


Abstract
The anesthetic etomidate modulates synaptic α1β2/3γ2 GABAA receptors via binding sites located in transmembrane β+/α− interfaces. Various approaches indicate that etomidate binds near β2/3M286 side chains, including recent cryogenic electron microscopy images in α1β2γ2L receptors under nonphysiologic conditions with ∼3.5-Å resolution. We hypothesized that substituted cysteine modification and protection experiments using variably sized n-alkyl-methanethiosulfonate (MTS) reagents could precisely estimate the distance between bound etomidate and β3M286 side chains in activated functional receptors. Using voltage-clamp electrophysiology in Xenopus oocytes expressing α1β3M286Cγ2L GABAA receptors, we measured functional changes after exposing GABA-activated receptors to n-alkyl-MTS reagents, from methyl-MTS to n-decyl-MTS. Based on previous studies using a large sulfhydryl reagent, we anticipated that cysteine modifications large enough to overlap etomidate sites would cause persistently increased GABA sensitivity and decreased etomidate modulation and that etomidate would hinder these modifications, reducing effects. Based on altered GABA or etomidate sensitivity, ethyl-MTS and larger n-alkyl-MTS reagents modified GABA-activated α1β3M286Cγ2L GABAA receptors. Receptor modification by n-propyl-MTS or larger reagents caused persistently increased GABA sensitivity and decreased etomidate modulation. Receptor-bound etomidate blocked β3M286C modification by n-propyl-MTS, n-butyl-MTS, and n-hexyl-MTS. In contrast, GABA sensitivity was unaltered by receptor exposure to methyl-MTS or ethyl-MTS, and ethyl-MTS modification uniquely increased etomidate modulation. These results reveal a “cut-on” between ethyl-MTS and n-propyl-MTS, from which we infer that -S-(n-propyl) is the smallest β3M286C appendage that overlaps with etomidate sites. Molecular models of the native methionine and -S-ethyl and -S-(n-propyl) modified cysteines suggest that etomidate is located between 1.7 and 3.0 Å from the β3M286 side chain.
SIGNIFICANCE STATEMENT Precise spatial relationships between drugs and their receptor sites are essential for mechanistic understanding and drug development. This study combined electrophysiology, a cysteine substitution, and n-alkyl-methanethiosulfonate modifiers, creating a precise molecular ruler to estimate the distance between a α1β3γ2L GABA type A receptor residue and etomidate bound in the transmembrane β+/α− interface.
Introduction
Precise structural models of drug interactions with functional receptor states are essential to advancing mechanistic understanding and for guiding drug design. Developing such models has proven particularly challenging for transmembrane drug sites in heteromultimeric transmembrane receptors. Cryogenic electron microscopy (cryo-EM) represents a promising approach (García-Nafría and Tate, 2020), with the caveats that drug-receptor complexes are imaged under nonphysiologic conditions and have uncertain relationships to functional states. High-precision structural studies in functional receptors represent a complementary approach.
GABAA receptors are members of the pentameric ligand-gated ion channel superfamily, the major inhibitory neurotransmitter-gated ion channels in the central nervous system, and key targets of various intravenous sedative-hypnotics, including etomidate, propofol, barbiturates, and alphaxalone (Hemmings et al., 2019). A variety of studies in typical synaptic heteromeric α1β2γ2L and α1β3γ2L GABAA receptors have identified multiple sets of transmembrane intersubunit binding sites that display selectivity for different intravenous hypnotics (Forman and Miller, 2016; Maldifassi et al., 2016). The first such sites to be located bind etomidate, propofol, and their analogs. Mutant function analysis, photolabeling, substituted cysteine modification and protection (SCAMP), and recent cryo-EM images have identified multiple amino acids that are near receptor-bound etomidate, including α1M236, α1L232, and α1T237 in the α1-M1 transmembrane helix; βN265 in the β2/3-M2 helix; and βM286, βF289, and βV290 in the β2/3-M3 helix (Krasowski et al., 1998; Siegwart et al., 2003; Li et al., 2006; Stewart et al., 2008, 2013b; Nourmahnad et al., 2016; Ziemba et al., 2018; Kim et al., 2020). These residues are located in the two outer transmembrane β+/α− intersubunit pockets formed by each pentameric receptor, where subunits are arranged counterclockwise β-α-β-α-γ when viewed from the synaptic space (Baumann et al., 2001).
cryo-EM images of presumably desensitized α1β2γ2L receptors in lipid nanodiscs show varying proximity of etomidate and propofol with the amino acid side chains listed above at ∼3.5-Å resolution (Kim et al., 2020). However, we lack estimates of distances between bound drugs and nearby amino acids in functional open-state receptors. Studies of receptor mutations and analogs of etomidate and propofol indicate that even small steric perturbations can affect binding affinity and/or efficacy (Krasowski et al., 2001; McGrath et al., 2018). However, anesthetic binding site mutations in GABAA receptors produce both local and allosteric effects (Szabo et al., 2019). Most anesthetic photolabels differ chemically from their parent drugs and undergo further chemical alterations and migration when photoactivated. SCAMP, which depends on covalent modification of cysteine sulfhydryl groups, is sensitive to steric interactions between modifying reagents and receptor-bound ligands. However, almost all SCAMP studies of anesthetic sites in GABAA receptors have used the sulfhydryl modifier p-chloromercuribenzene sulfonate (pCMBS), which is a rigid linear molecule over 9 Å long, larger than some anesthetics (Bali and Akabas, 2004; Stewart et al., 2013a,b; Nourmahnad et al., 2016; Stern and Forman, 2016; Ziemba et al., 2018).
We hypothesized that applying SCAMP with a series of n-alkyl-methanethiosulfonate (MTS) probes could provide a more precise approach to determining the distances between receptor-bound anesthetics and nearby amino acid side chains in functional receptors. This approach potentially provides spatial precision approaching 1.3 Å, with increments of one carbon-carbon bond at a dihedral angle of 109.5°. For initial trials, we studied etomidate interactions with α1β3γ2L receptors containing β3M286C mutations. Receptors with β2/3M286C mutations are readily modified by pCMBS, resulting in increased sensitivity to GABA (Bali and Akabas, 2004). In addition, bound etomidate blocks pCMBS modification at β3M286C and α1β2M286Cγ2L receptor interactions with etomidate having been extensively characterized (Stewart et al., 2013a). We postulated that β3M286C side chain modifications by small alkyl-MTS reagents might not overlap with etomidate sites, resulting in different functional effects than modification by larger probes, defining a “cut-on” size. Additionally, previous studies (Jenkins et al., 2001; McGrath et al., 2018) suggest that as β3M286C modifications increase in size, we might observe a cutoff at which large alkyl-MTS reagents are no longer accommodated in etomidate binding pockets.
We used two-microelectrode voltage-clamp electrophysiology to assess persistent functional changes that indicate disulfide bond formation after exposing GABA-activated α1β3M286Cγ2L receptors to n-alkyl-MTS reagents ranging in size from methyl-MTS to n-decyl-MTS. We anticipated that receptors with covalently modified β3M286C side chains overlapping etomidate sites would display persistently increased GABA sensitivity and reduced modulation by etomidate. We also tested whether adding etomidate during exposure to alkyl-MTS reagents blocked development of persistent functional effects associated with disulfide bond formation.
