


Visual Overview

Abstract
Alcohol dehydrogenases (ADHs) and aldehyde dehydrogenases (ALDHs) are vital enzymes involved in the metabolism of a variety of alcohols. Differences in the expression and enzymatic activity of human ADHs and ALDHs correlate with individual variability in metabolizing alcohols and drugs and in the susceptibility to alcoholic liver disease. MicroRNAs (miRNAs) function as epigenetic modulators to regulate the expression of drug-metabolizing enzymes. To characterize miRNAs that target ADHs and ALDHs in human liver cells, we carried out a systematic bioinformatics analysis to analyze free energies of the interaction between miRNAs and their cognate sequences in ADH and ALDH transcripts and then calculated expression correlations between miRNAs and their targeting ADH and ALDH genes using a public data base. Candidate miRNAs were selected to evaluate bioinformatic predictions using a series of biochemical assays. Our results showed that 11 miRNAs have the potential to modulate the expression of two ADH and seven ALDH genes in the human liver. We found that hsa-miR-1301-3p suppressed the expression of ADH6, ALDH5A1, and ALDH8A1 in liver cells and blocked their induction by ethanol. In summary, our results revealed that hsa-miR-1301-3p plays an important role in ethanol metabolism by regulating ADH and ALDH gene expression.
SIGNIFICANCE STATEMENT Systematic bioinformatics analysis showed that 11 microRNAs might play regulatory roles in the expression of two alcohol dehydrogenase (ADH) and seven aldehyde dehydrogenase (ALDH) genes in the human liver. Experimental evidences proved that hsa-miR-1301-3p suppressed the expression of ADH6, ALDH5A1, and ALDH8A1 in liver cells and decreased their inducibility by ethanol.
Introduction
Upon consumption, 80% of ethanol is absorbed in the upper small intestine and then transported to other organs. The liver is the primary site of ethanol metabolism. Excessive ethanol consumption is a common cause of liver injury and alcoholic liver disease (ALD) that encompasses liver manifestations, including fatty liver, fibrosis, cirrhosis, or even hepatocellular carcinoma (Adachi and Brenner, 2005; Tilg and Day, 2007). In China, the prevalence of ALD has increased considerably in recent years, which is correlated with an increase in alcohol consumption (Heo et al., 2019).
The major two-step oxidative metabolic pathway for ethanol begins with its oxidation to acetaldehyde with further oxidation to produce acetic acid (Dinis-Oliveira, 2016). The oxidation of ethanol to acetaldehyde is catalyzed by alcohol dehydrogenases (ADHs) and CYP2E1; this constitutes the rate-limiting step of ethanol metabolism. Subsequently, acetaldehyde is further oxidized to acetic acid by aldehyde dehydrogenases (ALDHs) using NAD as the cofactor, which is followed by elimination of the acetic acid end product through urine.
Obvious individual variability in ethanol metabolism is noted among humans, and this is partially attributed to differences in genetic background, age, sex, and health status (Liangpunsakul et al., 2016). For example, studies showed that functional genetic variants in ADH and ALDH genes affected the activities or the expression levels of ethanol-metabolizing enzymes, thus varying the catabolic rate of ethanol (Birley et al., 2009; Hartwell and Kranzler, 2019).
ADH and ALDH proteins represent two distinct families of enzymes. Specifically, the ADH family consists of multiple isozymes and allozymes that are divided into five (I-V) classes (Cederbaum, 2012), whereas the ALDH superfamily includes 19 functionally related family members (Vasiliou and Nebert, 2005). In addition to the primary function of ADHs and ALDHs members (especially ADHs and ALDH2) in ethanol metabolism, ADHs and ALDHs are also involved in metabolizing a large spectrum of biologically important alcohols and aldehydes, including drugs (Jelski and Szmitkowski, 2008).
The complexity of regulatory mechanisms underlying ethanol metabolism is not fully understood. Epigenetic regulatory mechanisms, including DNA methylation, noncoding RNAs, and histone modification, provide novel genotype-independent mechanisms to modulate gene expression, contributing to individual variability in the metabolism and toxicity of ethanol and other chemicals (Rodenhiser and Mann, 2006; Moss and Wallrath, 2007; Davison et al., 2009; Meng et al., 2012; McDaniel et al., 2014). The microRNAs (miRNAs), a class of noncoding single-stranded RNA molecules typically about 22 nucleotides in length, are well-recognized as important epigenetic mediators that regulate the expression of drug-metabolizing enzymes and nuclear receptors (Song and Wang, 2008; Yu, 2009; Nakajima and Yokoi, 2011; Dluzen and Lazarus, 2015). MiRNAs bind to the mRNA transcripts of target genes by partial sequence complementarity to miRNA response elements (MREs) that are most often located in the 3′-untranslated region (UTR) of target transcripts. Interactions between miRNAs and MREs generally result in either the enhanced degradation of mRNA target transcripts or decreased efficiency of translation.
Previous studies showed that ALDH5A1 expression is suppressed by the miRNA hsa-miR-29a-3p and that CYP2E1 expression is modulated by multiple miRNAs (Yu et al., 2015c; Wang et al., 2017). Specifically, miR-378a-5p and miR-214-3p bind to the 3′-UTR of CYP2E1 mRNA transcripts, causing decreased CYP2E1 protein expression and enzymatic activity (Mohri et al., 2010; Wang et al., 2017), whereas miR-552 suppresses CYP2E1 protein production by interacting with both the 3′-UTR and the promoter region of CYP2E1 (Miao et al., 2016). Although a few studies have reported associations between miRNAs and CYP2E1 and ALDH5A1 genes, to our knowledge no systematic analysis has been carried out to elucidate the functional roles of miRNAs in the expression of other genes in the catabolic pathway of alcohol, notably other ADH and ALDH genes expressed in the liver.
