
Abstract
Aims/hypothesis
In a previous study, we demonstrated that a creatine kinase (CK) modulates KATP channel activity in pancreatic beta cells. To explore phosphotransfer signalling pathways in more detail, we examined whether KATP channel regulation in beta cells is determined by a metabolic interaction between adenylate kinase (AK) and CK.
Methods
Single channel activity was measured with the patch–clamp technique in the inside-out (i/o) and open-cell attached (oca) configuration.
Results
The ATP sensitivity of KATP channels was higher in i/o patches than in permeabilised beta cells (oca). One reason for this observation could be that the local ATP:ADP ratio in the proximity of the channels is determined by factors not active in i/o patches. AMP (0.1 mmol/l) clearly increased open channel probability in the presence of ATP (0.125 mmol/l) in permeabilised cells but not in excised patches. This suggests that AK-catalysed ADP production in the vicinity of the channels is involved in KATP channel regulation. The observation that the stimulatory effect of AMP on KATP channels was prevented by the AK inhibitor P 1,P 5-di(adenosine-5′)pentaphosphate (Ap5A; 20 μmol/l) and abolished in the presence of the non-metabolisable ATP analogue adenosine 5′-(β,γ-imido)triphosphate tetralithium salt (AMP-PNP; 0.12 mmol/l) strengthens this idea. In beta cells from AK1 knockout mice, the effect of AMP was less pronounced, though not completely suppressed. The increase in KATP channel activity induced by AMP in the presence of ATP was outweighed by phosphocreatine (1 mmol/l). We suggest that this is due to an elevation of the ATP concentration by CK.
Conclusions/interpretation
We propose that phosphotransfer events mediated by AK and CK play an important role in determining the effective concentrations of ATP and ADP in the microenvironment of pancreatic beta cell KATP channels. Thus, these enzymes determine the open probability of KATP channels and eventually the actual rate of insulin secretion.
Similar content being viewed by others
Introduction
The KATP channel of pancreatic beta cells plays a predominant role in insulin secretion because it links beta cell metabolism to electrical activity that triggers Ca2+ influx and finally exocytosis. KATP channel activity is regulated by ATP derived from beta cell metabolism. The bulk ATP concentration in beta cells ranges between 3 and 5 mmol/l at high glucose concentration [1–3]. In excised patches half-maximal inhibition of beta cell KATP channels can be achieved in the micromolar range [4, 5]. In intact cells, ATP concentrations required for channel inhibition are somewhat higher but still below the physiological cytosolic ATP concentration [6, 7]. Therefore, ATP-sensitive K+ channels should be completely inhibited under physiological conditions; however, they are evidently active.
To reconcile the experimental observations with the physiological situation, it has been postulated that there must be additional regulatory mechanisms that determine the activity of KATP channels in beta cells. It is well accepted that ADP counteracts the inhibitory action of ATP [8–10]. While ATP exerts its inhibiting effect by binding at the inwardly rectifying potassium channel 6.2 (Kir6.2) subunit of the KATP channel, ADP interacts with the nucleotide-binding domains of the sulfonylurea receptor 1 (SUR1) subunit, thereby promoting channel opening. Therefore, ADP is at least as important as ATP in KATP channel regulation [11, 12]. However, the problem remains that bulk ATP and ADP concentrations would have to undergo large and rapid fuel-dependent changes to explain satisfactorily how the signal from glucose metabolism is transferred to the KATP channels. In particular, beta cell membrane potential oscillations, which include rhythmic changes in the open probability (P o) of KATP channels [13–15], cannot be explained by global concentration changes of ATP in the millimolar range. Thus, it seems much more likely that alterations in the microenvironment of the channels determine P o for the KATP channel. In the past decade, several suggestions have been put forward, with long-chain acyl-coenzyme A esters [16, 17], C16:0 sulfatide [18] and membrane phosphatidylinositol phosphates (especially phosphatidylinositol 4,5-bisphosphate (PIP2)) [19, 20] having been shown to increase the P o of pancreatic KATP channels by reducing their ATP sensitivity. These regulatory mechanisms are more likely to be of significance in receptor-dependent signalling pathways [19–21] or under conditions of pathophysiological stress [16, 17, 22, 23]. Recently, it has been proposed that the ATP sensitivity of pancreatic KATP channels is primarily determined by interaction of Mg-nucleotides with the nucleotide-binding domains of the SUR1 subunit [3].
