


Abstract
In adult rat ventricular cardiomyocytes, noradrenaline exerts dual effects on protein synthesis: increases via α1-adrenoceptors and decreases via β1-adrenoceptors. Carvedilol and bucindolol are β-blockers with additional α1-adrenoceptor blocking activities. We studied the effects of carvedilol and bucindolol on noradrenaline-induced protein synthesis (assessed by [3H]phenylalanine incorporation) in adult rat ventricular cardiomyocytes. Radioligand binding studies with [125I]iodocyanopindolol and [3H]prazosin revealed that carvedilol had a much higher affinity to α1-adrenoceptors than bucindolol (β1-/α1-adrenoceptor ratio for carvedilol, 1:2.7; for bucindolol, 1:43). Noradrenaline-evoked increases in protein synthesis were enhanced by propranolol (1 μM) and β1-adrenoceptor-selective antagonists bisoprolol (1 μM) and CGP 20712A [1-[2-((3-carbamoyl-4-hydroxy)phenoxy)-ethyl-amino]-3-[4-(1-methyl-4-trifluoromethyl-2-imidazolyl)phenoxy]-2-propranol methanesulfonate] (300 nM). Carvedilol (100 pM–10 μM) inhibited 1 μM noradrenaline-induced increase in protein synthesis with monophasic concentration-inhibition curves independent of whether CGP 20712A was present or not; Ki values for carvedilol were 5 to 6 nM. In contrast, bucindolol (100 pM–10 μM) inhibited l μM noradrenaline-induced increase in protein synthesis with a bell-shaped concentration-inhibition curve; it increased noradrenaline-induced protein synthesis at 10 nM, although at concentrations >100 nM it was inhibited. In the presence of 300 nM CGP 20712A or 1 μM propranolol, however, bucindolol inhibited 1 μM noradrenaline-induced increase in protein synthesis with monophasic concentration-inhibition curves; Ki values were 40 to 75 nM. On the other hand, both carvedilol and bucindolol inhibited 1 μM phenylephrine-induced protein synthesis with monophasic concentration-inhibition curves;Ki values were 4 (carvedilol) and 45 nM (bucindolol). These results indicate that, at low (β-adrenoceptor blocking) concentrations, bucindolol can enhance noradrenaline-induced protein synthesis whereas it is inhibited by carvedilol.
β-Adrenoceptor antagonists are now commonly used in the treatment of chronic heart failure patients (for review, see Bristow, 2000). Beneficial effects including reduction of mortality have been demonstrated not only for β1-adrenoceptor-selective antagonists such as metoprolol (MERIT-HF Study Group, 1999; Hjalmarson and Fagerberg, 2000) or bisoprolol (CIBIS-II Investigators and Committees, 1999) but also for the nonselective β-adrenoceptor antagonist carvedilol (CAPRICORN Investigators, 2001; Packer et al., 2001; Tendera and Ochala, 2001). On the other hand, a recent large trial with bucindolol, another nonselective β-adrenoceptor antagonist, failed to produce a significant reduction in mortality in patients with chronic heart failure (Bristow, 2000). Bucindolol (Hershberger et al., 1990) and carvedilol (Bristow et al., 1992) are β-adrenoceptor antagonists with a similar affinity at β1- and β2-adrenoceptors but differ in their affinities at α1-adrenoceptors. Carvedilol exhibited about 2- to 3-fold selectivity for β1- versus α1-adrenoceptors whereas bucindolol was found to have about 60 to 70 times higher affinity at β1- versus α1-adrenoceptors (Sponer and Feuerstein, 1999;Bristow, 2000). We have recently shown that, in adult rat ventricular cardiomyocytes, the noradrenaline-induced increase in rate of protein synthesis (a marker of the development of a hypertrophic phenotype) is composed of two components: an α1-adrenoceptor mediated increase in rate of protein synthesis, and a β1-adrenoceptor mediated reduction in rate of protein synthesis (Schäfer et al., 2001). The aim of this study was to find out whether the differences in affinities to α1-adrenoceptors of carvedilol and bucindolol might lead to differences in their interaction with noradrenaline-induced increase in rate of protein synthesis (assessed by [3H]phenylalanine incorporation in the cardiomyocytes) in adult rat ventricular cardiomyocytes.
Materials and Methods
Preparation of Cardiomyocyte Culture of Adult Rats.
