


Abstract
Cytochrome P450 (P450) enzyme expression patterns were determined for a panel of 60 human tumor cell lines, representing nine tumor tissue types, used by the National Cancer Institute (NCI) Anticancer Drug Screening Program. All 60 tumor cell lines displayed significant P450 activity, as well as P450 reductase activity, as determined using the general P450 substrate 7-benzyloxyresorufin. Cell line-specific P450 enzyme patterns were observed using three other P450 substrates, 7-ethoxycoumarin, coumarin, and 7-ethoxyresorufin, each of which was metabolized at a low rate. Using a pattern-matching computer program, COMPARE, correlative relationships were investigated between the arrays of P450 activities and the patterns of cytotoxicity exhibited by a large group of anticancer agents of proven or potential clinical utility. Significant negative correlations between the patterns of P450-dependent 7-benzyloxyresorufin metabolism activity and cell line chemosensitivity were observed for 10 standard anticancer agents (including 6 alkylating agents) and 55 investigational compounds, suggesting a role for P450 metabolism in the inactivation of these agents. Negative correlations between 7-ethoxycoumarinO-deethylation and cell line chemosensitivity to a group of topoisomerase inhibitors were also seen, again suggesting P450-dependent drug inactivation. P450 enzyme profiling may thus aid in interpreting the patterns of drug sensitivity and resistance in the NCI tumor cell panel, and may facilitate the identification of anticancer agents whose activity can be altered via cytochrome P450 metabolism.
The cytochrome P450s are a superfamily of hemeprotein monooxygenases that catalyze the oxidative metabolism of a large number of drugs, environmental carcinogens, as well as steroids and other endobiotics. Approximately 60 human cytochrome P450 genes are known (Nelson et al., 1996; Nelson, 1999) and encode proteins that exhibit major differences with respect to their catalytic specificities, tissue-specific patterns of expression, and interindividual differences. These differences result from genetic polymorphisms (Ingelman-Sundberg et al., 1999) and from the differential responsiveness of P450s2 to the large number of foreign chemical inducers and endogenous regulators that control P450 gene expression (Waxman, 1999). P450 enzymes are highly expressed in human liver and in certain extrahepatic tissues and have been studied extensively with respect to their roles in drug metabolism (Rendic and Di Carlo, 1997). Much less is known about the profiles of P450 expression in primary human tumor tissue and in cultured tumor cell lines, in part due to the very low enzyme levels present (Smith et al., 1993; Huang et al., 1996; Nakajima et al., 1996;Murray et al., 1999).
The expression of P450 enzymes in tumor tissue can have a major impact on the responsiveness of tumors to cancer chemotherapeutic drugs, owing to the central role that these enzymes play in the metabolism of numerous clinically useful anticancer agents (LeBlanc and Waxman, 1989). In the case of antitumor prodrugs, such as cyclophosphamide and ifosfamide, P450 metabolism is essential for therapeutic activity (Sladek, 1994). Indeed, the expression in human tumor cells of specific P450 enzymes that activate these oxazaphosphorine prodrugs can greatly sensitize the cells to drug cytotoxicity (Chase et al., 1998; Jounaidi et al., 1998). P450 enzymes can also impact the pharmacokinetics and the therapeutic activity of other classes of anticancer drugs, including those that are converted by P450 to metabolites that retain antitumor activity and those that are inactivated as a consequence of P450 metabolism (LeBlanc and Waxman, 1989; Kivisto et al., 1995).
During the past decade the U.S. National Cancer Institute (NCI) has used a panel of 60 human tumor cell lines, chosen to represent nine different tumor types, to carry out a large in vitro screening program for novel anticancer agents (Boyd and Paull, 1995; Monks et al., 1997). To date more than 60,000 compounds have been characterized with respect to their antitumor activity using this primary screen, generating a series of lead compounds for further investigation and evaluation (Weinstein et al., 1997). Although, as noted above, cytochrome P450 enzymes contribute to the metabolism of a large number of drug substrates and can have a large impact on a drug's anticancer activity, little is presently known about the P450 activity levels present in the individual tumor cell lines that constitute the NCI panel. P450 expression in tumor cells may lead to the localized production of intracellular drug metabolites, and may thereby either increase or decrease the cytotoxicity of test chemicals being evaluated. Characterization of P450 expression patterns within the NCI tumor cell line panel may thus provide insight into some of the factors that govern the cell line-specific and/or tumor type-dependent drug sensitivity and drug resistance patterns seen in these cells. The potential relevance of P450 expression in tumor cells with respect to effective anticancer drug screening is supported by our observation of a dramatic enhancement of the in vitro and in vivo cytotoxic action of the oxazaphosphorine anticancer drug cyclophosphamide when tumor cells are transduced to express the rat CYP2B1 gene (Chen and Waxman, 1995b) or its human P450 ortholog, CYP2B6 (Jounaidi et al., 1998).
In the present study, we sought to characterize the NCI human tumor cell line panel with the following goals: 1) to determine whether cytochrome P450 enzyme activities are expressed at a measurable level in the tumor cell lines constituting this panel; 2) to provide initial information regarding the individual P450 activities present in each of the cell lines; and 3) to search for any correlative relationship between the arrays of P450 enzyme activities displayed by the 60 cell lines and their patterns of chemosensitivity or chemoresistance toward ∼3000 compounds that have been characterized in the NCI in vitro screening program and shown to exhibit reproducible cytotoxic activity. Ultimately, these studies may help ascertain whether P450 enzyme expression patterns can aid in the interpretation of drug sensitivity patterns that individual tumor cell lines exhibit toward cytotoxic agents previously identified by the NCI drug screening program and whose mechanism of action is presently unknown. The present studies complement recent investigations of the same NCI tumor cell line panel that describe the expression of other enzymes potentially relevant to anticancer drug metabolism. These include glutathioneS-transferases and enzymes of glutathione metabolism (Tew et al., 1996), aldehyde dehydrogenases, which can contribute to the inactivation of the activated metabolites of a number of cancer chemotherapeutic agents including cyclophosphamide and ifosfamide (Sreerama and Sladek, 1997), and DT-diaphorase and other enzymes that catalyze bioreductive metabolism of a variety of quinones and related chemicals (Fitzsimmons et al., 1996).