In this study, we selected the ADH and ALDH genes expressed in the human liver and predicted potential miRNA-binding sites located in the 3′-UTRs of these target genes via in silico analyses. Furthermore, we identified hsa-miR-1301-3p as a candidate regulatory miRNA associated with ethanol metabolism. Finally, we tested the regulatory role of hsa-miR1301-3p on the suppression of ADH6, ALDH5A1, and ALDH8A1 using a series of in vitro and in vivo experimental approaches. Our results provided new clues to elucidate the epigenetic regulatory mechanisms influencing the expression of ethanol-metabolizing genes, thus adding a new layer of information toward understanding the interindividual variability in ethanol-induced liver injury and ALDs.
Materials and Methods
Chemicals and Reagents.
HepG2, Huh7, and 293T cells were obtained from the American Type Culture Collection (Manassas, VA). The hsa-miR-1301-3p mimic and miRNA negative control (NC) were purchased from Ribo Life Science (Shanghai, China). All oligonucleotides and primers used in our study were obtained from Sangon Biotech (Shanghai, China). All reporter gene vectors were produced by Generay Biotech (Shanghai, China). Rabbit anti-human antibodies against ADH6, ALDH5A1, and ALDH8A1 proteins and mouse anti-human antibodies against glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were obtained from Abcam (Cambridge, MA). Dual-Luciferase Reporter 1000 Assay System and TRIzol Reagent were purchased from Promega (Madison, WI). QuantiTect Reverse Transcription kit and Quanti Fast TB Green Real-Time Polymerase Chain Reaction (RT-PCR) kit were obtained from Qiagen (Valencia, CA). NCode miRNA First-Strand cDNA Synthesis kit, radioimmunoprecipitation assay buffer, and LipofectAMINE 2000 reagent were purchased from Life Technologies (Carlsbad, CA). Odyssey Western Blotting Kit was purchased from LI-COR Biosciences (Lincoln, NE). All other reagents were of analytical grade.
In Silico Analyses.
Nine ADH and 15 ALDH genes (Yang et al., 2013) were screened using mRNA data in 98 human liver tissues from The Cancer Genome Atlas (TCGA) data base (https://www.cancer.gov/about-nci/organization/ccg/research/structural-genomics/tcga) to select the alcohol-metabolizing enzymes expressed in human liver. The miRNAs potentially targeting alcohol-metabolizing genes were predicted using the miRTar.human data base (http://mirtar.mbc.nctu.edu.tw/human/), and the free energy of miRNA:mRNA duplexes was calculated by RNAhybrid algorithm (http://bibiserv2.cebitec.uni-bielefeld.de/rnahybrid), respectively. The correlations between alcohol metabolism genes and miRNAs were calculated by Pearson correlation analysis based on their RNA levels in liver tissues from TCGA data base.
Cell Culture, Transfection, and Ethanol Treatments.
HepG2, Huh7, and 293T cells were cultured in Dulbecco’s modified Eagle’s medium supplemented with 10% FBS at 37°C in a humidified 5% CO2 atmosphere. All cell lines were used at fewer than 10 passages at the time of the study.
The hsa-miR-1301-3p mimic and miRNA NC were transiently transfected into HepG2 and Huh7 cells at the final concentration of 20 nM using LipofectAMINE 2000 reagent. Total RNAs and proteins were extracted at 24 or 48 hours after transfection experiments. In ethanol exposure experiments, HepG2 and Huh7 cells transfected with miRNA mimics were cultured for 24 hours, treated with 50 mM ethanol for another 24 hours, and then harvested to obtain total RNAs and proteins for subsequent analyses. All experiments were carried out at least three times.
Fluorescent-Based RNA Electrophoretic Mobility Shift Assay.
Oligonucleotides and primers used in our study were obtained from Sangon Biotech, and their sequences were listed in Supplemental Table 1. The oligonucleotide for hsa-miR-1301-3p was dye-miR-1301-3p, which was synthesized and 5′-modified using DyLight 800 dye. The RNA oligonucleotides corresponding to the MREs of hsa-miR-1301-3p resident in 3′-UTR of ADH6, ALDH5A1, and ALDH8A1 were 5′-modified using Cy5.5 dye and designated as dye-ADH6, dye-ALDH5A1, and dye-ALDH8A1, respectively. In addition, 50-fold molar excesses of unlabeled oligonucleotides for negative control were included in competition assays.
Fluorescent-based RNA electrophoretic mobility shift assay (FREMSA) was carried out according to the protocol described in our previous report (Yu et al., 2020). Briefly, 200 fmol dye-miR-1301-3p and cognate dye-ADH6, dye-ALDH5A1, and dye-ALDH8A1 oligonucleotides were mixed in basic buffer containing 10 mM HEPES buffer (pH 7.3), 0.5% glycerol, 20 mM KCl, and 10 mM MgCl2, respectively. The mixtures were incubated for 20 minutes at room temperature and then separated by 10% native PAGE at 4°C. Interaction signals were detected using the Odyssey CLx Infrared Imaging System (LI-COR Biosciences).
Luciferase Reporter Gene Assays.
The core 3′-UTRs containing the MREs of hsa-miR-1301-3p located at the 3′-UTRs of ADH6, ALDH5A1, and ALDH8A1 were synthesized chemically; subcloned into the pMirGlo system; and designated as ADH6-WT, ALDH5A1-WT, and ALDH8A1-WT, respectively, to create wild-type reporter gene vectors. Furthermore, the mutant sequences that contained mutations in the MREs of hsa-miR-1301-3p in ADH6, ALDH5A1, and ALDH8A1, which obviously increased free energy of miRNA:mRNA duplexes, were synthesized, subcloned into the pMirGlo system, and designated as ADH6-Mut, ALDH5A1-Mut, and ALDH8A1-Mut, respectively. All resultant constructs were sequenced to confirm their authenticity.
