


Visual Overview
Abstract
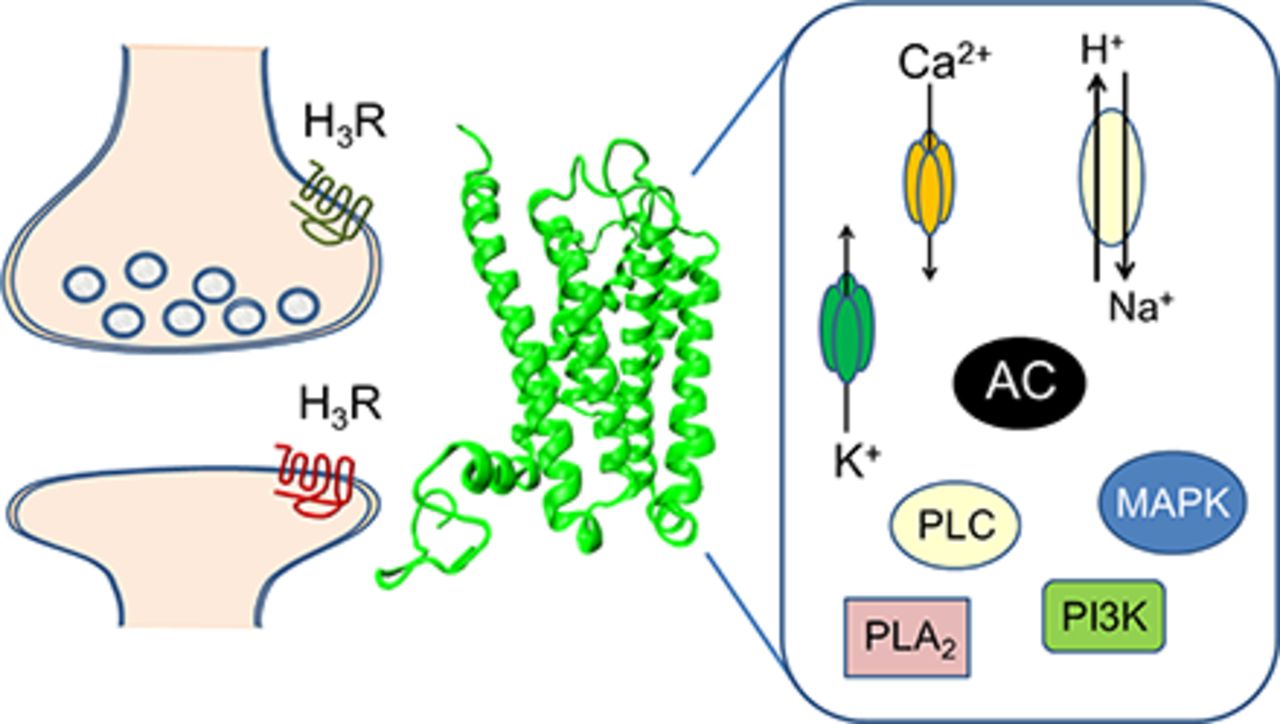
Among the four G protein–coupled receptors (H1–H4) identified as mediators of the biologic effects of histamine, the H3 receptor (H3R) is distinguished for its almost exclusive expression in the nervous system and the large variety of isoforms generated by alternative splicing of the corresponding mRNA. Additionally, it exhibits dual functionality as autoreceptor and heteroreceptor, and this enables H3Rs to modulate the histaminergic and other neurotransmitter systems. The cloning of the H3R cDNA in 1999 by Lovenberg et al. allowed for detailed studies of its molecular aspects. In this work, we review the characteristics of the H3R, namely, its structure, constitutive activity, isoforms, signal transduction pathways, regional differences in expression and localization, selective agonists, antagonists and inverse agonists, dimerization with other neurotransmitter receptors, and the main presynaptic and postsynaptic effects resulting from its activation. The H3R has attracted interest as a potential drug target for the treatment of several important neurologic and psychiatric disorders, such as Alzheimer and Parkinson diseases, Gilles de la Tourette syndrome, and addiction.
Footnotes
- Received April 15, 2016.
- Accepted August 24, 2016.
Research in J.-A. A.-M.’s laboratory is supported by Cinvestav and the Mexican Council for Science and Technology (Conacyt).
- Copyright © 2016 by The American Society for Pharmacology and Experimental Therapeutics
MolPharm articles become freely available 12 months after publication, and remain freely available for 5 years.Non-open access articles that fall outside this five year window are available only to institutional subscribers and current ASPET members, or through the article purchase feature at the bottom of the page.
|





{kind=link}