Materials and Methods
Isolation of Microsomes from Tumor Cell Lines.
The 60 human tumor cell lines used in this study are described elsewhere (Boyd, 1989; Monks et al., 1997) (see listing in Table 2, below). Cells at passage number ranging from 3 to 17 were harvested in mid-log growth phase. Microsomes were prepared from frozen cell pellets in procedures carried out at 0 to 4°C. Cell pellets were suspended in ice-cold 0.1 M KPi (pH 7.4) buffer containing 0.1 mM EDTA and 20% (v/v) glycerol. The samples were then homogenized, sonicated three times (2–10 s/sonication at a moderate instrument setting), and centrifuged at low speed (15 min at 8000g). The supernatant was then centrifuged for 1 h at 140,000g. Final microsomal pellets were resuspended in the above KPi/EDTA/glycerol buffer to give a protein concentration ≥5 mg/ml with a yield that ranged from 1 to 12 mg of microsomal protein per 5 × 108 cells. Microsomes were stored in aliquots at −80°C.
Summary of P450 activities measured in the NCI cell line panel
NADPH-Cytochrome P450 (Cytochrome c) Reductase Assay.
P450 reductase activities were measured spectrophotometrically at 550 nm. Cytochrome c (Sigma Chemical Co., St. Louis, MO), reduced by P450 reductase in the presence of NADPH, has a chromophore that absorbs visible light at 550 nm with ε = 21,000 M−1 cm−1. Reactions were carried out in a cuvette containing cytochrome c (42.7 μM), 0.3 M KPi buffer, pH 7.7, 25 μg of microsomal protein, and 120 μM NADPH in a final volume of 1 ml. Reactions were initiated by the addition of NADPH, and the change inA550 nm was monitored at room temperature for 5 min. Data obtained were shown to reflect initial reaction rates, and represent two to three replicate assays for each sample.
7-ECOD Assay.
Activity was assayed in 100 mM KPi buffer (pH 7.4), 20% glycerol, 0.1 mM EDTA, with 1 mM 7-ethoxycoumarin (Aldrich Chemical Co., Milwaukee, WI) and 200 μg of microsomal protein in a total volume of 200 μl. Reactions were initiated by adding NADPH to 1 mM. Reactions were incubated for 1 h at 37°C with gentle shaking then terminated by adding 25 μl of ice-cold 2 M HCl. The samples were then extracted twice with 450 μl of chloroform. The chloroform layers were combined and then back-extracted with 1 ml of 30 mM sodium borate, pH 9.2. The 7-hydroxycoumarin metabolite was determined fluorometrically (370-nm excitation wavelength, 450-nm emission wavelength) in comparison to authentic 7-hydroxycoumarin standard (Aldrich Chemical Co.). Data presented are based on two to three replicate assays for each sample.
Coumarin 7-Hydroxylation Assay.
Reaction mixtures contained reagents and microsomes assayed in the same conditions and concentrations as described for the 7-ECOD assay, but using 1 mM coumarin (Sigma Chemical Co.) in place of 7-ethoxycoumarin as the substrate. Enzyme incubation, extraction of the 7-hydroxycoumarin metabolite, and fluorometric analysis of enzyme activity were also performed using the same conditions as the 7-ECOD assay. Data shown generally reflect averages of duplicate assays for each sample.
Alkoxyresorufin O-Dealkylation Assays.
Reactions used to monitor 7-EROD and 7-BROD activities (total volume, 2.5 ml) were carried out in a 3-ml fluorometer cell containing a microstirring bar and 250 μg of microsomal protein. Samples were mixed with 4 μM substrate (7-ethoxyresorufin or 7-benzyloxyresorufin, delivered using 10 μl of 1 mM stock solution in dimethyl sulfoxide; Molecular Probes, Eugene, OR) in 0.1 M KPi (pH 7.4) and 0.1 mM EDTA buffer at room temperature. Reactions were started by the addition of NADPH to 250 μM. Formation of the fluorescent metabolite, resorufin, from either 7-ethoxyresorufin or 7-benzyloxyresorufin was measured at room temperature over an 8-min period using a Shimadzu RF-1501 fluorescence spectrophotometer (Shimadzu, Kyoto, Japan). Fluorescence was read at 550 nm (excitation) and 586 nm (emission). Activity values were quantitated using the 7-hydroxylated standard, resorufin (Molecular Probes). Data obtained were shown to correspond to initial reaction rates and generally represent two to three assays for each sample.
Drug Screening and COMPARE Analysis.
