


Abstract
A variety of polycyclic aromatic hydrocarbons and their dihydrodiol derivatives, arylamines, heterocyclic amines, and nitroarenes, were incubated with cDNA-based recombinant (Escherichia coli or Trichoplusia ni) systems expressing different forms of human cytochrome P450 (P450 or CYP) and NADPH-P450 reductase usingSalmonella typhimurium tester strain NM2009, and the resultant DNA damage caused by the reactive metabolites was detected by measuring expression of umu gene in the cells. Recombinant (bacterial) CYP1A1 was slightly more active than any of four CYP1B1 allelic variants, CYP1B1*1, CYP1B1*2, CYP1B1*3, and CYP1B1*6, in catalyzing activation of chrysene-1,2-diol, benz[a]anthracene-trans-1,2-, 3,4-, 5,6-, and 8,9-diol, fluoranthene-2,3-diol, dibenzo[a,l]pyrene, benzo[c]phenanthrene, and dibenz[a,h]anthracene and several arylamines and heterocyclic amines, whereas CYP1A1 and CYP1B1 enzymes had essentially similar catalytic specificities toward other procarcinogens, such as (+)-, (−)-, and (±)-benzo[a]pyrene-7,8-diol, 5-methylchrysene-1,2-diol, 7,12-dimethylbenz[a]anthracene-3,4-diol, dibenzo[a,l]pyrene-11,12-diol, benzo[b]fluoranthene-9,10-diol, benzo[c]chrysene, 5,6-dimethylchrysene-1,2-diol, benzo[c]phenanthrene-3,4-diol, 7,12-dimethylbenz[a]anthracene, benzo[a]pyrene, 5-methylchrysene, and benz[a]anthracene. We also determined activation of these procarcinogens by recombinant (T.ni) human P450 enzymes in S.typhimurium NM2009. There were good correlations between activities of procarcinogen activation by CYP1A1 preparations expressed in E. coli and T.ni cells, although basal activities with three lots of CYP1B1 in T. ni cells were very high without substrates and NADPH in our assay system. Using 14 forms of human P450s (but not CYP1B1) (in T. nicells), we found that CYP1A2, 2C9, 3A4, and 2C19 catalyzed activation of several of polycyclic aromatic hydrocarbons at much slower rates than those catalyzed by CYP1A1 and that other enzymes, including CYP2A6, 2B6, 2C8, 2C18, 2D6, 2E1, 3A5, 3A7, and 4A11, were almost inactive in the activation of polycyclic aromatic hydrocarbons examined here.
A variety of polycyclic aromatic hydrocarbons (PAHs1) are ubiquitous environmental carcinogens and have been shown to cause cellular transformations only after metabolic activation by so-called drug-metabolizing enzymes, such as cytochrome P450 (P450 or CYP) and epoxide hydrolase, that produce highly reactive carcinogenic electrophiles (Gelboin, 1980; Pelkonen and Nebert, 1982). In most cases, oxidation of PAHs by P450 enzymes is an initial step in the activation process. The resultant epoxide intermediates are usually more reactive than the parent compounds and have been shown to require further metabolism to evoke their critical carcinogenic potentials (Wood et al., 1976; Kapitulnik et al., 1978). These epoxide metabolites have been shown to be readily hydrolyzed to dihydrodiol metabolites (the suffix “dihydrodiol” or “diol” is used in the text to designate the prefix “dihydroxydihydro” for individual polycyclic hydrocarbons) by microsomal epoxide hydrolases and finally oxidized again by P450 enzymes to form highly reactive diol-epoxides (namely, bay- and fjord-region epoxides) that interact with DNA to initiate cell transformation (Conney, 1982). In these events, P450 enzymes play key roles, and efforts have been made to identify which P450 species are more active in catalyzing activation of PAHs and PAH dihydrodiols in experimental animals and humans (Pelkonen and Nebert, 1982; Shimada et al., 1989a,b; Gonzalez et al., 1990; Jacob et al., 1996; Shou et al., 1996a,b; Einolf et al., 1997).