293T cells were seeded into 96-well plates at a density of 3 × 104 cells/well and cultured till reaching approximately 80% confluence. Cells were transfected with constructed reporter gene vectors (100 ng/well) together with hsa-miR-1301-3p mimics (final concentration: 50 nM) or miRNA NC (final concentration: 50 nM), using the LipofectAMINE 2000 reagent. Both Firefly and Renilla luciferase activities were measured at 24 hours after transfection using the Dual-Luciferase Reporter 1000 Assay System, and transfection efficiencies were normalized to Renilla luciferase activity. Three independent experiments were conducted in triplicate.
RNA Extraction and Quantitative RT-PCR.
The TRIzol Reagent was used to extract total RNAs from HepG2 and Huh7 cells. Reverse transcription reactions of mRNA or miRNA were conducted using QuantiTect Reverse Transcription kit or NCode miRNA First-Strand cDNA Synthesis kit, respectively. The quantitative RT-PCR assays were carried out using Quanti Fast TB Green RT-PCR kit with the LightCycler 480 Detection System (Roche, Basel, Switzerland), and the primer pairs ADH6-F with ADH6-R, ALDH5A1-F with ALDH5A1-R, ALDH8A1-F with ALDH8A1-R, GAPDH-F with GAPDH-R, miR-1301-3p-F with the reverse primer supplemented in the NCode miRNA First-Strand cDNA Synthesis kit, and U6-F with U6-R were designed to detect RNA levels for ADH6, ALDH5A1, ALDH8A1, GAPDH, hsa-miR-1301-3p, and the small nuclear RNA U6 control, respectively. The fold changes for ADH6, ALDH5A1, or ALDH8A1 mRNA levels were calculated relative to GAPDH, whereas the fold change of hsa-miR-1301-3p was relative to snRNA U6 (Jiang et al., 2005). All experiments were carried out at least three times.
Western Blotting.
Radioimmunoprecipitation assay buffer was used to extract total proteins from HepG2 or Huh7 cells. Western blotting assays were carried out using the Odyssey Western Blotting Kit and analyzed using the Odyssey CLx Infrared Imaging System. All experiments were carried out at least three times.
Statistical Analyses.
One-way ANOVA on ranks test was used to test the differences between subgroups in luciferase assays and RNA or protein experiments, respectively. Data are shown as mean ± S.D. in the bar graphs, and P values of 0.05 were used as the cutoffs for statistical significance.
Results
Selection of miRNAs Potentially Targeting ADH and ALDH Genes.
RNA expression profiles of nine ADH and 15 ALDH genes (Yang et al., 2013) in human liver tissues were obtained from the TCGA data base. Among these genes, seven ADH genes, including ADH1A, ADH1B, ADH1C, ADH4, ADH5, ADH6, and ADHFE1 (Fig. 1A), and 14 ALDH genes, including ALDH1A1, ALDH1A2, ALDH1A3, ALDH1B1, ALDH2, ALDH3A1, ALDH3A2, ALDH3B1, ALDH4A1, ALDH5A1, ALDH6A1, ALDH7A1, ALDH8A1, and ALDH9A1 (Fig. 1B), were observed to be variably expressed in human liver. ADH1B and ALDH2 exhibited the highest expression, whereas ADHFE1 and ALDH1A2 were the ones with the lowest expression among the ADH and ALDH genes. Intriguingly, five ADH genes, including ADH1A, ADH1B, ADH1C, ADH4, and ADH6, and four ALDH genes, including ALDH1A1, ALDH3A1, ALDH1A3, and ALDH8A1 presented a wide range of expression levels in liver samples, indicating the individual variability in ethanol metabolism.
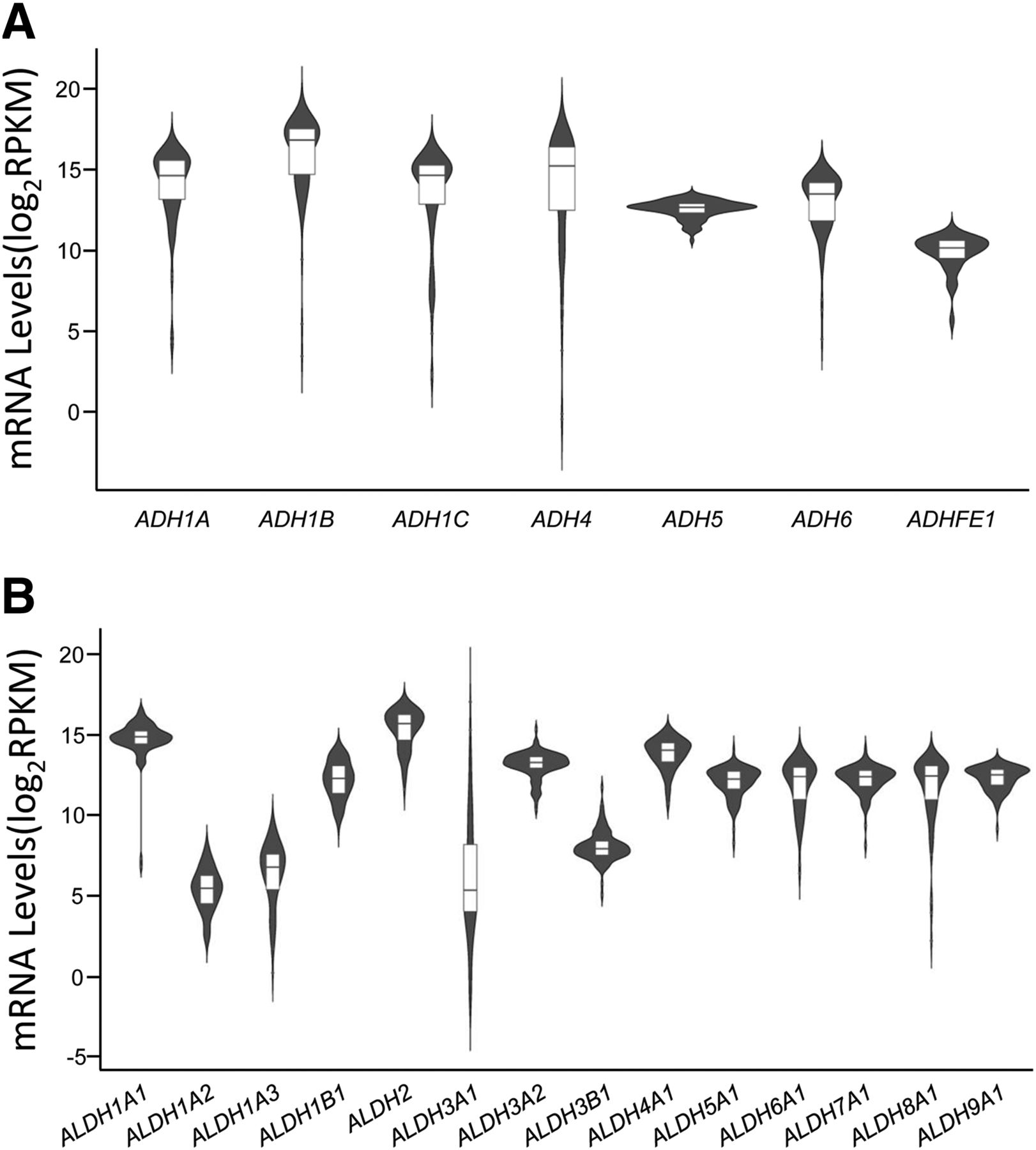
Expression status of ADH (A) and ALDH (B) genes in a liver data set obtained from the TCGA data base. RNA levels of nine ADH and 15 ALDH genes were obtained from 98 liver tissues from the TCGA data base. RPKM, reads per kilobase per million mapped reads.