The COMPARE program (Paull et al., 1989, 1995) has identified drugs with common mechanisms of action. The molecular target version of this program (Lee et al., 1994; Monks et al., 1997) was used here to analyze possible relationships between individual P450 enzyme activities and the cell line sensitivity patterns of standard agents. The standard agent database comprises 170 chemicals for which a considerable amount of information is available in terms of preclinical and/or clinical antitumor properties and presumed mechanism of action (Boyd and Paull, 1995; Paull et al., 1995). In addition, correlative analysis was undertaken between target patterns and a database of approximately 3000 active investigational compounds, whose cytotoxicity or growth-inhibitory activity has been confirmed in more than one series of in vitro cell line screenings. The relative sensitivities of the panel of 60 cell lines to a given compound, at a concentration causing 50% growth inhibition, are represented as a mean-graph pattern (Paull et al., 1989). In the molecular target version of the pattern recognition program COMPARE (Lee et al., 1994; Alvarez et al., 1995), each target measurement (e.g., P450 reductase or one of the measured P450 activities) is represented in a manner similar to a mean-graph and used as a seed to derive correlations between toxicity patterns in the various databases and the pattern of expression of the P450 enzymes. The compounds that correlated to the target pattern, either positively or negatively, were ranked by Pearson correlation coefficient (PCC) andp value. A positive PCC indicates that greater activity of the target enzyme may be associated with increased cell sensitivity to the drug. In contrast, a negative PCC implies that greater activity of the target enzyme may confer cellular resistance to the given drug. To evaluate compounds of possible interest, the uncorrected two-tailp value was set at <0.0012 for the standard agents (n = 170) and at <6.7E-05 for the investigational database (n = ∼3000), which reflects the equivalent of p < 0.2 after the Bonferroni adjustment for multiple comparisons. Using the assigned criteria, the probability of such occurrence from the selected database by random chance would be approximately 20%.
Correlative analysis was also carried out between the target P450 enzyme patterns and compounds classified according to each of six major clinical mechanisms (van Osdol et al., 1994): alkylating agents (n = 35); “anti-DNA agents” including compounds directly incorporated into DNA, polymerase inhibitors, and ribonucleotide reductase inhibitors (n = 16); nucleotide synthesis inhibitors including antifolates and antimetabolites (n = 19); topoisomerase I inhibitors, all of which are camptothecin analogs (n = 23); topoisomerase II inhibitors (n = 16); and tubulin active antimitotic agents (n = 13). Using compounds grouped within a single mechanism of action (van Osdol et al., 1994) to produce a general pattern of response, each target was correlated with these general mechanisms, and if the upper and lower limits of the 95% confidence interval pass through zero, then there was no positive or negative association suggested. However, a significant association may be suggested if positive correlations have a lower limit of >0.1 and negative confidence limits have an upper confidence limit of <−0.1.
Data presented in Figs. 1 through 5 are posted and can be accessed at the web sitehttp://dtp.nci.nih.gov/servlet/gcDisplaySearch?aliasStr=Waxman.
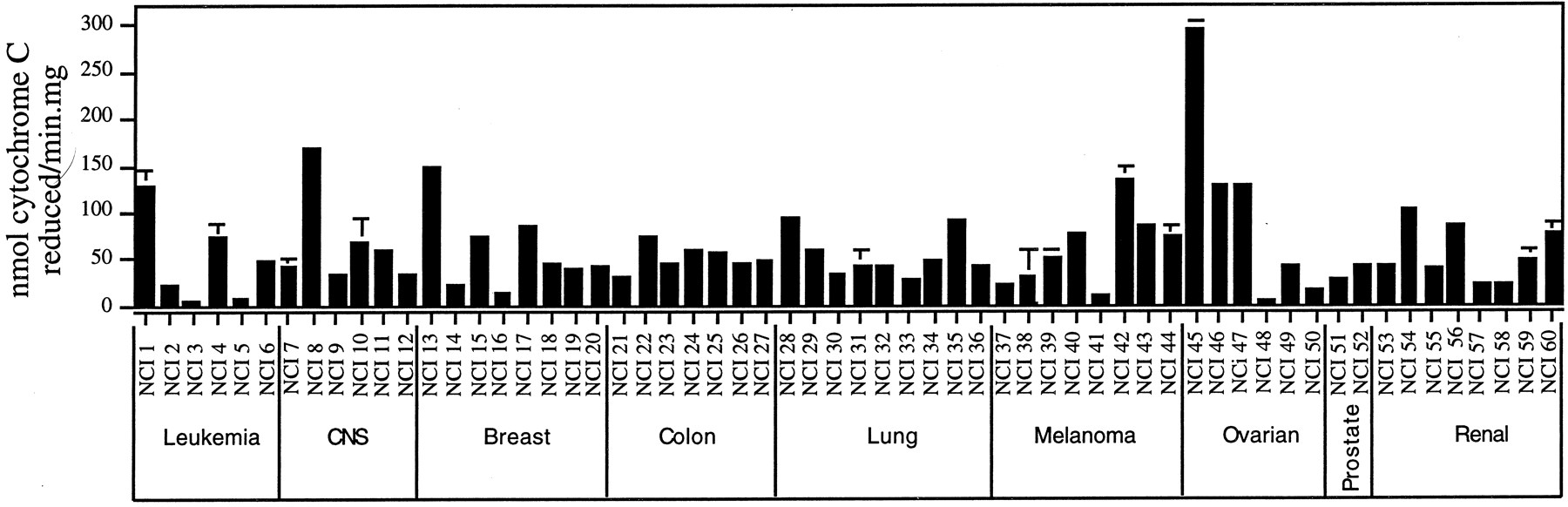
NADPH-P450 reductase activity of NCI human tumor cell line microsomes.
Enzyme activities were measured by monitoring NADPH-dependent cytochrome c reduction as described underMaterials and Methods. Data reflect mean ± range values for two to three replicate assays of each cell line using two independently prepared microsome aliquots. For many samples the error bars are too small to be seen. NCI cell lines are numbered as shown in Table 2.

7-BROD activity of NCI cell line microsomes.