Among the various forms of P450 determined so far, CYP1A1 and CYP1B1 have been shown to be the most important human P450 enzymes in the metabolic activation of PAHs and PAH dihydrodiols (Hall et al., 1989;Schmalix et al., 1993; Gautier et al., 1996; Shimada et al., 1996,1999a; Kim et al., 1998; Luch et al., 1998). Both enzymes are expressed predominantly in extrahepatic organs, such as lungs and mammary glands, and it has been suggested that levels of expression and catalytic properties of these P450 enzymes may determine different susceptibilities of individuals toward lung and breast cancers caused by these procarcinogens (Huang et al., 1996; Shimada et al., 1996;Shimada, 2000). Induction of enzymes by environmental chemicals and polymorphisms in P450 genes have been shown to be major factors causing differences in expression and altered catalytic activities in P450 enzymes, and the latter has been shown to be more important (Coughlin and Piper, 1999; Guengerich, 2000; Shimada, 2000). In fact, numerous epidemiological studies have suggested that genetic polymorphisms in CYP1A1 and CYP1B1 genes may produce differences in lung and breast cancer susceptibilities in humans (Kawajiri et al., 1990; Hayashi et al., 1991; Kawajiri and Fujii-Kuriyama, 1991; Nebert et al., 1999; Watanabe et al., 2000). Catalytic specificities of mutated enzymes have been determined in many laboratories using cDNA-based recombinant P450 systems to examine whether genetic polymorphisms cause alterations in protein catalytic activities toward environmental PAH procarcinogens (Guengerich et al., 1996; Doehmer et al., 1999; Shimada et al., 1999c, 2000). For example, replacement of isoleucine by valine at residue 462 in CYP1A1 produced a slight increase in the activity of B[a]P oxidation (Hayashi et al., 1991; Zhang et al., 1996). We have recently compared activities of activation of 12 PAHs, five heterocyclic amines, 2-aminofluorene, and 3-methoxy-4-aminoazobenzene by CYP1B1*1 and CYP1B1*3 in a tester strain Salmonella typhimurium NM2009 and found that the Leu432Val substitution causes small changes in the catalytic properties (Shimada et al., 1999c). However, it remains unclear whether other types of CYP1B1 genetic polymorphisms change catalytic specificities toward a variety of environmental procarcinogens.
In this study, we further compared activities of metabolic activation of a number of PAHs and PAH dihydrodiols and other procarcinogens by recombinant (Escherichia coli and Trichoplusia nicells) human P450 enzymes using a genotoxicity assay based onS. typhimurium NM2009 tester strain (Shimada et al., 1994a). The aims of the current study were 1) to determine differences in catalytic activities of CYP1A1 and CYP1B1 toward activation of seven PAHs, 21 PAH dihydrodiols, and 9-hydroxy-B[a]P to reactive metabolites in this assay system; 2) to compare activities of procarcinogen activation in CYP1B1*1 (Arg48Ala119Leu432Asn453), CYP1B1*2 (Gly48Ser119Leu432Asn453), CYP1B1*3 (Arg48Ala119Val432Asn453), and CYP1B1*62(Gly48Ser119Val432Asn453); 3) to examine differences in catalytic activities of human P450 enzymes using recombinant systems in E. coli andT. ni cells; and 4) to determine activities of other recombinant (T. ni) human P450s, including CYP1A2, 2A6, 2B6, 2C8, 2C9, 2C18, 2C19, 2D6, 2E1, 3A4, 3A5, 3A7, and 4A11 with PAHs and PAH dihydrodiols.
Materials and Methods
Chemicals.
B[a]P-7,8-diols [(±)-, (+)-, and (−)-],9-hydroxy-B[a]P, benz [a]anthracene-1,2-diol, benz[a]anthracene-trans-3,4-diol, benz[a]anthracene-cis-5,6-diol, and benz[a]anthracene-8,9-diol were purchased from the National Cancer Institute Chemical Carcinogen Repository Midwest Research Institute (Kansas City, MO). B[a]P and benz[a]anthracene were purchased from Sigma (St. Louis, MO) and Aldrich (Milwaukee, WI) respectively. 7,12-DMBA, DMBA-trans-3,4-diol, DMBA-cis-5,6-diol, chrysene-1,2-diol, 5-methylchrysene-1,2-diol, 5,6-dimethylchrysene-1,2-diol, dibenzo[a,l]pyrene-11,12-diol, benzo[g]chrysene-11,12-diol, and benzo[c]phenanthrene-3,4-diol were kindly donated by Dr. Stephen Hecht (University of Minnesota, Minneapolis, MN). Other chemicals and reagents used in this study were obtained from sources described previously or were of the highest qualities commercially available (Shimada et al., 1994b, 1999a,c).
Enzymes.
CYP1A1, CYP1A2, and four allelic variants of human CYP1B1, namely, CYP1B1*1 (Arg48Ala119Leu432Asn453), CYP1B1*2 (Gly48Ser119Leu432Asn453), CYP1B1*3 (Arg48Ala119Val432Asn453), and CYP1B1*6 (Gly48Ser119Val432Asn453) were expressed together with human NADPH-P450 reductase inE. coli, and the enzymes thus produced in membranes of the bacteria were used as described previously (Shimada et al., 2000, 2001). Yields of P450, as determined by the original spectral method (Omura and Sato, 1964), ranged between 40 and 250 nmol/l of medium, respectively, and all of the CYP1B1 preparations showed wavelength maxima at 446 nm for the reduced CO complex. NADPH-P450 reductase expression in these bacterial membranes ranged from 25 to 75 nmol/l of culture, and the P450/reductase ratio was determined to be 2 to 5.