We predicted the miRNAs that are potentially able to bind to the 3′-UTRs of seven ADH genes and 14 ALDH genes using the miRTar.human data base and then calculated the correlations between the expression of potential candidate miRNAs and the expression of their cognate targeting genes based on the liver expression data set from TCGA data base. As shown in Table 1, when the selection criteria were set as: 1) the free energy of binding is smaller than −20.0 kcal/mol and 2) the R between the expression of the miRNA and the expression of its target gene is smaller than −0.25 (r < −0.25) (Yu et al., 2015c), 11 mature miRNAs were considered as potential epigenetic modulators of ADH4, ADH6, ALDH1A3, ALDH1B1, ALDH3A2, ALDH4A1, ALDH5A1, ALDH8A1, and ALDH9A1, respectively.
miRNAs potentially targeting ADH and ALDH genes
Compared with other miRNAs, hsa-miR-1301-3p, hsa-miR-330-5p, and hsa-miR-93-5p showed stronger negative correlations (r < −0.4) (Akoglu, 2018) with the ADH or ALDH genes. Among which, hsa-miR-1301-3p exhibited the highest inverse correlations with mRNAs for ALDH8A1 (r = −0.480), ADH6 (r = −0.469), and ALDH5A1 (r = −0.459) and relatively strong binding affinities with its cognate MRE targets resident in these genes, suggesting its increased regulatory efficiency over the ethanol metabolism process. We then selected hsa-miR-1301-3p and its target genes, including ADH6, ALDH5A1, and ALDH8A1, for focused experiments to test the reliability of miRNA-gene interaction predictions made using bioinformatics (Table 1).
hsa-miR-1301-3p Interacted with ADH6, ALDH5A1, and ALDH8A1 Transcripts.
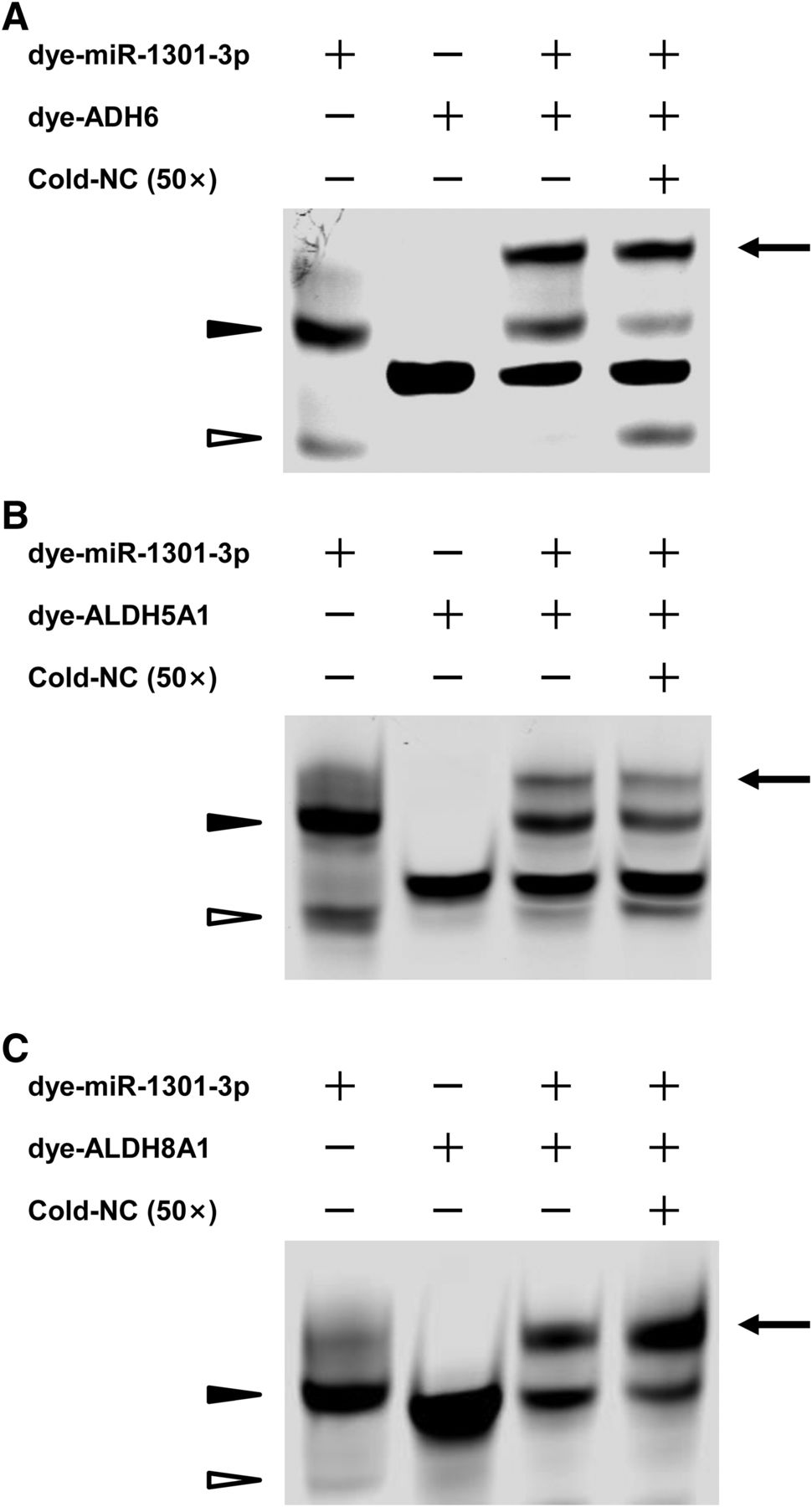
FREMSAs were applied to show the direct interactions between hsa-miR-1301-3p and its cognate mRNA targets in vitro (Fig. 2). A strong band formed by dimeric dye-miR-1301-3p was observed in the absence of any targets (Fig. 2, lane 1), which was due to the potential interactions among the dye-miR-1301-3p oligonucleotides (−20.2 kcal/mol, Supplemental Fig. 1). When the dye-ADH6, dye-ALDH5A1, and dye-ALDH8A1 oligonucleotide were added, the dye-miR-1301-3p displayed the mobility shift (the top band in Fig. 2, lane 3), and the bands formed by dimeric dye-miR-1301-3p were significantly reduced, respectively, because the mRNA oligonucleotides have the stronger binding ability to dye-miR-1301-3p (−28.3, −26.8, and −30.6 kcal/mol, respectively). The competition assays proved the sequence-specific interactions between hsa-miR-1301-3p and its cognate mRNA targets on ADH6, ALDH5A1, and ALDH8A1. The excess amount of unlabeled oligonucleotides for negative control failed to totally eliminate the densities of the miRNA:RNA complexes formed by dye-miR-1301-3p probe and its MREs in ADH6 and ALDH5A1, respectively (Fig. 2, A and B, lane 4). The nonspecific competitor enhanced the miRNA:RNA complexes formed by dye-miR-1301-3p and dye-ALDH8A1 oligonucleotides (Fig. 2C, lane 4); however, the reasons for this enhancement remain to be elucidated (Wolfgang et al., 1997; Yu et al., 2018).