Data shown reflect mean ± range for two to three assays of each cell line. Background activity values were measured for select cell line microsomes in assays carried out without NADPH or using bovine serum albumin in place of microsomal protein. Error bars are too small to be seen for many of the cell lines.
Results
NADPH-P450 Reductase Activity in the NCI Panel.
P450 reductase activity was readily measured in all 60 NCI cell line microsomes (Fig. 1). The mean specific activity for the panel of cell microsomes was 60.2 ± 6.1 nmol/min/mg of protein (mean ± S.E.), and the individual values ranged from 5 to 294 nmol of cytochrome c reduced/min/mg of protein. Because P450 reductase is an obligatory, and often rate-limiting, enzymatic component of microsomal P450 metabolism, P450 reductase levels are likely to be an important codeterminant of the P450 activity of those tumor cell lines that express one or more cytochrome P450 proteins. Analysis of P450 reductase activity levels on the basis of tumor cell type did not reveal any significant associations between P450 reductase activity and tissue of origin (data not shown).
COMPARE analysis revealed that the P450 reductase activity pattern of the cell line panel correlated negatively with cell sensitivity patterns to two established anticancer agents,l-asparaginase and fludarabine phosphate (p < 0.0012). In contrast, examination of the database of 3000 investigational compounds revealed no correlations that met the p value cutoff set for that group (seeMaterials and Methods).
7-ECOD Metabolism by NCI Cell Line Microsomes.
7-Ethoxycoumarin O-deethylation is a sensitive microsomal reaction that allows detection of two human P450s that metabolize this substrate at a high rate, CYP1A1 and CYP2E1. Several other human P450s metabolize 7-ethoxycoumarin at much lower rates (Table 1) (Waxman et al., 1991). Forty-nine of the 60 NCI cell lines were active in the 7-ECOD reaction (Fig. 2). 7-ECOD activity was below the limits of detection (<0.15 pmol/min/mg) in the other 11 cell lines. The melanoma cell line SK-MEL-2 and the ovarian cancer cell line OVCAR-4 were particularly active, followed by the colon cell line KM12. Overall, the highest average 7-ECOD activities were seen in the melanoma and prostate tumor cell line groups, while the lowest activities were seen in the central nervous system tumors and leukemia groups. However, these trends did not reach statistical significance as a consequence of heterogeneity of 7-ECOD activity within each tumor group.
P450 enzyme activities exhibited by a panel of human lymphocyte microsomes containing cDNA-expressed human P450s
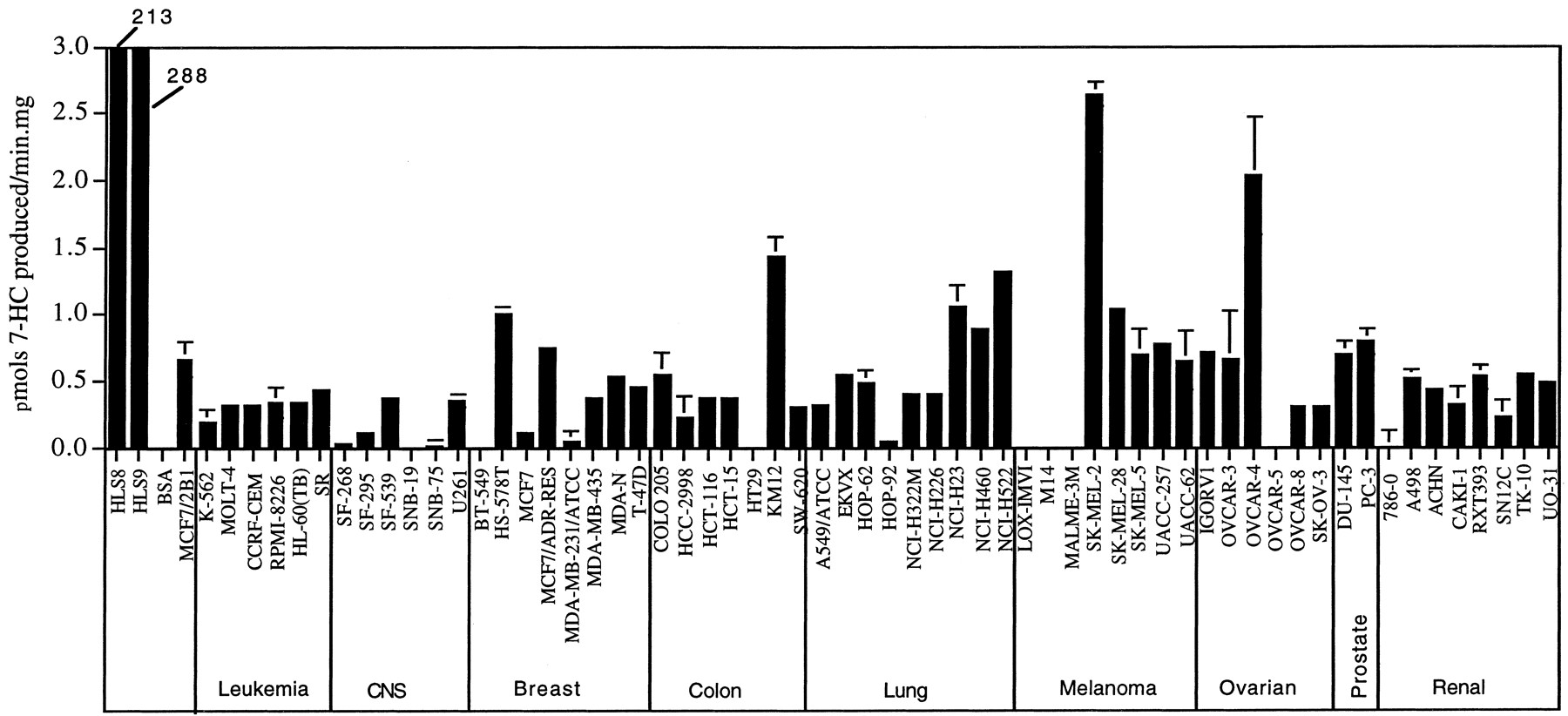
7-ECOD activity of NCI cell line microsomes.