Recombinant CYP1A1, 1A2, 1B1, 2A6, 2B6, 2C8, 2C9, 2C18, 2C19, 2D6, 2E1, 3A4, 3A5, 3A7, and 4A11 expressed in microsomes of T. ni cells infected with a baculovirus containing human P450 and NADPH-P450 reductase cDNA inserts were obtained from GENTEST (Woburn, MA); the P450 contents in these systems were described in the data sheets provided by the manufacturer. Three preparations of CYP1B1 (P-220 lot 1, 2, and 3) in T. ni cells were used.
Genotoxicity Assay.
P450-dependent activation of procarcinogens to reactive products that cause induction of umu gene expression in tester strainS. typhimurium NM2009 was determined as described previously (Shimada et al., 1989a, 1996). Standard incubation mixtures included P450 (10 pmol) and 5.0 μM procarcinogen in a final volume of 1.0 ml of 100 mM potassium phosphate buffer, pH 7.4, containing an NADPH-generating system consisting of 0.5 mM NADP+, 5 mM glucose 6-phosphate, and 0.5 unit of glucose 6-phosphate dehydrogenase/ml (Shimada et al., 1998a,b) and 0.75 ml of bacterial suspension. The induction of umu gene expression was monitored by measuring β-galactosidase activity usingo-nitrophenyl-β-d-galactopyranoside as a substrate (Miller, 1972) and is presented as units of β-galactosidase activity per minute per nanomole of P450 (Shimada et al., 1994a).
Other Assays.
7-Ethoxyresorufin and 7-ethoxycoumarin O-deethylation activities were determined by measuring formation of resorufin and 7-hydroxycoumarin, respectively, as described previously (Yamazaki et al., 1997; Shimada et al., 1998a,b; Yamazaki et al., 1998, 1999b). P450 and protein contents were estimated by the methods described (Lowry et al., 1951; Omura and Sato, 1964).
Statistical Analysis.
Kinetic parameters for the activation of procarcinogens by recombinant human P450 enzymes in S. typhimurium NM2009 were estimated by nonlinear regression analysis using the program KaleidaGraph (Synergy Software, Reading, PA).
Results
Metabolic Activation of PAHs and PAH Dihydrodiols and Other Procarcinogens by Recombinant (E. coli) CYP1A1 and Four CYP1B1 Allelic Variants in S.typhimurium NM2009.
CYP1A1 and four allelic variants of CYP1B1, namely, CYP1B1*1 (Arg48Ala119Leu432Asn453), CYP1B1*2 (Gly48Ser119Leu432Asn453), CYP1B1*3 (Arg48Ala119Val432Asn453), and CYP1B1*6 (Gly48Ser119Val432Asn453) expressed in E. coli together with human NADPH-P450 reductase were used for analysis of metabolic activation of PAHs and PAH dihydrodiols in S. typhimuriumNM2009 (Table 1). In this study, we did not determine the effects of cytochrome b5on the catalytic activities, since it has shown that no stimulation of cytochrome b5 is found in the reactions catalyzed by CYP1A1 and CYP1B1 in recombinant P450 systems (Shimada et al., 1998a, 2000; H. Yamazaki, M. Nakamura, T. Komatsu, K. Ohyama, S. Asahi, N. Shimada, F. P. Guengerich, T. Shimada, M. Nakajima, and T. Yokoi, unpublished data). Seven PAHs, 9-hydroxy-B[a]P, and 21 PAH dihydrodiols were examined. Essentially similar catalytic specificities were determined in CYP1A1 and four CYP1B1 variants when (+)-, (−)-, and (±)-B[a]P-7,8-diol, 5-methylchrysene-1,2-diol, 7,12-DMBA-3,4-diol, DB[a,l]P-11,12-diol, benzo[b]fluoranthene-9,10-diol, benzo[c]chrysene, 5,6-dimethylchrysene-1,2-diol, benzo[c]phenanthrene-3,4-diol, 7,12-DMBA, B[a]P, 5-methylchrysene, and benz[a]anthracene were used as substrates. CYP1A1 had relatively higher catalytic activities than CYP1B1 enzymes toward activation of chrysene-1,2-diol, benz[a]anthracene-trans-1,2-, 3,4-, 5,6-, and 8,9-diol, fluoranthene-2,3-diol, DB[a,l]P, benzo[c]phenanthrene, and dibenz[a,h]anthracene.