Our results show size-dependent effects of n-alkyl-MTS modification in activated α1β3M286Cγ2L receptors, providing an estimate of the distance between bound etomidate and the native methionine side chain.
Materials and Methods
Animals.
Female Xenopus laevis were housed and maintained in a veterinarian-supervised facility and used with approval from the Massachusetts General Hospital Institutional Animal Care and Use Committee (protocol 2010N000002). Oocytes for use in electrophysiology experiments were harvested via minilaparotomy from frogs anesthetized in 0.2% tricaine, in accordance with the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health.
Materials.
Alkyl-methanethiosulfonate (methyl-MTS, ethyl-MTS, n-propyl-MTS, n-butyl-MTS, n-hexyl-MTS, n-octyl-MTS, and n-decyl-MTS) reagents and pCMBS were purchased from Toronto Research Chemicals. Decyl-MTS was stored as a 500 mM stock in DMSO at −20°C, and other alkyl-MTS reagents were stored as 1 M stocks in DMSO at −20°C. pCMBS was diluted directly into electrophysiology buffer before use. Upon receipt and monthly thereafter, alkyl-MTS reagents and pCMBS were tested for sulfhydryl reactivity, as described by Karlin and Akabas (1998). New reagents were purchased when reactivity dropped more than 20% from the original. During experimental use, solutions of pCMBS and alkyl-MTS reagents in electrophysiology buffer were made fresh at least every 4 hours. R-etomidate (ETO), synthesized by Bachem America (Torrance, CA), was a gift from Prof. Douglas Raines (Department of Anesthesia Critical Care & Pain Medicine, Massachusetts General Hospital, Boston, MA) and was prepared as a 100 mM stock in DMSO and stored at −20°C. GABA and other chemicals were purchased from Sigma-Aldrich.
Molecular Biology.
Coding DNA for human GABAA receptor subunits α1, β3, and γ2L in pCDNA3.1 expression vectors was used. The β3M286C mutation was incorporated as described previously (Stewart et al., 2013a). Capped messenger RNAs were synthesized on linearized DNA templates using mMessage mMachine kits (Sigma-Aldrich) and stored in nuclease-free water at −80°C.
GABAA Receptor Expression in Xenopus Oocytes.
Briefly, oocytes were harvested from frogs and defolliculated with a combination of manual dissection and enzymatic digestion, as previously described (Forman and Stewart, 2012). Prepared oocytes were injected with 1 ng in total of a mixture of capped messenger RNAs encoding GABAA α1, β3 or β3M286C, and γ2L subunits at a 1:1:5 ratio of α:β:γ. After injection and before use in experiments, oocytes were incubated at 18°C for 18–48 hours in ND-96 (in millimolars: 96 NaCl, 2 KCL, 1.8 CaCl2, 1 MgCl2, 5 HEPES, pH 7.4) supplemented with 0.05 mg/ml gentamicin and 0.01 mg/ml ciprofloxacin.
Two-Electrode Voltage-Clamp Electrophysiology.
Experiments were performed at room temperature (21–23°C). Oocytes were placed in a low-volume (0.2 ml) flow chamber and impaled with borosilicate glass microelectrodes filled with 3 M KCl at a resistance of 0.5–2 MΩ. Cells were voltage-clamped at −50 mV (model OC-725C; Warner Instruments). Oocytes were superperfused with antibiotic-free ND-96 solutions delivered at 4 ml/min via computer-controlled valves (BPS-8; ALA Scientific Instruments) and a micromanifold (ALA-VM8; ALA Scientific Instruments). Current signals were digitized at 200 Hz using an RA834 interface and Laboratory Scribe 2 software (both from iWorx, Dover, NH). Digitized data were analyzed off-line using Clampfit8 software (Molecular Devices, San Jose, CA). Current sweeps were filtered with a 10-Hz low-pass Bessel function prior to further analysis and underwent data reduction for display in figures.
Substituted Cysteine Modification.
Covalent modification of the β3M286C residue was assessed using electrophysiology and was based on changes in both GABA sensitivity and etomidate modulation of α1β3M286Cγ2L receptors. Voltage-clamped oocyte currents were first elicited with 10 mM GABA for 15 seconds (sweep 1), followed by a 5-minute wash in ND-96. A second current sweep (sweep 2) was initially activated with 10 μM GABA for 30 seconds and then with 10 μM GABA + 10 μM ETO for 45–60 seconds, followed by another 5-minute ND-96 wash. Alkyl-MTS reagents were applied only to oocytes exhibiting two consecutive low:high GABA response ratios between 0.01 and 0.04 and differing by less than 20%, indicating stable baseline responses. Oocytes were exposed for varying durations to flowing solutions containing varying concentrations of each n-alkyl-MTS reagent combined with 3 mM GABA, followed by a wash in ND-96 for five or more minutes. Alkyl-MTS reagent exposure was calculated as concentration × time (e.g., 300 μM × 30 seconds = 9 mM × s). Postexposure current responses to 10 mM GABA and then 10 μM GABA without and with 10 μM ETO, with intervening ND-96 washes, were again recorded. Negative control studies in α1β3M286Cγ2L receptors used exposure to 3 mM GABA alone (no alkyl-MTS reagent). Another set of controls used wild-type α1β3γ2L receptors exposed to 1 mM GABA + n-alkyl-MTS reagents.
Alkyl-MTS Exposure Effects on GABA Sensitivity.
Low:high GABA response ratios were calculated from the peak current elicited by 10 μM GABA (the first peak in sweep 2) normalized to the peak current elicited with 10 mM GABA (sweep 1). Duplicate pre-exposure and postexposure low:high GABA response ratios from the same oocyte were averaged. For each oocyte, a GABA sensitivity modification ratio was calculated as the postexposure:pre-exposure ratio of averaged low:high GABA response ratios. GABA sensitivity modification ratios were measured in five oocytes for each experimental condition, which was defined by the n-alkyl-MTS reagent and its concentration × time exposure.
GABA sensitivity modification ratios for all seven n-alkyl-MTS reagents were compared under equivalent 9 mM × s exposure conditions (n-decyl-MTS and n-octyl-MTS: 50 μM for 180 seconds; n-hexyl-MTS: 100 μM for 90 seconds; methyl-MTS, ethyl-MTS, n-propyl-MTS, and n-butyl-MTS: 300 μM for 30 seconds). Each set of five oocytes for the seven different n-alkyl-MTS reagents was accompanied by five control oocytes exposed to 3 mM GABA alone, generating a set of 35 control values.
Alkyl-MTS Exposure Effects on Etomidate Enhancement.
Etomidate enhancement ratios were calculated from the two sequential peaks of sweep 2, elicited first with 10 μM GABA alone and then 10 μM GABA + 10 μM ETO, from the same recorded sweep used for low:high GABA response ratios. In individual oocytes, duplicate pre-exposure and postexposure enhancement ratios were measured, and the ETO enhancement modification ratio was calculated as the postexposure:pre-exposure ratio of averaged ETO enhancement ratios. ETO enhancement modification ratios were measured in five oocytes for each modification condition, which was defined by the n-alkyl-MTS reagent and its concentration × time exposure. ETO enhancement modification ratios for all seven n-alkyl-MTS reagents were compared under 9 mM × s exposure conditions (n = 5 per reagent). A set of 35 control values from oocytes exposed to GABA alone were also measured.
Etomidate Protection from Alkyl-MTS Modification.