Data reflect two to three replicate assays for each cell line. Background activity values, determined using bovine serum albumin in place of microsomal protein, were subtracted from each of the measured activities to give the indicated net specific activities. Error bars represent the range of specific activity values, and for many cell lines are too small to be seen. Also shown for comparison (samples at left) are the 7-ECOD activities of human liver microsome samples HLS8 and HLS9 and the 7-ECOD activity of an MCF7 cell line stably transfected with rat P450 2B1 (Chen et al., 1996). 7-HC, 7-hydroxycoumarin.
COMPARE analysis revealed that 7-ECOD activity exhibited weak negative associations with the cytotoxicity patterns of several established anticancer drugs, but none reached the defined level of statistical significance (see Materials and Methods). Examination of the database of 3000 investigational compounds identified one compound, a benzopyran (NSC 669781) whose cytotoxicity toward the cell panel showed a strong negative correlation with the pattern of 7-ECOD activity (PCC = −0.54 and p value = 2.7E-05), suggesting that this agent may be inactivated by CYP1A1 or CYP2E1, the two most active catalysts of the 7-ECOD reaction.
Coumarin 7-Hydroxylase Activity in the NCI Panel.
This reaction is actively catalyzed by CYP2A6, with CYP2B6 also catalyzing this reaction, albeit at a much lower rate (Table 1) (Waxman et al., 1991). Measurable coumarin 7-hydroxylase activity (≥0.1 pmol/min/mg) was observed in 43 of the 54 cell lines tested (Fig.3). A colon tumor cell line, HCT-15, showed highest activity (0.76 pmol of 7-hydroxycoumarin produced/min/mg), but no coumarin 7-hydroxylase activity was detected in several of the leukemia, lung, ovarian, and renal tumor cell lines (Fig. 3). No statistically significant correlations between the coumarin 7-hydroxylase activity pattern and the patterns of sensitivity to any of the established or investigational agents were detected, indicating that none of these compounds are metabolized by CYP2A6, or alternatively, that CYP2A6 does not contribute to the cellular sensitivity or resistance of these compounds at its relatively low level of expression in the tumor cell panel.

Coumarin 7-hydroxylase activity of NCI cell line microsomes.
Data reflect duplicate assays of each cell line, generally obtained using two independently prepared microsomal aliquots for each cell line. A bovine serum albumin activity background value was subtracted from the activity of each microsomal sample to give the net specific activities shown. The low activity values shown here were verified for select cell lines in (−)-NADPH control experiments. Error bars represent the range of specific activity values, and for many cell lines the error bars are too small to be seen. Shown for comparison at the left is the high coumarin 7-hydroxylase activity of cDNA-expressed CYP2A6. *Samples exhibited mean activities below the detection limit (∼0.1 pmol/min/mg). 7-HC, 7-hydroxycoumarin.
7-EROD Activity of the NCI Panel.
This P450 reaction is actively catalyzed by CYP1A1, with a lower but measurable activity exhibited by CYP1A2 and -1B1, as demonstrated using cDNA-expressed human P450 enzymes (Table 1). A broad range of chemical carcinogens, including several of the major chemical carcinogens found in cigarette smoke and auto exhaust, can induce CYP1A1 in various tissues, including lung. Although CYP1A1 is not commonly detected in cultured cell lines in the absence of exposure to P450 inducers, CYP1A1 activity has not been assayed in the NCI 60-cell line panel. In this context, it is notable that eight of the nine NCI lung cancer cell lines exhibited 7-EROD activity (Fig.4). The highest 7-EROD activities (∼3–5 pmol of resorufin produced/min/mg of protein) were observed for the ovarian cancer line OVCAR-4, for the colon cancer line KM12, and for two lung tumor cell lines, A549/ATCC and NCI-H23. 7-EROD activity was below the limit of detection (∼0.1 pmol/min/mg) for 19 of the 60 cell lines. Mean 7-EROD activity values from the seven colon tumor cell lines were ∼5-fold higher than that from the eight renal and six leukemia cell lines. The absolute level of 7-EROD activity in the panel was low, however, consistent with the general finding that CYP1A1 expression is low in cells not exposed to aromatic hydrocarbon inducers.
7-EROD activity of NCI cell line microsomes.
Data shown are based on two to three replicate assays for each cell line, generally using two independently prepared microsomal aliquots for each cell line. Several of the microsomal samples yielded negative slopes in the continuous 7-EROD fluorescent assay, resulting in calculated negative activity values (*). Error bars represent the range of specific activity values, and for many cell lines the error bars are too small to be seen. Shown for comparison at the left are the 7-EROD activities of human liver microsome sample HLS8 and cDNA-expressed human CYP1A2. Detection limit = 0.1 pmol/min/mg.
COMPARE analysis did not reveal any notable correlations between 7-EROD activity and anticancer drug sensitivity. One compound from the active database, the microtubule assembly inhibitor norhomohalichondrin B (NSC 700368), was negatively associated with 7-EROD activity.