We have demonstrated previously that a membrane-associated creatine kinase (CK) is involved in KATP channel regulation in native beta cells [24] and proposed that phosphocreatine (PCr) acts as a shuttle, transferring energy-rich phosphate to the plasma membrane where ADP is phosphorylated by a CK in close vicinity to the channels. It is tempting to speculate that several phosphotransfer enzymes together constitute a network that functions as a metabolic barrier. This enzyme network would on the one hand determine the actual ATP and ADP concentrations that the channels sense, and on the other hand shield beta cell KATP channels against bulk ATP and ADP concentrations. One good candidate for such an arrangement is an adenylate kinase (AK), which forms ADP using ATP and AMP as substrates. This enzyme would counterbalance ATP production by the CK. For cardiomyocytes it has been shown that macromolecular complexes consisting of CK, AK and KATP channels exist and that both enzymes are involved in the regulation of KATP channels [25, 26]. In the present paper we investigated whether AK activity could be involved in the regulation of KATP channels of pancreatic beta cells. We demonstrate here that the enzyme increases the P o of the channels and suggest that it is part of a metabolic barrier shielding the KATP channels from bulk ATP and ADP concentrations.
Methods
Islet and cell preparation
Experiments were performed on single pancreatic beta cells of fed male C57Bl/6N mice (20–30 g; Charles River, Sulzfeld, Germany) and AK1 knockout (AK1-KO) mice [27] (Bé Wieringa, Nijmegen, The Netherlands) that had been killed by CO2. Islets were isolated by collagenase digestion of the pancreas. Cells were dispersed in Ca2+-free medium and cultured up to 4 days in RPMI 1640 medium supplemented with 10% fetal calf serum, 100 U/ml penicillin and 100 μg/ml streptomycin [28].
Electrophysiology
Open-cell attached (oca) and inside-out (i/o) measurements were performed using an EPC-9 amplifier and the Pulse software package (both from Heka, Lambrecht, Germany). Data were stored on videotape for later analysis with Igor software (Wavemetrics, Lake Oswego, OR, USA). Channel activity in single channel recordings is given as NP o, the product of P o calculated by point-by-point analysis and the number of active channels in the patch (N). Each experiment was started in the cell-attached configuration with a bath solution containing (mmol/l): 140 NaCl, 5 KCl, 1.2 MgCl2, 2.5 CaCl2, 0.5 glucose, 10 HEPES, pH 7.4 adjusted with NaOH (solution 1). For oca and i/o measurements the bath solution was changed to (mmol/l): 130 KCl, 5 EGTA, 0.5 glucose, 10 HEPES, 1.5 MgCl2, pH 7.2 adjusted with KOH (solution 2). When ATP was added, this solution was adjusted to 3 mmol/l MgCl2. The pipette solution consisted of (mmol/l): 130 KCl, 4 MgCl2, 2 CaCl2, 10 EGTA, 0.65 Na2ATP, 20 HEPES, pH 7.15 adjusted with KOH.
The oca configuration was obtained as follows. After formation of a cell-attached patch, the bath solution was changed to solution 2 and the holding potential was set to −50 mV. A second pipette containing solution 2 with 0.05% saponin (w/v) was then brought to the other side of the cell. Positive pressure was applied to eject saponin-containing solution on to the surface of the cell. Permeabilisation was assessed by the development of a slight swelling of the cell and the increase in NP o of KATP channels due to wash-out of cellular ATP [29, 30]. The saponin-containing pipette was then withdrawn. Single KATP channel currents were recorded at 25° to 28°C.
Pipettes were pulled with a DMZ Puller (Zeitz Instruments, Munich, Germany) using borosilicate glass (Clark, Pangbourne, UK). They had resistances of 7–12 MΩ when filled with pipette solution.
Chemicals
HEPES was obtained from Roth (Karlsruhe, Germany), collagenase from Serva (Heidelberg, Germany) or Roche Diagnostics (Mannheim, Germany) and adenosine 5′-(β,γ-imido)triphosphate tetralithium salt (AMP-PNP) from Fluka (Seelze, Germany). RPMI 1640, fetal calf serum and penicillin–streptomycin were provided by Invitrogen (Karlsruhe, Germany). All other chemicals were purchased from Sigma-Aldrich (Seelze, Germany) and Merck (Darmstadt, Germany) in the purest form available.
Presentation of results
Electrophysiological experiments are illustrated by recordings that are representative for the indicated number of experiments performed with different cells. Beta cells of at least two different cell preparations were used for each series of experiments. Where possible, values in the text are given as means ± SEM for the indicated number of experiments. The statistical significance of differences between means was assessed by a Student’s t test for paired values when two samples were compared; multiple comparisons were made by ANOVA followed by Student–Newman–Keuls test. A value of p ≤ 0.05 was considered to represent a statistically significant difference.