For n-alkyl-MTS reagents that significantly enhanced GABA sensitivity relative to control conditions (all except methyl-MTS and ethyl-MTS), we tested whether the presence of ETO during n-alkyl-MTS exposures blocked the development of modification effects. After assessing premodification low:high GABA response ratios and 5-minute wash, oocytes (n = 5 per alkyl-MTS reagent) were exposed to 300 μM ETO for 30 seconds and then to 300 μM ETO + 3 mM GABA + n-alkyl-MTS reagents (9 mM × s), as described above. After additional ND-96 washes of 5 minutes each, postmodification low:high GABA response ratios were reassessed in duplicate.
Propyl-MTS Modification Effects on Etomidate-Dependent Enhancement.
We assessed the effects of propyl-MTS modification on the apparent efficacy and potency of etomidate as a positive modulator of α1β3M286Cγ2L receptor activation. ETO enhancement ratios were measured over a range of ETO concentrations (3–300 μM) in a set of five oocytes, both before and after a 30-second exposure to 3 mM GABA + 300 μM n-propyl-MTS (9 mM × s). In individual oocytes, ETO enhancement ratios were measured in duplicate with each ETO concentration.
Sequential Methyl-MTS and pCMBS Exposures.
To test whether methyl-MTS formed stable disulfide bonds at the β3M286C sulfhydryl, we tested whether methyl-MTS exposure could prevent a subsequent exposure to pCMBS from modifying α1β3M286Cγ2L receptors. In each oocyte (n = 5), baseline low:high GABA response and ETO enhancement ratios were measured (T1) and repeated after a 5-minute ND-96 wash (T2). Oocytes were then exposed to 3 mM GABA + 300 µM methyl-MTS for 30 seconds (9 mM × s), followed by a 5-minute wash in ND-96. Low:high GABA response and ETO enhancement ratios were again assessed in duplicate with a 5-minute wash between each sweep (T3, T4). Oocytes were then exposed to 3 mM GABA + 100 µM pCMBS for 30 seconds, followed by a 5-minute ND-96 wash and two more measurements of low:high GABA response and ETO enhancement ratios (T5, T6). For comparison, we made duplicate measurements of GABA response and ETO enhancement ratios in another set of five oocytes before (T1, T2) and after the initial 30-second exposure to 3 mM GABA + 100 µM pCMBS, then 5-minute washes (T3, T4) followed by a 30-second exposure to 3 mM GABA + 300 μM methyl-MTS and wash (T5, T6).
Molecular Modeling.
Molecular models of methionine, cysteine, and modified cysteines were constructed using a Web-based version of Java Script Molecular Editor (http://biomodel.uah.es/en/DIY/JSME/draw.en.htm) (Bienfait and Ertl, 2013). The molecular models were visualized, energy-minimized, and measured using JSMol, an open-source Hypertext Markup Language 5 viewer for chemical structures in three dimensions (http://wiki.jmol.org/index.php/JSmol).
Curve Fitting and Statistical Analysis.
Curve fitting and statistical analyses were performed in Prism 8.4.3 (GraphPad Software). Results are reported and displayed as means ± S.D.
ETO-dependent enhancement data before and after propyl-MTS modification were fit using nonlinear least squares to a logistic dose-response function [Y = 1 + (Max − 1)/(1 + 10^((log EC50 − log X)))], where Y is the enhancement ratio, X is the ETO concentration, Max is maximal enhancement, and EC50 is the half-maximal effect ETO concentration. Fitted parameters for pre- and postexposure data were compared using F tests.
Single-variable ANOVA analysis and Dunnett’s multiple comparisons tests were used to compare GABA sensitivity modification ratios and ETO enhancement modification ratios for each of the seven n-alkyl-MTS reagents at 9 mM × s exposure (n = 5 per reagent) with controls exposed to GABA alone (n = 35).
We inferred that ETO sterically interacted with the n-alkyl-MTS reagent if its presence during exposure significantly reduced the low:high GABA response modification ratio. These and other comparisons between two experimental groups were based on Student’s t tests.
Statistically significant results were inferred if P < 0.05 (α = 0.05).
Results
Persistent Effects on GABA Sensitivity after α1β3M286Cγ2L GABAA Receptor Exposure to n-Alkyl-MTS Reagents.
Our initial experiments tested whether exposures of heterologously expressed α1β3M286Cγ2L receptors to n-alkyl-MTS reagents combined with maximally activating GABA (3 mM) resulted in persistent increases in GABA sensitivity, as previously observed with pCMBS modification (Stewart et al., 2013a). For each n-alkyl-MTS reagent, we tested a range of concentrations, and using voltage-clamp electrophysiology in Xenopus oocytes, we compared the ratio of currents elicited by low GABA (10 μM ≈ EC2-3; concentration eliciting 2-3% of maximal current response) to maximal-activating GABA before and after reagent exposure (e.g., Fig. 1A). Control experiments were performed using α1β3M286Cγ2L receptors exposed to GABA alone (no MTS reagent). Additional controls were performed in oocytes expressing wild-type α1β3γ2L receptors exposed to high GABA combined with alkyl-MTS reagents and tested with appropriately adjusted low (3 μM) and high (1 mM) GABA activation.