Left ventricular cardiomyocytes were isolated from 12-week-old male Wistar rats exactly as recently described (Pönicke et al., 2000). Freshly isolated cardiomyocytes were gently diluted in sterile culture medium M199, pH 7.4, supplemented with 10% newborn calf serum. To study [3H]phenylalanine incorporation, the cardiomyocyte suspension was seeded into 12-well plates (16,000 cells per well), which had been coated with 4% fetal calf serum in medium M199 for 24 h at 37°C (in a humidified incubator at 95% air/5% CO2) and incubated for 16 h at 37°C. Thereafter, the cultures were rinsed with serum-free Hanks' balanced salt solution to remove damaged, rounded, and nonattached cells, and the rod-shaped cells were cultured in serum-free medium M199 supplemented with 2 mM l-carnitine, 5 mM taurine, 5 mM creatine, and antibiotics (100 U ml−1 penicillin and 100 μg ml−1 streptomycin). To prevent growth of nonmyocytes the culture medium was supplemented with 10 μM cytosine-β-d-arabinofuranoside.
[3H]Phenylalanine Incorporation.
Protein synthesis by cardiomyocytes was assessed as incorporation of [3H]phenylalanine into cells as recently described (Pönicke et al., 2000). Briefly, after addition of [3H]phenylalanine (0.5 μCi ml−1) at 37°C and the addition of various concentrations of noradrenaline or phenylephrine in the presence or absence of the various antagonists, cells were cultured for 16 h at 37°C in 95% air/5% CO2. Ascorbate (100 μM) was always present in the medium during this incubation period as an antioxidant. Thereafter cells were washed with ice-cold 0.9% NaCl solution to remove attached radioactivity and incubated for 24 h at 4°C with 10% trichloroacetic acid. Acid-insoluble precipitates were washed again with 10% trichloroacetic acid and twice with 0.9% NaCl. The remaining precipitate on the culture dishes was solubilized in 1 N NaOH supplemented with 0.1% sodium dodecyl sulfate at room temperature for 24 h, and incorporation of radioactivity into acid-insoluble cell mass was determined by the use of a liquid scintillation counter (Beckman LS 6000; Beckman Coulter, Inc., Fullerton, CA). We have recently shown that under these experimental conditions [3H]phenylalanine incorporation was paralleled by increases in protein mass, cell volume, and cross-sectional area of the cells indicating hypertrophic growth of the cardiomyocytes (Schäfer et al., 2001).
Radioligand Binding Studies.
Affinities of carvedilol and bucindolol to β1- and β2-adrenoceptors were determined by (−)-[125I]iodocyanopindolol binding to membranes from rat heart left ventricle (in the presence of 50 nM ICI 118,551 = homogeneous population of β1-adrenoceptors) and rat lung [in the presence of 300 nM CGP 20712A (1-[2-((3-carbamoyl-4-hydroxy)phenoxy)-ethyl-amino]-3-[4-(1-methyl-4-trifluoromethyl-2-imidazolyl)phenoxy]-2-propranol methanesulfonate] = homogenous population of β2-adrenoceptors; Brown et al., 1992). Crude membrane fractions were prepared by standard homogenization and centrifugation, and (−)-[125I]iodocyanopindolol binding was performed in 10 mM Tris, 154 mM NaCl buffer, pH 7.4, in a total volume of 250 μl for 90 min at 37°C; nonspecific binding was defined as binding in the presence of 1 μM CGP 12177 [4-(3′-tert-butylamino-2′-hydroxypropoxy)-benzimidazole-2 hydrochloride] (Michel et al., 1987). Affinities of carvedilol and bucindolol for α1-adrenoceptors were assessed by [3H]prazosin binding to rat liver membranes. [3H]Prazosin binding was performed in 50 mM Tris, 1 mM EDTA buffer, pH 7.4, in a total volume of 1 ml for 45 min at 25°C; nonspecific binding was defined by 10 μM phentolamine (Michel et al., 1993).
Drugs.
l-[2,3,4,5,6-3H]Phenylalanine (specific activity, 5.03 TBq mmol−1) was purchased from Amersham Buchler (Braunschweig, Germany), (−)-[125I]iodocyanopindolol (specific activity, 81.4 TBq mmol−1) was purchased from PerkinElmer Life Sciences (Zaventem, Belgium), (−)-noradrenaline bitartrate, l-phenylephrine hydrochloride, l-phenylalanine, cytosine-β-d-arabinofuranoside, sodium dodecyl sulfate, trypsin (crude), l-carnitine, taurine, creatine, propranolol, and ICI 118,551 were purchased from Sigma-Aldrich (Deisenhofen, Germany). CGP 20712A and CGP 12177 was kindly donated by Ciba-Geigy (Basel, Switzerland). Carvedilol and bucindolol hydrochloride were kindly provided by Dr. Eliot H. Ohlstein (GlaxoSmithKline, King of Prussia, PA). Hanks' balanced salt solution, culture medium M199, and penicillin-streptomycin were obtained from Invitrogen (Eggenstein, Germany). All other chemicals were of the highest purity grade commercially available.
