangements in the extracellular part. The active, NbSmo8-bound SMO presents with upward displacement of TM6 and extracellular loop 3 and a minor tilt in the CRD compared with inactive SMO (Deshpande et al., 2019). Similarly, the Gi-bound SMO presents with a TM6 rearrangement manifesting in outward movement and an upward shift of TM6 (Qi et al., 2019). Interestingly, TM6 appears to link CRD and the intracellular regions that undergo larger conformational changes physically, thereby potentially explaining complex allosteric modulation of receptor activity by the CRD itself and molecules interacting with the CRD and lower binding pockets. As mentioned previously, the structures of the active state of SMO confirm the concept of a common molecular switch in class F receptors, which we identified recently (Wright et al., 2019). The polar interactions between the conserved basic residue R/K6.32 and the backbone of TM7 that are common in inactive class F receptor structures are broken in the active SMO structures as a result of the outward movement of TM6 (Deshpande et al., 2019; Qi et al., 2019; Wright et al., 2019). Furthermore, the distance between R/K6.32 and the aromatic electron system of W7.55 is increased, indicating also that the π-cation interactions as part of the molecular switch are weakened (Deshpande et al., 2019; Qi et al., 2019).
Furthermore, the SMO-Gi structure revealed a different arrangement of the α5 helix of the Gi protein that leads to a less-pronounced TM6 outswing compared with the Gi-coupled class A receptor (e.g., cannabinoid 1 receptor; PDB ID: 6N4B) (Krishna Kumar et al., 2019). Interestingly, the SAG21k- and cholesterol-bound SMO (PDB ID: 6O3C) (Deshpande et al., 2019) represents a unique ligand-bound receptor structure in which three ligands are bound simultaneously to a single receptor. The SAG21k occupies the upper ligand binding site in the 7TM core just on top of a cholesterol bound to the lower binding site in the 7TM core of SMO. Furthermore, a second cholesterol occupies the lipophilic binding groove in the CRD (Fig. 2). Thus far, the majority of the cyclopamine- or sterol-bound SMO structures solved (Wang et al., 2013, 2014; Weierstall et al., 2014; Byrne et al., 2016; Zhang et al., 2017b; Huang et al., 2018) differ substantially with regard to ligand binding poses since these small molecules have been reported to bind the CRD only, the CRD and the upper 7TM core site, the CRD and the lower 7TM core site, or only the upper 7TM core site (Fig. 2). It cannot be ruled out that these differences reflect different purification and reconstitution approaches used in the various studies, but it also underlines the high complexity of SMO receptor activation mechanisms (Luchetti et al., 2016; Myers et al., 2017; Hedger et al., 2019). As opposed to other GPCRs, it still remains obscure which binding sites on SMO present orthosteric or allosteric sites, and what the functional role of the CRD could be as an allosteric modulator (Byrne et al., 2016; Kozielewicz et al., 2019). Interestingly, additional sites have been implicated, such as a cytoplasmic binding pocket comprised of cytoplasmic facing portions of TM1, TM3, TM6, and TM7, which was identified by docking of oxysterols to human SMO (Raleigh et al., 2018). Along these lines, the nature of endogenous SMO ligands, cholesterol, and naturally occurring oxysterols is also not fully understood since these molecules may serve as orthosteric agonists or allosteric modulators. Interestingly, there is a possibility that under certain cellular conditions with sufficiently high cholesterol levels, SMO signaling may occur, in fact, in the absence HH stimulation.

SMO contains (at least) three small molecule binding pockets. The pharmacology of SMO is complex. The receptor model was derived from the active, NbSmo8-bound crystal structure of human SMO (PDB ID: 6O3C) bound to two cholesterol molecules and a small molecule agonist called SAG21k (Deshpande et al., 2019). The full-length receptor is shown as a cartoon with round helices and an opaque surface. The N-terminal CRD is shown in red. The structured linker domain is shown in green. Ligands are shown as spheres in yellow and the three main ligand-binding sites are encircled in bright yellow. The gray part of the receptor presents the transmembrane domain. A clear distinction of orthosteric and allosteric sites is thus far not possible because the endogenous ligand of SMO and its primary binding site are not identified. In addition, cooperativity between binding sites remains obscure. PyMOL (The PyMOL Molecular Graphics System, version 2.0; Schrödinger, LLC) was used for visualization.