Kinetic analysis of activation of polycyclic aromatic hydrocarbons and their dihydrodiol derivatives by recombinant (bacterial) CYP1A1 and four CYP1B1 allelic variants in S. typhymurium NM2009
Four CYP1B1 allelic variants had similar catalytic specificities toward activation of PAHs used in this study, except that CYP1B1*2 and CYP1B1*6 had slightly higher activities in catalyzing 7,12-DMBA-3,4-diol, DB[a,l]P-11,12-diol, benzo[j]fluoranthene-4,5-diol, and 9-hydroxy-B[a]P than CYP1B1*1 and CYP1B1*3 (Fig. 1).

Metabolic activation of polycyclic aromatic hydrocarbons and their dihydrodiol derivatives by membranes of E. coli expressing CYP1B*1 (A and a), CYP1B1*2 (B and b), CYP1B1*3 (C and c), CYP1B1*6 (D and d), and CYP1A1 (E and e) in S. typhimurium NM2009.
P450 and substrate concentration used were 10 nM and 5 μM, respectively, and incubation time used was 120 min. DNA damage caused by reactive metabolites of chemicals was determined by measuring β-galactosidase activity in the cells. Results are presented as means of duplicate determinations.
Seven heterocyclic amines and other aryl-, aminoazo-, and nitro-compounds were also used to examine catalytic specificities of CYP1A1 and CYP1B1 variants in S. typhimuriumNM2009 (Fig. 2). CYP1A1 had a tendency to have slightly higher activities than CYP1B1 enzymes in the activation of these compounds, except that 2-nitropyrene activation was catalyzed more efficiently by CYP1B1s than CYP1A1.

Metabolic activation of heterocyclic and aryl amines by membranes of E. coli expressing CYP1B1*1 (A), CYP1B1*2 (B), CYP1B1*3 (C), CYP1B1*6 (D), and CYP1A1 (E) in S. typhimurium NM2009.
Substrate concentration used was 5 μM in all cases. Results are presented as means of duplicate determinations.
Kinetic Analysis of Activation of PAHs and PAH Dihydrodiols and MeIQ by Recombinant (E. coli) CYP1A1 and CYP1B1 and Its Variants in S.typhimurium NM2009.
Kinetic analysis of the activation of four PAHs and their dihydrodiols by CYP1A1 and CYP1B1 and its allelic variants was performed inS. typhimurium NM2009 at substrate concentrations of 1, 3.1, 6.2, 12.5, and 25 μM for PAHs and 0.6, 1.2, 1.8, and 2.4 μM for PAH dihydrodiols (Table 1). In these cases, the dihydrodiol derivatives were more rapidly catalyzed to yield reactive metabolites by CYP1A1 and CYP1B1 enzymes than were the parent compounds. Since the former chemicals were more cytotoxic than the PAHs when they were activated by CYP1A1 and 1B1, we used substrate concentrations of PAH-dihydrodiols between 0.6 and 2.4 μM.Km values for the activation of four dihydrodiols by CYP1A1 and CYP1B1 variants were essentially similar, and the enzyme efficiencies (Vmax/Km ratio) were not so different in these P450 enzymes.Km values andVmax values for the activation of B[a]P by CYP1A1 and CYP1B1 enzymes were larger and smaller, respectively, than those of (±)-B[a]P-7,8-diol, and thus enzyme efficiency was markedly higher with (±)-B[a]P-7,8-diol. In contrast,Km values for the activation of 7,12-DMBA by these P450 enzymes were slightly higher than for 7,12-DMBA-3,4-diol, although the Vmax values were larger in the latter cases. Interestingly, CYP1B1 enzymes did not efficiently activate DB[a,l]P but activated DB[a,l]P-11,12-dihydrodiol at high rates. This is in contrast to the case of CYP1A1, in that this P450 catalyzed both DB[a,l]P and its dihydrodiol to active metabolites. Similar tendencies were also seen in the activation of 5-methylchrysene and its 1,2-dihydrodiol by these P450 enzymes.
We also determined kinetics of activation of the (+)- and (−)-enantiomers of B[a]P-7,8-diol and of MeIQ by CYP1A1, CYP1A2, and CYP1B1*1 in S. typhimurium NM2009 (Fig. 3). For the activation of (+)- and (−)-B[a]P-7,8-diol, these three P450 enzymes had essentially similar Km values, althoughVmax values with CYP1A2 were markedly lower than those of CYP1A1 and CYP1B1*1. In contrast, theKm value for the activation of MeIQ by CYP1A2 (∼0.04 μM) was very low compared with those by CYP1A1 (2.3 μM) and CYP1B1*1 (2.3 μM).