Statistical Evaluations.
Data given are means ± S.E.M. of n experiments. Experimental data for agonist-induced increases in rate of protein synthesis were analyzed by fitting sigmoidal curves to the experimental data using the GraphPad Prism 3.0 program (GraphPad Software, San Diego, CA); in these calculations the bottom of the curves were fixed to 100% (i.e., no increases in rate of protein synthesis above control), the Hill slopes were fixed 1.0. Antagonist concentration-inhibition curves were also analyzed by the GraphPad Prism 3.0 program. Statistical significance of difference was analyzed by paired two-tailed Student's t test. Ap value less than 0.05 was considered to be significant. All statistical calculations were performed with the GraphPad Prism 3.0 program.
Results
Affinities of Carvedilol and Bucindolol at α1- and β-Adrenoceptors.
Carvedilol (10 pM–10 μM) and bucindolol (10 pM–10 μM) inhibited [125I]iodocyanopindolol binding (100 pM) to membranes from rat left ventricle (in the presence of 50 nM ICI 118,551 = homogeneous population of β1-adrenoceptors) and from rat lung (in the presence of 300 nM CGP 20712A = homogenous population of β2-adrenoceptors) with concentration-inhibition curves that were best described by a one-site fit. From these curves,Ki values of 0.7 to 1.0 nM for carvedilol and 1.2 to 1.8 nM for bucindolol were calculated (see Table1). Studies with human cardiac membranes had suggested that binding of carvedilol and bucindolol to β-adrenoceptors can be modulated by guanine nucleotides (Hershberger et al., 1990; Bristow et al., 1992; Maack et al., 2000). In the present study, however, addition of 100 μM GTP to the incubation medium did not affect the concentration-inhibition curves of carvedilol or bucindolol, neither at rat β1- nor at rat β2-adrenoceptors (Table 1), in accordance with recently published data from Willette et al. (1999). Carvedilol (100 pM–10 μM) and bucindolol (100 pM–10 μM) inhibited [3H]prazosin binding (1 nM) to rat liver membranes with monophasic concentration-inhibition curves; affinity of carvedilol to rat α1-adrenoceptors was, however, about 30 times higher than that of bucindolol (Table 1).