The CRD appeared flexible in the SMO-Gi complex, resulting in poor resolution by cryo-electron microscopy not allowing secure conclusions regarding its position in the ternary complex with heterotrimeric G protein (Qi et al., 2019). On the other hand, the CRD of human SMO is well resolved in the crystal structure of the SMO-NbSmo8 complex, indicating that the previously reported reorientation of the CRD in Xenopus leavis SMO does not accompany receptor activation (Huang et al., 2018; Deshpande et al., 2019; Schulte and Kozielewicz, 2019).
Pathway-Selective Conformations
Identification of a conserved molecular switch in class F receptors provided a molecular mechanism for constitutive and ligand-induced activation of class F receptors in agreement with what is known from other GPCRs (Trzaskowski et al., 2012; Wright et al., 2019). It is known that WNTs can induce WNT/β-catenin in parallel to β-catenin–independent signaling, for example, via WNT/extracellular signal–regulated kinases 1/2 signaling in mouse, primary microglia cells (Halleskog and Schulte, 2013), and other biologic systems (Schlange et al., 2007; Samarzija et al., 2009). Employing genome editing to create a cell system deficient in signaling through heterotrimeric G proteins, we have recently shown that WNT-3A maintains its ability to induce WNT/β-catenin signaling efficiently in the complete absence of heterotrimeric G protein signaling (Bowin et al., 2019). While these findings strongly argue against involvement of heterotrimeric G proteins in the WNT-induced WNT/β-catenin signaling, they further support the concept of conformation-driven, functional selectivity (Fig. 3). It is impossible for FZDs that carry a mutation in the conserved R/K6.32 molecular switch, which presents with enhanced potency toward G protein coupling, to interact with DVL and mediate DVL-dependent signaling pathways such as the WNT/β-catenin branch (Wright et al., 2019). Along the same line, we have previously described FZD mutations that exhibit selective DVL over G protein preference or vice versa (Kilander et al., 2014b; Strakova et al., 2017), underlining that pathway selection between G protein–dependent signaling and DVL-dependent pathways originates in a conformational selection in agreement with the recently developed concept of GPCR bias or functional selectivity (Kenakin, 2019). Especially, the Y250F2.39 mutation in FZD4, which presents with impaired ability to recruit DVL, is conceptually of interest. This residue is engaged in a polar network with H3484.46 and W3524.50 stabilizing an overall receptor conformation, which is essential for effective FZD-DVL interaction (Strakova et al., 2017; Yang et al., 2018). While mutation of this residue interferes with FZD-DVL interaction, FZD4 Y250F2.39 maintains its ability to interact with heterotrimeric G proteins, underlining that distinct conformations are required for complex formation with either DVL or heterotrimeric G protein (Arthofer et al., 2016; Strakova et al., 2017). This argumentation and the similarity between the SMO-NbSmo8 and SMO-Gi complex structures also further support the implication by Deshpande et al. (2019) that NbSmo8 indeed stabilizes the active, G protein–bound SMO. Looking forward, it will now be interesting to explore if there are alternative active conformations for SMO, for example, in complex with β-arrestin or GPCR kinases (Chen et al., 2004b; Zhao et al., 2016; Schulte and Wright, 2018). Excitingly, the concept of conformational selection comes with opportunities for biased ligands to target class F receptors in a pathway-selective manner similar to what has been successful in the case of class A GPCRs (Kenakin, 2019).

The concept of conformational selection and functional selectivity. In accordance with what is known for class A and B receptors (Latorraca et al., 2017; Kenakin, 2019), we present here how conformational selection either driven by endogenous ligands of the receptors, selective class F receptor-targeting drugs, intrinsic constitutive activity, or intracellular binding partners can contribute to pathway-selective signal initiation. Colors highlight receptor conformations that are stabilized by interaction, for example, with heterotrimeric G proteins, DVL, or arrestins (Schulte and Wright, 2018). Other, yet unidentified binding partners could stabilize additional active states of class F receptors. The theoretical number of active conformations is unlimited. FZD6 models (Wright et al., 2019) were modified using PyMOL (The PyMOL Molecular Graphics System, version 2.0; Schrödinger, LLC). The CRD is not shown.