Kinetic analysis of the activation of (+)-B[a]P-7,8-diol (A), (−)-B[a]P-7,8-diol (B), and MeIQ (C) by membranes of E. coli expressing CYP1A1 (●), CYP1B1*1 (○), and CYP1A2 (▪) in S. typhimurium NM2009 tester strain.
Experimental conditions were the same as in Table 1.
Problems in Use of Recombinant (T.ni) CYP1B1 in Assays of Procarcinogen Activation inS. typhimurium NM2009.
Since 15 sets of human P450s coexpressing human NADPH-P450 reductase in T. ni cells were already available from GENTEST, catalytic specificities of these P450 enzymes were determined for the activation of PAHs and PAH dihydrodiols in ourumu assay system. Initially, we compared activities with CYP1A1 and 1B1 expressed in T. ni cells in the activation of (±)-B[a]P-7,8-diol (Fig.4). Unexpectedly, CYP1B1 in T. ni cells was found to have high basal activities in the expression of umu gene expression in S. typhimurium NM2009 tester strain. It was found that CYP1B1 caused an increase in β-galactosidase activities in S. typhimurium NM2009 in the absence of (±)-B[a]P-7,8-diol and NADPH. These increases in the activities were not seen when the enzyme was heat-inactivated at 90°C for 2 min (Fig. 4, B and D). The basal increases in β-galactosidase activities were further enhanced when (±)-B[a]P-7,8-diol was activated by CYP1B1 in the presence of NADPH. These unexpected findings were not seen with CYP1A1 (Fig. 4, A and C).

Effects of concentrations of recombinant (insect) P450 (A and B) and NADPH (C and D) on induction of umu gene expression in S. typhimurium NM2009.
A and B, different concentrations of CYP1A1 and CYP1B1, respectively, were first treated with (○) or without (▵ and ▪) heat inactivation (90°C for 2 min) and then the activation of (±)-B[a]P-7,8-diol (at 5 μM) was determined in the presence (▪) or absence (▵) of 100 μM NADPH by measuring β-galactosidase activities in the bacterial cells. C and D, different concentrations of NADPH were incubated in intact (▵ and ▪) or heat-inactivated (○) CYP1A1 and CYP1B1, respectively, in the presence (▪) or absence (▵) of (±)-B[a]P-7,8-diol (at 5 μM) and the induced umu gene expression was determined as described above.
Other P450 preparations in T. ni cells did not have high basal β-galactosidase activities in S. typhimurium NM2009 with or without heat inactivation of the enzymes (Table 2). However, all three lots of T. ni CYP1B1 preparations had high basal β-galactosidase activities without treatment of enzymes at 90°C for 2 min.
Effects of inactivation (at 90°C for 2 min) of recombinant (T. ni cells) P450 preparations on umu gene expression in S. typhymurium NM2009
We also determined activities of 7-ethoxyresorufin and 7-ethoxycoumarin O-deethylations by CYP1B1 in T. ni cells and found that it catalyzed the former reaction at rates of 12 nmol of resorufin formed/min/nmol of CYP1B1 and the latter at rates of 15 nmol/min/nmol of CYP1B1. Neither reaction proceeded when the NADPH-generating system was removed from the reaction mixture. We also found that α-naphthoflavone (at 5 μM concentration) decreased both activities catalyzed by CYP1B1 in T. nicells (more than 90%). These inhibition patterns were very similar to those obtained in CYP1B1 in E. coli and CYP1A1 enzymes in E. coli and T. ni cells.
Metabolic Activation of PAHs and PAH Dihydrodiols by 14 Forms of Recombinant Human P450 Expressed in T.ni Cells.
Good correlations were observed between CYP1A1 preparations obtained from systems of E. coli and T. ni cells in catalyzing activation of seven PAHs, 9-hydroxy-B[a]P, and 21 PAH dihydrodiols (r = 0.96) (Fig. 5). We further determined activation of PAHs and PAH dihydrodiols by 14 forms of recombinant (T. ni cells) P450 enzymes (Fig.6), except for CYP1B1, because the preparation caused high basal β-galactosidase activities inS. typhimurium NM2009. At least five human P450s were found to have activities with these procarcinogens in this assay system, and CYP1A1 was the highest in catalyzing these reactions followed by CYP1A2. CYP2C9 was found to activate all of the PAH compounds examined at much slower rates than those catalyzed by CYP1A1 and 1A2. CYP2C19 activated some of the PAH compounds at very low rates. CYP3A4 activated several of PAH dihydrodiols but did not activate PAH parent compounds at significant rates. Other P450 enzymes (e.g., CYP2A6, 2B6, 2C8, 2C18, 2D6, 2E1, 3A5, 3A7, and 4A11) did not activate the procarcinogens examined in this assay system.