ATP concentration–response curves were fitted with the Hill equation: \( y = {100} \mathord{\left/ {\vphantom {{100} {{\left[ {1 + {\left( {x \mathord{\left/ {\vphantom {x {{\text{IC}}}}} \right. \kern-\nulldelimiterspace} {{\text{IC}}}_{{50}} } \right)}^{h} } \right]}}}} \right. \kern-\nulldelimiterspace} {{\left[ {1 + {\left( {x \mathord{\left/ {\vphantom {x {{\text{IC}}}}} \right. \kern-\nulldelimiterspace} {{\text{IC}}}_{{50}} } \right)}^{h} } \right]}} \), where y equals NP o of the KATP channel in the test solution (taken as percentage of NP o in the solution without ATP), x represents the ATP concentration used and IC50 is the ATP concentration that causes half-maximal block, h is the Hill coefficient. IC50 is given ±SD.
Results
Different ATP sensitivity of KATP channels in permeabilised beta cells vs cell-free patches
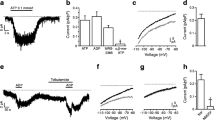
In Fig. 1, KATP channel inhibition in dependence of the ATP concentration is compared in beta cells permeabilised by saponin (=oca) with patches in the i/o configuration. The pores formed by permeabilisation in the oca configuration allowed us to establish a range of intracellular nucleotide concentrations by extracellular application of ATP, AMP etc. [29, 30]. ATP concentrations of 3, 10, 30, 100 and 300 μmol/l were applied. NP o under control conditions (without ATP) was taken as 100%. In each experiment the inhibitory effectiveness of a certain ATP concentration was determined first in the oca configuration and afterwards in the i/o configuration. The data reveal a clear right-shift of the curve obtained in the oca configuration vs that measured in the i/o configuration. The values for half-maximal inhibition amounted to 6.6 ± 0.7 μmol/l (i/o) and 27.5 ± 14.8 μmol/l (oca) (n = 4–8 for different ATP concentrations).

KATP channel sensitivity to ATP inhibition is different in permeabilised cells and excised patches. The figure shows the relation between the ATP concentration and NP o measured in the oca configuration (open circles) and in i/o patches (closed circles). Control values before addition of ATP were taken as 100%. n = 4–8 for different ATP concentrations. *p < 0.05, **p < 0.01
Regulation of the KATP channel by an AK
To test whether an AK is involved in the regulation of KATP channels of pancreatic beta cells, we added AMP to cells in the presence of ATP after formation of the oca configuration. AK-mediated production of ADP should increase NP o of KATP channels. Figure 2a shows that 0.1 mmol/l AMP added in the presence of 0.125 mmol/l ATP clearly increased channel activity. To quantify the effect, NP o under control conditions (ATP 0.125 mmol/l) in each experiment was taken as 1 and changes were referred to this value. AMP increased normalised NP o from 1 to 5.95 ± 1.07 (n = 5, p < 0.001 vs control). Removal of AMP reversed the effect. Normalised NP o amounted to 0.92 ± 0.41 (n = 4, p < 0.01 vs value with AMP) after wash-out of AMP.

KATP channels are activated by AMP in (a) oca patches but not in (b) i/o patches. a Representative experiment showing that NP o of beta cell KATP channels markedly increased when 0.1 mmol/l AMP was applied in the presence of 0.125 mmol/l ATP. The number of open channels is indicated by the arrowheads on the left-hand side. The lower traces show parts of the experiment before and during application of AMP and after wash-out at extended time scales. The box above indicates the AK-catalysed reaction responsible for the channel opening. b Representative experiment showing that in cell-free patches AMP (0.1 mmol/l) did not increase KATP channel activity in the presence of ATP (0.1 mmol/l). The number of open channels is indicated by the arrowheads on the left-hand side
Lack of AK-mediated KATP channel regulation in excised patches
To evaluate whether the discrepancy in ATP sensitivity of the KATP channel between i/o and oca was linked to AK activity, we repeated the experiments shown in Fig. 2a in i/o experiments. Addition of 0.1 mmol/l AMP in the presence of 0.1 mmol/l ATP did not alter channel activity under these conditions (Fig. 2b). Moreover, these data rule out the possibility that the stimulatory effect of AMP on pancreatic KATP channels in the oca configuration is a direct effect of addition of this metabolite. The slight reduction of NP o in the presence of AMP (to 0.72 ± 0.26; n = 5, NS vs control) is likely to be channel rundown because it persisted after wash-out (0.34 ± 0.06, n = 4, p < 0.05 vs control). In control experiments under the same experimental conditions as above, 0.2 mmol/l ADP was still able to open KATP channels in the presence of ATP 0.1 mmol/l (67.68 ± 33.03 vs 1 under ATP alone, n = 5) (data not shown). Thus, ADP remains capable of increasing the channel activity under conditions where AMP fails to do so. This indicates that the failing effect of AMP in i/o experiments is due to the lack of AK activity.
