
Abstract
Class A G-protein-coupled receptors (GPCRs) are a large family of membrane proteins that mediate a wide variety of physiological functions, including vision, neurotransmission and immune responses1,2,3,4. They are the targets of nearly one-third of all prescribed medicinal drugs5 such as beta blockers and antipsychotics. GPCR activation is facilitated by extracellular ligands and leads to the recruitment of intracellular G proteins3,6. Structural rearrangements of residue contacts in the transmembrane domain serve as ‘activation pathways’ that connect the ligand-binding pocket to the G-protein-coupling region within the receptor. In order to investigate the similarities in activation pathways across class A GPCRs, we analysed 27 GPCRs from diverse subgroups for which structures of active, inactive or both states were available. Here we show that, despite the diversity in activation pathways between receptors, the pathways converge near the G-protein-coupling region. This convergence is mediated by a highly conserved structural rearrangement of residue contacts between transmembrane helices 3, 6 and 7 that releases G-protein-contacting residues. The convergence of activation pathways may explain how the activation steps initiated by diverse ligands enable GPCRs to bind a common repertoire of G proteins.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others


Change history
16 August 2016
A minor change was made to Extended Data Fig. 2e.
References
Bockaert, J. & Pin, J. P. Molecular tinkering of G protein-coupled receptors: an evolutionary success. EMBO J. 18, 1723–1729 (1999)
Lagerström, M. C. & Schiöth, H. B. Structural diversity of G protein-coupled receptors and significance for drug discovery. Nat. Rev. Drug Discov. 7, 339–357 (2008)
Rosenbaum, D. M., Rasmussen, S. G. & Kobilka, B. K. The structure and function of G-protein-coupled receptors. Nature 459, 356–363 (2009)
Venkatakrishnan, A. J. et al. Molecular signatures of G-protein-coupled receptors. Nature 494, 185–194 (2013)
Overington, J. P., Al-Lazikani, B. & Hopkins, A. L. How many drug targets are there? Nat. Rev. Drug Discov. 5, 993–996 (2006)
Katritch, V., Cherezov, V. & Stevens, R. C. Structure-function of the G protein-coupled receptor superfamily. Annu. Rev. Pharmacol. Toxicol. 53, 531–556 (2013)
Rasmussen, S. G. et al. Crystal structure of the β2 adrenergic receptor-Gs protein complex. Nature 477, 549–555 (2011)
Choe, H. W. et al. Crystal structure of metarhodopsin II. Nature 471, 651–655 (2011)
Dror, R. O. et al. Activation mechanism of the β2-adrenergic receptor. Proc. Natl Acad. Sci. USA 108, 18684–18689 (2011)
Huang, W. et al. Structural insights into μ-opioid receptor activation. Nature 524, 315–321 (2015)
Kruse, A. C. et al. Activation and allosteric modulation of a muscarinic acetylcholine receptor. Nature 504, 101–106 (2013)
Lebon, G. et al. Agonist-bound adenosine A2A receptor structures reveal common features of GPCR activation. Nature 474, 521–525 (2011)
Standfuss, J. et al. The structural basis of agonist-induced activation in constitutively active rhodopsin. Nature 471, 656–660 (2011)
Flock, T. et al. Universal allosteric mechanism for Gα activation by GPCRs. Nature 524, 173–179 (2015)
Manglik, A. et al. Structural insights into the dynamic process of β2-adrenergic receptor signaling. Cell 161, 1101–1111 (2015)
Oldham, W. M. & Hamm, H. E. Heterotrimeric G protein activation by G-protein-coupled receptors. Nat. Rev. Mol. Cell Biol. 9, 60–71 (2008)
Dror, R. O. et al. Signal transduction. Structural basis for nucleotide exchange in heterotrimeric G proteins. Science 348, 1361–1365 (2015)
Isberg, V. et al. GPCRdb: an information system for G protein-coupled receptors. Nucleic Acids Res . 44 (D1), D356–D364 (2016)
Gonzalez, A., Cordomí, A., Caltabiano, G. & Pardo, L. Impact of helix irregularities on sequence alignment and homology modeling of G protein-coupled receptors. ChemBioChem 13, 1393–1399 (2012)