Conclusions and Future Perspectives
This review aims to summarize the state of the art of our understanding of class F receptor activation fueled by the recent structural and functional dissection of active SMO in a ternary complex (Deshpande et al., 2019; Qi et al., 2019; Wright et al., 2019). The implications of this more detailed insight into SMO and FZD activation for understanding class F receptor biology, disease-associated mutations in class F receptors, and future efforts to develop more selective, potentially pathway-biased class F receptor-targeting drugs are exciting and promising. While some general concepts and molecular mechanisms of receptor activation emerge, several resulting key questions remain. How is it possible to assess the contribution of the different small molecule ligand binding pockets on SMO (Fig. 2) for receptor activation and how can that knowledge be exploited to target SMO therapeutically, potentially avoiding resistance-prone mutations? How are the different sterol-targeted binding sites driving receptor activation in vivo and how does that knowledge translate to FZDs regarding the localization and function of the lipid modification of WNTs for ligand binding and receptor activation? How does WNT binding to the CRD drive receptor activation? How does the CRD contribute to agonist-induced and constitutive receptor activation? How does the accessibility and local concentration of intracellular binding partners contribute to ligand-induced pathway selection? With the advances of structural analysis of TM receptor complexes using, for example, cryo-electron microscopy, gene editing technology, and the optimization of sensitive resonance energy transfer assays to assay protein-protein interactions and receptor dynamics in living cells, future work will shed even more light on the activation mechanisms defining this enigmatic receptor family. The hope is that the new knowledge will not only result in better understanding of the biology of class F receptors but also in assay development for innovative drug screening technologies, efficient drug discovery of class F–targeting, and potentially pathway-biased compounds.
Acknowledgments
The authors thank the Swedish National Infrastructure for Computing–National Supercomputer Centre in Linköping, Sweden, and High Performance Computing Centre North in Umeå, Sweden, for providing the computational resources for the work in the Schulte Laboratory.
Authorship Contributions
Participated in research design: Kozielewicz, Turku, Schulte.
Wrote or contributed to the writing of the manuscript: Kozielewicz, Turku, Schulte.
Footnotes
- Received July 29, 2019.
- Accepted September 25, 2019.
The work in the Schulte Laboratory was financially supported by the Karolinska Institutet, the Swedish Research Council [Grant 2017-04676], Swedish Cancer Society [Grant CAN2017/561], Novo Nordisk Foundation [Grant NNF17OC0026940], Stiftelsen Olle Engkvist Byggmästare [Grant 2016/193], Wenner-Gren Foundations [Grant UPD2018-0064], and Emil and Wera Cornells Stiftelse.
Abbreviations
- CRD
- cysteine-rich domain
- DVL
- dishevelled
- FZD
- Frizzled
- GLI
- glioma-associated homolog
- GPCR
- G protein-coupled receptor
- HH
- Hedgehog
- IL
- intracellular loop
- LRP5/6
- lipoprotein receptor-related protein 5/6
- PCP
- planar cell polarity
- PDB ID
- Protein Data Bank identification
- SAG
- smoothened agonist
- SMO
- Smoothened
- TM
- transmembrane
- WNT
- Wingless/Int-1
- Copyright © 2020 by The American Society for Pharmacology and Experimental Therapeutics
References
In this issue





{kind=link}
{kind=link}
{kind=link}
Jump to section
- Article
- Abstract
- Introduction
- Class F Receptors and Their Ligands
- A Short Overview of Molecular Mechanisms of WNT and HH Signaling
- Frizzleds: Ligand-Receptor Interaction
- SMO: Ligand-Receptor Interaction—a Mystery
- Signal Initiation: Different Concepts
- SMO in Ternary Complex with Heterotrimeric Gi or Nanobody NbSmo8
- Pathway-Selective Conformations
- Conclusions and Future Perspectives
- Acknowledgments
- Authorship Contributions
- Footnotes
- Abbreviations
- References
- Figures & Data
- Info & Metrics
- eLetters