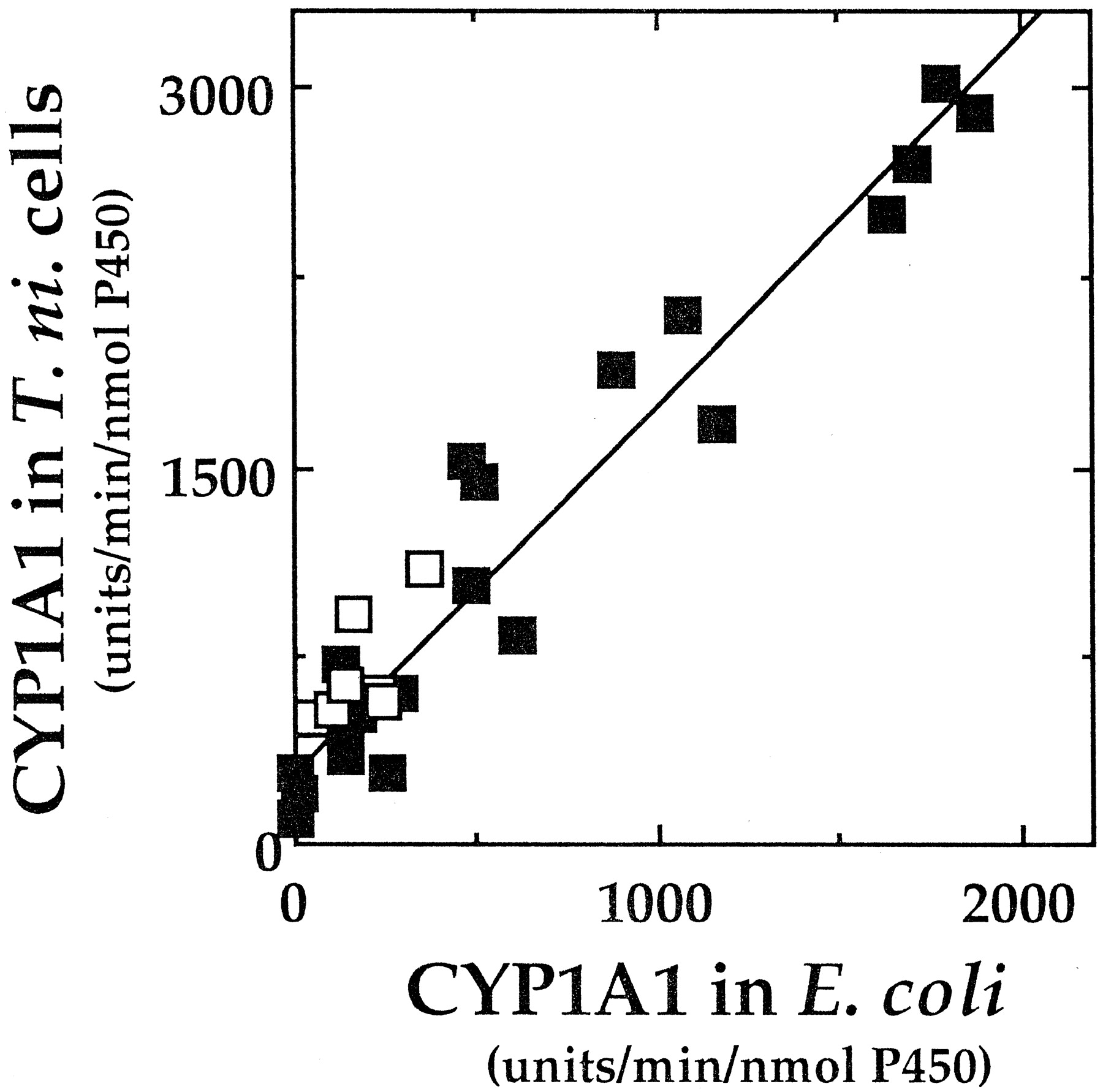
Correlation between activities of induction or umu gene expression by seven PAHs (■) and 21 PAH dihydrodiol derivatives (▪) in recombinant CYP1A1 expressed in E. coli and T. ni cells.

Metabolic activation of PAHs and PAH dihydrodiol derivatives by microsomes of T. ni cells expressing different forms of human P450 together with human NADPH-P450 reductase.
Results are presented as means of duplicate determinations.