Affinities (nM) of carvedilol and bucindolol at rat heart β1-, rat lung β2-, and rat liver α1-adrenoceptors
Effect of Carvedilol and Bucindolol on Noradrenaline-Induced Increase in Rate of Protein Synthesis in Cardiomyocytes of Adult Rats.
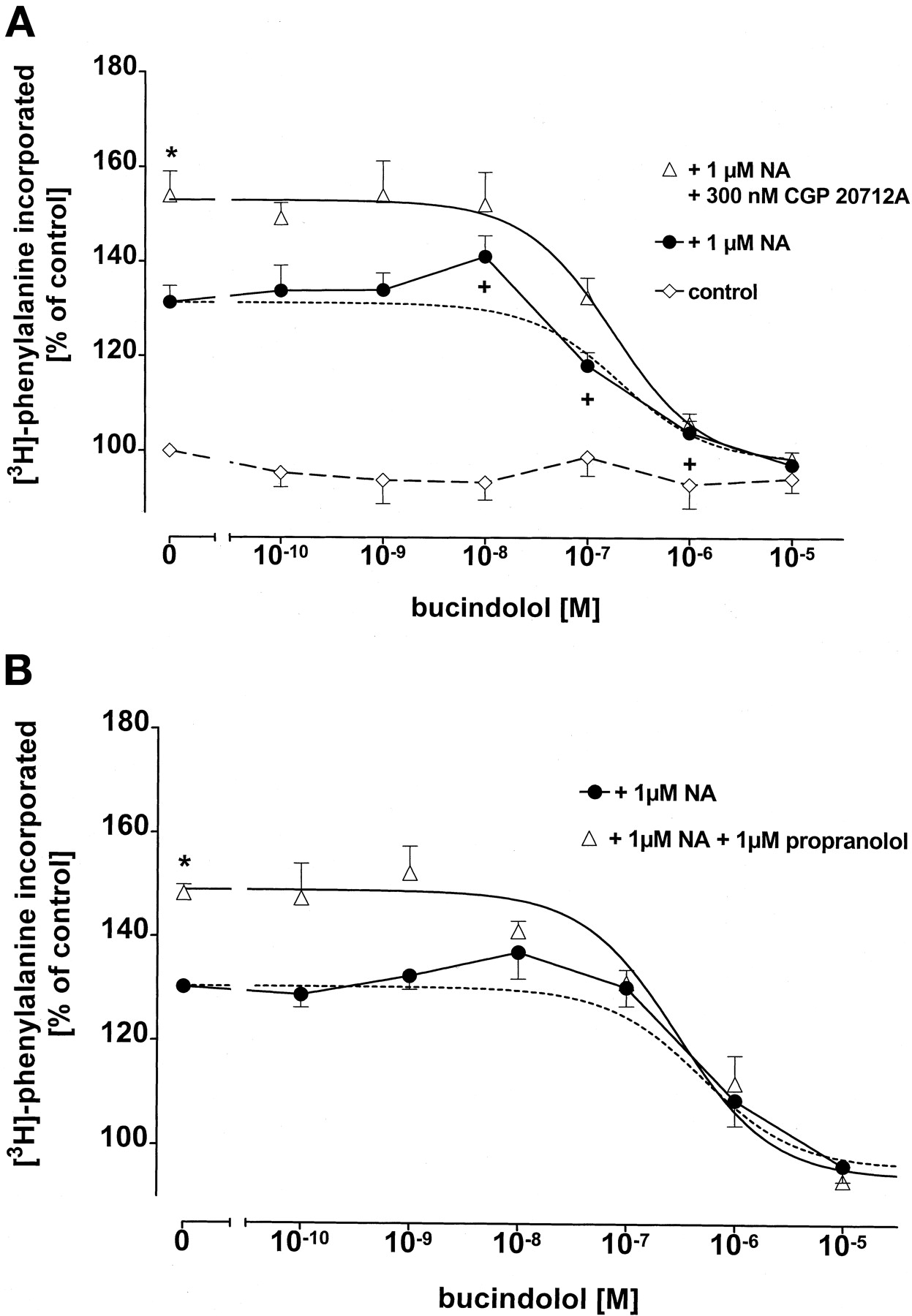
We have recently shown that noradrenaline (NA, 1 nM–10 μM) in adult rat ventricular cardiomyocytes concentration-dependently increases the rate of protein synthesis (assessed as [3H]phenylalanine incorporation); this NA effect can be enhanced by the β-adrenoceptor antagonists propranolol (nonselective), CGP 20712A and atenolol (β1-adrenoceptor-selective; Schäfer et al., 2001). In the present study, NA (1 nM–10 μM) concentration-dependently increased [3H]phenylalanine incorporation into cardiomyocytes (Fig. 1), maximal increase amounted to 130 ± 1% of control, pEC50value was 6.0 ± 0.2. The β1-adrenoceptor antagonist bisoprolol (1 μM) led to a significant increase in rate of protein synthesis at each NA concentration (NA ≥ 100 nM), maximal increase was 151 ± 5% of control. In addition, the concentration-effect curve for NA was significantly shifted to the left; pEC50 value in the presence of bisoprolol was 7.2 ± 0.2 (p < 0.05, Fig. 1). Bucindolol (100 pM–10 μM) influenced 1 μM NA-induced increase in rate of protein synthesis with a bell-shaped concentration-inhibition curve (Fig. 2A); at concentrations up to 10 nM, it increased NA-induced protein synthesis whereas at concentrations >10 nM, NA-induced protein synthesis was inhibited. Addition of 300 nM CGP 20712A to the incubation medium significantly (p < 0.05) increased 1 μM NA-induced protein synthesis from 131 ± 4% to 154 ± 5%; in the presence of CGP 20712A, bucindolol (100 pM–10 μM) inhibited NA-induced protein synthesis with a monophasic concentration-inhibition curve and aKi value of 38.4 ± 8.0 nM (n = 9, Fig. 2A). A monophasic concentration-inhibition curve for bucindolol (Ki value 72.5 ± 16 nM, n = 4) was also observed when in the cardiomyocytes β-adrenoceptors were antagonized by 1 μM propranolol (Fig. 2B).