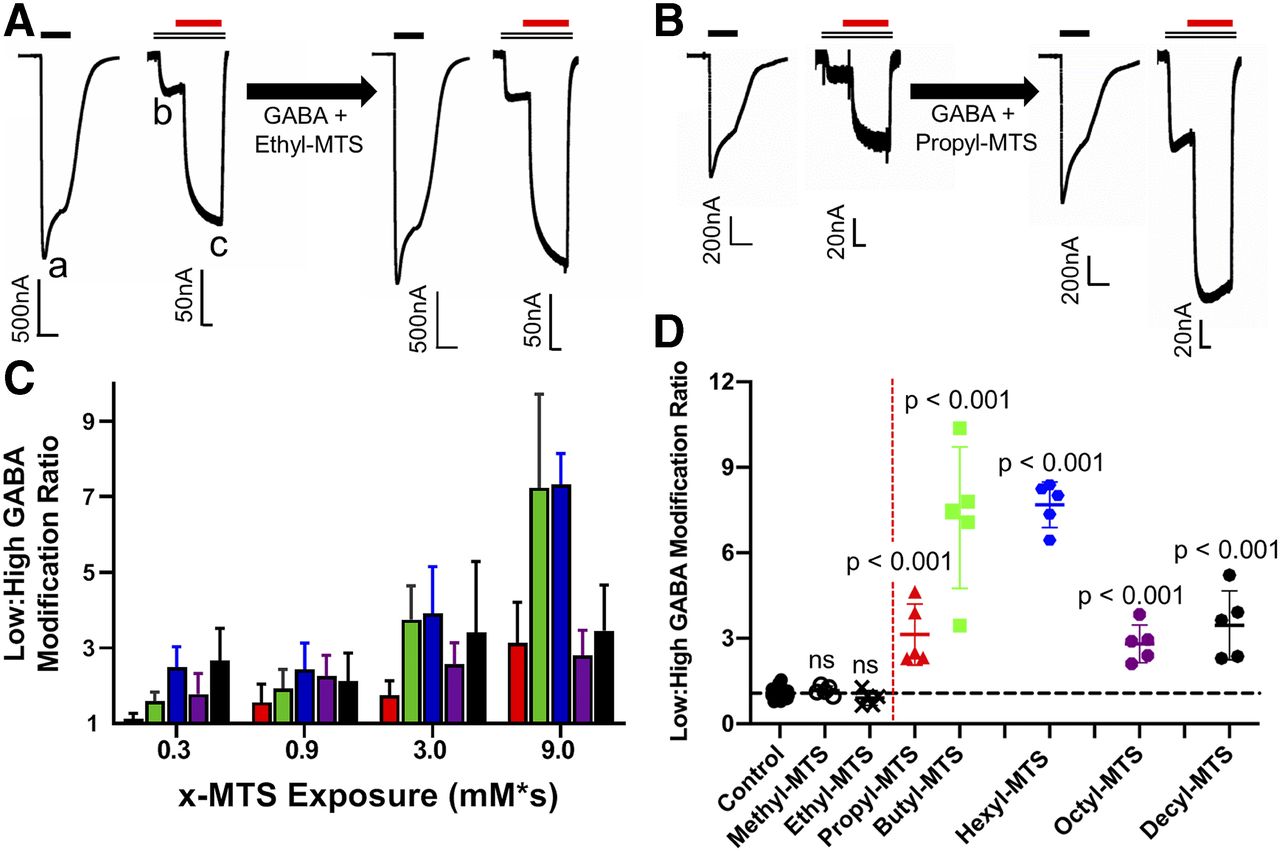
GABA sensitivity of α1β3M286Cγ2L receptors after n-alkyl-MTS exposure. (A) Ethyl-MTS exposure (300 μM × 30 seconds) produces no significant changes in GABA sensitivity. Left traces are paired pre-exposure sweep 1 and sweep 2. Right traces are post-exposure sweeps recorded from the same oocyte (note different scaling of sweep 1 vs. sweep 2 in each pair). Bars above traces represent applications to 10 mM GABA (solid black), 10 μM GABA (double black), and 10 μM etomidate (solid red). Recorded current peaks are labeled a, b, and c. Low:high GABA response ratio = b/a, and etomidate enhancement ratio = c/b. (B) n-Propyl-MTS exposure (300 μM × 30 seconds = 9 mM/s) produces increased GABA sensitivity and decreased etomidate modulation. Paired sweeps (note different scaling of sweep 1 vs. sweep 2) and drug applications are arranged as in (A). (C) n-Alkyl-MTS reagents produce exposure-dependent increases in GABA sensitivity. Bars represent means ± S.D. (n = 5 for each n-alkyl-MTS reagent); red = n-propyl-MTS; green = n-butyl-MTS; blue = n-hexyl-MTS; purple = n-octyl-MTS; black = n-decyl-MTS. (D) GABA sensitivity changes after equivalent (9 mM × s) n-alkyl-MTS exposures. Symbols represent individual oocyte results (open black circles = methyl-MTS; black x's = ethyl-MTS; red triangles = n-propyl-MTS; green squares = n-butyl-MTS; blue hexagons = n-hexyl-MTS; purple octagons = n-octyl-MTS; black circles = n-decyl-MTS) and lines represent means ± S.D. (n = 5 for each n-alkyl-MTS reagent). P values were calculated by ANOVA and Dunnett’s tests against controls (n = 35). The horizontal line is the control mean, and the vertical dashed red line indicates the cut-on of GABA sensitivity effect between ethyl-MTS and n-propyl-MTS.
Control experiments in oocytes expressing α1β3γ2L receptors exposed to GABA plus n-alkyl-MTS reagents or pCMBS (up to 90 mM × s with methyl-MTS and 9 mM × s for others) produced no significant changes in low:high GABA response ratios (data not shown). In oocytes expressing α1β3M286Cγ2L receptors prior to alkyl-MTS reagent exposure, the average low:high GABA response ratio was 0.021 ± 0.0090 (mean ± S.D.; n = 70). In control oocytes expressing α1β3M286Cγ2L receptors and exposed to GABA alone, no significant change in average low:high GABA response ratio was observed (postexposure:pre-exposure ratio = 1.1 ± 0.17; mean ± S.D.; n = 35; P = 0.13 by two-tailed paired Student’s t test).
After α1β3M286Cγ2L receptor exposure to 3 mM GABA + methyl-MTS up to 1 mM for 90 seconds, low:high GABA response ratios remained similar to controls (90 mM × s modification ratio, mean ± S.D. = 1.5 ± 0.35; n = 5; P = 0.20; and 9 mM × s modification ratio = 1.2 ± 0.15; n = 5; P = 0.99; data not shown). Similarly, GABA sensitivity remained close to control after exposures to GABA + ethyl-MTS at 300 μM for 30 seconds or longer (Fig. 1A; 9 mM × s modification ratio = 0.90 ± 0.26; n = 5; P = 0.99).
Exposing α1β3M286Cγ2L receptors to GABA + n-propyl-MTS resulted in increased low:high GABA response ratios that persisted after 15 minutes or more of washout (Fig. 1B). Receptor sensitivity to GABA increased with n-propyl-MTS exposure (concentration × time; Fig. 1C, red bars). After exposure to GABA + 300 μM n-propyl-MTS for 30 seconds (9 mM × s), average low:high GABA response ratios were about 3-fold higher than pre-exposure ratios (Fig. 1D; modification ratio mean ± S.D. = 3.1 ± 1.1; n = 5; P < 0.001). Greater exposure to n-propyl-MTS (1 mM × 30 seconds = 30 mM × s) did not further increase the GABA modification response ratio.
The GABA sensitivity effects of modification by n-alkyl-MTS reagents larger than n-propyl-MTS were also significant and increased with exposure (Fig. 1C). Comparing exposures to GABA + 9 mM × s for all tested n-alkyl-MTS reagents (Fig. 1D), GABA sensitivity modification ratios were highest with both n-butyl-MTS (mean ± S.D. = 7.2 ± 2.5; n = 5; P < 0.001 vs. control) and n-hexyl-MTS (mean ± S.D. = 7.7 ± 0.80; n = 5; P < 0.001 vs. control). These effects are similar to the maximal effect of GABA + pCMBS exposure (Stewart et al., 2013a). The effects of 9 mM × s exposures to n-octyl-MTS (mean ± S.D. = 2.8 ± 0.65; n = 5) and n-decyl-MTS (mean ± S.D. = 3.5 ± 1.2; n = 5), although also significantly higher than controls (P < 0.001 for both), were comparable to that of n-propyl-MTS (Fig. 1D).
Persistent Effects on Etomidate Modulation after α1β3M286Cγ2L GABAA Receptor Exposure to n-Alkyl-MTS Reagents.
We assessed persistent changes in etomidate sensitivity after exposure of α1β3M286Cγ2L receptors to n-alkyl-MTS reagents combined with GABA. For each n-alkyl-MTS reagent, we used voltage-clamp electrophysiology in Xenopus oocytes, comparing the ratio of currents elicited by 10 μM GABA + 10 μM etomidate to that elicited by 10 μM GABA alone, before and after reagent exposure and washout. Control experiments were again performed using exposures to 3 mM GABA without MTS reagents.
In oocytes expressing α1β3M286Cγ2L receptors prior to alkyl-MTS exposure, addition of 10 μM ETO to 10 μM GABA enhanced currents over 4-fold (mean + S.D. = 4.4 ± 1.5; n = 70). After control exposures to 3 mM GABA alone, ETO enhancement ratios varied but, on average, slightly declined relative to control (Fig. 2A; post:pre-exposure ratio mean ± S.D. = 0.87 ± 0.10; n = 35).