The hsa-miR-1301-3p interacted with ADH6 (A), ALDH5A1 (B), and ALDH8A1 (C) mRNA in vitro. Lanes 1 and 2 indicated the mobility of dye-miR-1301-3p and dye-ADH6, dye-ALDH5A1, and dye-ALDH8A1 oligonucleotides, respectively; lane 3 indicated the mobility status of the miRNA:mRNA complex formed by dye-miR-1301-3p oligonucleotides with dye-ADH6, dye-ALDH5A1, and dye-ALDH8A1 mRNA oligonucleotides, respectively; lane 4 revealed the mobility shift status of miRNA:mRNA complex in the presence of excess unlabeled nonspecific competitors. NC, nonspecific competitor. The arrow, miRNA:mRNA complex; hollow triangle, monomeric dye-miR-1301-3p; solid triangle, dimeric dye-miR-1301-3p; experiments were carried out at least three times. WT, wild type.
hsa-miR-1301-3p Suppressed Luciferase Activity Produced by MRE Sequences Derived from ADH6, ALDH5A1, and ALDH8A1 3′-UTRs.
Core sequences containing MREs or mutants within the MREs located at 3′-UTRs (Fig. 3A) of ADH6, ALDH5A1, or ALDH8A1 were introduced into luciferase reporter gene vectors and then transfected into 293T cells together with hsa-miR-1301-3p mimics, respectively. As shown in Fig. 3B, hsa-miR-1301-3p mimics dramatically reduced reporter gene activity for constructs containing the MREs derived from the 3′-UTRs of ADH6 (51.5%, P < 0.01), ALDH5A1 (38.9%, P < 0.01), and ALDH8A1 (37.3%, P < 0.01) compared with that in cells treated with the miRNA negative control. Constructs containing mutated MREs for hsa-miR-1301-3p in ADH6, ALDH5A1 and ALDH8A1 (Fig. 3A) were also transfected into 293T cells together with hsa-miR-1301-3p mimics; however, no inhibitory effects were observed. All these evidences proved that hsa-miR-1301-3p was able to bind to its MREs in ADH6, ALDH5A1, and ALDH8A1 3′-UTRs.

The hsa-miR-1301-3p inhibited luciferase reporter gene expression. (A) Free energy analyses of miRNA:mRNA duplex formed by hsa-miR-1301-3p and core or mutated response elements in 3′-UTRs of ADH6, ALDH5A1, and ALDH8A1, respectively, by the RNAhybrid algorithm. (B) Constructs containing the wild-type or mutated sequence of the core 3′-UTRs of ADH6, ALDH5A1, and ALDH8A1 genes were transiently transfected into 293T cells, respectively, together with 50 nM hsa-miR-1301-3p mimic or miRNA negative control. Cells were harvested at 24 hours after transfection. Three independent experiments were conducted in triplicate, and data were shown as mean ± S.D. **P < 0.01 vs. miRNA negative control.