Inhibition of the AK by P 1,P 5-di(adenosine-5′)pentaphosphate
If an AK is responsible for the increase of NP o in oca experiments after addition of AMP (0.1 mmol/l), the effect should be prevented by the AK inhibitor P 1,P 5-di(adenosine-5′)pentaphosphate (Ap5A) [31, 32]. Indeed, Fig. 3 shows that addition of 20 μmol/l Ap5A completely suppressed channel activation by AMP in the oca configuration. Channel NP o, taken as 1 under control conditions (ATP 0.125 mmol/l + Ap5A 20 μmol/l), was even decreased with AMP to 0.54 ± 0.17 (n = 6, p < 0.01 vs control), falling subsequently to 0.31 ± 0.11 after wash-out (n = 5, p < 0.01 vs control). The further decline after wash-out is probably due to channel rundown. Diadenosine polyphosphates have been found to directly inhibit KATP channel activity in cardiac cells and pancreatic beta cells [33]. Therefore, in experiments with Ap5A, AMP was added when a constant channel activity was reached in the presence of ATP and Ap5A. Furthermore, we ruled out the possibility that Ap5A prevents channel activation by ADP. To do this, we applied ADP (0.2 or 0.5 mmol/l) in oca patches in the presence of 0.125 mmol/l ATP and 20 μmol/l Ap5A (Fig. 4). Again, channel NP o in the presence of ATP was taken as 1. In the presence of Ap5A, it amounted to 1.80 ± 0.37 in the series with 0.2 mmol/l ADP and 0.93 ± 0.24 in the series with 0.5 mmol/l ADP, increasing to 25.39 ± 13.10 and 20.83 ± 7.63, respectively after addition of ADP (n = 4 and n = 5). This shows that ADP is able to activate KATP channels in the presence of the inhibitor.

The effect of AMP can be blocked by the AK inhibitor Ap5A. In an oca experiment AMP (0.1 mmol/l) was not able to increase KATP channel activity when the AK was inhibited by 20 μmol/l Ap5A (with 0.125 mmol/l ATP). The number of open channels is indicated by the arrowheads on the left-hand side

Ap5A does not prevent channel activation by ADP. Ap5A (20 μmol/) in the presence of 0.125 mmol/l ATP does not prevent 0.5 mmol/l ADP from augmenting the channel activity in oca experiments. The number of open channels is indicated by the arrowheads on the left-hand side
KATP channels of AK1-KO murine beta cells still exhibit regulation by AMP
AK1-KO mice (first described by Janssen et al. [27]) lack the cytosolic AK1 isoform, which could be responsible for the AK activity that regulates the beta cell KATP channel as shown above. Therefore, we performed oca experiments as in Fig. 2 to explore whether the regulation by AMP is abolished in AK1-KO mice. Contrary to our expectation, this was not the case and AMP still increased NP o in the AK1-KO cells (Fig. 5). However, the increase was smaller than in wild-type beta cells (AMP increased the normalised NP o from 1 to 2.65 ± 0.33 in AK1-KO cells, n = 5, p < 0.001 vs control, p < 0.05 vs AMP in wild-type beta cells). These data suggest that AK1 is not the only AK isoform responsible for KATP channel regulation in pancreatic beta cells.

KATP channels in AK1-KO mice are still activated by AMP in oca experiments. In the presence of 0.125 mmol/l ATP, 0.1 mmol/l AMP is still able to activate KATP channels in AK1-KO mice, but to a lesser extent than in wild-type beta cells. The number of open channels is indicated by the arrowheads on the left-hand side
AMP does not activate KATP channels in the presence of a non-metabolisable ATP analogue (AMP-PNP)
If activity of an AK is responsible for the opening of the KATP channels, as suggested from Fig. 2, the reaction would not be expected to take place in the presence of a non-metabolisable ATP analogue. According to this expectation, addition of 0.1 mmol/l AMP did not increase NP o when the non-metabolisable ATP analogue AMP-PNP (0.12 mmol/l) was used instead of ATP in oca experiments (Fig. 6). Channel NP o was taken as 1 in the presence of AMP-PNP and decreased to 0.41 ± 0.14 with AMP (n = 4, p < 0.01 vs control; wash-out 0.36 ± 0.10, n = 4, p < 0.01 vs control). In three of these experiments, AMP (0.1 mmol/l) was subsequently tested for an effect in the presence of ATP (0.05–0.125 mmol/l) and indeed elicited a five- to eightfold increase in channel activity (data not shown). This strengthens the argument that the presence of a metabolisable nucleotide is needed as the substrate for AK-mediated phosphotransfer to produce the channel openings.