Effect of the β1-selective adrenoceptor antagonist bisoprolol (1 μM) on NA-induced [3H]phenylalanine incorporation in ventricular cardiomyocytes of 12-week-old male (n = 5) Wistar rats. Ordinate scale, [3H]phenylalanine incorporation as percentage of basal incorporation; abscissa scale, molar concentrations of noradrenaline. Basal [3H]phenylalanine incorporation amounted to 1062 ± 83 cpm in control cells (n= 5). ★, p < 0.05 versus NA-stimulated control cells. Values are means and vertical lines show S.E.M.

A, effect of bucindolol in the presence or absence of the β1-selective adrenoceptor antagonist CGP 20712A (300 nM) on 1 μM NA-induced [3H]phenylalanine incorporation in ventricular cardiomyocytes of 12-week-old male Wistar rats (n = 9). Ordinate scale, [3H]phenylalanine incorporation as percentage of basal incorporation; abscissa scale, molar concentrations of bucindolol. Basal [3H]phenylalanine incorporation amounted to 874 ± 73 cpm in control cells (n = 9).+, p < 0.05 versus NA control (without bucindolol); ★, p < 0.05 versus without CGP 20712A. B, effect of bucindolol in the presence or absence of the nonselective β-adrenoceptor antagonist propranolol (1 μM) on 1 μM NA-induced [3H]phenylalanine incorporation in ventricular cardiomyocytes (n = 4). Ordinate scale, [3H]phenylalanine incorporation as percentage of basal incorporation; abscissa scale, molar concentrations of bucindolol. Basal [3H]phenylalanine incorporation amounted to 912 ± 118 cpm in control cells (n = 4). ★, p < 0.05 versus without propranolol. In panels A and B, values are means and vertical lines show S.E.M; the dotted lines represent a mathematical fit of the data to a theoretical one-site model. Control = effects of the different bucindolol concentrations on basal [3H]phenylalanine incorporation.
We next tested whether bucindolol might have also dual effects on the concentration-response curve for NA-induced protein synthesis. As shown in Fig. 3, in the presence of the low concentration of 10 nM bucindolol, NA (100 nM–10 μM) caused a higher increase in [3H]phenylalanine incorporation at each concentration than in the absence of bucindolol whereas in the relatively high concentration of 1 μM bucindolol caused a significant shift to the right of the concentration-response curve of NA; at 10 μM NA, the increase in protein synthesis was significantly reduced from 133.9 ± 7.9% to 116.0 ± 6.3% (n = 5,p < 0.05; Fig. 3).

Effect of bucindolol (10 nM and 1 μM) on NA-induced [3H]phenylalanine incorporation in ventricular cardiomyocytes of 12-week-old male Wistar rats (n = 5). Ordinate scale, [3H]phenylalanine incorporation as percentage of basal incorporation. Abscissa scale, molar concentrations of noradrenaline. Basal [3H]phenylalanine incorporation amounted to 723 ± 122 cpm in untreated control cells (n = 5). ★, p < 0.05 versus NA-stimulated control cells. Values are means and vertical lines show S.E.M.
In contrast to bucindolol, carvedilol (100 pM–10 μM) inhibited 1 μM NA-induced increase in rate of protein synthesis with a monophasic concentration-inhibition curve (Fig. 4A) independent of whether or not β1-adrenoceptors were blocked with CGP 20712A (300 nM);Ki values for carvedilol were 5.0 ± 1.3 nM (n = 7) in the absence and 5.7 ± 1.4 nM (n = 7) in the presence of 300 nM CGP 20712A. Accordingly, carvedilol did not exert a dual effect on the concentration-response curve for NA-induced protein synthesis; the low (10 nM) concentration of carvedilol was without effect whereas the higher (100 nM and 1 μM) concentrations of carvedilol shifted the concentration-response curve for NA to the right to higher concentrations (Fig. 4B).