Etomidate modulation of α1β3M286Cγ2L receptors after n-alkyl-MTS exposure. (A) Etomidate (10 μM) enhancement modification ratios after equivalent (9 mM/s) n-alkyl-MTS exposures. Symbols represent individual oocyte results (open black circles = methyl-MTS; black x's = ethyl-MTS; red triangles = n-propyl-MTS; green squares = n-butyl-MTS; blue hexagons = n-hexyl-MTS; purple octagons = n-octyl-MTS; black circles = n-decyl-MTS), and lines represent means ± S.D. (n = 5 for each n-alkyl-MTS reagent). P values were calculated by ANOVA and Dunnett’s tests against controls (n = 35). The horizontal line is the control mean, and the vertical dashed red line indicates the cut-on of reduced etomidate modulation effect between ethyl-MTS and n-propyl-MTS. (B) Etomidate-dependent modulation of α1β3M286Cγ2L receptor activation before and after 9 mM × s n-propyl-MTS modification. Symbols represent means ± S.D. (n = 5 each). Lines through data represent fits to logistic functions. Pre-exposure (black circles) EC50 = 38 μM (95% CI [confidence interval] = 22–67 μM), max = 32-fold (95% CI = 27–38), and postexposure (red squares) EC50 = 34 μM (95% CI = 21–57 μM), max = 15-fold (95% CI = 13–17).
After receptor exposure to 3 mM GABA + n-propyl-MTS (9 mM × s) and buffer wash, average normalized (post:pre) ETO enhancement modification ratios fell by more than 30% from baseline and significantly more than controls (Figs. 1B and 2A; mean ± S.D. = 0.63 ± 0.11; n = 5; P < 0.001). Greater exposure to n-propyl-MTS (1 mM × 30 seconds = 30 mM × s) did not further decrease ETO modulation. Comparing etomidate-dependent enhancement of 10 μM GABA-elicited currents before versus after n-propyl-MTS modification (Fig. 2B) revealed that the etomidate EC50 was unchanged (pre- and post-EC50 values are 38 and 34 μM, respectively; P > 0.05 by F test), whereas maximal etomidate modulation was reduced by about 50% (P < 0.001 by F test).
After receptor exposures to 3 mM GABA + methyl-MTS at up to 1 mM for 90 seconds, etomidate enhancement ratios remained similar to the control value (Fig. 2A; 9 mM × s post:pre ratio mean ± S.D. = 0.75 ± 0.077; n = 5; P = 0.11 by ANOVA). Surprisingly, exposures to GABA + ethyl-MTS at 300 μM for 30 seconds resulted on average in persistently increased etomidate modulation relative to control (Fig. 2A; mean ± S.D. = 1.0 ± 0.14; n = 5; P = 0.004).
With n-propyl-MTS and larger reagents, exposures to GABA + 9 mM × s n-alkyl-MTS all persistently reduced etomidate sensitivity, with etomidate enhancement modification ratios mirroring those for low:high GABA response (Fig. 2A): n-butyl-MTS (mean ± S.D. = 0.28 ± 0.077; n = 5; P < 0.001), n-hexyl-MTS (mean ± S.D. = 0.24 ± 0.057; n = 5; P < 0.001), n-octyl-MTS (mean ± S.D. = 0.52 ± 0.096; n = 5; P < 0.001), and n-decyl-MTS (mean ± S.D. = 0.49 ± 0.13; n = 5; P < 0.001). The ETO enhancement modification ratios for n-butyl-MTS and n-hexyl-MTS correspond to ETO enhancement ratios near 1.0, reflecting near absence of etomidate modulation in modified receptors. Greater exposure to n-butyl-MTS (1 mM × 30 second = 30 mM × s) did not further reduce ETO modulation. The effects of n-octyl-MTS and n-decyl-MTS exposure were intermediate between those observed with n-propyl-MTS and those observed with n-butyl-MTS or n-hexyl-MTS.
Etomidate Block of n-Alkyl-MTS Modification Effects.
Because covalent modification requires proximity of reactants to the β3M286C side chain, evidence that etomidate inhibits modification implies that it sterically obstructs n-alkyl-MTS reagent access to the targeted sulfhydryl. We therefore compared the effects of GABA + n-alkyl-MTS reagent exposures with and without the addition of 300 μM etomidate, which occupies a large fraction of sites in α1β3M286Cγ2L receptors (Stewart et al., 2013a). For n-propyl-MTS and larger reagents, we used persistently increased GABA sensitivity (low:high modification ratios) to evaluate covalent modification and inferred steric hindrance if the presence of etomidate significantly reduced the effects of exposure to GABA + n-alkyl-MTS. These experiments were not performed for methyl-MTS or ethyl-MTS, which do not alter GABA sensitivity.
Etomidate protection from covalent modification by n-hexyl-MTS is illustrated in Fig. 3, A and B. The dramatic postexposure versus pre-exposure increase in α1β3M286Cγ2L receptor GABA sensitivity produced by exposure to GABA + n-hexyl-MTS (Fig. 3A) is much reduced by addition of etomidate during exposure to modifier (Fig. 3B). Similar evidence of significant etomidate protection was found with n-propyl-MTS and n-butyl-MTS modification (Fig. 3C). However, etomidate did not significantly reduce the effects of exposures to GABA + n-octyl-MTS or n-decyl-MTS (Fig. 3C).

Etomidate inhibition of n-alkyl-MTS modification at β3M286C. (A) n-Hexyl-MTS exposure (100 μM × 90 seconds) produces large increases in GABA sensitivity in α1β3M286Cγ2L receptors. Left traces are paired pre-exposure sweep 1 and sweep 2 (note scaling of sweep 1 vs. sweep 2; low:high ratio = 0.017), and right traces are postexposure sweeps recorded from the same oocyte (low:high ratio = 0.12). Bars above traces represent applications to 10 mM GABA (solid black), 10 μM GABA (double lines), and 10 μM etomidate (solid red). (B) Addition of etomidate (300 μM) reduces n-hexyl-MTS exposure effects. Pre-exposure low:high ratio = 0.014 vs. postexposure low:high ratio = 0.027. (C) Summary of equivalent n-alkyl-MTS exposure (9 mM × s) effects without (left columns) vs. with ETO (right columns). Symbols represent individual oocyte results (red triangles = propyl-MTS; green squares = butyl-MTS; blue hexagons = hexyl-MTS; purple octagons = octyl-MTS; black circles = decyl-MTS), and columns/lines indicate means ± S.D. (n = 5 for each condition). P values were calculated from unpaired two-tailed Student’s t tests.
For ethyl-MTS, we used 10 μM ETO enhancement ratios (Fig. 2A) to assess persistent effects of modification in the absence versus presence of etomidate. To control for the possibility of incomplete etomidate washout in the protection experiments, we changed the control condition by sequentially exposing oocytes expressing α1β3M286Cγ2L receptors to GABA + ethyl-MTS and then to 300 μM etomidate, with each exposure followed by a 5-minute wash in buffer. Control experiments that included a postmodification etomidate exposure showed an average modification ratio different from that reported above (Fig. 2A). The presence of 300 μM etomidate during GABA + ethyl-MTS exposure significantly reduced the persistent change in etomidate modulation observed after sequential exposures to GABA + ethyl-MTS and then to 300 μM etomidate (control mean ± S.D. = 0.89 ± 0.10 vs. protection 0.72 ± 0.047; P = 0.011 by two-tailed unpaired Student’s t test).