Discussion
We previously compared the abilities of recombinant CYP1B1*3 expressed in yeast with CYP1A1 and 1A2 purified from membranes ofE. coli in activating a variety of PAHs and PAH dihydrodiols, arylamines, heterocyclic amines, and nitroarenes inS. typhimurium NM2009 (Shimada et al., 1999c). In these cases, rabbit, rat, and human NADPH-P450 reductase were added to the P450 systems to determine P450 catalytic activities. Conclusions reached in that study showed that CYP1B1*3, CYP1A1, and CYP1A2 play key roles in the activation of a variety of environmental procarcinogens determined so far (Shimada et al., 1999c). However, to compare catalytic activities of these P450 enzymes more precisely, recombinant P450 enzymes obtained from the same cDNA-based systems coexpressing human NADPH-P450 reductase are required to be used (Guengerich et al., 1996, 1997; Crespi and Penman, 1997; Shimada et al., 1999b, 2000).
In this study, we examined metabolic activation of PAH and PAH-dihydrodiols in two cDNA-based systems expressing human P450s and NADPH-P450 reductase in E. coli and T. ni cells. Several studies using these systems have shown that high catalytic rates of substrate oxidations by these P450 enzymes can be achieved and it has also been reported that individual forms of P450 in these systems show essentially similar catalytic rates toward typical drug and xenobiotic substrates (Guengerich et al., 1996;Shimada et al., 1999b, 2000; Yamazaki et al., 1999a; H. Yamazaki, M. Nakamura, T. Komatsu, K. Ohyama, S. Asahi, N. Shimada, F. P. Guengerich, T. Shimada, M. Nakajima, and T. Yokoi, unpublished data). We compared catalytic differences in procarcinogen activation by CYP1A1 and CYP1B1 and its variants obtained from membranes of E. coli by measuring umu gene expression in a tester strain S. typhimurium NM2009. These results suggested that CYP1A1 has relatively higher catalytic activities than CYP1B1 and its variants in the activation of chrysene-1,2-diol, benz[a]anthracene-1,2-, 3,4-, 5,6-, and 8,9-diol, fluoranthene-2,3-diol, DB[a,l]P, benzo[c]phenanthrene, and dibenz[a,h]anthracene, whereas in the activation of other PAHs and PAH dihydrodiols, these enzymes had essentially similar catalytic specificities (Fig. 1). Assays of carcinogenic aryl and heterocyclic amines revealed slightly higher, but not so different, activities by CYP1A1 in comparison with those catalyzed by CYP1B1 enzymes (Fig. 2). These results support the view that both CYP1A1 and CYP1B1 enzymes have important roles in the metabolism of a variety of environmental procarcinogens, particularly PAHs and PAH dihydrodiols that have been shown to cause cancers in extrahepatic organs such as lung and breast (Pelkonen and Nebert, 1982;Hecht, 1999; Shimada, 2000).
Kinetic analysis of the activation of B[a]P, 7,12-DMBA, DB[a,l]P, and 5-methylchrysene and their dihydrodiol derivatives suggests that both CYP1A1 and CYP1B1 enzymes have higher Vmax values andVmax/Km ratios in the activation of PAH dihydrodiols than for the parent compounds. Since we determined formation of various kinds of reactive metabolites of PAH and PAH-dihydrodiols that are genotoxic in S. typhimurium NM2009, the possibility exists that the kinetic parameters may be different from our values when the formations of individual types of reactive metabolites are determined chemically. The dihydrodiols have been shown to be further converted to highly reactive diol-epoxides by P450 enzymes, and in these processes CYP1A1 and CYP1B1 were found to be the key enzymes (Shimada et al., 1996, 1999c). We also obtained evidence suggesting that CYP1A1 and 1B1 have important roles in the activation of parent PAH compounds as well as their dihydrodiol derivatives.
Numerous studies have shown that a variety of DNA reactive metabolites are formed on incubation of PAHs with P450 enzymes (Wood et al., 1976;Gelboin, 1980; Pelkonen and Nebert, 1982; Hall et al., 1989; Shimada et al., 1989b; McManus et al., 1990; Rojas et al., 1992; Shields et al., 1993; Shou et al., 1994). In the case of B[a]P, P450s produce several reactive metabolites, e.g., 1,2-, 2,3-, 4,5-, 7,8-, and 9,10-epoxides, which are readily hydrolyzed to dihydrodiol metabolites if epoxide hydrolase is present (Gelboin, 1980; Pelkonen and Nebert, 1982). It is, however, not known at present time which reactive metabolites of B[a]P, 7,12-DMBA, DB[a,l]P, and 5-methylchrysene produced by human P450s cause DNA damage in our assay system. 9-Hydroxy-B[a]P has been suggested to be oxidized to a reactive 4,5-epoxide metabolite (Pelkonen and Nebert, 1982) and was shown in this study to be activated by CYP1B1*2 and CYP1B1*6 more rapidly than other P450 forms.