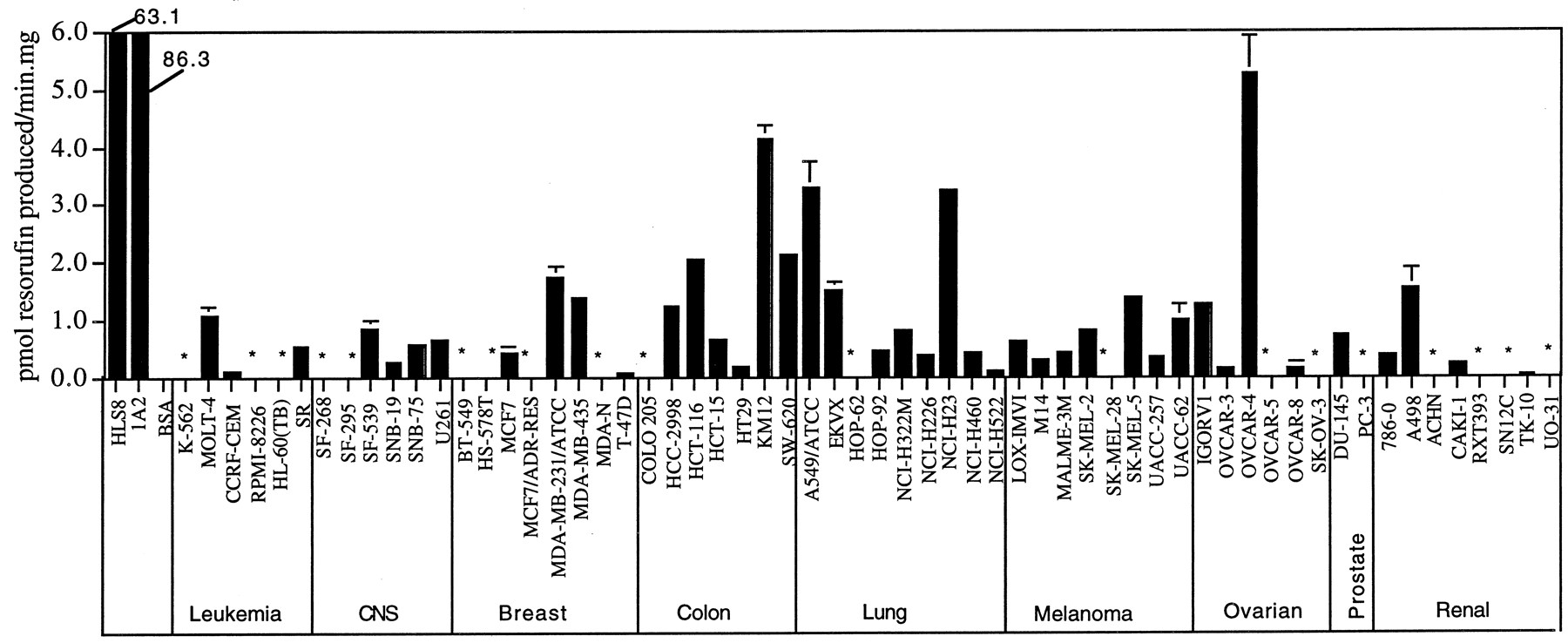
7-BROD Activity Patterns in the NCI Cell Line Panel.
The 7-BROD assay is a more general assay for cytochrome P450 enzymes than the three other P450 enzyme assays used in this study. This assay measures the activity of multiple human P450s, including CYP1A1, -1B1, -2B6, -2C8, -2C9, -2C19, -2D6, and -3A4 (Table 1). All 60 of the NCI tumor cell lines showed 7-BROD activity that was readily measurable and well above background (Fig. 5), as verified in control incubations containing bovine serum albumin in place of cell microsomes, or complete assay mixtures incubated without NADPH. Analysis of the 7-BROD activity data on the basis of the tissue of tumor origin indicated that the lowest 7-BROD activities were present in the six leukemia cell lines.
By COMPARE analysis, 7-BROD activity was negatively correlated with 10 standard anticancer agents (p < 0.002), six of which are classified as alkylating agents (Table 3). Moreover, COMPARE analysis using the investigational database identified 55 active compounds, all of which were negatively correlated with 7-BROD activity (PCC ≥ 0.49 and p < 6.7E-05). These negative correlations suggest that increased 7-BROD activity may confer resistance to these agents, perhaps via a P450-dependent metabolic inactivation reaction.
Correlation between 7-BROD activity pattern and standard agent sensitivity patterns
Overall Comparisons of P450 Activity Patterns.
Table 2 presents an overall comparison of the four microsomal P450 enzyme activities and P450 reductase activity measured for the NCI tumor cell line panel. Sixteen of the tumor cell lines gave good positive activities for all four P450 activities, indicating that multiple P450 enzymes are likely to be expressed in each of these cell lines. The ovarian cancer cell line OVCAR-4 gave the highest overall P450 activity with the four substrates tested. The high P450 activity of this cell line is not solely a reflection of its high P450 reductase activity, as several other cell lines in the panel displayed similarly high P450 reductase activity but did not exhibit the consistently high P450 enzyme activities seen in OVCAR-4.
Cell lines positive for 7-ECOD metabolism were not necessarily positive for coumarin 7-hydroxylation [e.g., cell lines K-562, RPMI-8226, HL60(TB)], consistent with the broader catalytic participation of human P450s in the 7-ECOD reaction (cf. Table 1). Several strong 7-ECOD positive cell lines, such as the breast cell line HS-578T, the melanoma cell line SK-MEL-28, and the prostate cell line PC-3, did not show measurable 7-EROD activity (Table 2), suggesting that these cell lines are functionally devoid of CYP1A activity. Several cell lines (SNB-19, M14, NALME-3M, and 786–0) were negative for 7-ECOD assay but showed substantial activities in the coumarin hydroxylase, 7-EROD, and 7-BROD assays. These findings demonstrate that unique P450 expression patterns characterize each of the individual tumor cell lines.