In addition, ALDH5A1 was also predicted to be modulated by another five miRNAs, including hsa-miR-149-3p, hsa-miR-149-5p, hsa-miR-18a-5p, hsa-miR-330-5p, and hsa-miR-93-3p. Among these, hsa-miR-330-5p and hsa-miR-149-3p exhibited the lower free energies of miRNA:mRNA duplexes (less than −30.0 kcal/mol) compared with the other miRNAs, indicating their stronger abilities to bind to the corresponding MREs, respectively. To further validate the reliability of miRNA-gene interaction predictions, hsa-miR-330-5p was selected, and its interaction with ALDH5A1 3′-UTR was proved by both FREMSA and luciferase reporter gene assays (Supplemental Fig. 2).
hsa-miR-1301-3p Suppressed Endogenous ADH6, ALDH5A1, and ALDH8A1 Production in Hepatic Cells.
To investigate the regulatory roles of hsa-miR-1301-3p in the expression of endogenous ADH6, ALDH5A1, and ALDH8A1, we transfected hsa-miR-1301-3p mimics or the miRNA negative control into HepG2 and Huh7 cells and then measured the RNA and protein levels for ADH6, ALDH5A1, and ALDH8A1 at 24 hours after transfection. Compared with HepG2 and Huh7 cells transfected with the miRNA negative control, cells transfected with hsa-miR-1301-3p mimics showed elevated levels of hsa-miR-1301-3p (Fig. 4A). Importantly, quantitative RT-PCR assays showed that mRNA levels for ADH6, ALDH5A1, and ALDH8A1 were statistically significantly suppressed in HepG2 and Huh7 cells treated with ectopic hsa-miR-1301-3p (Fig. 4B, 18%, 22%, and 33% in HepG2 cells and Fig. 4C, 34%, 37%, and 38% in Huh7 cells, respectively; all P < 0.05). Likewise, Western blot assays showed that endogenous ALDH5A1 and ALDH8A1 protein levels were reduced in hepatic cells treated with exogenous hsa-miR-1301-3p (Fig. 4D, ALDH5A1 and ALDH8A1 protein levels were 45% and 23% in HepG2 cells and 29% and 60% in Huh7 cells, respectively; all P < 0.05). Although hsa-miR-1301-3p transfection suppressed ADH6 protein levels in HepG2 cells (23%, P < 0.05), we were unable to demonstrate a statistically significant reduction in Huh7 cells, presumably because of reduced expression in this cell line under normal conditions.

The hsa-miR-1301-3p inhibited endogenous ADH6, ALDH5A1, and ALDH8A1 expression in HepG2 cells and Huh7 cells. The hsa-miR-1301-3p mimics or miRNA negative control (both at the final concentration of 20 nM) were transfected into HepG2 and Huh7 cells, respectively. (A) The transfection of hsa-miR-1301-3p mimics dramatically elevated the hsa-miR-1301-3p levels in both HepG2 and Huh7 cells, compared with the transfection of miRNA negative control. Downregulated mRNA levels of ADH6, ALDH5A1, and ALDH8A1 were observed after the transfection of hsa-miR-1301-3p mimics in HepG2 (B) or Huh7 cells (C). (D) Protein levels of ALDH5A1 and ALDH8A1 were statistically significantly suppressed by exogenous hsa-miR-1301-3p mimics in both HepG2 and Huh7 cells. ADH6 proteins were statistically significantly decreased by exogeneous hsa-miR-1301-3p in HepG2 cells, whereas the detection of ADH6 proteins failed in Huh7 cells under our experimental conditions. Each assay was carried out in triplicate, and data were shown as mean ± S.D. The fold changes of miRNA, mRNAs, and proteins induced by exogenous hsa-miR-1301-3p transfection were calculated by defining their levels in the cells transfected with miRNA negative control (miR-NC) as 1. *P < 0.05 vs. miRNA negative control. **P < 0.01 vs. miRNA negative control.
hsa-miR-1301-3p Suppressed Ethanol-Dependent Induction of ADH6, ALDH5A1, and ALDH8A1 Expression.
It is well-known that ADH and ALDH genes are inducible by ethanol exposure. We then investigated whether ectopic hsa-miR-1301-3p can suppress the induction of ADH6, ALDH5A1, and ALDH8A1 by ethanol. As shown in Fig. 5A, the cells exposed to 50 mM ethanol resulted in a reduction in the expression of endogenous hsa-miR-1301-3p (52% and 45% in HepG2 and Huh7 cells, respectively; both P < 0.05), whereas the transfection of ectopic hsa-miR-1301-3p dramatically elevated the level of hsa-miR-1301-3p even under ethanol exposure.

The hsa-miR-1301-3p suppressed ethanol-induced ADH6, ALDH5A1, and ALDH8A1 expression in HepG2 and Huh7 cells. (A) Ethanol exposure (at the final concentration of 50 mM) resulted in the reduction of endogenous hsa-miR-1301-3p levels in HepG2 and Huh7 cells. **P < 0.01 vs. untreated group. ##P < 0.01 vs. ethanol treatment group transfected with miRNA negative control. *P < 0.05 vs. ethanol treatment group transfected with hsa-miR-1301-3p mimics. (B) Exogenous hsa-miR-1301-3p was able to reduce the mRNA levels of ADH6, ALDH5A1, and ALDH8A1 that increased by ethanol exposure in HepG2 and Huh7 cells. *P < 0.05 vs. control group. #P < 0.05; ##P < 0.01 vs. ethanol treatment group transfected with miRNA negative control. (C) Exogenous hsa-miR-1301-3p was able to reduce the protein levels of ADH6, ALDH5A1, and ALDH8A1 that increased by ethanol exposure in HepG2 cells and able to decrease the proteins of ALDH5A1 and ALDH8A1 that increased by ethanol exposure in Huh7 cells. *P < 0.05; **P < 0.01 vs. control group. #P < 0.05 vs. ethanol treatment group transfected with miRNA negative control. Each assay was performed in triplicate.