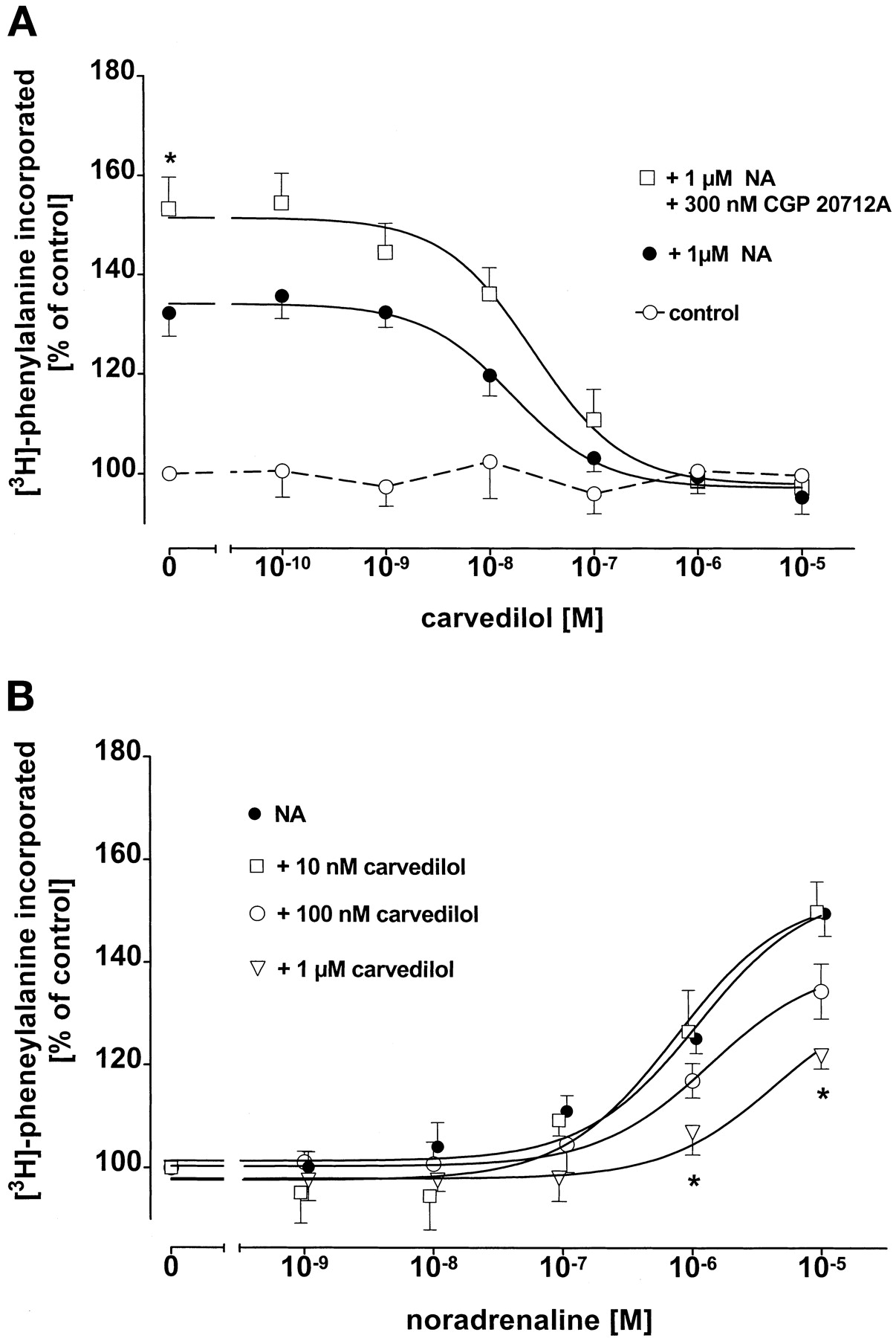
A, effect of carvedilol in the presence or absence of the β1-selective adrenoceptor antagonist CGP 20712A (300 nM) on 1 μM NA-induced [3H]phenylalanine incorporation in ventricular cardiomyocytes of 12-week-old male Wistar rats (n = 7). Ordinate scale, [3H]phenylalanine incorporation as percentage of basal incorporation; abscissa scale, molar concentrations of carvedilol. Basal [3H]phenylalanine incorporation amounted to 797 ± 104 cpm in control cells (n = 7). ★,p < 0.05 versus without CGP 20712A. Control = effects of the different carvedilol concentrations on basal [3H]phenylalanine incorporation. B, effect of carvedilol (10 nM, 100 nM, and 1 μM) on NA-induced [3H]phenylalanine incorporation in ventricular cardiomyocytes (n = 7). Ordinate scale, [3H]phenylalanine incorporation as percentage of basal incorporation. Abscissa scale, molar concentrations of noradrenaline. Basal [3H]phenylalanine incorporation amounted to 1034 ± 143 cpm in control cells (n = 7). ★,p < 0.05 versus NA-stimulated control cells. In panels A and B, values are means and vertical lines show S.E.M.