Does Methyl-MTS Modify β3M286C?
Among the alkyl-MTS reagents we studied, methyl-MTS exposures produced no persistent changes in GABA or etomidate sensitivity of α1β3M286Cγ2L receptors. These negative findings imply either that covalent modification produced no functional effect or that covalent modification did not occur. If methyl-MTS failed to modify α1β3M286Cγ2L receptors, then subsequent exposure to GABA + pCMBS should produce a large effect on receptor function, whereas modification without functional effects by methyl-MTS should prevent subsequent pCMBS modification and associated functional effects. We assessed persistent changes in GABA sensitivity (low:high response ratios) for functional evidence of pCMBS modification in these experiments (Fig. 4). As noted above, exposing α1β3M286Cγ2L receptors to 3 mM GABA + methyl-MTS (9 mM × s) produced no evident change in GABA low:high response ratio (Fig. 4A, left and middle sweeps, and Fig. 4B, T1–T4). Subsequent exposure of the same oocytes to GABA + pCMBS (100 μM × 30 seconds) resulted in large, persistent increases in GABA sensitivity (Fig. 4A, right sweeps, and Fig. 4B, T5 and T6), suggesting that methyl-MTS failed to modify β3M286C sulfhydryls that were subsequently modified by pCMBS. In contrast, initial exposure to GABA + pCMBS produced large increases in GABA sensitivity that were not significantly reversed by subsequent exposures to GABA + methyl-MTS (Fig. 4, C and D). These results support the inference that methyl-MTS does not modify the engineered sulfhydryl in α1β3M286Cγ2L receptors.

Sequential exposures to pCMBS indicate that methyl-MTS does not modify α1β3M286Cγ2L receptors. (A) Initial 3 mM GABA + 9 mM × s methyl-MTS exposure produces no significant change in GABA sensitivity, whereas subsequent GABA + 3 mM × s pCMBS exposure produces large GABA and ETO sensitivity changes. Paired sweeps were recorded from the same oocyte (note different scaling of sweep 1 vs. sweep 2). Bars above current traces represent applications to 10 mM GABA (solid black), 10 μM GABA (double lines), and 10 μM etomidate (solid red). (B) Summary of results for initial methyl-MTS exposure followed by pCMBS exposure. Symbols represent low:high GABA response ratios (mean ± S.D.; n = 5). T1/T2, T3/T4, and T5/T6 are sets of replicate tests separated by 5-minute buffer washes. (C) Initial 3 mM GABA + 3 mM × s pCMBS exposure produces large GABA and ETO sensitivity changes, and subsequent GABA + 9 mM × s methyl-MTS exposure produces no significant change. Sweep pairs are from a single oocyte, and drug exposures are indicated as in (A). (D) Summary of results for initial pCMBS exposure followed by methyl-MTS exposure. Symbols represent low:high GABA response ratios (mean ± S.D.; n = 5). T1/T2, T3/T4, and T5/T6 are sets of replicate tests separated by 5-minute buffer washes.
Discussion
Summary of Findings.
We found that exposure to different n-alkyl-MTS reagents produced different functional effects in α1β3M286Cγ2L receptors that fall into distinct size-related groups. In particular, n-propyl-MTS, n-butyl-MTS, and n-hexyl-MTS all produced the predicted effects for modifications that overlap the etomidate site. In contrast, our results with methyl-MTS suggest that it did not modify β3M286C, whereas modification by ethyl-MTS uniquely enhanced etomidate sensitivity relative to the unmodified cysteine. Compared with n-hexyl-MTS, exposures to n-octyl-MTS or n-decyl-MTS produced smaller effects on GABA and etomidate sensitivity that were not significantly blocked by etomidate.
Persistent Functional Effects Indicate Disulfide Bond Formation.
Previous studies (Stewart et al., 2013a) show that GABAA receptors containing β2/3M286C mutations retain sensitivity to positive allosteric modulation by etomidate without direct etomidate activation, features attributable to reduced etomidate efficacy relative to wild-type receptors. Covalent modification by pCMBS at β2/3M286C produces large functional changes and is blocked by etomidate. These features suggest that α1β3M286Cγ2L receptors are a good model for assessing the interactions of various other sulfhydryl modifiers with bound etomidate. No persistent effects were observed in wild-type α1β3γ2L receptors exposed to GABA + n-alkyl-MTS reagents. In contrast, exposure to six of the seven tested n-alkyl-MTS reagents produced persistent effects in α1β3M286Cγ2L receptors that grew with increasing exposure (concentration × time). Additionally, α1β3M286Cγ2L receptors exposed to GABA without MTS reagents displayed only small changes in GABA and etomidate sensitivity. In short, the development of substantial persistent functional changes required the presence of both the β3M286C mutation and an alkyl-MTS reagent other than methyl-MTS. Thus, persistent functional effects were attributable to the formation of stable disulfide bonds.
A Cut-On Effect Is Evident between Ethyl-MTS and n-Propyl-MTS.
Under the same exposure conditions (9 mM × s), ethyl-MTS and n-propyl-MTS produced different effects in two outcomes we assessed. Exposure of α1β3M286Cγ2L receptors to ethyl-MTS induced no significant change in GABA sensitivity but uniquely induced persistently increased etomidate sensitivity relative to unexposed receptors. Exposure to GABA + ethyl-MTS in the presence of etomidate significantly reduced the resulting change in etomidate sensitivity. These findings fulfill only one of the conditions for β3M286C modifications that overlap with etomidate sites. In comparison, n-propyl-MTS exposure resulted in persistently enhanced GABA sensitivity and reduced etomidate sensitivity, whereas receptor-bound etomidate blocked modification. Maximal modification by n-propyl-MTS (9 mM × s) reduced the magnitude of etomidate modulation (i.e., efficacy) without altering apparent etomidate potency (EC50). These observations are all expected consequences of the modified cysteine side chains overlapping etomidate sites.