Lack of activation of KATP channels by AMP in the presence of AMP-PNP. AMP (0.1 mmol/l) fails to increase the NP o of KATP channels in oca experiments where a non-metabolisable ATP analogue (AMP-PNP, 0.12 mmol/l) is used instead of ATP. The number of open channels is indicated by the arrowheads on the left-hand side
Effect of PCr
Assuming an interaction between AK and CK in pancreatic beta cells, one would expect PCr to reduce the AMP-induced channel activity because CK forms ATP by transferring phosphate from PCr to ADP. We observed that PCr (1 mmol/l) completely reversed the increase in NP o that was evoked by the activity of the AK (Fig. 7). As in the experiments presented in Fig. 2, normalised NP o was increased about sixfold after addition of AMP (6.31 ± 1.16, n = 3, p < 0.01 vs control). Subsequent application of PCr decreased the NP o to levels equal to those in the presence of ATP alone (1.02 ± 0.21, n = 3, p < 0.01 vs value with AMP). The effect was reversible upon removal of PCr.

PCr reverses the activation of KATP channels by AMP in oca patches. PCr (1 mmol/l) counteracts the effect of AMP (0.1 mmol/l). The boxes above show the reactions catalysed by AK and CK. The number of open channels is indicated by the arrowheads on the left-hand side
Discussion
For native beta cells it has been widely accepted that glucose enhances the intracellular ATP concentration and the ATP:ADP ratio. However, this concept does not explain how glucose metabolism and global intracellular ATP and ADP concentrations are interlinked with KATP channel activity regulation. Experimental data regarding this regulatory network are inconsistent [34–38]; moreover, the high intracellular ATP concentration as such questions the simple assumption that bulk ATP and ADP concentrations regulate beta cell KATP channel activity [3].
One important finding of our work presented here is the right-shift of the concentration–response curve for KATP channel inhibition by ATP in the oca configuration vs excised patches. This supports the idea that the P o is regulated by the ATP and ADP concentration in the microenvironment of the channels rather than by the total cellular ATP and ADP concentration. Interestingly, a similar result has been reported for cardiomyocytes [30] and very recently for pancreatic beta cells [3]. The different ATP sensitivity of KATP channels in oca and excised patches of heart cells was explained by augmented ATP consumption by ATPases and the presence of glycolytic enzymes associated with KATP channels in the oca configuration [30]. In beta cells the difference in the ATP effectiveness between the diverse patch configurations was explained by altered Mg-nucleotide stimulation in oca vs i/o patches at the nucleotide-binding domains of SUR1 [3]. In addition to the above, our data strongly suggest that high-energy phosphoryl exchange between adenosine phosphonucleotides is involved in the regulation of the KATP channel activity in pancreatic beta cells.
Theoretically, nucleotides could also affect beta cell function by stimulating purinergic receptors or changing the activity of the AMP-activated protein kinase (AMPK). Activation of purinergic receptors by binding nucleotides at the extracellular site is reported to increase insulin secretion [39]. However, potentiation of insulin secretion is not reconcilable with the AMP-induced opening of KATP channels and thus we rule out the possibility that the observed effects were due to interaction of the nucleotides with purinergic receptors. AMP also activates the general fuel-sensor AMPK, which has been shown to inhibit glucose oxidation and ATP synthesis in beta cells [40]. However, our results with a non-metabolisable ATP analogue, where AMP failed to increase KATP channel activity in oca measurements (Fig. 6), argue against the possibility that AMPK is involved in the regulation observed in Fig. 2.
AKs reversibly catalyse phosphotransfer between AMP, ADP and ATP. Several isoforms have been identified: these are localised in mitochondria and cytosol or are plasma membrane-bound (for review see [41]) and contribute to the regulation of adenine nucleotide concentration(s) in microdomains of the cell [41].
According to Smink and co-workers [42], different isoforms of the mRNA for AK are found in mouse pancreatic islets. Moderate expression of mRNA for AK1, AK5 and AK7 as well as an enriched expression of mRNA for AK2 and AK3 have been observed so far. For human primary beta cells a moderate expression of mRNA for AK1 has been described whereas the expression of mRNA for AK2 is enriched and that of AK3L1 is only low. No data are available yet for the expression of other isoforms.
Our assumption that AK activity determines local ATP concentration in pancreatic beta cells is based on the following observations: (1) that NP o of KATP channels is increased by AMP in the presence of ATP; (2) that the effect is only observed in permeabilised cells with intact enzyme activity, but not in small excised patches (see below), where ADP is still able to open the channels; (3) that the effect is suppressed by the AK inhibitor Ap5A; and (4) that AMP only activates KATP channels in the presence of ATP, but not in the presence of the non-metabolisable analogue AMP-PNP. Moreover, we demonstrated that PCr counteracted AK-induced KATP channel activation, suggesting the involvement of CK activity. This confirms our previous observation of the existence of CK-mediated phosphotransfer in pancreatic beta cells [24].
The lack of effect of AMP in i/o patches points to a role for cytosolic and/or mitochondria-associated AK isoforms. However, Larsson et al. [43] observed an increase in channel activity with AMP in cell-free macro-patches with extremely low pipette resistances (2–3 vs 8–11 MΩ in our experiments). Macro patches may facilitate the accumulation of nucleotides generated by AK activity or could result in the excision of cytosolic components. According to this concept, we also observed an increase of channel openings by AMP in i/o patches using pipettes with resistances lower than 4 MΩ (data not shown).