Four genetic polymorphisms in human CYP1B1 gene have been shown to cause amino acid replacements at Arg48Gly, Ala119Ser, Leu432Val, and Asn453Ser (Shimada, 2000; Watanabe et al., 2000). Our previous studies have suggested that replacement of Leu by Val at residue 432 (change of CYP1B1*1 to CYP1B1*3) does not cause significant alterations in the activation of 19 procarcinogens in S. typhimurium NM2009 (Shimada et al., 1999c). In this study, we found no large differences in the activities of procarcinogens by CYP1B1*2 and *6 compared with other CYP1B1 variants in catalyzing activation of some procarcinogens, with exception that CYP1B1*2 and CYP1B1*6 had slightly higher activities for 7,12-DMBA-3,4-diol, DB[a,l]P-11,12-diol, benzo[j]fluoranthene-4,5-diol, and 9-hydroxy-B[a]P than did CYP1B1*1 and CYP1B1*3 (Fig. 1). However, it remains unclear whether amino acid replacement at residue 453 of CYP1B1 causes alterations in its catalytic specificities.
We used two recombinant P450 systems in this study. There were good correlations between two CYP1A1 preparations expressed in E. coli and T. ni cells for the activation of a variety of PAH compounds in our assay system. However, in all three lots of CYP1B1 preparations expressed in T. ni cells (obtained from GENTEST) high basal activities of β-galactosidase in S. typhimurium NM2009 were determined. These basal activities did not require NADPH and substrate but were lost following heat inactivation of the enzyme at 90°C for 2 min. These CYP1B1 preparations were catalytically active; they catalyzed 7-ethoxyresorufin and 7-ethoxycoumarinO-deethylation at high rates, and these activities were completely inhibited by typical inhibitor α-naphthoflavone (Shimada and Okuda, 1988; Shimada et al., 1989a,b). Fourteen other P450 preparations in T. ni cells used did not have such high basal β-galactosidase activities and could be used to examine carcinogen activation in our assay system.
Among the 14 forms of human P450 enzymes expressed in T. ni cells tested, CYP1A1, CYP1A2, CYP2C9, CYP2C19, and CYP3A4 activated a variety of PAHs and PAH dihydrodiols with CYP1A1 had the highest activities followed by CYP1A2. Our earlier work showed that CYP3A4 catalyzes activation of some of the PAH dihydrodiols in theumu assay (Shimada et al., 1989a,b), confirmed in the present study in which recombinant CYP3A4 (in T. ni cells) is involved in the oxidation of at least 10 dihydrodiols of PAHs to reactive metabolites. Interestingly, CYP3A4 did not catalyze PAH parent compounds at high rates, whereas CYP2C9 had activities toward both PAHs and PAH dihydrodiols. These findings support the previous work (Yun et al., 1992) in which both CYP2C9 and CYP3A4 catalyzed 3-hydroxylation of B[a]P in human liver microsomes. CYP2C19 was less active with some of the PAH compounds, but had finite activity. Other P450 enzymes, CYP2A6, 2B6, 2C8, 2C18, 2D6, 2E1, 3A5, 3A7, and 4A11, did not catalyze any activities with these procarcinogens.
In conclusion, we studied the activation of a variety of PAHs and PAH dihydrodiols and other procarcinogens by human P450 enzymes in anS. typhimurium NM2009 tester strain using individual P450 enzymes in E. coli andT. ni cells. The results obtained support the importance of CYP1A1 and CYP1B1 in the activation of PAHs and PAH dihydrodiols; other P450 enzymes such as CYP1A2, CYP2C9, and CYP3A4 have abilities to catalyze PAH compounds at much slower rates. We also reported small differences between CYP1A1 and CYP1B1 in the activation of a variety of procarcinogen compounds determined. However, it should be mentioned that when recombinant CYP1B1 in T. ni cells was used for the umu assay, high basal β-galactosidase activities were determined in the absence of NADPH and procarcinogens. Further work will be needed to address this problem.
Footnotes
-
This work was supported in part by grants from the Ministry of Education, Science, and Culture of Japan and the Ministry of Health and Welfare of Japan (T.S.); by United States Public Health Service Grants R35 CA44353 and P30 ES00267 (F.P.G.); and by the Kathleen Cuningham Foundation for Breast Cancer Research (E.M.J.G.).
-
↵2 Nomenclature suggested by Oscarson et al. (http://www.imm.ki.se/CYPalleles).
- Abbreviations used are::
- PAH
- polycyclic aromatic hydrocarbon
- P450
- general term for cytochrome P450
- CYP
- individual forms of P450
- B[a]P
- benzo[a]pyrene
- 7,12-DMBA
- 7,12-dimethylbenz[a]anthracene
- DB[a,l]P
- dibenzo[a,l]pyrene
- MeIQ
- 2-amino-3,5-dimethylimidazo[4,5-f]quinoline
- Received March 26, 2001.
- Accepted May 15, 2001.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}