The levels of ADH6, ALDH5A1, and ALDH8A1 mRNA transcripts were elevated upon ethanol exposure (1.19-fold, 1.35-fold, and 1.16-fold in HepG2 cells and 1.18-fold, 1.33-fold, and 1.12-fold in Huh7 cells, respectively; all P < 0.05), whereas they were statistically significantly decreased after transfection with hsa-miR-1301-3p mimics with ethanol exposure (decreased by 14%, 42%, and 13% in HepG2 cells and 30%, 11%, and 17% in Huh7 cells, respectively; all P < 0.05) (Fig. 5B). Similar regulatory trends were also observed in the protein levels for ADH6, ALDH5A1, and ALDH8A1 with the elevations upon ethanol exposure (1.62-fold, 1.27-fold, and 1.40-fold in HepG2 cells and 1.37-fold for ALDH5A1 and 1.39-fold for ALDH8A1 in Huh7 cells, respectively; all P < 0.05) and the reduction after transfection with hsa-miR-1301-3p mimics and ethanol exposure (reduced by 29%, 22%, and 14% in HepG2 cells and 25% for ALDH5A1 and 22% for ALDH8A1 in Huh7 cells, respectively; all P < 0.05) (Fig. 5C).
Discussion
ADHs and CYP2E1 catalyze the conversion of ethanol to acetaldehyde, and ALDHs catalyze the transformation of acetaldehyde to acetic acid; regulating the expression of these enzymes appropriately is vital for normal ethanol metabolism (Birley et al., 2009; Hartwell and Kranzler, 2019). The balance between the rates of accumulation and elimination of acetaldehyde (a known mutagen inducing DNA damage) is crucial for the pathologic processes associated with ethanol-induced hepatotoxicity, ALDs, and liver cancer (Tan et al., 2017).
Based on the results from a previous study (Yang et al., 2013) and in silico analysis, ADHs and ALDHs were found to be variably expressed in the human population, consistent with interindividual variability in alcohol and drug metabolism, and variable susceptibility toward ALDs among humans. Comprehensive mechanisms involved in the regulation of these enzymes are not fully understood, although it is presumed that genetic variations for ADH, ALDHs, and CYP2E1 contribute to interindividual variability in their expression among humans.
In this study, we applied a systematic approach using in silico bioinformatics methods to identify 11 miRNAs that could potentially regulate two ADH and seven ALDH genes in human liver based on in silico calculations of free energies of miRNAs binding to transcripts and inverse correlations between the expression of these miRNAs and their putative target genes using data from a publicly available data base. Of these 11 candidate regulatory miRNAs, hsa-miR-1301-3p was selected for focused study because the expression of this miRNA in human liver exhibited strong inverse correlations with the expression of several ethanol-metabolizing genes. Our study showed that hsa-miR-1301-3p suppresses the expression of ADH6, ALDH5A1, and ALDH8A1 genes in human hepatic cells with or without ethanol exposure using a series of biochemical assays. These results provide new insight into the epigenetic regulation of ethanol metabolism.
More than 100 algorithms (Henry et al., 2014) have been developed to predict potential mRNA targets for regulation by miRNAs. However, the failure of carefully controlled experiments performed in vitro to confirm predicted miRNA/mRNA target regulatory interactions reveals current limitations of in silico miRNA target predictions made without experimental validation (Thomas et al., 2010; Fan and Kurgan, 2015; Li and Zhang, 2015). Over the past several years we have successfully characterized and confirmed the biochemical functions of many miRNAs in the regulation of drug metabolizing enzymes and transporters (Yu et al., 2015a,b,c, 2018; Jin et al., 2016; Chen et al., 2017; Wang et al., 2017; Zeng et al., 2017; Knox et al., 2018) by integrating a combination of in silico, in vitro, and in vivo approaches to collect and analyze multiple layers of information.
In this study, in silico analyses predicted that 11 miRNAs, showing a high binding affinity with target sequences and a statistically significantly inverse correlation with their targets at the mRNA level, may potentially modulate two ADHs (ADH4 and ADH6) and seven ALDHs (ALDH1A3, ALDH1B1, ALDH3A2, ALDH4A1, ALDH5A1, ALDH8A1, and ALDH9A1) in the oxidative alcohol-aldehyde-acid metabolic pathway. Furthermore, hsa-miR-1301-3p was selected for focused study because the expression of this miRNA in the human liver exhibited the strongest inverse correlations with the expression of ADH6, ALDH5A1, and ALDH8A1 genes. Another important rationale for the selection of hsa-miR-1301-3p is that we recognize the benefits of “killing many birds with one stone”; in other words, a specific miRNA affecting multiple target genes within the same pathway should have an increased regulatory efficiency over that pathway. This rationale could be physiologically or pharmacologically significant for the major substrates of ADH6, ALDH5A1, and ALDH8A1 enzymes, including ethanol, succinic semialdehyde, or 2-amino-muconic semialdehyde (Marchitti et al., 2008).
Our FREMSA results showed the direct interactions between hsa-miR-1301-3p and its cognate targets in ADH6, ALDH5A1, and ALDH8A1 transcripts in vitro, and further competition assays and reporter gene assays proved the interactions were sequence-specific. Transfection assays showed that ectopic hsa-miR-1301-3p indeed suppressed endogenous ADH6, ALDH5A1, and ALDH8A1 production in liver cells. Together, our experimental evidence confirmed our prediction that hsa-miR-1301-3p interferes with the expression of these enzymes involved in alcohol and aldehyde metabolism.