Compared with beta cells, the contribution of phosphotransfer mediated by AK and other enzymes to the regulation of KATP channels in heart cells is much better understood [44, 45]. Carrasco and co-workers [26] showed in cardiomyocytes that AMP activates KATP channels in the presence of ATP in the oca configuration. This effect was reversed by PCr and abolished in cells from AK1-KO mice. Thus, in heart cells the AK1 isoform seems to be involved in KATP channel regulation together with a cytosolic CK isoform. Carrasco et al. [26] have further demonstrated that the AK1 is physically associated with a KATP channels in heart cells, an observation described before for a CK [25].
In beta cells from AK1-KO mice, by contrast, an AK-dependent regulation of the KATP channel is preserved. However, this regulation is less pronounced than in wild-type beta cells. Possibly, therefore, another AK activity than AK1 must be involved in the regulation of beta cell KATP channels. Alternatively, AK1 is the only relevant KATP channel-regulating isoform in wild-type beta cells and other AK isoforms can take over this function in AK1-KO cells to compensate for its loss.
Olson and co-workers [46] have shown that increasing the glucose concentration from substimulatory to stimulatory values inhibits the AK-catalysed phosphorylation of AMP by ATP in HIT-T15 cells. This would result in reduced formation of ADP and thus enhanced closure of KATP channels by ATP, a situation that actually occurs after glucose stimulation in beta cells. Very importantly, the authors were able to demonstrate that the glucose-induced decrease in AK-catalysed phosphotransfer occurs prior to or at the time of KATP channel closure, and not as a consequence of events provoked by KATP channel inhibition, including membrane depolarisation, Ca2+ influx and insulin secretion. The authors reported [46] that glucose-induced changes of total intracellular ATP and ADP are not detectable or are marginal in HIT-T15 cells. However, overall ATP metabolic flux is increased due to the increased energy demand of glucose-stimulated secretion, resulting in augmentation of ATP hydrolytic consumption and ATP regeneration. Taken together, these data [46] strongly support the view that not bulk ATP and ADP, but the concentration of the nucleotides in the microenvironments of KATP channels determine the P o of the channels.
Interestingly, Ap n As were reported to be synthesised in beta cells and to increase with rising glucose concentrations [47]. Thus, beta cell endogenous AK inhibitors may form a link between rising glucose concentrations and inhibition of AK.
For pancreatic beta cells we propose a model (Fig. 8) in which AK- and CK-mediated exchange of high-energy phosphoryl groups functions as a metabolic barrier shielding beta cell KATP channels against total cellular ATP and ADP concentrations. According to this model, ATP produced in the glycolytic–mitochondrial pathway is transferred to the KATP channels via a PCr shuttle system and re-equilibration by coupled AK helps to determine the local ATP and ADP concentrations that govern channel behaviour. Future work should help to reveal the role of partitioning of phosphotransfer enzymes and mitochondrial distribution in cell cortical areas underneath sites of KATP in the cell membrane.

Model illustrating the involvement of AK and CK activities in phosphotransfer networking and the regulation of beta cell KATP channels. Cr creatine, mitoCK mitochondrial CK
Abbreviations
- AK1-KO:
-
adenylate kinase 1 knockout
- AK:
-
adenylate kinase
- AMPK:
-
AMP-activated protein kinase
- AMP-PNP:
-
adenosine 5′-(β,γ-imido)triphosphatetetralithium salt
- Ap5A:
-
P 1,P 5-di(adenosine-5′)pentaphosphate
- CK:
-
creatine kinase
- i/o:
-
inside-out
- P o :
-
open probability
- oca:
-
open-cell attached
- PCr:
-
phosphocreatine
- SUR1:
-
sulfonylurea receptor 1
References
Ashcroft FM (1998) Exciting times for PIP2. Science 282:1059–1060
Detimary P, Dejonghe S, Ling Z, Pipeleers D, Schuit F, Henquin JC (1998) The changes in adenine nucleotides measured in glucose-stimulated rodent islets occur in beta cells but not in alpha cells and are also observed in human islets. J Biol Chem 273:33905–33908