Effects of Carvedilol and Bucindolol on Phenylephrine-Induced Increase in Rate of Protein Synthesis.
In a final set of experiments, we studied the effects of carvedilol and bucindolol on phenylephrine (PE)-induced increase in rate of protein synthesis because PE is an α1-adrenoceptor agonist that exerts β-adrenoceptor agonist activity only in concentrations >1 μM. PE (1 μM) increased protein synthesis in the adult rat cardiomyocytes to 150 ± 3% (Fig.5, A and B, n = 20), i.e., a value in a range similar to that induced by 1 μM NA in the presence of CGP 20712A in the same cardiomyocytes preparations (cf. Figs. 2A and 4A); 300 nM CGP 20712A did not affect the increase in protein synthesis induced by 1 μM PE [maximal increase in the presence of CGP 20712A was 148 ± 10% (n = 4, Fig. 5A), respectively, 153 ± 6% (n = 4, Fig.5B)]. Both bucindolol and carvedilol inhibited 1 μM PE-induced protein synthesis with monophasic concentration-inhibition curves;Ki values were 44.7 ± 6.3 nM (n = 10) for bucindolol and 3.5 ± 0.4 nM (n = 10) for carvedilol (Fig. 5B).

Effect of bucindolol (panel A, n= 10) or carvedilol (panel B, n = 10) in the presence or absence of the β1-selective adrenoceptor antagonist CGP 20712A (300 nM) on 1 μM PE-induced [3H]phenylalanine incorporation in ventricular cardiomyocytes of 12-week-old male Wistar rats (n = 10). Ordinate scale, [3H]phenylalanine incorporation as percentage of basal incorporation. Abscissa scale, molar concentrations of bucindolol (A) or carvedilol (B). Basal [3H]phenylalanine incorporation amounted to 1099 ± 137 cpm (A, n = 10) and 939 ± 111 cpm (B,n = 10) in control cells. In panels A and B, values are means and vertical lines show S.E.M. Control = effects of the different bucindolol (A) and carvedilol (B) concentrations on basal [3H]phenylalanine incorporation.
Discussion
Adult rat cardiomyocytes contain α1- (Brown et al., 1985; Buxton and Brunton, 1986) and β1- and β2-adrenoceptors (Im et al., 1984; Xiao and Lakatta, 1993; for a recent review, see Steinberg, 1999). We have recently shown that noradrenaline, an α1-, α2-, and β1-adrenoceptor agonist, exerts dual growth-promoting effects on the cardiomyocytes. It increases rate of protein synthesis via α1-adrenoceptor stimulation and decreases rate of protein synthesis via β1-adrenoceptor stimulation in a cyclic AMP-dependent fashion (Schäfer et al., 2001). Accordingly, β-adrenoceptor antagonists such as propranolol, CGP 20712A, and atenolol increased noradrenaline-induced protein synthesis whereas β-adrenoceptor agonists such as isoprenaline, dobutamine, or xamoterol decreased noradrenaline-induced protein synthesis (Brodde et al., 2001; Schäfer et al., 2001). Moreover, forskolin and dibutyryl cyclic AMP inhibited noradrenaline-induced protein synthesis whereas RpcAMPS, an inhibitor of protein kinase A, could enhance it (Schäfer et al., 2001; Brodde et al., 2001). On the other hand, the increases in rate of protein synthesis evoked by phenylephrine, a “pure” α1-adrenoceptor agonist (at least in concentrations up to 1 μM), were not affected by β-adrenoceptor antagonists but were inhibited by the β-adrenoceptor agonists isoprenaline, dobutamine, and xamoterol (Brodde et al., 2001).
Carvedilol and bucindolol are nonselective third-generation β-blockers (Bristow, 2000) with additional α1-adrenoceptor blocking activity. Our radioligand binding data confirm data from the literature (Hershberger et al., 1990; Bristow et al., 1992) that carvedilol has a much higher affinity to α1-adrenoceptors than bucindolol. We found a β1-/α1-adrenoceptor ratio for carvedilol of 1:2.7 and for bucindolol of 1:43 (cf. Table 1).