Comparison of the breast carcinoma cell line MCF7 with its doxorubicin (Adriamycin)-resistant subline MCF7/ADR-RES revealed a decrease in P450 reductase activity, which has been seen earlier (Chen and Waxman, 1995a). Also seen were decreases in 7-EROD and 7-BROD activity and an increase in 7-ECOD activity (Table 2). Multiple changes in gene expression characterize MCF7/ADR-RES cells, but these changes do not necessarily contribute to the cell's drug-resistant phenotype (Chen and Waxman, 1995a).
To further investigate which specific P450 enzymes are expressed and at what relative levels, we used a panel of seven anti-human P450 antibodies (anti-CYP1A1, -1A2, -2A6, -2B6, -2C, -2E1, and -3A) to analyze microsomes prepared from 11 of the tumor cell lines by Western blotting using enhanced chemiluminescence detection. In all cases, strong P450 form-specific signals were obtained when human liver microsomes (20 μg of protein) or cDNA-expressed P450s (1–5 pmol) were used as positive controls on the blots. However, none of the Western blots showed detectable P450 protein signals with the tumor cell microsomes (50–75 μg of protein) (data not shown). This observation is in agreement with the very low level of corresponding P450 mRNAs seen in the same tumor cell line panel by microarray-based expression profiling (Ross et al., 2000).
With the exception of 7-BROD activity, the P450 activity values presently described for the NCI cell line panel are all very low when compared with typical human liver microsomal P450 activities. For example, the 7-ECOD activities of the most active NCI tumor cell line microsomes correspond to only ∼1% of the specific catalytic activity of typical human liver microsomes assayed in our laboratory (Figs. 2and 4). This may be reflected in the lack of compounds whose cytotoxicity patterns correlate with these target activity patterns; furthermore, it is consistent with our inability to detect in these cells P450 enzyme proteins by Western blotting, as noted above.
Associations with Mechanisms of Action and Other Molecular Targets.
Correlative analyses were carried out to identify any possible associations between the measured cell panel P450 activity patterns and anticancer drugs grouped according to each of six major clinical mechanisms of action (van Osdol et al., 1994). Potentially meaningful negative correlations were found between 7-ECOD activity patterns and a group of 23 camptothecin analog topoisomerase I inhibitors (95% confidence limits from −0.34 to −0.22). The 7-ECOD activity pattern was more weakly associated (negatively) with the topoisomerase II inhibitor group (95% confidence limits from −0.25 to −0.11). The P450 catalysts of 7-ECOD activity might therefore confer resistance to compounds with topoisomerase I or II inhibition activities. 7-BROD activity was negatively correlated and therefore potentially associated with resistance to topoisomerase I-inhibiting camptothecin analogs (95% confidence limits from −0.32 to −0.20), anti-DNA agents (95% confidence limits from −0.32 to −0.18), and alkylating agents (95% confidence limits from −0.30 to −0.21). This latter association was further emphasized by the significant association between 7-BROD activity and six standard alkylating agents (Table3), as noted above.
Finally, analyses were carried out to determine whether any of the four sets of P450 activity measurements were coordinately expressed with any of the molecular targets previously measured in the NCI cell line panel for which data are presently available in the public domain. These include various oncogenes, tumor suppressor genes, drug transporters, cytokines, cell cycle molecules, DNA repair enzymes, and metabolic enzymes (Monks et al., 1997). No associations were found, other than correlations between P450 reductase and 7-BROD activity and between 7- EROD and 7-BROD activity (p < 0.05). The former correlation supports the conclusion that P450 reductase can be rate limiting for P450-dependent metabolism in the NCI cell line panel.
Discussion
The ultimate goal of the NCI anticancer drug screening program is to discover new anticancer agents by large-scale screening of libraries of compounds for their ability to inhibit tumor cell growth in a broad-based panel of human tumor cell lines. The cytotoxicity of agents identified in these in vitro assays can be greatly affected by the expression of one or more cellular components, such as transport proteins or enzymes that can activate or deactivate the potential anticancer drug (Weinstein et al., 1997). Characterization of the NCI panel with respect to these “drug responsiveness” determinants may help identify factors that determine the selectivity of a given agent for certain tumor cell lines in the panel. These studies may also aid in the discovery of new molecular targets and may facilitate the design of new chemotherapeutic strategies based on the altered expression (either increased or decreased expression) of the drug response determinant. Efforts have been made in recent years to profile the expression patterns of various drug responsiveness determinants in the NCI panel, including aldehyde dehydrogenases (Sreerama and Sladek, 1997), various reductases (Fitzsimmons et al., 1996), glutathione-associated enzymes (Tew et al., 1996), the drug transporters mdr-1/P-glycoprotein and multidrug resistance-associated protein (Alvarez et al., 1995, 1998), the tumor suppressor gene p53 (O'Connor et al., 1997), and the inhibitor of apoptosis protein family (Tamm et al., 1998). These studies have led to several interesting findings. For example, a high correlation was observed between drug sensitivity of a series of compounds found in NCI's open database of ∼30,000 compounds and the P-glycoprotein-dependent mechanism of resistance (Alvarez et al., 1995). The validity of this approach was established in experiments demonstrating that the compounds identified in this manner are, in fact, P-glycoprotein substrates (Alvarez et al., 1995). Similarly, the correlation between transcript levels of γ-glutamyl cysteine synthetase and certain standard agents has suggested an association between the capacity of cells to synthesize glutathione and their resistance to alkylating agents (Tew et al., 1996).