Tarasov AI, Girard CAJ, Ashcroft FM (2006) ATP sensitivity of the ATP-sensitive K+ channel in intact and permeabilized pancreatic beta cells. Diabetes 55:2446–2454
Sturgess NC, Ashford ML, Carrington CA, Hales C (1986) Single channel recordings of potassium currents in an insulin-secreting cell line. J Endocrinol 109:201–207
Cook DL, Hales CN (1984) Intracellular ATP directly blocks K+ channels in pancreatic B-cells. Nature 311:271–273
Niki I, Ashcroft FM, Ashcroft SJH (1989) The dependence on intracellular ATP concentration of ATP-sensitive K-channels and of Na,K-ATPase in intact HIT-T15 beta cells. FEBS Lett 257:361–364
Speier S, Yang SB, Sroka K, Rose T, Rupnik M (2005) KATP-channels in beta-cells in tissue slices are directly modulated by millimolar ATP. Mol Cell Endocrinol 230:51–58
Kakei M, Kelly RP, Ashcroft SJH, Ashcroft FM (1986) The ATP sensitivity of K+ channels in rat pancreatic beta cells is modulated by ADP. FEBS Lett 208:63–66
Dunne MJ, West-Jordan JA, Abraham RJ, Edwards RHT, Petersen OH (1988) The gating of nucleotide-sensitive K+ channels in insulin-secreting cells can be modulated by changes in the ratio ATP4-/ADP3- and by nonhydrolyzable derivatives of both ATP and ADP. J Membr Biol 104:165–177
Nichols CG, Shyng SL, Nestorowicz A et al (1996) Adenosine diphosphate as an intracellular regulator of insulin secretion. Science 272:1785–1787
Tucker SJ, Gribble FM, Zhao C, Trapp S, Ashcroft FM (1997) Truncation of Kir6.2 produces ATP-sensitive K+ channels in the absence of the sulphonylurea receptor. Nature 387:179–183
Aguilar-Bryan L, Bryan J (1999) Molecular biology of adenosine triphosphate-sensitive potassium channels. Endocr Rev 20:101–135
Larsson O, Kindmark H, Bränström R, Fredholm B, Berggren PO (1996) Oscillations in KATP channel activity promote oscillations in cytoplasmic free Ca2+ concentration in the pancreatic beta cell. Proc Natl Acad Sci USA 93:5161–5165
Rolland JF, Henquin JC, Gilon P (2002) Feedback control of the ATP-sensitive K+ current by cytosolic Ca2+ contributes to oscillations of the membrane potential in pancreatic beta-cells. Diabetes 51:376–384
Kanno T, Rorsman P, Göpel SO (2002) Glucose-dependent regulation of rhythmic action potential firing in pancreatic beta-cells by KATP-channel modulation. J Physiol 545:501–507
Bränström R, Leibiger IB, Leibiger B, Corkey BE, Berggren PO, Larsson O (1998) Long chain coenzyme A esters activate the pore-forming subunit (Kir6.2) of the ATP-regulated potassium channel. J Biol Chem 273:31395–31400
Gribble FM, Proks P, Corkey BE, Ashcroft FM (1998) Mechanism of cloned ATP-sensitive potassium channel activation by oleoyl-CoA. J Biol Chem 273:26383–26387
Buschard K, Blomqvist M, Mansson JE, Fredman P, Juhl K, Gromada J (2006) C16:0 sulfatide inhibits insulin secretion in rat beta cells by reducing the sensitivity of KATP channels to ATP inhibition. Diabetes 55:2826–2834
Baukrowitz T, Schulte U, Oliver D et al (1998) PIP2 and PIP as determinants for ATP inhibition of KATP channels. Science 282:1141–1144
Shyng SL, Nichols CG (1998) Membrane phospholipid control of nucleotide sensitivity of KATP channels. Science 282:1138–1141
Lin CW, Yan F, Shimamura S, Barg S, Shyng SL (2005) Membrane phosphoinositides control insulin secretion through their effects on ATP-sensitive K+ channel activity. Diabetes 54:2852–2858
Riedel MJ, Boora P, Steckley D, de Vries G, Light PE (2003) Kir6.2 polymorphisms sensitize beta-cell ATP-sensitive potassium channels to activation by acyl CoAs: a possible cellular mechanism for increased susceptibility to type 2 diabetes? Diabetes 52:2630–2635
Lin YW, MacMullen C, Ganguly A, Stanley CA, Shyng SL (2006) A novel KCNJ11 mutation associated with congenital hyperinsulinism reduces the intrinsic open probability of beta-cell ATP-sensitive potassium channels. J Biol Chem 281:3006–3012
Krippeit-Drews P, Bäcker M, Düfer M, Drews G (2003) Phosphocreatine as a determinant of KATP channel activity in pancreatic beta cells. Pflügers Arch 445:556–562
Crawford RM, Ranki HJ, Botting CH, Budas GR, Jovanovic A (2001) Creatine kinase is physically associated with the cardiac ATP-sensitive K+ channel in vivo. FASEB J 16:102–104
Carrasco AJ, Dzeja PP, Alekseev AE et al (2001) Adenylate kinase phosphotransfer communicates cellular energetic signals to ATP-sensitive potassium channels. Proc Natl Acad Sci USA 98:7623–7628