Isberg, V. et al. Generic GPCR residue numbers—aligning topology maps while minding the gaps. Trends Pharmacol. Sci. 36, 22–31 (2015)
Breton, B. et al. Multiplexing of multicolor bioluminescence resonance energy transfer. Biophys J . 99, 4037–4046 (2010)
Galés, C. et al. Probing the activation-promoted structural rearrangements in preassembled receptor-G protein complexes. Nat. Struct. Mol. Biol. 13, 778–786 (2006)
Greasley, P. J., Fanelli, F., Rossier, O., Abuin, L. & Cotecchia, S. Mutagenesis and modelling of the α1b-adrenergic receptor highlight the role of the helix 3/helix 6 interface in receptor activation. Mol. Pharmacol. 61, 1025–1032 (2002)
Gabilondo, A. M., Krasel, C. & Lohse, M. J. Mutations of Tyr326 in the β2-adrenoceptor disrupt multiple receptor functions. Eur. J. Pharmacol. 307, 243–250 (1996)
Böhm, S. K. et al. Identification of potential tyrosine-containing endocytic motifs in the carboxyl-tail and seventh transmembrane domain of the neurokinin 1 receptor. J. Biol. Chem. 272, 2363–2372 (1997)
Tello, J. A. et al. Congenital hypogonadotropic hypogonadism due to GnRH receptor mutations in three brothers reveal sites affecting conformation and coupling. PLoS One 7, e38456 (2012)
Isom, D. G. & Dohlman, H. G. Buried ionizable networks are an ancient hallmark of G protein-coupled receptor activation. Proc. Natl Acad. Sci. USA 112, 5702–5707 (2015)
Lans, I., Dalton, J. A. & Giraldo, J. Helix 3 acts as a conformational hinge in class A GPCR activation: An analysis of interhelical interaction energies in crystal structures. J. Struct. Biol. 192, 545–553 (2015)
Berman, H. M. et al. The Protein Data Bank. Nucleic Acids Res. 28, 235–242 (2000)
Venkatakrishnan, A. J. et al. Structured and disordered facets of the GPCR fold. Curr. Opin. Struct. Biol. 27, 129–137 (2014)
Yuan, S., Filipek, S., Palczewski, K. & Vogel, H. Activation of G-protein-coupled receptors correlates with the formation of a continuous internal water pathway. Nat Comms . 5, 4733 (2014)
Sun, D. et al. AAscan, PCRdesign and MutantChecker: a suite of programs for primer design and sequence analysis for high-throughput scanning mutagenesis. PLoS One 8, e78878 (2013)
Acknowledgements
We thank C. Chothia, G. Chalancon, N. Latysheva, R. Henderson, A. Sente, K. Kruse, A. Singhal and S. Chavali for their comments on this manuscript. We acknowledge LMB International Scholarship (in support of A.J.V.), St. John’s College Benefactor Scholarship (A.J.V.), MRC Centenary Early Career Award (A.J.V.), the Medical Research Council (MC_U105185859) (M.M.B., S.B., T.F.); (MC_U105197215) (C.G.T., G.L.), Boehringer Ingelheim Fonds (T.F.), Heptares Therapeutics Ltd (G.L.), ATIP-Avenir program (G.L.), Swiss National Science Foundation (grant number 310030_153145) (G.F.X.S.); (31003A_146520) (X.D.); (31003A_159748) (D.B.V.), Swiss National Science Foundation Doc.Mobility fellowship P1EZP3_165219 (F.M.H.), the Human Frontiers Science Program (RGP0034/2014) (G.F.X.S.), the Canadian Institute for Health Research (MOP-10501) (M.B.) and the COST Action CM1207 (GLISTEN) (X.D., G.F.X.S., C.G.T., G.L.). M.M.B. is a Lister Institute Research Prize Fellow. M.B. holds a Canada Research Chair.
Author information
Authors and Affiliations
Contributions
A.J.V. and M.M.B. designed the project, analysed the data, interpreted the results and wrote the manuscript, with inputs from other authors. A.J.V. collected data, wrote scripts and performed the contact fingerprinting analyses. X.D. and G.L. helped with interpretation of the activation pathways. T.F. helped with the collection of disease mutations. S.B. performed the sequence analysis of GPCRs. D.B.V., M.B., F.M.H. and T.M. designed and performed the mutagenesis and BRET assays and analysed the data. G.F.X.S. and C.G.T. provided general input on the project. M.M.B. supervised the project.