Although some ADH and ALDH isoforms are inducible by alcohol and aldehyde, the functional significance of most ADH and ALDH enzymes in alcohol and aldehyde metabolism still needs to be elucidated. It was reported that the induction of class I ADH by ethanol involves liver transcriptional factors, such as CCAAT/enhancer-binding proteins (He et al., 2002) and sterol regulatory element-binding protein-1c (He et al., 2004). ADH6 may be regulated by hormonal changes or by exposure to xenobiotics because of the existence of glucocorticoid response elements (Yasunami et al., 1991) and other regulatory elements, such as CCAAT/enhancer-binding protein α (Humphrey and Kuciauskas, 2006) in its promoter region, but the induction of ADH6 by alcohol has not been noted previously. Similarly, although some isoforms of human ALDH are inducible, no previous study reported that ALDH5A1 and ALDH8A1 are induced by ethanol. It is worth noting that ethanol can stimulate the expression of ADH6, ALDH5A1, and ALDH8A1 genes, which is in company with a statistically significant reduction of endogenous hsa-miR-1301-3p levels in HepG2 and Huh7 cells by ethanol exposure, suggesting that the induction of ADH6, ALDH5A1, and ALDH8A1 may be mediated by the suppression of hsa-miR-1301-3p. Excessive ectopic hsa-miR-1301-3p could inhibit the induction of ADH6, ALDH5A1, and ALDH8A1 in liver cells, further echoing the suppression effect on ADH6, ALDH5A1, and ALDH8A1 by hsa-miR-1301-3p.
ADH6, which encodes a class V ADH, is mainly expressed in the liver. Many attempts to purify and characterize the human ADH6 protein have failed. Although computational studies predicted distinct structural changes existing in ADH6, compared with other ADH enzymes (Fagerberg et al., 2014), recombinant human ADH6 protein expressed in Escherichia coli cells showed comparable properties with other ADH isozymes (Chen and Yoshida, 1991). Under our experimental conditions, ADH6 protein was only detected in HepG2 cells but not in Huh7 cells, indicating that distinctive expression patterns are present in different human hepatic cell lines.
Both ALDH5A1 and ALDH8A1 belong to the superfamily of ALDHs. Besides metabolizing acetaldehyde (Muzio et al., 2012; Davis et al., 2018; Niimi et al., 2018), ALDHs were reported to modulate cell proliferation and differentiation by synthesizing retinoic acid, betaine, and γ-aminobutyric acid (Jackson et al., 2011). Deregulated ALDH5A1 expression occurred in a variety of cancers (Tian et al., 2017). The knockout of the ALDH5A1 gene in mice resulted in a significant reduction of glutathione, suggesting that ALDH5A1 might be associated with mitochondrial function (Sauer et al., 2007). ALDH8A1 was identified as a marker for liver diseases, including cirrhosis and liver cancer (Grinberg et al., 2014), but its functional significance still needs to be elucidated.
Our research strategy is associated with some limitations. First, we only focused on the miRNAs whose expression correlated inversely with the expression of ethanol-metabolizing enzymes in human liver, which is considered to be the classic regulatory mechanism for many miRNAs (i.e., a miRNA inhibits the expression of a target gene by binding to an MRE present in the 3′-UTR of the mRNA transcript). However, accumulating evidences proved that some miRNAs might increase gene expression by interacting with promoters, 5′-UTRs, and the protein-coding regions involving different molecular mechanisms (Ning et al., 2019). Second, we employed a strict threshold (−20 kcal/mol) for the free energy of binding between miRNA:RNA duplexes, which may result in the omission of some regulatory miRNAs that interact with targeting sequences via a non–seed-sequence-dependent manner (Piscioli et al., 1985). Third, it is well-known that one miRNA may target multiple genes, whereas one gene may be targeted by multiple miRNAs. Therefore, the overall effect of the 11 miRNAs, rather than one miRNA, may play an essential regulatory role in the metabolism of ethanol. In the future work, we would validate the regulatory effects of other 10 predicted miRNAs and measure the production of acetaldehyde and acetate, two useful benchmarks to test the activities of ADHs and ALDHs, respectively, to evaluate the overall regulatory roles of multiple miRNAs.
In summary, we predicted 11 regulatory miRNAs that may target ADH and ALDH genes in human liver based on systematic bioinformatic analyses and in vitro and in vivo experiments. We identified hsa-miR-1301-3p as a new epigenetic factor involved in the regulation of the alcohol-aldehyde-acetate metabolism pathway by suppression of ADH6, ALDH5A1, and ALDH8A1. Our results provide a new clue to interpret the interactions between miRNAs and ethanol metabolism.
Authorship Contributions
Participated in research design: Yu.
Conducted experiments: X. Wang, Y. Zhao, Luo, Xu, X. Li, Jin, Y. Wang.
Performed data analysis: C. Li, Feng, Chen, Hou, Q. Zhao, J. Zhao, Ning, Zheng.
Wrote or contributed to the writing of the manuscript: X. Wang, Y. Zhao, Ning, Yu.
Footnotes
- Received February 4, 2020.
- Accepted May 27, 2020.
↵1 X.W. and Y.Z. contributed equally to this work.
This study was supported and funded by the National Key Research and Development Program of China (SQ2017YFC1600201), National Natural Science Foundation of China (91743113, 81973075, and 81903354), and Young Taishan Scholars Program of Shandong Province (tsqn201812046).
Disclaimer: The information in this paper is not an official guidance or policy statement of the US Food and Drug Administration (FDA). No official support or endorsement by the US FDA is intended or should be inferred.
↵
 This article has supplemental material available at molpharm.aspetjournals.org.
This article has supplemental material available at molpharm.aspetjournals.org.

Abbreviations
- ADH
- alcohol dehydrogenase
- ALD
- alcoholic liver disease
- ALDH
- aldehyde dehydrogenase
- -F
- forward
- FREMSA
- fluorescent-based RNA electrophoretic mobility shift assay
- GAPDH
- glyceraldehyde-3-phosphate dehydrogenase
- miRNA
- microRNA
- MRE
- miRNA response element
- Mut
- mutant
- NC
- negative control
- -R
- reverse
- RT-PCR
- real-time polymerase chain reaction
- TCGA
- The Cancer Genome Atlas
- UTR
- untranslated region
- U.S. Government work not protected by U.S. copyright





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}