This study presents the first systematic investigation of human P450 enzyme activity patterns in a large panel of human tumor cell lines. These initial findings, with the support of future, more comprehensive studies using additional P450 form-selective enzyme assays (Chang and Waxman, 1998) and more sensitive detection methods for P450 expression (e.g., reverse transcriptase-polymerase chain reaction), may provide for a more complete understanding of the characteristic patterns of anticancer drug responsiveness exhibited by each cell line. Such information may be helpful in the identification of lead anticancer agents that are inactivated, or alternatively, are activated via P450 metabolism. This concept is supported by the uniformly negative correlation between cellular 7-BROD activity and each of 10 standard antitumor agents and 55 other confirmed active anticancer drugs, which indicates a role for this P450 activity in determining cellular resistance to these groups of agents. P450 activity can also enhance chemosensitivity, as previously demonstrated with the human tumor cell line MCF7/2B1, a derivative of MCF7 breast carcinoma that expresses P450 form CYP2B1 at a relatively low level yet exhibits significant cytotoxic sensitization to cyclophosphamide, both in vitro and in a nude mouse xenograft model (Chen and Waxman, 1995b; Chen et al., 1996). Tumor cell lines engineered to express specific P450s could potentially be very helpful if incorporated into the NCI anticancer drug screening program in identifying novel anticancer prodrugs that undergo P450 metabolism but which would likely escape detection using the current panel of human tumor cell lines.
Recently, Fitzsimmons et al. (1996) reported NADPH-P450 reductase activity in S9 supernatant fractions prepared from the same panel of 60 NCI tumor cell lines examined in this study. The S9 fraction P450 reductase activities reported in that study are severalfold lower than the microsomal P450 reductase activities described in this report, consistent with the expected enrichment of P450 reductase in the microsomal fraction. In addition, the pattern of P450 reductase activities in this panel of NCI cell lines reported by Fitzsimmons et al. (1996) is different from what we observed in our study. Although the basis for this discrepancy is uncertain, it is possible that other interfering enzyme activities present in the S9 supernatant could be a contributing factor. The pattern of P450 reductase activity seen in the present study was reasonably well correlated with the toxicity pattern of two standard agents (l-asparaginase and fludarabine phosphate), which was not found in the study of Fitzsimmons et al. (1996).
The correlations between the arrays of P450 enzyme activities and the patterns of toxicity of standard agents or database compounds described in this report may aid in the design of further experiments to evaluate new hypotheses regarding the role of P450 enzymes in metabolism of select anticancer agents. Follow-up studies of the metabolism of standard agents and database compounds selected by the COMPARE algorithm using panels of cDNA-expressed human P450 enzymes are likely to be informative in this regard. Similarly, it will be of interest to further investigate three tumor cell lines that are reported to be sensitive to either cyclophosphamide or ifosfamide, namely, renal carcinoma cell line RXF-393 and nonsmall cell lung carcinoma cell lines NCI-H226 and NCI-H522, in view of the apparent requirement for P450 activity to manifest the latent cytotoxic potential of these anticancer prodrugs (Sreerama and Sladek, 1997). Conceivably, such studies may reveal the expression in these cells of one or more oxazaphosphorine-activating human P450 enzymes (Chang et al., 1993; Roy et al., 1999). Finally, caution should be exercised when extrapolating the present findings to in vivo tumor models. An earlier report (Smith et al., 1993) indicated that P450 expression patterns in human colon and breast tumor lines can change in response to a variety of factors when the cells are grown as xenografts in immunodeficient mice. In addition, the cellular profile of P450 activities measured in tumor cell lines such as the NCI 60 panel may be very different from those present in vivo in the tumor tissue of origin due to a variety of factors, including the variable loss of expression of individual P450s that is often seen in cultured cells (Alexandre et al., 1990). Cellular P450 activities can also be influenced by many factors including cell passage number, growth phase, culture conditions, and origin of the tumor tissue used initially to develop the cell line. Thus, while the P450 activity profiles described in this study may not be representative of the corresponding parent tumor tissue, they are nevertheless informative with respect to interpretation of the drug sensitivity patterns of the cell lines constituting this anticancer drug screening panel.
Footnotes
-
Send reprint requests to: Dr. David J. Waxman, Dept. of Biology, Boston University, 5 Cummington St., Boston, MA. E-mail:djw{at}bio.bu.edu
-
↵1 Current address: Central Research Division, Pfizer Inc., Groton, CT.
-
These studies were supported in part by Contract SAIC 97-CX-50351A and by National Institutes of Health Grant CA49248 (to D.J.W.). Support was provided in whole or in part with Federal funds from the National Cancer Institute, National Institutes of Health, under Contract N01-CO-56000.
-
Disclaimer: The content of this article does not necessarily reflect the views or policies of the Department of Health and Human Services, nor does mention of trade names, commercial products, or organization imply endorsement by the U.S. government.
- Abbreviations used are::
- P450 or CYP
- cytochrome P450
- NCI
- U.S. National Cancer Institute
- P450 reductase
- NADPH-cytochrome P450 oxidoreductase
- 7-ECOD
- 7-ethoxycoumarinO-deethylase
- 7-EROD
- 7-ethoxyresorufinO-deethylase
- 7-BROD
- 7-benzyloxyresorufinO-debenzylase
- PCC
- Pearson correlation coefficient
- Received July 6, 2000.
- Accepted October 26, 2000.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}