In the present study, in adult rat ventricular cardiomyocytes, bucindolol—in contrast to carvedilol—exerted a dual effect on noradrenaline-induced increase in rate of protein synthesis. At low, rather selective, β-adrenoceptor blocking concentrations (≤10 nM), bucindolol caused an increase in noradrenaline-induced protein synthesis (similar to what is observed with other β-adrenoceptor antagonists; see above) while at higher (now α1-adrenoceptor blocking) concentrations, it inhibited noradrenaline-induced increase in rate of protein synthesis. The increase in noradrenaline-induced protein synthesis is due to the β-adrenoceptor blocking activity of bucindolol because it completely disappeared when noradrenaline effects were assessed in the presence of 300 nM CGP 20712A or 1 μM propranolol; on the other hand, the inhibitory effects of bucindolol on noradrenaline-induced protein synthesis is due to its α1-adrenoceptor blocking activity since nearly identical concentration-inhibition curves for bucindolol were obtained inhibiting increase in rate of protein synthesis induced by noradrenaline in the presence of 300 nM CGP 20712A (now acting solely at α1-adrenoceptors) or by phenylephrine (a pure α1-adrenoceptor agonist).
In contrast to bucindolol, carvedilol did not exert dual effects on noradrenaline-induced increase in rate of protein synthesis; it inhibited noradrenaline-induced increases in rate of protein synthesis in the presence as well as in the absence of 300 nM CGP 20712A with monophasic concentration-inhibition curves and nearly identicalKi values (see Fig. 4A). In addition, similar Ki values were also obtained for carvedilol-evoked inhibition of phenylephrine-induced increase in rate of protein synthesis; this supports the view that the inhibitory effect of carvedilol is due to its α1-adrenoceptor blocking activity.
It should be noted, however, that bucindolol (1 μM) and carvedilol (100 nM and 1 μM) caused shifts to the right of the concentration-response curve for noradrenaline-induced increase in rate of protein synthesis that were by far less than could be expected from their affinities for α1-adrenoceptors. Thus, at a concentration of 1 μM, carvedilol should cause about a 200-fold shift to the right, but the shift observed was only about 20-fold; similarly, bucindolol at a concentration of 1 μM should cause about a 25- to 30-fold shift to the right, but the shift observed was only about 10-fold. However, at these rather high concentrations, both carvedilol and bucindolol will inhibit also β-adrenoceptors and, by this, induce a shift to the left of the concentration- response curve for noradrenaline. As a consequence, these β-adrenoceptor blocking activities will partly counteract the α1-adrenoceptor blocking effects of both β-adrenoceptor antagonists; this is very likely the reason why the shifts to the right induced by bucindolol and carvedilol are less than could be expected from their α1-adrenoceptor blocking activities.
In conclusion, carvedilol and bucindolol differentially affect noradrenaline-induced increase in rate of protein synthesis in adult rat ventricular cardiomyocytes. Bucindolol exerts a dual effect with low (pure β-adrenoceptor blocking) concentrations increasing noradrenaline-induced protein synthesis and high (now α1-adrenoceptor blocking) concentrations inhibiting noradrenaline-induced increase in rate of protein synthesis. In contrast, carvedilol does not exert such dual effects. It causes no increase but only inhibition of noradrenaline-induced increase in rate of protein synthesis. However, how and/or whether these differential effects of carvedilol and bucindolol on noradrenaline-induced development of a hypertrophic phenotype might contribute to their different effects on mortality in long-term treatment of patients with chronic heart failure (see Introduction) remains to be elucidated.
Acknowledgments
The skillful technical assistance of I. Adler, A. Beilfuß, A. Dunemann, A. Hauser, and M. Niebisch is gratefully acknowledged.
Footnotes
-
This work was supported by grants from the Deutsche Forschungsgemeinschaft (Bonn, Germany; SFB TR 2-01) and by GlaxoSmithKline Pharmaceuticals (King of Prussia, PA).
- Abbreviations:
- CGP 20712A
- 1-[2-((3-carbamoyl-4-hydroxy)phenoxy)-ethyl-amino]-3-[4-(1-methyl-4-trifluoromethyl-2-imidazolyl)phenoxy]-2-propranol methanesulfonate
- CGP 12177
- 4-(3′-tert-butylamino-2′-hydroxypropoxy)-benzimidazole-2 hydrochloride
- NA
- noradrenaline
- PE
- phenylephrine
- ICI 118,551
- (±)-1-[2,3-(dihydro-7-methyl-1H-inden-4-yl)oxy]-3-[1-methylethyl)amino]-2-butanol hydrochloride
- Received July 25, 2001.
- Accepted December 5, 2001.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}