Janssen E, Dzeja PP, Oerlemans F et al (2000) Adenylate kinase 1 gene deletion disrupts muscle energetic economy despite metabolic rearrangement. EMBO J 19:6371–6381
Plant TD (1988) Properties and calcium-dependent inactivation of calcium currents in cultured mouse pancreatic B-cells. J Physiol 404:731–747
Kakei M, Noma A, Shibasaki T (1985) Properties of adenosine-triphosphate-regulated potassium channels in guinea-pig ventricular cells. J Physiol 363:441–462
Nichols CG, Lederer WJ (1990) The regulation of ATP-sensitive K+ channel activity in intact and permeabilized rat ventricular myocytes. J Physiol 423:91–110
Lienhard GE, Secemski II (1973) P1,P5-Di(adenosine-5′)pentaphosphate, a potent multisubstrate inhibitor of adenylate kinase. J Biol Chem 248:1121–1123
Elvir-Mairena JR, Jovanovic A, Gomez LA, Alekseev AE, Terzic A (1996) Reversal of the ATP-liganded state of ATP-sensitive K+ channels by adenylate kinase activity. J Biol Chem 271:31903–31908
Jovanovic A, Alekseev AE, Terzic A (1997) Intracellular diadenosine polyphosphates. A novel family of inhibitory ligands of the ATP-sensitive K+ channel. Biochem Pharmacol 54:219–225
Matschinsky FM, Landgraf R, Ellerman J, Kotler-Brajtburg J (1972) Glucoreceptor mechanisms in islets of Langerhans. Diabetes 21:555–569
Malaisse WJ, Hutton JC, Kawazu S, Herchuelz A, Valverde I, Sener A (1979) The stimulus-secretion coupling of glucose-induced insulin release. XXXV. The links between metabolic and cationic events. Diabetologia 16:331–341
Trus M, Warner H, Matschinsky F (1980) Effects of glucose on insulin release and on intermediary metabolism of isolated perifused pancreatic islets from fed and fasted rats. Diabetes 29:1–14
Ghosh A, Ronner P, Cheong E, Khalid P, Matschinsky FM (1991) The role of ATP and free ADP in metabolic coupling during fuel-stimulated insulin release from islet beta-cells in the isolated perfused rat pancreas. J Biol Chem 266:22887–22892
Malaisse WJ, Sener A (1987) Glucose-induced changes in cytosolic ATP content in pancreatic islets. Biochim Biophys Acta 927:190–195
Farret A, Vignaud M, Dietz S, Vignon J, Petit P, Gross R (2004) P2Y purinergic potentiation of glucose-induced insulin secretion and pancreatic beta-cell metabolism. Diabetes 53(Suppl 3):S63–S66
Rutter GA, Da Silva Xavier G, Leclerc I (2003) Roles of 5′-AMP-activated protein kinase (AMPK) in mammalian glucose homoeostasis. Biochem J 375:1–16
Dzeja PP, Terzic A (2003) Phosphotransfer networks and cellular energetics. J Exp Biol 206:2039–2047
Smink LJ, Helton EM, Healy BC et al (2005) T1DBase, a community web-based resource for type 1 diabetes research. Nucleic Acids Res 33:D544–D549 (database issue)
Larsson O, Ämmälä C, Bokvist K, Fredholm B, Rorsman P (1993) Stimulation of the KATP channel by ADP and diazoxide requires nucleotide hydrolysis in mouse pancreatic beta-cells. J Physiol 463:349–365
Dzeja PP, Terzic A (1998) Phosphotransfer reactions in the regulation of ATP-sensitive K+ channels. FASEB J 12:523–529
Selivanov VA, Alekseev AE, Hodgson DM, Dzeja PP, Terzic A (2004) Nucleotide-gated KATP channels integrated with creatine and adenylate kinases: Amplification, tuning and sensing of energetic signals in the compartmentalized cellular environment. Mol Cell Biochem 256–257:243–256
Olson LK, Schroeder W, Robertson RP, Goldberg ND, Walseth TF (1996) Suppression of adenylate kinase catalyzed phosphotransfer precedes and is associated with glucose-induced insulin secretion in intact HIT-T15 cells. J Biol Chem 271:16544–16552
Ripoll C, Martin F, Manuel Rovira J, Pintor J, Miras-Portugal MT, Soria B (1996) Diadenosine polyphosphates. A novel class of glucose-induced intracellular messengers in the pancreatic beta-cell. Diabetes 45:1431–1434
Acknowledgements
This work was supported by the Deutsche Forschungsgemeinschaft (Dr 225/6-3).
Duality of interest
The authors declare that there is no duality of interest associated with this manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Schulze, D.U., Düfer, M., Wieringa, B. et al. An adenylate kinase is involved in KATP channel regulation of mouse pancreatic beta cells. Diabetologia 50, 2126–2134 (2007). https://doi.org/10.1007/s00125-007-0742-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00125-007-0742-9