Corresponding authors
Additional information
Reviewer Information Nature thanks D. Gloriam and the other anonymous reviewer(s) for their contribution to the peer review of this work.
Extended data figures and tables
Extended Data Figure 1 Sequence analysis of positions involved in the conserved rearrangement during receptor activation.
The alignment of 311 non-olfactory class A human GPCRs were obtained from GPCRdb. Percentage of residue pairs making a contact in the inactive state only (a, orange spectrum; five bins) and the active state only (b, green spectrum; five bins). The set of large hydrophobic/aromatic residues was defined to include the following amino acids: Val, Leu, Ile, Met, Phe, Tyr, Trp. c, The percentage of amino acids occupying residue positions 3x46, 6x37 and 7x53 involved in the conserved rearrangement is shown. d, The odds of finding a large hydrophobic or aromatic residue at a pair/triplet of positions that form a contact during the conserved rearrangement. See Methods for details.
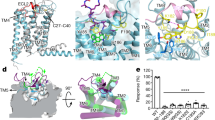
Extended Data Figure 2 Signalling profile of Vasopressin V2 receptor mutants using BRET-based biosensors.
a, b, Activation of Gs (a) and Gq (b) by wild-type and mutant vasopressin V2 receptors. The residues at positions 3x46, 6x37 and 7x53 were mutated to alanine separately and in combination and their G protein signalling response was measured using BRET-based biosensors. The data points represent the mean ± SEM of n = 3–4 experiments. Gs is the primary and Gq the secondary cognate G protein of the vasopressin V2 receptor. For both G proteins, the activation is monitored by the decrease in BRET resulting from the separation between Gα and Gβγ. The G protein response was markedly reduced for all mutants, pointing towards reduced receptor activity. The expression levels of the mutant receptors were quantified using a fluorescent dye bound to the SNAP tag of surface-expressed receptors. While the Y325A (Y7x53A), T273A (T6x37A) and T273A/Y325A (T6x37A/Y7x53A) proteins were expressed at wild-type levels, the expression of M133A (M3x46A) and its combinations showed reduced expression levels. As a comparison, the signalling of reduced levels of wild-type vasopressin V2 receptor (% of DNA transfected) for Gs and Gq is shown in c and d, respectively. This indicates that the reduction in expression level alone cannot explain the reduced signalling of Y325A (Y7x53A), T273A (T6x37A) and the Y325A (Y7x53A)/T273A (T6x37A) combination. Thus, the reduced signalling activity of Y325A (Y7x53A), T273A (T6x37A) and T273A/Y325A (T6x37A/Y7x53A) is due to a change in the ability of the receptor to promote G protein activation. In the case of receptor mutants (single, double and triple) involving M133A (M3x46A), both the G protein response and the expression level of the receptor are markedly reduced. This may be because position 3x46 is involved in mediating contacts that are important for both the active and the inactive state, and mutating this position might simultaneously affect receptor biogenesis and downstream response. e, The increases in EC50 (along with the decrease in maximal responses) are consistent with a reduced ability of the receptor to activate G proteins.
Supplementary information
Supplementary Data
This file contains Supplementary Table 1. (XLSX 37 kb)
Supplementary Data
This file contains Supplementary Table 2. (XLSX 44 kb)
Supplementary Data
This file contains Supplementary Table 3. (XLSX 41 kb)
Rights and permissions
About this article

Cite this article
Venkatakrishnan, A., Deupi, X., Lebon, G. et al. Diverse activation pathways in class A GPCRs converge near the G-protein-coupling region. Nature 536, 484–487 (2016). https://doi.org/10.1038/nature19107
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature19107
This article is cited by
-
Beyond the G protein α subunit: investigating the functional impact of other components of the Gαi3 heterotrimers
Cell Communication and Signaling (2023)
-
Two-step structural changes in M3 muscarinic receptor activation rely on the coupled Gq protein cycle
Nature Communications (2023)
-
Computational investigation of functional water molecules in GPCRs bound to G protein or arrestin
Journal of Computer-Aided Molecular Design (2023)
-
Gαi-derived peptide binds the µ-opioid receptor
Pharmacological Reports (2023)
-
Ultrafast structural changes direct the first molecular events of vision
Nature (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