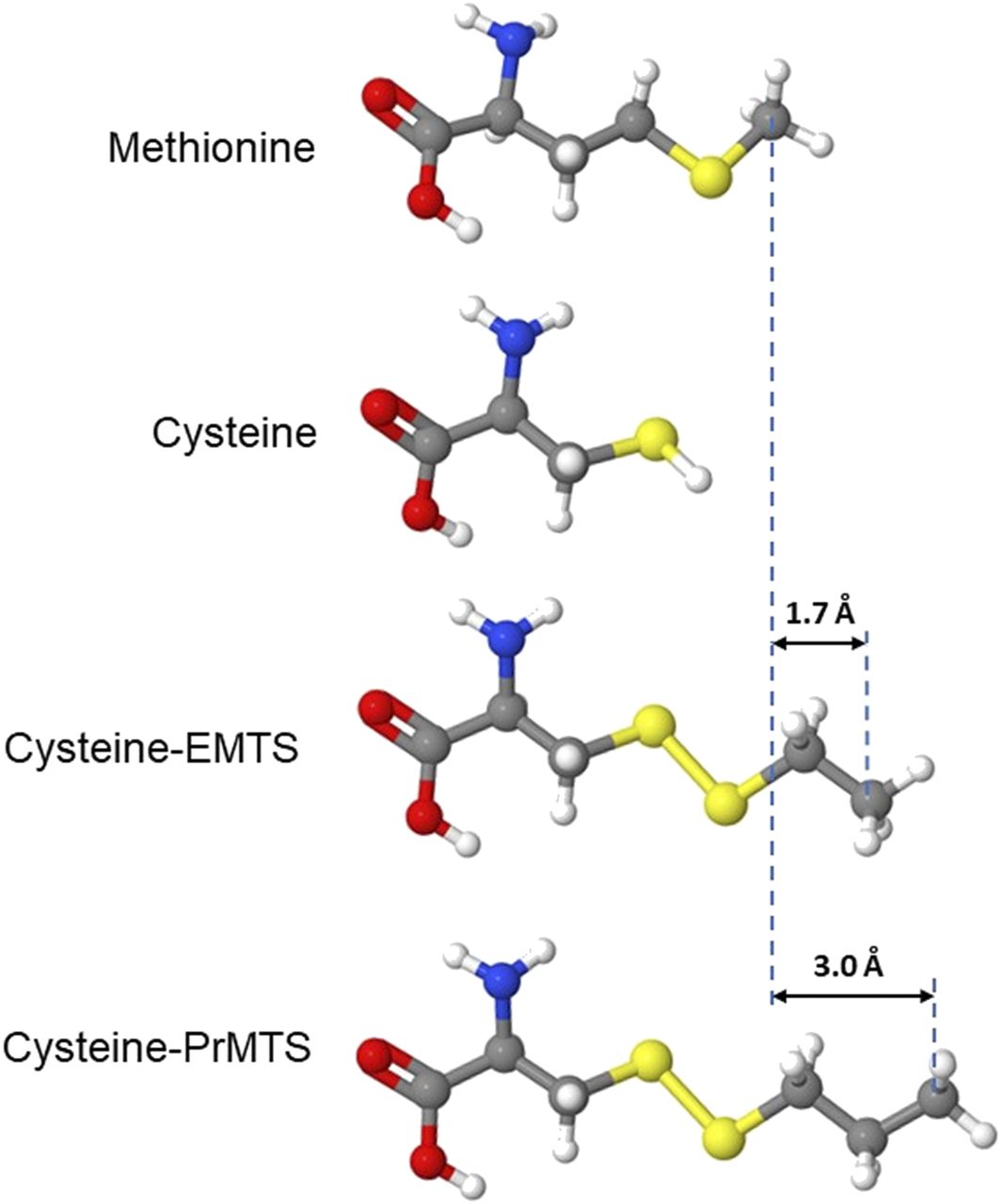
The transition of effects produced by ethyl-MTS versus propyl-MTS modification represents a clear cut-on linked to modifier size with a length difference of ∼1.3 Å. The n-propyl-MTS modification is the smallest cysteine modification for which we can confidently infer overlap with the etomidate sites. Molecular modeling (Fig. 5) indicates that the Cys-S-S-(n-propyl) side chain is 3.0 Å longer than the native methionine, representing an upper limit on the distance between the β3M286 side chain and receptor-bound etomidate. This distance is far closer than that inferred from SCAMP studies with pCMBS. It is in excellent agreement with recent cryo-EM images of etomidate bound to α1β2γ2L receptors, which show van der Waals contacts between the β2M286 side chain and etomidate (Kim et al., 2020). Our current result confirms the utility of this approach as a precise molecular ruler (Johnson et al., 2003) and represents an important confirmation in functional open-state receptors to the cryo-EM images, which were obtained under nonphysiologic conditions with average spatial resolution of 3.5 Å.
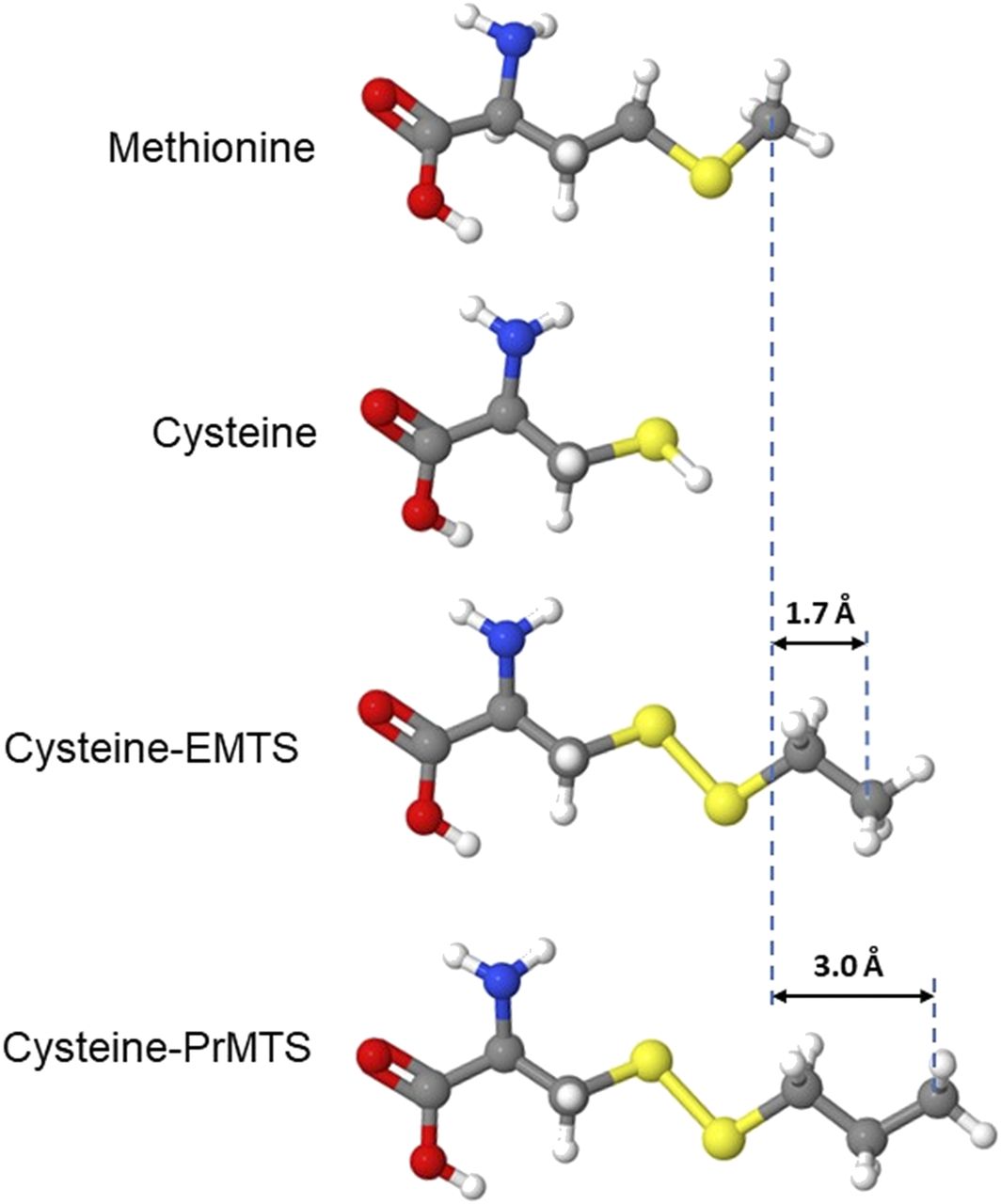
Modeling modified side chains at β3M286C provides estimates of the distance between etomidate and the native methionine in activated receptors. Aligned molecular models of methionine, cysteine, and cysteine side chains modified by ethyl-MTS (EMTS) and n-propyl-MTS (PrMTS) are shown in ball-and-stick format. Differences in molecular lengths between methionine and modified cysteines are also indicated.
Our results with ethyl-MTS modification and etomidate protection are ambivalent regarding overlap with the etomidate sites. The ethyl-modified receptor shows greater sensitivity to etomidate than the unmodified β3M286C mutant, suggesting net attractive van der Waals interactions rather than net steric exclusion. However, mutant receptors modified with ethyl-MTS show less etomidate modulation than wild-type receptors. This may reflect the different chemical properties and/or different sizes of the side chains. We infer from ethyl-MTS results that 1.7 Å is an approximate lower limit for the distance between the native methionine side chain and bound etomidate (Fig. 5). Another caveat is that bound etomidate may shift its position relative to the β3286 side chain in mutants. Additionally, cryo-EM structures for α1β2γ2L receptors differ from α1β3γ2L (Laverty et al., 2019), possibly because of the technical details of their preparation in lipid nanodiscs.
Biphasic Size-Dependent Effects of n-Alkyl-MTS Reagents in α1β3M286Cγ2L Receptors.
Exposure to the smallest alkyl-MTS reagent, methyl-MTS, at concentrations up to 1 mM for 90 seconds, produced no persistent changes in the function of α1β3M286Cγ2L receptors. Exposure to methyl-MTS and wash followed by pCMBS produced persistent functional changes similar to primary exposure to pCMBS (Fig. 4), indicating that methyl-MTS exposure does not produce stable disulfide bonds in α1β3M286Cγ2L receptors.
Exposure of α1β3M286Cγ2L receptors to n-butyl-MTS or n-hexyl-MTS both produced persistent functional effects similar to those caused by n-propyl-MTS but larger in magnitude and comparable to the effects of pCMBS modification. These effects include dramatically increased GABA sensitivity and loss of etomidate modulation. Consistent with our hypothesis that these effects reflect the modified side chains overlapping the etomidate sites, bound etomidate during exposure to these reagents significantly reduces their effects, presumably by blocking access to and reaction with the cysteine sulfhydryl group.
Exposing α1β3M286Cγ2L receptors to GABA + n-octyl-MTS or n-decyl-MTS produced persistently increased GABA sensitivity and reduced etomidate modulation, with effect magnitudes comparable to those of n-propyl-MTS. However, adding etomidate during exposures to these reagents did not significantly reduce their effects. We speculate that, because of their hydrophobicity, both n-octyl-MTS and n-decyl-MTS partition into membrane lipids and intracellular compartments of oocytes and wash out of these compartments more slowly than etomidate. Thus, in etomidate protection studies, these alkyl-MTS reagents may persist in oocyte membranes and modify the β3M286C residue after etomidate washout has occurred.
Both size and hydrophobicity, which covary in the n-alkyl-MTS series, may influence the magnitude of persistent functional changes we observed. Similar inferences emerged from studies of βM286 mutations that reduced propofol modulation (Krasowski et al., 2001). Indeed, incorporating βM286W mutations mimics etomidate binding and obliterates etomidate efficacy (Stewart et al., 2008). Tryptophan is about 1.4 Å longer than the native methionine, but it is more bulky and rigid and has aromatic properties. These observations are consistent with etomidate located within 3 Å of native βM286 but also suggest that side chain properties other than length are important. Additionally, hydrophobic interactions between the amphiphilic β+/α− anesthetic sites and n-alkyl-MTS reagents probably influence the effective concentrations and spatial orientations of these reagents. The smallest reagent that modified the engineered cysteine, ethyl-MTS, produced no change in GABA sensitivity. With larger modifiers up to n-hexyl-MTS, larger effects, including obliteration of etomidate modulation, were observed. However, some of our results also suggest that the efficiency of modification may influence the effect size for modifications that overlap the etomidate sites, especially for methyl-MTS and the largest reagents we tested. Our results suggest that n-propyl, n-butyl-MTS, and n-hexyl-MTS were efficient modifiers, whereas n-octyl-MTS and n-decyl-MTS modified less efficiently. The weak effects of these larger reagents might reflect their limited aqueous solubility and resulting slow modification rate, or a size-dependent steric cutoff in binding to the etomidate sites. Further studies using MTS reagents that produce similarly sized but differentially hydrophobic modifications could determine which of these factors matter.
Precise Localization of Anesthetic Sites in Functional GABAA Receptors.
Our current results demonstrate that a SCAMP strategy using n-alkyl-MTS modifiers can provide precise estimates of distances between receptor-bound anesthetics and nearby amino acid side chains in functional GABAA receptors. Establishing upper-limit distance estimates for several more side chains in the β+/α− transmembrane pockets that form etomidate sites could define an excluded volume that constrains the space occupied by bound etomidate within structural models of activated receptor states. Based on these initial studies, selecting appropriate amino acid targets for this purpose will need to weigh the magnitudes of n-alkyl-MTS modification effects as an important factor in SCAMP sensitivity. Studies in α1β3M286Cγ2L receptors that determine proximity to propofol and other drugs that act through the β+/α− sites are also of interest. Prior SCAMP studies suggest that etomidate, which is larger than propofol, also binds near more amino acid side chains in their shared sites (Nourmahnad et al., 2016). Extension of this approach is also feasible for other transmembrane intersubunit sites where anesthetics bind to GABAA receptors (Chiara et al., 2013; Ziemba et al., 2018) and for other drug-receptor systems.
In conclusion, cryo-EM is providing unprecedented information on ligand poise as well as binding site configuration in GABAA and other transmembrane receptors, but with limited spatial resolution and uncertain relationship to functional states. SCAMP studies such as those we report here represent a complementary high-precision approach to probe spatial relationships between ligands and functional receptors.
Acknowledgments
We thank Youssef Jounaidi for assistance with molecular biology, Douglas Raines for supplying R-etomidate, and both Kieran Bhave and Megan McGrath (all of the Department of Anesthesia Critical Care and Pain Medicine, Massachusetts General Hospital) for assistance with oocyte harvests and frog husbandry.
Authorship Contributions
Participated in research design: Fantasia, Nourmahnad, Forman.
Conducted experiments: Fantasia, Nourmahnad, Halpin.
Performed data analysis: Fantasia, Forman.
Wrote or contributed to the writing of the manuscript: Fantasia, Forman.
Footnotes
- Received December 18, 2020.
- Accepted March 10, 2021.
This work was supported by a grant from National Institutes of Health National Institute of General Medical Sciences to S.A.F. [R01 GM089745].
The authors declare no conflicts of interest.
↵
 This article has supplemental material available at molpharm.aspetjournals.org.
This article has supplemental material available at molpharm.aspetjournals.org.

Abbreviations
- cryo-EM
- cryogenic electron microscopy
- ETO
- R-etomidate
- GABAA
- GABA type A
- MTS
- methanethiosulfonate
- pCMBS
- p-chloromercuribenzene sulfonate
- SCAMP
- substituted cysteine modification and protection
- Copyright © 2021 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}